
Method Article
Липидомика тканей грызунов
В этой статье
Резюме
Липидомика на основе масс-спектрометрии дробовика обеспечивает чувствительный количественный снимок широкого спектра классов липидов одновременно в одном измерении из различных тканей грызунов.
Аннотация
Липиды играют жизненно важную роль в качестве основных компонентов всех прокариотических и эукариотических клеток. Постоянные технологические усовершенствования в масс-спектрометрии сделали липидомику мощным аналитическим инструментом для мониторинга липидомных составов тканей как при гомеостатических состояниях, так и при болезненных состояниях. В этой статье представлен пошаговый протокол для метода анализа липидов дробовика, который поддерживает одновременное обнаружение и количественное определение нескольких сотен молекулярных видов липидов в различных образцах тканей и биожидкостей с высокой пропускной способностью. В этом методе используется автоматизированная нанопоточная прямая инъекция общего липидного экстракта, снабженного мечеными внутренними стандартами, без хроматографического разделения в масс-спектрометрический прибор высокого разрешения. Начиная с субмикрограммового количества ткани грызунов, анализ рассеянного склероза занимает 10 минут на образец и охватывает до 400 липидов из 14 классов липидов в легочной ткани мыши. Представленный здесь метод хорошо подходит для изучения механизмов заболевания, а также для выявления и количественного определения биомаркеров, которые указывают на раннюю токсичность или благотворное воздействие на ткани грызунов.
Введение
Сигаретный дым (КС) признан основным фактором риска, связанным с развитием хронических воспалительных заболеваний легких, включая карциному легких, бронхит и хроническую обструктивную болезнь легких (ХОБЛ)1. Помимо воздействия на легкие, воздействие КС играет важную роль в развитии других заболеваний, таких как атеросклеротическая ишемическая болезнь сердца и заболевание периферических сосудов1. Сердечно-сосудистые заболевания, наряду с ХОБЛ, являются первой и третьей по значимости причинами смерти во всем мире, соответственно. Подходы к оценке токсикологического риска исторически основывались на использовании моделей животных, таких как грызуны. Модели крыс и мышей in vivo, предназначенные только для носа или всего тела, обычно используются для изучения долгосрочных последствий воздействия CS.
Например, как правило, 6 месяцев воздействия дыма вызывают гистологические и функциональные нарушения в легких мышей, которые имитируют нарушения заболевания человека, включая эмфизему, ремоделирование дыхательных путей и легочную гипертензию, хотя изменения относительно мягкие по сравнению с теми, которые наблюдаются у длительно курильщиков2. Как в тканях животных, так и в тканях человека в исследованиях наблюдался широкий спектр молекулярных изменений в ответ на воздействие CS, включая реакции окислительного стресса, воспаление и структурные изменения тканей 3,4. Неудивительно, что воздействие CS также оказывает далеко идущее влияние на липидом легких, включая воздействие на липиды сурфактанта, липидные сигнальные медиаторы и структурные липиды 4,5,6.
Чтобы охарактеризовать объемные изменения липидов, вызванные длительным воздействием CS на легкие мышей, мы провели быстрый и количественный масс-спектрометрический анализ прямой инфузии дробовика. После введения метода липидомики дробовика в 2005году 7 этот метод был эффективно использован для расширения наших знаний о липидном клеточном метаболизме 8,9,10 в модельных системах, таких как дрожжи 11, Caenorhabditis elegans12 и Drosophila melanogaster13, а также в широком диапазоне типов образцов млекопитающих, таких как клеточные линии 14, 15,16 различные ткани человека17,18 и грызунов19,20 и биологические жидкости21,22.
За последние десятилетия исследования выявили сложность клеточных реакций на изменения окружающей среды, включая тысячи взаимосвязанных белков, липидов и метаболитов. Это ясно дало понять, что использование современных аналитических методов имеет важное значение для получения углубленного представления о молекулярных механизмах и для раскрытия полной величины экзогенных физиологических воздействий. В этом контексте всеобъемлющие количественные липидные отпечатки, полученные с помощью подходов к липидомике, могут эффективно дополнить наши знания о липидном клеточном метаболизме 8,9,10.
Что касается воздействия CS как фактора риска для ряда заболеваний, токсикологические подходы к оценке риска исторически основывались на использовании моделей животных, таких как грызуны. Дробовик липидомики МС предоставляет быстрый, чувствительный и количественный аналитический инструмент для оценки возмущения липидома в различных типах образцов. Уникальной особенностью липидомики дробовика является автоматизированный анализ с прямой инъекцией общего липидного экстракта с добавлением меченых внутренних стандартов без хроматографического разделения в прибор MS с высоким разрешением с использованием проводящего наночипа, генерирующего наноспрей с ионизацией электрораспылением (ESI)23.
Информация об отношении массы к заряду, которая одновременно получается в режиме MS1, обеспечивает общий липидный след всех интактных эндогенных липидов. Опционально режим MS2/MS3, в котором родительские липидные молекулы фрагментируются и анализируются, предоставляет дополнительную структурную информацию. Анализ данных требует специализированного программного обеспечения и включает в себя деконволюцию спектров и присвоение пиков в объединенных спектрах, что приводит к идентификации липидов и выяснению предполагаемой химической структуры. Кроме того, абсолютное количественное определение может быть выполнено путем добавления меченой смеси внутренних стандартов, содержащей, по меньшей мере, один липидный стандарт для каждого интересующего класса липидов. В целом, с помощью данной методики анализ РС, занимающий несколько минут на образец, может охватывать идентификацию и количественное определение до 800 липидов из 14 классовлипидов 24 в тканях грызунов.
протокол
Все процедуры с участием животных проводились на объекте, аккредитованном Ассоциацией по оценке и аккредитации Международной организации по уходу за лабораторными животными и лицензированном Агропродовольственным и ветеринарным управлением Сингапура, с одобрения Институционального комитета по уходу за животными и их использованию и в соответствии с Национальным консультативным комитетом по исследованиям на лабораторных животных Руководящие принципы по уходу за животными и их использованию в научных целях (NACLAR, 2004).
1. Сбор образцов
- Выполните расслоение легких мыши через 3 месяца и 6 месяцев воздействия мышей ApoE-/- . Соберите ткани через 16-24 часа после воздействия, заморозьте их в жидком азоте и храните при -80 ° C до начала рабочего процесса липидомики. Обеспечьте сбор в общей сложности девяти образцов из каждой группы вскрытия «омиксов».
2. Ткань - измельчение образцов
- Подготовьте магнитный молоток и расходные материалы для шлифовки тканей. Предварительно охладите тканевые мешочки, щипцы, закрытые трубки для переноса тканей с колпачками, пластиковый шпатель «V» и пластиковые пробирки для сбора 2 мл на сухом льду. Установите соответствующий держатель трубки на магнитный молотковый инструмент. Включите магнитный молоток и установите силу удара на Высокий.
- Загрузите образец в тканевый мешочек; При необходимости вставьте образец через верхнее отверстие тканевого мешочка с помощью предварительно охлажденных щипцов. Поместите образец в центр гибкого пакета, подальше от краев.
- Запечатайте тканевый пакет, навинчивая предварительно охлажденную трубку для сбора на верхнюю часть тканевого пакета.
- Заморозьте образец, погрузив гибкий мешочек в жидкий азот на 10 с.
- Вентиляция сборной трубки; Ослабьте трубку на 1/4 оборота для вентиляции, чтобы избежать разрыва пакета при ударе магнитного молотка.
- Загрузите замороженный тканевый мешочек на магнитный молоток.
- Примените магнитный молоток, нажав кнопку активации , чтобы измельчить образец. Если в пакете остались неизмельченные кусочки ткани, снова заморозьте образец в жидком азоте и повторите повторный удар.
- Извлеките тканевый мешочек из магнитного молотка, удерживая измельченный образец на дне. Чтобы образец не расплавился, немедленно заморозьте его снова.
- Перенесите образец в сборную пробирку. Быстро переверните пробирку так, чтобы тканевый мешочек оказался наверху, и постучите по пакету, чтобы перенести частицы ткани в нижнюю часть пробирки для сбора.
ПРИМЕЧАНИЕ: Этот шаг следует сделать быстро, чтобы избежать расплавления ткани. - Перенесите тканевую аликвоту 10 мг в пробирку объемом 2 мл для дальнейшего липидомного анализа и храните образец при -80 ° C до тех пор, пока не будут выполнены дальнейшие шаги.
3. Гомогенизация тканей
- Включите тканевый лизер и установите частоту встряхивания на 30 Гц и продолжительность встряхивания на 2 минуты. Приготовьте буферный раствор для гомогенизации тканей: 150 мМ бикарбоната аммония в воде с рН ~ 8,2.
- Выньте образцы из морозильной камеры, поместите на лед и добавьте четыре шарика из нержавеющей стали в пробирки с тканевым порошком. Добавьте 0,5 мл 150 мМ буфера бикарбоната аммония к каждому образцу.
- Поместите пробирки с образцами в держатели тканевых лизеров. Уравновесьте оба держателя одинаковым количеством трубок. Закрепите держатели в руке, удерживающей салфетный лизер. Разрушают ткани.
- Поместите пробирки на лед и визуально убедитесь, что в пробирке нет остатков тканевых агрегатов. Если разрушение тканей неполное (видны агрегаты), повторите процесс разрушения, выполнив еще один цикл лечения. Поместите образцы на лед.
- Создайте аликвоту 1 из 100 мкл образца в пробирке объемом 1,5 мл, предназначенной для определения белка. Центрифугируют его при 18 200 × г в течение 5 мин при 10 °С.
- Определите содержание белка аликвоты 1 с помощью анализа Брэдфорда (см. раздел 4). Храните образцы на льду.
- Создайте аликвоту 2 20-100 мкл исходного гомогената в новой микропробирке объемом 2 мл для экстракции липидов (см. раздел 5). Если образцы не анализируются немедленно, держите их на льду до тех пор, пока не будет выполнена экстракция.
ПРИМЕЧАНИЕ: Окончательный объем экстракции должен быть определен на основе общего количества белка. Она должна быть в пределах 0,015-0,045 мг белка на общую аликвоту.
4. Анализ определения белка Брэдфорда
ПРИМЕЧАНИЕ: Исследуемые образцы должны быть рандомизированы до начала анализа, и рандомизация соблюдается на всех этапах обработки образцов.
- Разбавьте надосадочную жидкость центрифугированной аликвотой 1 2x буфером из бикарбоната аммония.
ПРИМЕЧАНИЕ: Коэффициент разбавления зависит от концентрации тканевого гомогената. Для более концентрированных растворов применяют более высокий коэффициент, чтобы определенная концентрация попадала в динамический диапазон анализа. - Подготовьте стандартную кривую бычьего сывороточного альбумина (BSA) в соответствии с таблицей 1. Вихревая стандартные трубки. Центрифугируют пробирки при 18 200 × г в течение 15 с при комнатной температуре.
- Из предварительно подготовленных стандартных пробирок перенесите по 6 мкл каждой заготовки и стандартов на 96-луночную пластину с плоским дном в соответствии с расположением пластины, показанным на рисунке 1.
- Перенесите предварительно подготовленные образцы на 96-луночную плоскодонную плиту в соответствии с схемой плиты, описанной на рисунке 1.
- Добавьте 250 мкл реагента Брэдфорда в каждую лунку с помощью многоканальной пипетки и перемешайте. Инкубировать 5 мин.
- Измерьте коэффициент поглощения на длине волны 595 нм с помощью планшетного считывателя.
- Рассчитайте концентрацию белка в аликвотах на основе стандартной кривой.
ПРИМЕЧАНИЕ: Коэффициент разбавления, используемый в начале процедуры, следует учитывать при расчете концентраций белка.
5. Экстракция BUME
ПРИМЕЧАНИЕ: Избегайте использования пластиковых пробирок с низкой связью для экстракции липидомики, так как сильные органические растворители высвобождают пластификаторы и создают прочный фон во время анализа образца.
- Подготовьте пробирки объемом 1,5 мл со 100 мкл раствора бикарбоната аммония в каждой пробирке общей заготовки (TB), внутренней стандартной заготовки (ISB) и образцов контроля качества (QC), учитывая, что 1 образец QC помещается между каждым набором из 10 образцов. Храните все образцы на льду.
- Добавьте 10 мкл объединенной плазмы человека ко всем образцам контроля качества. Добавьте 10 мкл внутреннего стандартного раствора во все ISB, QC и исследуемые образцы (т. е. все образцы, кроме TB).
ПРИМЕЧАНИЕ: Для контроля качества используйте любую объединенную коммерчески доступную плазму K2EDTA. Количественно определите концентрацию липидов в объединенной плазме, используя упомянутый протокол при получении, и сохраните эти диапазоны в качестве эталона для всех анализов, в которых используется материал контроля качества. Используйте плазму NIST только для более целенаправленных применений или для глобальной проверки метода, где необходимо учитывать точность анализа. - Добавьте 300 мкл смеси бутанола/метанола с температурой -20 °C (3/1, об./об.) к каждому образцу. Взболтать при 450 × г в течение 10 мин при комнатной температуре на термомиксере.
- Добавьте 300 мкл смеси гептан/этилацетат (3/1, об./об.). Встряхните при 450 × г в течение 10 мин при комнатной температуре на термомиксере.
- Добавьте 300 мкл 1% смеси уксусной кислоты. Встряхните в течение 5 мин при температуре 450 × г при комнатной температуре на термомиксере. Центрифуга при 2 800 × г в течение 5 мин при комнатной температуре. Перенесите 360 мкл верхней фазы в новую самоблокирующуюся трубку 2 мл 2.
ПРИМЕЧАНИЕ: Жидкостно-жидкостная экстракция может привести к смещению положения границы фаз в зависимости от образца, что может привести к небольшому расхождению объема для верхней фазы. При необходимости отрегулируйте объем сбора для верхней фазы. - Добавьте 320 мкл гептана/этилацетата (3/1, об./об.) в пробирку 1 водной фазы. Взболтать при 450 × г в течение 5 мин при комнатной температуре на термомиксере. Центрифугу при 2 800 × г в течение 5 мин при комнатной температуре. Перенесите 320 мкл верхней фазы и соедините с фракцией из шага 5.11.
ПРИМЕЧАНИЕ: При необходимости отрегулируйте громкость сбора для верхней фазы. - Добавьте 250 мкл гептана/этилацетата (3/1, об./об.) в водную фазу в пробирке 1. Взболтать при 450 × г в течение 5 мин при комнатной температуре на термомиксере. Центрифуга при 2 800 × г в течение 5 мин при комнатной температуре. Перенесите 200 мкл верхней фазы и соедините с фракциями из шага 5.11 и шага 5.15 в пробирке 2.
ПРИМЕЧАНИЕ: При необходимости отрегулируйте громкость сбора для верхней фазы. - Выпарить досуха при 35 °C в вакуумном концентраторе.
ПРИМЕЧАНИЕ: Высушенные образцы можно хранить при температуре -20 °C до анализа, если это необходимо. - Повторно растворяют в 300 мкл раствора смеси МС (7,5 мМ ацетат аммония в изопропаноле/метаноле/хлороформе, раствор 4:2:1). Вкрутите каждую трубку в течение 5 с, чтобы убедиться, что все растворилось. Центрифуга при 18 200 × г в течение 5 мин при 4 °C.
- Перенесите аликвоту 50 мкл в микролитровую пластину в соответствии с расположением пластины на рисунке 2 для анализа положительной ионизационной моды (столбцы 1-6) и разбавьте 50 мкл раствора смеси MS.
- Переведите аликвоту 20 мкл в пластину MTP в соответствии с расположением пластины на рисунке 2 для анализа отрицательных ионизационных режимов (столбцы 7-12) и разбавьте 80 мкл смеси MS. Перед анализом оберните тарелку фольгой и держите при температуре 4 °C.
6. Анализ рассеянного склероза
- Калибруйте масс-спектрометр (МС) как в положительном, так и в отрицательном режимах в соответствии с инструкциями производителя не более чем за 5 дней до анализа.
- Установите наноисточник прямой инфузии заранее, убедившись, что он правильно выровнен по транспортному капилляру MS. Установите чип сопла 4,1 мкм в держатель микросхемы и настройте его в программном обеспечении.
- Для настройки метода положительного и отрицательного режимов используйте следующие параметры: давление газа 1,25 фунта на квадратный дюйм; напряжение источника 1,1 кВ; 5 мкл объема впрыска образца; замыкание выходного контакта Rel1 на 2,5 с; 5 мин времени доставки; охлаждение пластины при 4 °C.
- Настройте очередь последовательности анализа для робота прямой инфузии в положительном режиме.
- Настройте метод MS для положительного режима , используя следующие настройки MS:
- Установите температуру капилляров на уровне 250 °C и уровень RF объектива S-lens на уровне 65,0. Выполняйте полное сканирование MS от 0 до 1 минуты в диапазоне 550-1000 м/з с разрешением 140 000 с автоматической регулировкой усиления 1 ×106 и максимальным временем впрыска 50 мс. Применим замок массой 680.48022.
- Установите независимый от данных метод сбора данных MS/MS в диапазоне времени от 1 до 5 минут с разрешением 17 500 с фиксированной первой массой 250 м/z. Используйте автоматическую регулировку усиления для MS2 при 1 × 105 и максимальном времени впрыска 64 мс, энергии столкновения 20 NCE и окне изоляции 1 м/z. Используйте список масс включения от 550 до 1 000 м/z с шагом массы 1 Да.
- Для съемки в отрицательном режиме установите температуру капилляров на уровне 250 °C и уровень RF объектива S-Lens на уровне 65,0 в файле настройки MS.
- Используйте следующие настройки метода MS: режим полного сканирования от 0 до 1 минуты при разрешении 140 000 м/з, охватывающий диапазон 400-940 м/з, автоматическая регулировка усиления 1 × 106; максимальное время впрыска 50 мс; Используйте массу замка 529.46262.
- Установите сбор данных MS2 в режиме DIA между 1 и 5 минутами прогона с разрешением 17 500 с фиксированной первой массой 150 м/z; автоматизированная регулировка усиления 1 × 105; 64 мс максимального времени впрыска; и 35NCE энергии столкновения. Используйте список масс включения от 400 до 940 с шагом массы 1 Да.
- Создайте очередь последовательности анализа в программном обеспечении MS в положительном режиме. Подождите, пока сбор образца не будет инициирован сигналом замыкания контакта, поступающим от робота прямой инфузии для каждого образца.
- Когда анализ положительного режима будет завершен, создайте очередь последовательности для робота прямой инфузии в отрицательном режиме.
- Настройте очередь последовательности анализа в программном обеспечении MS в отрицательном режиме.
7. Обработка данных
- Чтобы преобразовать файлы данных из положительного и отрицательного режимов из .raw формата в формат .mzML, используйте программное обеспечение конвертера.
ПРИМЕЧАНИЕ: Файлы данных из положительного и отрицательного режимов должны быть преобразованы отдельно (из-за их разных настроек захвата и разного порога шума).- Заполните следующие настройки, чтобы выполнить преобразование файлов: коэффициент порога шума равен 3 и 5 для MS и MS2 соответственно; активируйте функцию удаления шума как для MS, так и для MS2, сжатие данных, среднее сканирование MS2 и пиковый выбор. Установите пороговые значения TIC для положительных и отрицательных мод (например, 10 000 и 5 000; должны быть определены на основе уровня шума конкретного образца, производимого конкретным прибором). Генерируйте отчеты XLC и PDF в конце обработки.
- Выполните идентификацию липидов. Определите следующие (пример) параметры импорта файла: окно выделения ±0,5 Да (зависит от окна выбора в методе MS); временной диапазон 2-300 с (зависит от времени сканирования в методе MS); отсутствие калибровочных масс для МС и МС2; передавать и округлять в большую сторону диапазон масс из файлов метаданных программного обеспечения Peak by Peak (min mass [MS]) и (min mass [MS2]); допуск 3 ppm и 10 ppm для MS и MS2 соответственно; держите пустым пороговое поле MS и MS2, частотный фильтр, смещение MS1 и PMO; Выбрана изотопная коррекция для МС и МС2; передавать градиент разрешения из файла метаданных конвертера как (Разрешение линейной подгонки [MS]) и как (Разрешение линейной подгонки [MS2]).
ПРИМЕЧАНИЕ: Идентификация липидов в положительном и отрицательном режимах должна выполняться отдельно из-за их различных настроек приобретения. - После того, как данные будут импортированы, перейдите в меню «Выполнить » и загрузите файлы MFQL для идентификации липидов.
ПРИМЕЧАНИЕ: Для получения подробной информации о структуре MFQL см. публикацию Herzog et al.25. Ознакомьтесь с некоторыми примерами файлов MFQL на https://wiki.mpi-cbg.de/lipidx/MFQL_library и ознакомьтесь с обсуждением. Все MFQL, используемые для обработки данных, представлены в Дополнительных материалах. - Количественно определите идентифицированные виды на уровне МС путем деления интенсивности массы предшественника липидного признака на соответствующую интенсивность меченого внутреннего стандарта, а затем умножения частного на концентрацию меченого внутреннего стандарта. Нормализуйте конечную концентрацию липидов на количество общего белка, измеренное с помощью анализа Брэдфорда.
8. Процедура проверки качества
ПРИМЕЧАНИЕ: Процедура проверки качества является важным шагом для проверки технической воспроизводимости метода. С этой целью коммерческую объединенную плазму анализируют для определения эндогенных уровней различных липидов в течение 5 дней в 15 идентичных аликвотах на партию (всего пять партий). Обратите внимание, что в этом случае конвейер оценки данных был создан в виде веб-приложения Shiny с использованием статистической программной среды R.
- Определите эталонный диапазон приемки как среднее значение, рассчитанное для всех пяти партий ±3 стандартными отклонениями.
- Применяйте правила приемки «пройдено» для каждой аналитической партии: 90% референтных мишеней должны пройти, учитывая, что одно эталонное соединение представляет один класс липидов. Например, если в проверку контроля качества включено 15 эталонных целей, убедитесь, что 13 целей проходят для принятия партии.
ПРИМЕЧАНИЕ: Другими словами, 68% образцов контроля качества на эталонное соединение должны пройти для принятия данных для этого класса липидов. - Если исследуемая партия не соответствует критериям приемки для проверки контроля качества, повторите процедуру.
9. Оценка (относительных) количеств конъюгированных жирных кислот
- Для каждого класса липидов оцените количество конъюгированных жирных кислот на основе их интенсивности MS2.
ПРИМЕЧАНИЕ: Из-за различий в фрагментации и ионизации полученные значения предназначались только для сравнения относительной численности между группами выборки, а не, например, для оценки относительного вклада конкретной жирной кислоты в общий пул конъюгированных жирных кислот класса липидов. - Нормализуйте интенсивность MS2 фрагментов жирных кислот на среднюю интенсивность фрагментов жирных кислот соответствующего внутреннего стандарта. Основываясь на концентрации внутреннего стандарта и измеренной массе ткани, выведите «нормированную концентрацию» для конъюгированных жирных кислот. Суммируйте эти значения по классу липидов, чтобы оценить общее количество каждой конъюгированной жирной кислоты на образец.
10. Статистический анализ
- Проведите статистический анализ. Подгонка линейной модели для каждого условия и соответствующей контрольной группы и вычисление значений p на основе t-статистики для данных, преобразованных в log2. Используйте метод частоты ложных открытий Бенджамини-Хохберга для коррекции нескольких эффектов тестирования.
ПРИМЕЧАНИЕ: Липиды со скорректированными значениями p < 0,05 считаются дифференцированно обильными. Здесь статистический анализ проводился в статистической среде R26.
Результаты
Здесь, в качестве репрезентативных результатов, мы представляем данные, иллюстрирующие, как липидомика дробовика может быть использована для изучения влияния CS на легкие мышей. Протокол анализа липидов дробовика был успешно применен для исследования in vivo для сравнительного анализа двух групп образцов: тканей легких, рассеченных у девяти самок мышей Apoe-/-, подвергшихся воздействию CS (группа 3R4F; положительный контроль), и равного количества мышей, содержащихся в условиях свежего воздуха (фиктивная группа) в качестве отрицательного контроля, собранного через 3 месяца, 4 месяца и 6 месяцев времени воздействия. Животные содержались под фильтрованным и кондиционированным свежим воздухом при температуре 22 °C ± 2 °C и влажности 55% ± 15% и питались гранулированным диетой с гамма-облучением. Световой режим поддерживался на уровне 12 ч в сутки. В каждой клетке содержалось до восьми мышей. Эталонные сигареты 3R4F для обработки воздействия были приобретены в Университете Кентукки для получения аэрозоля 3R4F CS (600 мкг общего аэрозоля твердых частиц TPM/L) для целевой концентрации воздействия, эквивалентной 28 мкг никотина/л.CS дым от сигарет 3R4F производился с использованием 30-портовых ротационных коптильных машин, согласно Phillips et al.27 . В соответствии с Протоколом интенсивного курения Министерства здравоохранения Канады к сигаретам 3R4F применялось 55 мл объема затяжки, одна затяжка в 30 с и 100% закупорка вентиляционных отверстий, чтобы обеспечить достаточное воздействие. Все дополнительные подробности о дизайне исследования были сообщены ранее27,28.
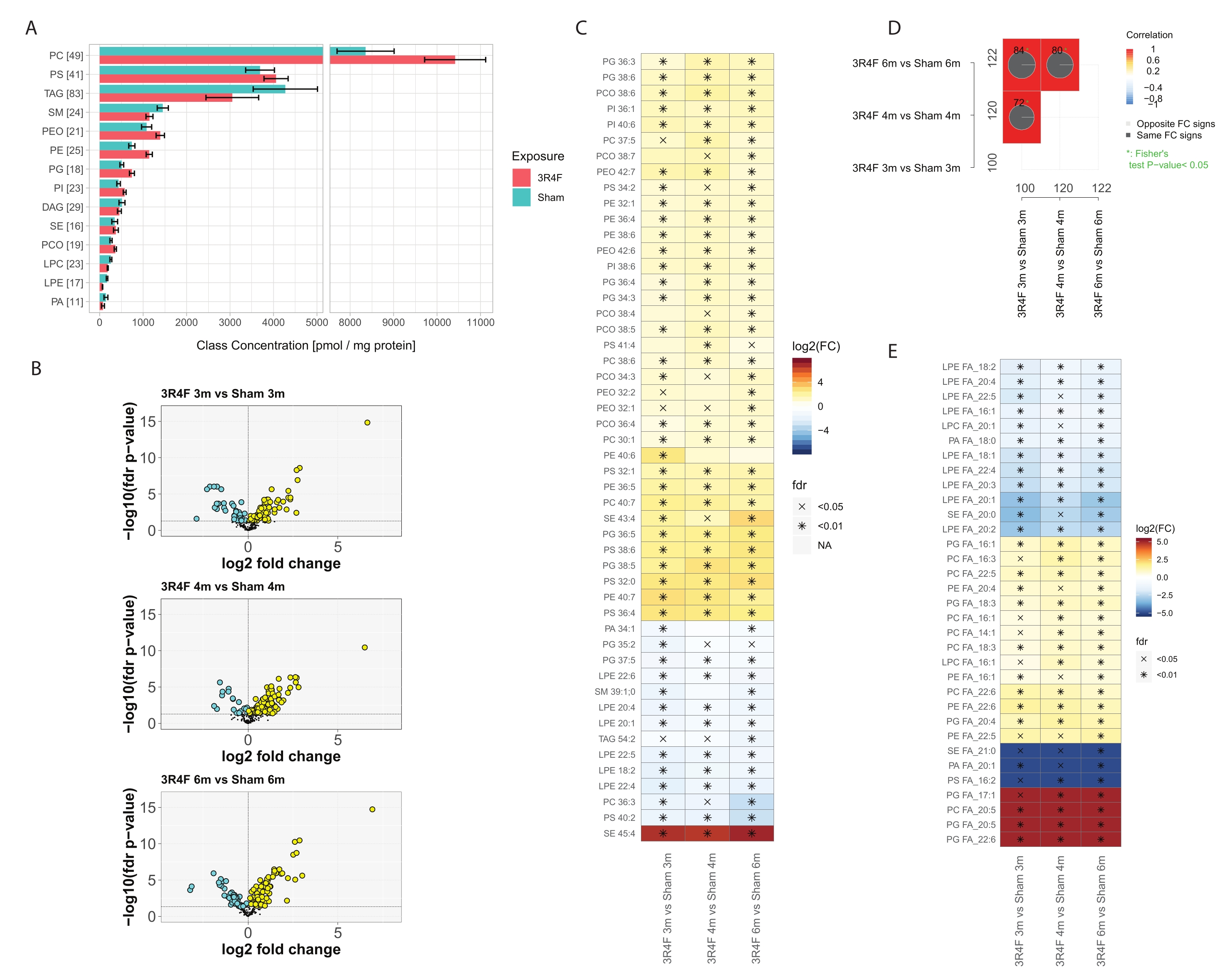
В целом, ~ 400 молекулярных видов липидов из 14 наиболее распространенных классов липидов были идентифицированы и количественно оценены (рис. 3). Общая нормализованная концентрация липидов по классу была немного повышена для классов липидов PE, PEO, PC, PCO, PI и PG, и некоторое снижение наблюдалось для липидов SM, LPE и PA (рис. 3A), в то время как для TAG и PS различий не наблюдалось. Интересно, что повышенные уровни липидов плазмалогена PCO и PEO в группе CS были зарегистрированы в воспалительных клетках29 . Одной из внутриклеточных функций липидов плазмалогена является антиоксидантная активность. При воздействии активных форм кислорода (АФК) и азота (РНС) сообщалось о селективном окислении плазмалогенов по сравнению с диацилфосфолипидами30. Связь с енил-эфиром в химической структуре плазмалогенов чувствительна к радикальной атаке на окислительный стресс31. Основными продуктами радикальной атаки являются гидроксилированные эйкозатетраеновые кислоты, фосфолипиды 2-моноацилглицерина, пентадеканол, муравьиные кислоты, α-гидроксиальдегиды различной длины цепи, 1-формил-2-арахидоноилглицерофосфолипиды и лизофосфолипиды.
Эти результаты показывают, что среди молекулярных особенностей, основанных на общей молекулярной формуле, воздействие CS значительно повлияло на 100-120 соединений (рис. 3D). Детальная деконволюция информации о фрагментации MS2 и нормализация интенсивности сигнала фрагмента позволили приблизительно определить общее количество каждой конъюгированной жирной кислоты (ЖК), представленной в каждом классе липидов (рис. 3E), и сравнить эти данные между группами, подвергшимися воздействию, и контрольной группой. Наблюдалось явное снижение как полностью насыщенных ЖК, так и ненасыщенных ЖК, в основном в контексте лизолипидов. Напротив, полиненасыщенные жирные кислоты (ПНЖК) в составе классов фосфолипидов PC, PE и PG были повышены в группе CS для всех временных моментов. Крайние случаи ПК и ПГ, конъюгированные с эйкозапентановой (ЭПК) и докозагексаеновой кислотой (ДГК), были обнаружены исключительно в группе воздействия 3R4F CS (рис. 3E, красный цвет).

Рисунок 1: Расположение планшета для распределения образцов в анализе Брэдфорда. Заготовки и стандартные образцы размещаются в трех экземплярах в графах 1-3; Неизвестные образцы помещаются в двух экземплярах в графах 4-11. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2: Расположение 96-луночного микротитровального планшета для масс-спектрометрического анализа дробовика. Распределение холостых, стандартных, неизвестных и КК образцов для сбора в режиме положительной ионизации отображено на левой стороне планшета (столбцы 1-6); Все образцы для режима отрицательной ионизации размещены справа (столбцы 7-12). Сокращения: pos = положительный; QC = контроль качества; neg = отрицательный; TB = общее пустое. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3: Анализ липидомики легких мышей, подвергшихся воздействию сигаретного дыма. Мыши Apoe-/− подвергались воздействию CS от эталонной сигареты 3R4F или свежего воздуха (фиктивный). А) Концентрации в классе липидов и количество видов липидов в каждом классе. Для каждого класса в скобках указано количество количественных видов липидов. Средние и 95% доверительные интервалы для каждого класса липидов, отдельно для каждого типа воздействия (агрегированные по трем временным точкам). (B) Графики вулканов, показывающие величину эффекта (изменение log2 раза) и значимость (значение p с поправкой на -log10 FDR) количественных видов липидов для сравнения 3R4F CS и фиктивного в три момента времени. Значительно повышенные липиды отмечены желтым цветом; значительно сниженные липиды отмечены в голубом цвете (скорректированное FDR значение p < 0,05). (C) Дифференциальная численность 25 видов липидов с наибольшими абсолютными средними изменениями складок. Изменения log2 fold для сравнения 3R4F CS и фиктивного сравнения имеют цветовую кодировку, и указывается статистическая значимость: *: значение p с поправкой на FDR < 0,01; X: значение p с поправкой на FDR < 0,05. (D) Сравнительные графики. Коэффициенты корреляции для сравнений с кратным изменением имеют цветовую кодировку, и указывается количество общих дифференциально распространенных липидов (общие числа на полях). Круговые диаграммы показывают процент общих дифференциально распространенных липидов с одинаковым направлением изменения (знак FC). Звездочка указывает на значительное перекрытие дифференциально обильных липидов. (E) Дифференцированное содержание конъюгированных ЖК по классу липидов. Тепловая карта, как показано на панели C , для конъюгированных жирных кислот со значительными различиями в численности между 3R4F и фиктивным воздействием во всех трех временных точках (скорректированное FDR значение p < 0,05). Количество каждой жирной кислоты на класс липидов оценивали на основе интенсивности MS2 фрагментов жирных кислот. Сокращения: CS = сигаретный дым; FDR = частота ложных обнаружений; FC = изменение крата; FA = жирные кислоты; ПК = фосфатидилхолин; PS = фосфатидилсерин; TAG = триацилглицерин; SM = сфингомиелин; ПЭО = плазмалоген фосфатидилэтаноламин; ПЭ = фосфатидилэтаноламин; PG = фосфатидилглицерин; PI = фосфатидилинозитол; DAG = диацилглицерин; SE = эфиры стерина/холестерина; PCO = плазмалоген фосфатидилхолин; ЛПК = лизофосфатидилхолин; LPE = лизофосфатидилэтаноламин; PA = фосфатидная кислота. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
| Конечная концентрация БСА (мг/мл) | Раствор BSA в дозе 2 мг/мл (мкл) | Буфер на основе бикарбоната аммония (мкл) |
| 1 | 50 | 50 |
| 0.75 | 37.5 | 62.5 |
| 0.5 | 25 | 75 |
| 0.25 | 12.5 | 87.5 |
| 0.125 | 12.5 | 187.5 |
| 0.0625 | 50 раствора 0,125 мг/мл | 50 |
| 0 | 0 | 100 |
Таблица 1: Схема разбавления внутреннего стандарта BSA для калибровочной кривой для анализа Брэдфорда. Аббревиатура: BSA = бычий сывороточный альбумин.
Дополнительная информация: Файлы MFQL, используемые для идентификации липидов LipidXplorer. Пакет .zip состоит из отдельных обязательных файлов MFQL, каждый из которых именуется следующим образом: <аббревиатура класса липидов>_<режим ионизации>_MS2.mfql (например, PE_neg_MS2.mfql). Имена файлов для маркированной внутренней стандартной идентификации включают тег D7 (D9) в <аббревиатуру липидов> (например, PED7_neg_MS2.mfql). Эти файлы MFQL используются для проверки количества дейтерия во внутреннем стандарте. Пожалуйста, нажмите здесь, чтобы загрузить этот файл.
Обсуждение
Несмотря на то, что достижения в области рассеянного склероза привели к появлению различных методов точного мониторинга многих видов липидов, предыдущее липидное профилирование различных тканей млекопитающих не показало последовательных результатов, и, следовательно, конкретные тканеспецифические функции липидов остаются неясными. По сравнению с функциональным анализом белков, для которого доступны более надежные подходы к нокаутированию экспрессии конкретных соединений, большинство липидов не могут быть селективно отключены или сверхэкспрессированы в тканях, что затрудняет функциональную оценку липидов. Расширенное профилирование концентраций липидомов в тканях может обеспечить альтернативный подход к выявлению связей между циркулирующими липидами и заболеваниями человека.
При оценке комплексных методов липидомики, способных качественно и количественно охватить эндогенный липидный профиль любой ткани мыши, мы отдали предпочтение методу дробовиковой липидомики. В целом, возможны два противоположных типа анализа образцов: либо полностью нецелевой скрининг липидов с использованием разделения на основе жидкостной хроматографии (ЛХ) с дальнейшим масс-спектрометрическим обнаружением для выявления общей липидной сложности образца, либо целевые подходы, которые в основном позволяют очень точно определить конкретные липиды, представляющие интерес. В отличие от этого, сильной стороной представленного рабочего процесса липидомики является быстрый всесторонний охват сотен эндогенных липидов из заранее определенных классов липидов, который все еще может быть выполнен надежным полуколичественным образом.
Эффективность ионизации различных классов липидов зависит от структуры и может сильно варьироваться в зависимости от различных условий экспериментальной ионизации. В отличие от методов разделения на основе LC, анализ дробовика сводит к минимуму эти различия за счет прямой одновременной инфузии всего липидного экстракта в тех же условиях ионизации в прибор MS. Использование изотопно меченных аналогов липидов или структурно сходных неэндогенных стандартов позволяет проводить полуколичественную оценку всех классов липидов. Дробовиковая липидомика обеспечивает низкую меж- и внутрипрогонную вариабельность при анализе МС; В результате этот метод дает более низкие коэффициенты вариации21 , чем методы, основанные на нецелевой жидкостной хроматографии, которые требуют нескольких стандартов для адекватного количественного определения. Важно отметить, что, хотя в текущем методе не используется внешняя калибровочная кривая, метод по-прежнему считается полностью количественным32.
Один уровень меченого внутреннего стандарта (или немеченого стандарта, который не экспрессируется эндогенно) для каждого класса липидов достаточен для количественной оценки большинства липидов. Лишь в нескольких публикациях сообщалось о частичной валидации метода липидомики дробовика. Например, в работах Gryzbek et al.17 и Surma et al.21 были подготовлены обратные калибровочные кривые с использованием внутренних стандартов и матрицы с фиксированным количеством образцов. Линейность оценивали с помощью линейной регрессии логарифмически преобразованных количеств липидов и их интенсивности и сообщали как R2 и наклон соответственно. Предел обнаружения (LOD) и предел количественного определения (LOQ) были определены с помощью взвешенной линейной регрессии, основанной на отношении сигнал/шум 3 для LOD и 10 для LOQ. Для большинства классов липидов LOQ определялся между 2-9,8 пмоль для жировой ткани и 0,05-5 мкМ в плазме. В обоих случаях для получения оценок для всех липидов в классе использовались неэндогенные единичные внутренние стандарты для каждого класса. Однако в этой работе мы не предоставляем LOD/LOQ из-за нескольких проблем: эндогенная матрица не свободна от соединений, а суррогатная матрица для тканей недоступна - при этом всплеск небольших известных количеств стандартов невозможен. Мы не применяем классический подход к таргетной количественной оценке с использованием ряда калибровочных кривых конкретного соединения, нормализованного его идентичным изотопно меченным стандартом, из-за отсутствия чистых эталонов и очень ограниченной доступности изотопно меченных липидов. Орбитальные детекторы автоматически выполняют преобразование переходного сигнала, применяя преобразование Фурье, и некоторый сигнал уже подставляется - в результате нижний диапазон концентраций будет линейным только до некоторого минимального сигнала, ниже которого молекула больше не будет обнаруживаться. Значения сигнал/шум программного обеспечения Xcalibur зависят от отношения m/z молекулы; В результате соединения каждого класса липидов, содержащие различные комбинации жирных кислот, будут иметь разные значения шума. Кроме того, значения LOQ/LOQ строго связаны с типом матрицы, и когда количественная оценка липидома проводится в различных тканях грызунов, это должно быть отражено оценкой LOQ для каждого типа ткани в отдельности.
Фактически, метод предлагает высокий диапазон линейной динамической количественной оценки до четырех порядковвеличины 33 и очень хорошую чувствительность для охвата наиболее важных эндогенных структурных липидов, которая может быть дополнительно увеличена за счет технических усовершенствований в получении МС32. Коэффициенты вариации средних концентраций липидов были в основном ниже 15%, что означает, что липидомика дробовика соответствует требованиям Управления по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) в качестве метода, который следует рассматривать для исследований надлежащей лабораторной практики (GLP) и надлежащей клинической практики (GCLP)34.
Однако следует отметить, что из-за их разной полярности на некоторые классы липидов гораздо больше влияет вклад специфических конъюгированных ЖК. Это приводит к искажению интенсивности реакции в эквимолярных смесях, содержащих широкий спектр сопряженных ЖК, что приводит к смещению количественного определения, как подчеркивается Koivusalo et al.35 для классов фосфолипидов. Следует отметить, что эти авторы представили данные для широкого диапазона ЖК длиной цепи от 24 до 48, что, вероятно, не отражает ситуацию в реальном биологическом образце. Реакция для полиненасыщенных видов была на 40% выше, чем для полностью насыщенных видов, но этот эффект наблюдался только для более высоких концентраций. Когда смесь постепенно разбавляли, эффект ненасыщенности постепенно уменьшался и практически исчезал при 0,1 пмоль/мкл на вид. Кроме того, все измерения проводились на приборах с ионными ловушками и тройными квадруполями, а не на оборудовании Q-Exactive.
Еще одним преимуществом представленного рабочего процесса является его техническая гибкость, которая позволяет адаптироваться к конкретным требованиям проекта. Например, любой протокол экстракции липидов, такой как модифицированные методы Блая и Дайера36, метил-трет-бутилового эфира 37 или бутанол-метанола38, может быть использован для получения общего липидного экстракта перед анализом МС. Основным ограничением хлороформ-метанольной экстракции является то, что нижняя фаза содержит липидную фракцию, что усложняет рутинную работу и особенно автоматизацию; Кроме того, необходимо учитывать токсичность хлороформа. Экстракция трет-бутилового эфира широко используется для липидного анализа образцовплазмы 37, и был предложен автоматизированный вариант21. В данном случае мы выбрали метод BUME, поскольку он обеспечивает еще лучшее восстановление для шипованных липидов классов38 PG, PI, PA и PS, более низкий расход растворителя и возможность автоматизации39, в то время как общие концентрации, количественно определенные для образцов тканей, извлеченных всеми тремя методами, были сопоставимы. Кроме того, несмотря на то, что в текущей работе отбор проб выполнялся вручную, также можно получить воспроизводимые и точные результаты из больших наборов проб путем автоматизированной пробоподготовки и экстракции липидов в 96-луночном формате40,41. Это позволяет внедрять липидомный анализ в масштабные клинико-токсикологические исследования.
В текущей работе мы выполнили получение МС положительных и отрицательных мод отдельно без переключения полярности, как описано Schuhmann et al.42. Стабильность сигнала наномата лучше в отрицательном режиме для чуть менее концентрированного раствора, чем в положительном режиме. Кроме того, мы разработали полностью прослеживаемую процедуру с помощью программного преобразователя файлов .raw в mzML, которая предоставляет значения, указываемые в программном обеспечении LipidXplorer - при этом не требуются ручные расчеты наклона разрешения. Мы также применили различные подстановки настроек шума, потому что в положительном режиме уровни шума спектров выше, чем в отрицательном режиме. Все этапы были оптимизированы для прослеживаемого и высокопроизводительного рутинного анализа.
Для идентификации анализ липидомики дробовика может использовать уникальное поведение различных классов липидов, которые образуют уникальные аддукты в разных режимах полярности. В этом методе частицы ПК и ПЭ с одинаковой молекулярной массой, которые перекрываются в режиме положительной ионизации, могут быть полностью разделены в режиме отрицательной ионизации, поскольку ПК образует аддукты ацетата или формиата, а ПЭ ионизируется в депротонированной форме. Кроме того, возможна (частичная) структурная деконволюция для метода, использующего не только молекулярную формулу, но и объемную структуру жирных кислот. Например, идентификация FA по уровню общего углерода и общему количеству ненасыщенных работает для всех фосфолипидов, DAG, TAG и лизофосфолипидов. Количественное определение каждой изомерной формы снизу вверх может быть частично выполнено для некоторых классовфосфолипидов 43 , но является гораздо более сложным для DAG и TAG из-за неодинакового сигнального отклика различных FA в спектрах MS2.
Важно также подчеркнуть необходимость внедрения соответствующих процедур контроля качества, полностью согласующихся с недавними инициативами в этой области44. Поскольку мы хотим обеспечить надлежащую прослеживаемость и воспроизводимость данных между лабораторией и внутри нее, был предпринят ряд шагов, таких как надлежащая рандомизация образцов для всех этапов анализа, работа со стандартными смесями, сертифицированными поставщиками, включение образцов контроля качества, процедуры проверки приемки или отбраковки партии, а также создание внутренней базы данных для отслеживания эффективности контроля качества в долгосрочной перспективе. Кроме того, этим инициативам соответствует потребность в стандартизированном методе решения проблемы стабильности образца. В целом, большинство структурных липидов не подвержены окислению липидов, что более актуально для оксилипинов, окисленных липидов и полиненасыщенных жирных кислот, которые, следовательно, гораздо более чувствительны к условиям хранения и обработки. Однако правильная оценка стабильности образца по-прежнему является технически сложной задачей.
Однако этот протокол имеет ограничение, когда речь идет об определении субмолекулярных уровней соединений. Учитывая, что нет разделения общего экстракта, все изоформы липидов с одинаковой молекулярной массой, но разным составом жирных кислот объединяются в анализе MS. Для большинства классов можно добиться частичной деконволюции структуры с помощью коэффициентов фрагментации остаточных фрагментов жирных кислот в MS2. Однако независимая количественная оценка каждой изоформы остается особенно сложной задачей из-за больших различий в фрагментационном поведении различных изоформ и того факта, что чистые химические стандарты не доступны в достаточно большом разнообразии для определения значений компенсации. Другим ограничением является то, что процесс ESI неизбежно генерирует артефакты, что приводит к искусственной генерации пиков для некоторых липидов, таких как DAG, Pas и FA, что может привести к ошибочной количественной оценке.
Далее мы суммируем наиболее важные части протокола, основанные на нашем опыте. Первый связан с тем, что каждый тип ткани мыши имеет уникальный липидный профиль как с точки зрения количества липидов, так и с точки зрения соотношения классов. При этом исходные количества ткани на основе общего содержания белка перед экстракцией должны быть тщательно определены, чтобы не насыщать сигнал МС и не выходить из динамического диапазона количественного определения из-за агрегации липидов при высоких концентрациях33 или - в противоположном полюсе - обеспечить достаточный сигнал МС для охвата основных липидных соединений для каждого класса липидов.
Вторым важным аспектом является обеспечение правильного совмещения положения выходного отверстия чипа наноисточника прямой инфузии с трансферным капилляром масс-спектрометра. Учитывая, что полная калибровка масс-спектрометра в обоих режимах выполняется еженедельно, переключение между калибровочным источником и настройкой чипа наноисточника может быть причиной резкой изменчивости интенсивности сигнала из-за смещения во время установки.
Еще одной важной частью протокола является тщательное обращение с внутренним набором стандартов. Поскольку эта смесь содержит значительное количество дихлорметана, после вскрытия ее следует быстро употребить, чтобы избежать длительного хранения и многократного использования, приводящего к испарению и искусственному изменению концентрации. Кроме того, важно последовательное обращение со стандартной смесью после извлечения из хранилища при температуре −20 °C, поскольку перепады температур могут привести к несоответствиям объема во время пипетирования пипетками на воздушной подушке. Один из вариантов может заключаться в замене дихлорметана в стандартном буфере ресуспендирования чистым метанолом, что могло бы повысить удобство обработки, но могло бы отрицательно сказаться на растворимости некоторых классов липидов и, таким образом, снизить точность количественной оценки этих классов липидов.
Последней важной частью является обработка данных. Рабочий процесс обработки данных сочетает в себе программное преобразование Peak by Peak из формата .raw в формат .mzML, применение усреднения сканирования MS2 и фильтрации шума MS1 и MS2, а также пикового выбора и сжатия данных. В качестве альтернативы программное обеспечение Proteowizard также можно использовать для преобразования данных, но в этом случае несколько настроек в LipidXplorer должны быть определены вручную. Вся сложность липидомики дробовика особенно сконцентрирована на этапе деконволюции спектров прямого впрыска MS1 и MS2. Программное обеспечение LipidXplorer с открытым исходным кодом импортирует преобразованные спектры из формата файла mzML на основе точности массы, массового разрешения и наклона его изменения с увеличением m/z. Программное обеспечение объединяет несколько отдельных спектров МС и МС / МС, полученных в ходе анализа. После этого он выравнивает отдельные пики в разных прогонах выборки, и в каждом кластере выровненных пиков он заменяет их массы их единой взвешенной по интенсивности средней массой, в то время как их содержание в каждом файле данных остается нетронутым. Репрезентативные массы выровненных кластеров пиков и отдельные пиковые интенсивности хранятся в базе данных главного сканирования. База данных главного сканирования содержит все спектры MS1 и MS2, сгенерированные для всех образцов в партии, и может быть деконволютирована для идентификации липидов с помощью запросов, написанных на языке запросов молекулярной фрагментации (MFQL).
В целом, метод охватывает идентификацию липидов DAG, TAG и SE на основе положительной моды и липидов PC, PE, PS, PI, PA, PG, SM, LPC и LPE на основе получения отрицательной моды. Во время идентификации липидов проводится изотопная коррекция для MS1 и MS2, а скорректированные интенсивности сообщаются в выходном файле .xlsx. В качестве альтернативы доступно несколько других программ для обработки данных дробовика, таких как ALEX45 и LipidHunter46.
Жирные кислоты - основные ядерные и клеточные мембранные компоненты - хранятся в виде фосфолипидов для дальнейшего превращения в биологически активные молекулы. Они могут быть преобразованы ферментами лизофосфолипидацилтрансферазы в лизофосфолипиды через путь47 Лендса. Например, известно, что фермент LPCAT3 демонстрирует высокую специфичность для включения АА в промежуточные продукты лизофосфатидилхолина и лизофосфатидилсерина. Относительно высокая экспрессия этих ферментов была зарегистрирована в воспалительных клетках, таких как альвеолярные макрофаги и эпителиальные клеткибронхов 47, что приводило к высвобождению больших относительных пропорций АА-содержащих фосфолипидов в этих клетках. Однако после их высвобождения из мембранных фосфолипидов фосфолипазой А2 превращение ПНЖК в различные типы липидных медиаторов катализируется многими ферментами48. Кроме того, эти ПНЖК могут стать субстратом для производства провоспалительных и противовоспалительных простагландинов посредством действия ферментов циклооксигеназы (ЦОГ)-1 и ЦОГ-247. Фосфолипиды являются одним из источников липидных медиаторов, высвобождаемых в определенных местах, где эти медиаторы должны демонстрировать свои биологические эффекты.
Легкое представляет собой сложный орган, состоящий из нескольких типов клеток, каждый из которых играет перекрывающуюся и нишевую роль в содействии нормальному развитию и функционированию легких. Было проведено всего несколько исследований для выделения и профилирования различных типов клеток в легких мыши или человека (например, альвеолярных клеток 2-го типа)49,50; Другие основные типы клеток легких не были охарактеризованы. Еще одно интересное исследование51 было проведено для выделения и проведения липидного анализа эндотелиальных, эпителиальных, мезенхимальных и смешанных иммунных клеток в легких мыши. Было замечено, что концентрация ПНЖК, включенных в PCO и PG, была обогащена в иммунных клетках. Учитывая тот факт, что CS индуцирует увеличение иммунных клеток в легких (в 4-5 раз), что оценивается с помощью бронхоальвеолярного лаважа (BAL)52, общие изменения липидов, наблюдаемые в этом исследовании, могут быть объяснены кумулятивным увеличением рекрутирования иммунных клеток в легких мыши.
В заключение, наблюдаемое искажение мононенасыщенных жирных кислот и ПНЖК, включенных в фосфолипиды, может отражать избыток определенных ЖК в клетке и/или представлять собой внутриклеточный ресурс предшественников оксилипина, которые перепродуцируются при окислительном стрессе и воспалительных состояниях. Тем не менее, дополнительные данные о свободных ЭПК, ДГК и других оксилипинах необходимы для прояснения этого момента и выходят за рамки применения текущего метода.
Раскрытие информации
Все авторы являются сотрудниками компании «Филип Моррис Интернэшнл». Компания «Филип Моррис Интернэшнл» является единственным источником финансирования и спонсором этого проекта.
Благодарности
Авторы хотели бы поблагодарить исследовательскую группу и особенно отметить техническую помощь и поддержку групп биоисследований и аэрозолей в PMI R&D, Philip Morris International Research Laboratories Pte. Ltd., Сингапур, и PMI R&D, Philip Morris Products S.A., Невшатель, Швейцария. Авторы благодарят Сэма Ансари за управление биобанкингом и выражают признательность за поддержку Синдхуре Бхаргави Гопале Редди за редактирование черновика рукописи.
Материалы
| Name | Company | Catalog Number | Comments |
| 1, 5, and 2 mL self-lock tubes | Eppendorf | 30120086, 30120094 | |
| 3 mm stainless still beads | Qiagen | 69997 | |
| 4.1 µm nozzle chip | Advion | HD-D-384 | |
| Acetic acid | Sigma Aldrich | 45754-100ML-F | |
| Ammonium acetate | Honeywell | 14267-25G | |
| Ammonium bicarbonate | Sigma Aldrich | 09830-500G | |
| Bovine serum albumin standard, 2 mg/mL | Thermo Scientific | 23209 | |
| Butanol | Honeywell | 33065-2.5L | |
| Chloroform | Sigma Aldrich | 650498-1L | |
| Dichloromethane | Honeywell | 34856-1L | |
| Ethyl acetate | Honeywell | 33211 | |
| Greiner CELLSTAR 96 well plates | Sigma | M9686 | |
| Heptane | Sigma Aldrich | 34873-2.5L | |
| Isopropanol | Fisher Scientific | A461 | |
| Methanol | Fisher Scientific | A456 | |
| Mouse pooled plasma | BioIvt | ||
| Mouse SPLASH standard | Avanti Polar Lipids | 330710X | Internal standard |
| Nunc 96-flat bottom well transparent plates | VWR | 62409-068 | |
| Plastic spatula | Sigma | Z560049-300EA | |
| Quick Start Bradford 1x Dye reagent | BioRad | 5000205 | |
| Serum diluent | Sigma Aldrich | D5197 | |
| Equipment/software | |||
| CryoPrep CP02 impactor instrument | Covaris | Magnetic hammer | |
| Centrifuge 5427R | Eppendorf | . | Centrifuge |
| ChipSoft 8.3 | Advion Biosciences | . | Software to set up method and acquisition on the Triversa nanomate robot |
| LipidXplorer 1.2.8.1 | N/A | . | Software to identify lipids |
| Peak By Peak | SpectroSwiss | . | Software to convert .raw data from MS to .mzml format |
| ProteoWizard | ProteoWizard | . | Alternative (open source) software to convert .raw data from MS to .mzml format |
| Q-Exactive MS | Thermo Fisher | . | High resolution orbitrap mass spectrometer |
| Qiagen Tissue Lyser II | Qiagen | . | Tissue lyser |
| SpeedVac SPD140DDA | Thermo Fisher | . | Vacuum concentrator |
| Tecan Infinite M nano plus | Tecan | . | Plate reader |
| ThermoMixer C | Eppendorf | . | Thermomixer |
| TriVersa Nanomate | Advion Biosciences | . | Direct infusion nano-source |
| Xcalibur 4.3 | Thermo Scientific | . | Software to set up method and acquisition on the Q-Exactive MS |
Ссылки
- Salehi, N., Janjani, P., Tadbiri, H., Rozbahani, M., Jalilian, M. Effect of cigarette smoking on coronary arteries and pattern and severity of coronary artery disease: A review. Journal of International Medical Research. 49 (12), 3000605211059893 (2021).
- Churg, A., Sin, D. D., Wright, J. L. Everything prevents emphysema: Are animal models of cigarette smoke-induced chronic obstructive pulmonary disease any use. American Journal of Respiratory Cell and Molecular Biology. 45 (6), 1111-1115 (2011).
- Lee, G., Walser, T. C., Dubinett, S. M. Chronic inflammation, chronic obstructive pulmonary disease, and lung cancer. Current Opinion in Pulmonary Medicine. 15 (4), 303-307 (2009).
- Titz, B., et al. Effects of cigarette smoke, cessation, and switching to two heat-not-burn tobacco products on lung lipid metabolism in C57BL/6 and Apoe-/- mice-An integrative systems toxicology analysis. Toxicological Sciences. 149 (2), 441-457 (2016).
- Morissette, M. C., Shen, P., Thayaparan, D., Stampfli, M. R. Disruption of pulmonary lipid homeostasis drives cigarette smoke-induced lung inflammation in mice. European Respiratory Journal. 46 (5), 1451-1460 (2015).
- Jubinville, E., et al. Interplay between cigarette smoking and pulmonary reverse lipid transport. European Respiratory Journal. 50 (3), 1700681 (2017).
- Han, X., Gross, R. W. Shotgun lipidomics: Electrospray ionization mass spectrometric analysis and quantitation of cellular lipidomes directly from crude extracts of biological samples. Mass Spectrometry Reviews. 24 (3), 367-412 (2005).
- Titz, B., et al. Multi-omics systems toxicology study of mouse lung assessing the effects of aerosols from two heat-not-burn tobacco products and cigarette smoke. Computational and Structural Biotechnology Journal. 18, 1056-1073 (2020).
- Hartung, T., et al. Systems toxicology: Real world applications and opportunities. Chemical Research in Toxicology. 30 (4), 870-882 (2017).
- Talikka, M., et al., Lyubimov, A., et al. Systems Toxicology. Encyclopedia of Drug Metabolism and Interactions. , (2016).
- Ejsing, C. S., et al. Global analysis of the yeast lipidome by quantitative shotgun mass spectrometry. Proceedings of the National Academy of Sciences of the United States of America. 106 (7), 2136-2141 (2009).
- Papan, C., et al. Systematic screening for novel lipids by shotgun lipidomics. Analytical Chemistry. 86 (5), 2703-2710 (2014).
- Carvalho, M., et al. Effects of diet and development on the Drosophila lipidome. Molecular Systems Biology. 8, 600 (2012).
- Strittmatter, N., et al. Shotgun lipidomic profiling of the NCI60 cell line panel using rapid evaporative ionization mass spectrometry. Analytical Chemistry. 88 (15), 7507-7514 (2016).
- Lydic, T. A., et al. Rapid and comprehensive 'shotgun' lipidome profiling of colorectal cancer cell derived exosomes. Methods. 87, 83-95 (2015).
- Sampaio, J. L., et al. Membrane lipidome of an epithelial cell line. Proceedings of the National Academy of Sciences of the United States of America. 108 (5), 1903-1907 (2011).
- Grzybek, M., et al. Comprehensive and quantitative analysis of white and brown adipose tissue by shotgun lipidomics. Molecular Metabolism. 22, 12-20 (2019).
- Stegemann, C., et al. Comparative lipidomics profiling of human atherosclerotic plaques. Circulation: Cardiovascular Genetics. 4 (3), 232-242 (2011).
- Tajima, Y., et al. Lipidomic analysis of brain tissues and plasma in a mouse model expressing mutated human amyloid precursor protein/tau for Alzheimer's disease. Lipids in Health and Disease. 12, 68 (2013).
- Gross, R. W., Han, X. Shotgun lipidomics of neutral lipids as an enabling technology for elucidation of lipid-related diseases. American Journal of Physiology: Endocrinology and Metabolism. 297 (2), 297-303 (2009).
- Surma, M. A., et al. An automated shotgun lipidomics platform for high throughput, comprehensive, and quantitative analysis of blood plasma intact lipids. European Journal of Lipid Science and Technology. 117 (10), 1540-1549 (2015).
- Heiskanen, L. A., Suoniemi, M., Ta, H. X., Tarasov, K., Ekroos, K. Long-term performance and stability of molecular shotgun lipidomic analysis of human plasma samples. Analytical Chemistry. 85 (18), 8757-8763 (2013).
- Zhang, S., Van Pelt, C. K. Chip-based nanoelectrospray mass spectrometry for protein characterization. Expert Review of Proteomics. 1 (4), 449-468 (2004).
- Surma, M. A., et al. Mouse lipidomics reveals inherent flexibility of a mammalian lipidome. Scientific Reports. 11 (1), 19364 (2021).
- Herzog, R., Schwudke, D., Shevchenko, A. LipidXplorer: Software for quantitative shotgun lipidomics compatible with multiple mass spectrometry platforms. Current Protocols in Bioinformatics. 43, 11-30 (2013).
- R: A language and environment for statistical computing. R: Foundation for Statistical Computing Available from: https://www.R-project.org/ (2018)
- Phillips, B., et al. A 7-month cigarette smoke inhalation study in C57BL/6 mice demonstrates reduced lung inflammation and emphysema following smoking cessation or aerosol exposure from a prototypic modified risk tobacco product. Food and Chemical Toxicology. 80, 328-345 (2015).
- Phillips, B., et al. A six-month systems toxicology inhalation/cessation study in ApoE(-/-) mice to investigate cardiovascular and respiratory exposure effects of modified risk tobacco products, CHTP 1.2 and THS 2.2, compared with conventional cigarettes. Food and Chemical Toxicology. 126, 113-141 (2019).
- Zemski Berry, K. A., Murphy, R. C. Electrospray ionization tandem mass spectrometry of glycerophosphoethanolamine plasmalogen phospholipids. Journal of the American Society for Mass Spectrometry. 15 (10), 1499-1508 (2004).
- Engelmann, B. Plasmalogens: Targets for oxidants and major lipophilic antioxidants. Biochemical Society Transactions. 32, 147-150 (2004).
- Nagan, N., Zoeller, R. A. Plasmalogens: Biosynthesis and functions. Progress in Lipid Research. 40 (3), 199-229 (2001).
- Southam, A. D., Weber, R. J., Engel, J., Jones, M. R., Viant, M. R. A complete workflow for high-resolution spectral-stitching nanoelectrospray direct-infusion mass-spectrometry-based metabolomics and lipidomics. Nature Protocols. 12 (2), 310-328 (2016).
- Yang, K., Han, X. Accurate quantification of lipid species by electrospray ionization mass spectrometry - Meet a key challenge in lipidomics. Metabolites. 1 (1), 21-40 (2011).
- Zullig, T., Trotzmuller, M., Kofeler, H. C. Lipidomics from sample preparation to data analysis: a primer. Analytical and Bioanalytical Chemistry. 412 (10), 2191-2209 (2020).
- Koivusalo, M., Haimi, P., Heikinheimo, L., Kostiainen, R., Somerharju, P. Quantitative determination of phospholipid compositions by ESI-MS: Effects of acyl chain length, unsaturation, and lipid concentration on instrument response. Journal of Lipid Research. 42 (4), 663-672 (2001).
- Bligh, E. G., Dyer, W. J. A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology. 37 (8), 911-917 (1959).
- Matyash, V., Liebisch, G., Kurzchalia, T. V., Shevchenko, A., Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. Journal of Lipid Research. 49 (5), 1137-1146 (2008).
- Lofgren, L., Forsberg, G. B., Stahlman, M. The BUME method: A new rapid and simple chloroform-free method for total lipid extraction of animal tissue. Scientific Reports. 6, 27688 (2016).
- Lofgren, L., et al. The BUME method: A novel automated chloroform-free 96-well total lipid extraction method for blood plasma. Journal of Lipid Research. 53 (8), 1690-1700 (2012).
- Jung, H. R., et al. High throughput quantitative molecular lipidomics. Biochimica et Biophysica Acta. 1811 (11), 925-934 (2011).
- Lavrynenko, O., et al. Ceramide ratios are affected by cigarette smoke but not heat-not-burn or e-vapor aerosols across four independent mouse studies. Life Sciences. 263, 118753 (2020).
- Schuhmann, K., et al. Shotgun lipidomics on a LTQ Orbitrap mass spectrometer by successive switching between acquisition polarity modes. Journal of Mass Spectrometry. 47 (1), 96-104 (2012).
- Schuhmann, K., et al. Quantitative fragmentation model for bottom-up shotgun lipidomics. Analytical Chemistry. 91 (18), 12085-12093 (2019).
- Lipidomics Standards Initiative Consortium. Lipidomics needs more standardization. Nature Metabolism. 1 (8), 745-747 (2019).
- Husen, P., et al. Analysis of lipid experiments (ALEX): A software framework for analysis of high-resolution shotgun lipidomics data. PloS One. 8 (11), 79736 (2013).
- Ni, Z., Angelidou, G., Lange, M., Hoffmann, R., Fedorova, M. LipidHunter identifies phospholipids by high-throughput processing of LC-MS and shotgun lipidomics datasets. Analytical Chemistry. 89 (17), 8800-8807 (2017).
- Zemski Berry, K. A., Murphy, R. C., Kosmider, B., Mason, R. J. Lipidomic characterization and localization of phospholipids in the human lung. Journal of Lipid Research. 58 (5), 926-933 (2017).
- Ghosh, M., Tucker, D. E., Burchett, S. A., Leslie, C. C. Properties of the Group IV phospholipase A2 family. Progress in Lipid Research. 45 (6), 487-510 (2006).
- Besnard, V., et al. Deletion of Scap in alveolar type II cells influences lung lipid homeostasis and identifies a compensatory role for pulmonary lipofibroblasts. Journal of Biological Chemistry. 284 (6), 4018-4030 (2009).
- Plantier, L., et al. Activation of sterol-response element-binding proteins (SREBP) in alveolar type II cells enhances lipogenesis causing pulmonary lipotoxicity. Journal of Biological Chemistry. 287 (13), 10099-10114 (2012).
- Kyle, J. E., et al. Cell type-resolved human lung lipidome reveals cellular cooperation in lung function. Scientific Reports. 8 (1), 13455 (2018).
- Lugg, S. T., Scott, A., Parekh, D., Naidu, B., Thickett, D. R. Cigarette smoke exposure and alveolar macrophages: mechanisms for lung disease. Thorax. 77 (1), 94-101 (2022).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены