Method Article
Lipidomica shotgun di tessuti di roditori
In questo articolo
Riepilogo
La lipidomica basata sulla spettrometria di massa shotgun fornisce un'istantanea quantitativa sensibile di un ampio spettro di classi lipidiche simultaneamente in una singola misurazione da vari tessuti di roditori.
Abstract
I lipidi svolgono un ruolo vitale come componenti essenziali di tutte le cellule procariotiche ed eucariotiche. I costanti miglioramenti tecnologici nella spettrometria di massa hanno reso la lipidomica un potente strumento analitico per il monitoraggio delle composizioni del lipidoma tissutale negli stati omeostatici e patologici. Questo articolo presenta un protocollo passo-passo per un metodo di analisi dei lipidi shotgun che supporta il rilevamento e la quantificazione simultanea di alcune centinaia di specie lipidiche molecolari in diversi campioni di tessuto e biofluido ad alta produttività. Questo metodo sfrutta l'iniezione diretta automatizzata di nano-flusso di un estratto lipidico totale arricchito con standard interni etichettati senza separazione cromatografica in uno strumento di spettrometria di massa ad alta risoluzione. A partire da quantità inferiori al microgrammo di tessuto dei roditori, l'analisi della SM richiede 10 minuti per campione e copre fino a 400 lipidi da 14 classi lipidiche nel tessuto polmonare del topo. Il metodo qui presentato è adatto per studiare i meccanismi della malattia e identificare e quantificare i biomarcatori che indicano tossicità precoce o effetti benefici all'interno dei tessuti dei roditori.
Introduzione
Il fumo di sigaretta (CS) è riconosciuto come un principale fattore di rischio associato allo sviluppo di malattie infiammatorie croniche del polmone, tra cui carcinoma polmonare, bronchite e broncopneumopatia cronica ostruttiva (BPCO)1. Oltre all'impatto polmonare, l'esposizione alla CS svolge un ruolo significativo nello sviluppo di altre malattie come la malattia coronarica aterosclerotica e la malattia vascolare periferica1. Le malattie cardiovascolari, insieme alla BPCO, sono rispettivamente la prima e la terza causa di morte in tutto il mondo. Gli approcci di valutazione del rischio tossicologico si sono storicamente basati sull'uso di modelli animali come i roditori. I modelli di ratto e topo in vivo solo naso o full-body sono comunemente usati per studiare gli effetti a lungo termine dell'esposizione alla CS.
Ad esempio, generalmente, 6 mesi di esposizione al fumo inducono anomalie istologiche e funzionali nei polmoni murini che imitano quelli delle malattie umane, tra cui enfisema, rimodellamento delle vie aeree e ipertensione polmonare, sebbene i cambiamenti siano relativamente lievi rispetto a quelli osservati nei fumatori umani a lungo termine2. Sia nei tessuti animali che in quelli umani, gli studi hanno osservato una vasta gamma di cambiamenti molecolari in risposta all'esposizione a CS, tra cui risposte allo stress ossidativo, infiammazione e cambiamenti strutturali del tessuto 3,4. Non sorprende che l'esposizione a CS abbia anche un impatto di vasta portata sul lipidoma polmonare, compresi gli effetti sui lipidi tensioattivi, sui mediatori della segnalazione lipidica e sui lipidi strutturali 4,5,6.
Per caratterizzare i cambiamenti lipidici di massa indotti dall'esposizione a lungo termine al CS dei polmoni di topo, abbiamo eseguito un'analisi di spettrometria di massa a infusione diretta rapida e quantitativa. Dopo l'introduzione del metodo di lipidomica shotgun nel 20057, questo metodo è stato efficacemente utilizzato per estendere le nostre conoscenze sul metabolismo cellulare lipidico 8,9,10 in sistemi modello come lievito 11, Caenorhabditis elegans12 e Drosophila melanogaster13, nonché in una vasta gamma di tipi di campioni di mammiferi come le linee cellulari 14, 15,16 vari tessuti umani17,18 e roditori19,20 e fluidi corporei21,22.
Negli ultimi decenni, gli studi hanno rivelato la complessità delle risposte cellulari ai cambiamenti ambientali, coinvolgendo migliaia di proteine, lipidi e metaboliti interconnessi. Ciò ha chiarito che l'utilizzo di tecniche analitiche all'avanguardia è essenziale per ottenere una visione approfondita dei macchinari molecolari e per scoprire l'intera entità degli impatti fisiologici esogeni. In questo contesto, le impronte lipidiche complete e quantitative prodotte dagli approcci lipidomici shotgun possono effettivamente aggiungere alle nostre conoscenze sul metabolismo cellulare lipidico 8,9,10.
Per quanto riguarda l'esposizione al CS come fattore di rischio per diverse malattie, gli approcci di valutazione del rischio tossicologico si sono storicamente basati sull'uso di modelli animali come i roditori. Shotgun lipidomics MS fornisce uno strumento analitico rapido, sensibile e quantitativo per valutare la perturbazione del lipidoma in numerosi tipi di campioni. La caratteristica unica della lipidomica shotgun è l'analisi automatizzata a iniezione diretta di un estratto lipidico totale - spiked con standard interni etichettati - senza separazione cromatografica in uno strumento MS ad alta risoluzione utilizzando un nanochip conduttivo che genera un nanospray di ionizzazione elettrospray (ESI)23.
Le informazioni sul rapporto massa-carica acquisite simultaneamente nella modalità MS1 forniscono l'impronta lipidica totale di tutti i lipidi endogeni intatti. Opzionalmente, la modalità MS2/MS3, in cui le molecole lipidiche madri vengono frammentate e analizzate, fornisce ulteriori informazioni strutturali. L'analisi dei dati richiede un software specializzato e include la deconvoluzione degli spettri e le assegnazioni dei picchi negli spettri raggruppati che portano all'identificazione dei lipidi e alla delucidazione della struttura chimica putativa. Inoltre, la quantificazione assoluta può essere eseguita mediante spiking di una miscela di standard interni etichettati contenente almeno uno standard lipidico per classe lipidica di interesse. Nel complesso, con la presente tecnica, l'analisi della SM che richiede pochi minuti per campione può coprire l'identificazione e la quantificazione di un massimo di 800 lipidi da 14 classi lipidiche24 nei tessuti dei roditori.
Protocollo
Tutte le procedure che coinvolgono animali sono state eseguite in una struttura accreditata dall'Associazione per la valutazione e l'accreditamento della cura degli animali da laboratorio internazionale e autorizzata dall'Autorità agroalimentare e veterinaria di Singapore, con l'approvazione di un comitato istituzionale per la cura e l'uso degli animali e in conformità con le linee guida del comitato consultivo nazionale per la ricerca sugli animali da laboratorio sulla cura e l'uso degli animali per scopi scientifici (NACLAR, 2004).
1. Raccolta dei campioni
- Eseguire dissezioni polmonari di topo dopo 3 mesi e 6 mesi di esposizione di topi ApoE−/− . Raccogliere i tessuti 16-24 ore dopo l'esposizione, congelarli in azoto liquido e conservarli a -80 °C prima di iniziare il flusso di lavoro lipidomico. Garantire la raccolta di un totale di nove campioni da ciascun gruppo di dissezione "omica".
2. Tessuto - polverizzazione dei campioni
- Preparare il martello magnetico e i materiali di consumo per la macinazione dei tessuti. Sacchetti di tessuto preraffreddato, pinze, tubi di trasferimento del tessuto con cappuccio, una spatola di plastica a "V" e 2 mL di tubi di raccolta della plastica su ghiaccio secco. Installare il supporto del tubo corrispondente su uno strumento martello magnetico. Accendere il martello magnetico e impostare la potenza d'impatto su Alta.
- Caricare il campione in una sacca di tessuto; Inserire il campione attraverso l'apertura superiore della sacca di tessuto utilizzando una pinza preraffreddata, se necessario. Posizionare il campione al centro della sacca flessibile, lontano dai bordi.
- Sigillare la sacca di tessuto avvitando un tubo di raccolta preraffreddato sulla parte superiore della sacca di tessuto.
- Congelare il campione immergendo la busta flessibile in azoto liquido per 10 s.
- Sfiatare il tubo di raccolta; Allentare il tubo per un giro di 1/4 per lo sfiato per evitare di rompere la sacca quando il martello magnetico impatta.
- Caricare la sacca di tessuto congelata sul martello magnetico.
- Applicare il martello magnetico premendo il pulsante di attivazione per polverizzare il campione. Se nella sacca rimangono pezzi di tessuto non polverizzati, congelare nuovamente il campione in azoto liquido e ripetere per un secondo impatto.
- Rimuovere la sacca di tessuto dal martello magnetico, mantenendo il campione polverizzato sul fondo. Per evitare che il campione si sciolga, congelarlo di nuovo immediatamente.
- Trasferire il campione in una provetta di raccolta. Capovolgere rapidamente il tubo in modo che la sacca di tessuto sia nella parte superiore e toccare la sacca per trasferire le particelle di tessuto nella parte inferiore del tubo di raccolta.
NOTA: Questo passaggio dovrebbe essere fatto rapidamente per evitare lo scioglimento del tessuto. - Trasferire un'aliquota tissutale da 10 mg in una provetta da 2 ml per un'ulteriore analisi lipidomica e conservare il campione a -80 °C fino a ulteriori fasi.
3. Omogeneizzazione dei tessuti
- Accendere lo strumento tissue lyser e impostare la frequenza di agitazione a 30 Hz e la durata dell'agitazione a 2 min. Preparare la soluzione tampone di omogeneizzazione tissutale: 150 mM di bicarbonato di ammonio in acqua, con un pH di ~8,2.
- Estrarre i campioni dal congelatore, metterli sul ghiaccio e aggiungere quattro perle di acciaio inossidabile nei tubi contenenti la polvere di tessuto. Aggiungere 0,5 mL di tampone bicarbonato di ammonio da 150 mM a ciascun campione.
- Posizionare le provette nei supporti del lyser tissutale. Controbilanciare entrambi i supporti con lo stesso numero di tubi. Fissare i supporti nel braccio di tenuta del lyser tissutale. Interrompere il tessuto.
- Posizionare i tubi sul ghiaccio e controllare visivamente che non siano presenti aggregati tissutali residui nel tubo. Se la rottura dei tessuti è incompleta (si vedono aggregati), ripetere il processo di interruzione eseguendo un altro ciclo di trattamento. Posizionare i campioni sul ghiaccio.
- Creare un'aliquota 1 di 100 μL del campione in una provetta da 1,5 mL dedicata alla determinazione delle proteine. Centrifugare a 18.200 × g per 5 minuti a 10 °C.
- Determinare il contenuto proteico dell'aliquota 1 mediante il test di Bradford (vedere paragrafo 4). Conservare i campioni su ghiaccio.
- Creare un'aliquota 2 di 20-100 μL dell'omogenato madre in un nuovo microtubo da 2 mL per l'estrazione dei lipidi (vedere paragrafo 5). Se i campioni non vengono analizzati immediatamente, tenerli in ghiaccio fino a quando non viene eseguita l'estrazione.
NOTA: Il volume finale di estrazione deve essere definito sulla base della quantità totale di proteine. Deve essere compreso tra 0,015 e 0,045 mg di proteine per aliquota totale.
4. Saggio di determinazione della proteina di Bradford
NOTA: i campioni di studio devono essere randomizzati prima dell'inizio dell'analisi e la randomizzazione è rispettata per tutte le fasi dell'elaborazione del campione.
- Aliquota centrifugata diluita 1 surnatante 2x con tampone bicarbonato di ammonio.
NOTA: Il fattore di diluizione dipende dalla concentrazione dell'omogenato tissutale. Per soluzioni più concentrate, applicare un fattore più elevato in modo che la concentrazione determinata rientri nell'intervallo dinamico del test. - Preparare una curva standard dell'albumina sierica bovina (BSA) secondo la Tabella 1. Vortice i tubi standard. Centrifugare i tubi a 18.200 × g per 15 s a temperatura ambiente.
- Dai tubi standard precedentemente preparati, trasferire 6 μL ciascuno del bianco e degli standard su una piastra a fondo piatto a 96 pozzetti secondo il layout della piastra mostrato nella figura 1.
- Trasferire i campioni precedentemente preparati sulla piastra a fondo piatto a 96 pozzetti secondo il layout della piastra descritto nella figura 1.
- Aggiungere 250 μL di Bradford Reagent a ciascun pozzetto utilizzando una pipetta multicanale e miscelare. Incubare per 5 min.
- Misurare l'assorbanza a una lunghezza d'onda di 595 nm utilizzando un lettore di piastre.
- Calcolare le concentrazioni proteiche nelle aliquote in base alla curva standard.
NOTA: Il fattore di diluizione utilizzato all'inizio della procedura deve essere considerato nel calcolo delle concentrazioni proteiche.
5. Estrazione del BUME
NOTA: Evitare l'uso di tubi di plastica a basso legame per l'estrazione lipidomica, poiché i solventi organici forti rilasciano plastificanti e creano uno sfondo forte durante l'analisi del campione.
- Preparare provette da 1,5 mL con 100 μL di soluzione di bicarbonato di ammonio in ciascuna provetta del bianco totale (TB), del bianco standard interno (ISB) e dei campioni di controllo qualità (QC), considerando che 1 campione di controllo qualità viene posizionato tra ogni set di 10 campioni. Conservare tutti i campioni sul ghiaccio.
- Aggiungere 10 μL di plasma umano aggregato a tutti i campioni di controllo qualità. Aggiungere 10 μL di soluzione standard interna in tutti i campioni ISB, QC e di studio (cioè tutti i campioni tranne TB).
NOTA: Per il materiale di controllo qualità, utilizzare qualsiasi plasma K2EDTA disponibile in commercio. Quantificare la concentrazione lipidica nel plasma aggregato utilizzando il protocollo menzionato al momento della ricezione e mantenere questi intervalli come riferimenti per tutte le analisi in cui viene utilizzato il materiale QC. Utilizzare il plasma NIST solo per applicazioni più mirate o per la verifica globale del metodo, in cui è necessario affrontare l'accuratezza del test. - Aggiungere 300 μL di miscela butanolo/metanolo (3/1, v/v) a ciascun campione. Agitare a 450 × g per 10 minuti a temperatura ambiente sul termomixer.
- Aggiungere 300 μL di miscela eptano/acetato di etile (3/1, v/v). Agitare a 450 × g per 10 minuti a temperatura ambiente sul termomixer.
- Aggiungere 300 μL di miscela di acido acetico all'1%. Agitare per 5 minuti a 450 × g a temperatura ambiente sul termomixer. Centrifugare a 2.800 × g per 5 minuti a temperatura ambiente. Trasferire 360 μL della fase superiore in un nuovo tubo autobloccante da 2 mL 2.
NOTA: l'estrazione liquido-liquido può comportare lo spostamento delle posizioni del limite di fase in modo dipendente dal campione, che può comportare una leggera discrepanza di volume per la fase superiore. Se necessario, regolare il volume di raccolta per la fase superiore. - Aggiungere 320 μL di eptano/acetato di etile (3/1, v/v) al tubo 1 in fase acquosa. Agitare a 450 × g per 5 minuti a temperatura ambiente sul termomixer. Centrifugare a 2.800 × g per 5 minuti a temperatura ambiente. Trasferire 320 μL della fase superiore e combinare con la frazione dal punto 5.11.
NOTA: se necessario, regolare il volume di raccolta per la fase superiore. - Aggiungere 250 μL di eptano/acetato di etile (3/1, v/v) alla fase acquosa nel tubo 1. Agitare a 450 × g per 5 minuti a temperatura ambiente sul termomiscelatore. Centrifugare a 2.800 × g per 5 minuti a temperatura ambiente. Trasferire 200 μL della fase superiore e combinare con le frazioni del passo 5.11 e del passo 5.15 nel tubo 2.
NOTA: se necessario, regolare il volume di raccolta per la fase superiore. - Evaporare fino all'essiccazione a 35 °C in un concentratore sotto vuoto.
NOTA: I campioni essiccati possono essere conservati a -20 °C fino all'analisi, se necessario. - Ridissolvere in 300 μL di soluzione di miscela MS (7,5 mM di acetato di ammonio in isopropanolo/metanolo/cloroformio, soluzione 4:2:1). Vortice ogni tubo per 5 s per garantire che tutto sia sciolto. Centrifugare a 18.200 × g per 5 min a 4 °C.
- Trasferire un'aliquota di 50 μL nella piastra da microlitri secondo la disposizione della piastra nella figura 2 per l'analisi della modalità di ionizzazione positiva (colonne 1-6) e diluire con 50 μL di soluzione di miscela MS.
- Trasferire un'aliquota di 20 μL nella piastra MTP secondo la disposizione della piastra in Figura 2 per l'analisi della modalità di ionizzazione negativa (colonne 7-12) e diluire con 80 μL di miscela MS. Avvolgere la piastra con un foglio di alluminio e conservare a 4 °C prima dell'analisi.
6. Analisi della SM
- Calibrare uno spettrometro di massa (MS) sia in modalità positiva che negativa in conformità con le istruzioni del produttore 5 giorni o meno prima dell'analisi.
- Installare la nano-sorgente di infusione diretta in anticipo, assicurandosi che sia correttamente allineata contro il capillare di trasferimento della MS. Installare un chip ugello da 4,1 μm nel supporto del chip e configurarlo nel software.
- Utilizzare i seguenti parametri per l'impostazione del metodo in modalità positiva e negativa: pressione del gas di 1,25 psi; tensione di sorgente di 1,1 kV; 5 μL di volume di iniezione del campione; chiusura contatto uscita Rel1 per 2,5 s; 5 min di tempo di consegna; raffreddamento a piastre a 4 °C.
- Impostare la coda della sequenza di analisi per il robot di infusione diretta in modalità positiva.
- Impostare il metodo MS per la modalità positiva utilizzando le seguenti impostazioni MS:
- Impostare la temperatura capillare a 250 °C e il livello RF dell'obiettivo S a 65,0. Esegui l'acquisizione MS a scansione completa da 0 a 1 minuto nell'intervallo 550-1000 m/z con risoluzione 140.000 con controllo automatico del guadagno di 1 ×106 e tempo di iniezione massimo di 50 ms. Applicare la massa di blocco di 680,48022.
- Impostare il metodo di acquisizione MS/MS indipendente dai dati tra l'intervallo di tempo di 1 e 5 minuti con una risoluzione di 17.500 con una prima massa fissa di 250 m/z. Utilizzare un controllo automatico del guadagno per MS2 a 1 × 105 e un tempo massimo di iniezione di 64 ms, un'energia di collisione di 20 nce e una finestra di isolamento di 1 m/z. Utilizzare l'elenco delle masse di inclusione da 550 a 1.000 m/z, con un passo di massa di 1 Da.
- Per l'acquisizione in modalità negativa, impostare la temperatura capillare a 250 °C e il livello RF S-Lens a 65,0 nel file MS tune.
- Utilizzare le seguenti impostazioni del metodo MS: modalità di acquisizione full-scan da 0 a 1 min con risoluzione 140.000 che copre l'intervallo 400-940 m/z, controllo automatico del guadagno di 1 × 106; tempo massimo di iniezione di 50 ms; Utilizzare una massa di blocco di 529,46262.
- Impostare l'acquisizione MS2 in modalità DIA tra 1 e 5 minuti della corsa con una risoluzione di 17.500 con una prima massa fissa di 150 m/z; controllo automatico del guadagno di 1 × 105; 64 ms di tempo massimo di iniezione; e 35NCE di energia di collisione. Utilizzare l'elenco di massa di inclusione da 400 a 940 con un passo di massa di 1 Da.
- Creare la coda della sequenza di analisi nel software MS in modalità positiva . Attendere che l'acquisizione del campione venga attivata da un segnale di chiusura del contatto proveniente dal robot di infusione diretta per ciascun campione.
- Al termine dell'analisi in modalità positiva, creare la coda di sequenza per il robot di infusione diretta in modalità negativa .
- Impostare la coda della sequenza di analisi nel software MS in modalità negativa .
7. Trattamento dei dati
- Per convertire i file di dati dalle modalità positiva e negativa dal formato .raw al formato .mzML, utilizzare il software di conversione.
NOTA: i file di dati delle modalità positiva e negativa devono essere convertiti separatamente (a causa delle diverse impostazioni di acquisizione e della diversa soglia di rumore).- Compilare le seguenti impostazioni per eseguire la conversione dei file: il fattore di soglia del rumore è 3 e 5 rispettivamente per MS e MS2; attivare la funzione di rimozione del rumore sia per MS che per MS2, compressione dei dati, scansioni MS2 medie e peak picking. Impostare soglie TIC per le modalità positiva e negativa (ad esempio, 10.000 e 5.000; da definire in base al livello di rumore del particolare campione prodotto da uno strumento specifico). Generare report XLC e PDF al termine dell'elaborazione.
- Eseguire l'identificazione dei lipidi. Definire i seguenti parametri (di esempio) delle impostazioni di importazione dei file: finestra di selezione ±0,5 Da (dipende dalla finestra di selezione nel metodo MS); intervallo di tempo 2-300 s (dipende dal tempo di scansione nel metodo MS); nessuna massa di calibrazione per MS e MS2; trasferire e arrotondare l'intervallo di massa dai file di metadati del software Peak by Peak (min mass [MS]) e (min mass [MS2]); tolleranza di 3 ppm e 10 ppm per MS e MS2, rispettivamente; tenere vuoto il campo soglia MS e MS2, il filtro di frequenza, l'offset MS1 e il PMO; Correzione isotopica per MS e MS2 selezionati; trasferire il gradiente di risoluzione dal file di metadati del convertitore come (Resolution linear fit [MS]) e as (Resolution linear fit [MS2]).
NOTA: L'identificazione dei lipidi nelle modalità positiva e negativa deve essere eseguita separatamente a causa delle loro diverse impostazioni di acquisizione. - Dopo aver importato i dati, vai al menu Esegui e carica i file MFQL per l'identificazione dei lipidi.
NOTA: Per informazioni dettagliate sulla struttura degli MFQL, fare riferimento alla pubblicazione di Herzog et al.25. Vedere alcuni esempi di file MFQL in https://wiki.mpi-cbg.de/lipidx/MFQL_library e vedere la discussione. Tutti gli MFQL utilizzati per l'elaborazione dei dati sono forniti nei Materiali supplementari. - Quantificare le specie identificate a livello di SM dividendo l'intensità di massa del precursore della caratteristica lipidica per la rispettiva intensità dello standard interno marcato e quindi moltiplicando il quoziente per la concentrazione dello standard interno marcato. Normalizzare la concentrazione lipidica finale per quantità di proteina totale misurata dal test di Bradford.
8. Procedura di controllo del controllo qualità
NOTA: Una procedura di controllo della qualità è un passo essenziale per verificare la riproducibilità tecnica del metodo. A tal fine, viene analizzato un plasma aggregato commerciale per determinare i livelli endogeni di diversi lipidi nell'arco di 5 giorni in 15 aliquote identiche per lotto (totale di cinque lotti). Si noti che, in questo caso, è stata creata una pipeline di valutazione dei dati come applicazione Web Shiny utilizzando l'ambiente software statistico R.
- Definire l'intervallo di accettazione di riferimento come il valore medio calcolato per tutti e cinque i lotti ±3 deviazioni standard.
- Applicare le regole di accettazione "pass" per ogni lotto analitico: il 90% dei target di riferimento deve essere superato, considerando che un composto di riferimento rappresenta una classe lipidica. Ad esempio, se nella verifica del controllo qualità sono inclusi 15 target di riferimento, assicurarsi che 13 target passino affinché il lotto venga accettato.
NOTA: In altre parole, il 68% dei campioni di QC per composto di riferimento deve passare affinché i dati per questa classe lipidica siano accettati. - Se il lotto di studio non soddisfa i criteri di accettazione per il controllo del controllo QC, ripetere la procedura.
9. Stima delle quantità (relative) di acidi grassi coniugati
- Per ogni classe lipidica, stimare le quantità di acidi grassi coniugati in base alle loro intensità MS2.
NOTA: A causa delle differenze di frammentazione e ionizzazione, i valori derivati sono stati pensati solo per confronti di abbondanza relativa tra gruppi di campioni piuttosto che, ad esempio, per stimare il contributo relativo di uno specifico acido grasso al pool complessivo di acidi grassi coniugati di una classe lipidica. - Normalizzare le intensità MS2 dei frammenti di acidi grassi in base all'intensità media dei frammenti di acidi grassi dello standard interno corrispondente. Sulla base della concentrazione dello standard interno e del peso del tessuto misurato, derivare una "concentrazione normalizzata" per gli acidi grassi coniugati. Sommare questi valori per classe lipidica per stimare la quantità totale di ciascun acido grasso coniugato per campione.
10. Analisi statistica
- Eseguire analisi statistiche. Adatta un modello lineare per ogni condizione e il gruppo di controllo corrispondente e calcola i valori p da una statistica t per i dati trasformati in log2. Utilizzare il metodo del tasso di falsa scoperta di Benjamini-Hochberg per correggere gli effetti di test multipli.
NOTA: I lipidi con valori p aggiustati < 0,05 sono considerati differenzialmente abbondanti. In questo caso, l'analisi statistica è stata eseguita nell'ambiente statistico R26.
Risultati
Qui, come risultati rappresentativi, presentiamo i dati che illustrano come la lipidomica del fucile può essere utilizzata per studiare gli effetti della CS sui polmoni dei topi. Il protocollo di analisi lipidica shotgun è stato applicato con successo per uno studio in vivo per l'analisi comparativa di due gruppi di campioni: tessuti polmonari sezionati da nove topi femmina Apoe-/− esposti a CS (gruppo 3R4F; controllo positivo) e un numero uguale di topi mantenuti in condizioni di aria fresca (gruppo sham) come controllo negativo raccolto a 3 mesi, Punti temporali di esposizione di 4 mesi e 6 mesi. Gli animali sono stati tenuti sotto aria fresca filtrata e condizionata a 22 °C ± 2 °C di temperatura e 55% ± 15% di umidità e alimentati con una dieta a pellet irradiata con raggi gamma. Il regime di luce è stato mantenuto a 12 ore al giorno. Sono stati ospitati fino a otto topi per ogni gabbia. Le sigarette di riferimento 3R4F per il trattamento di esposizione sono state acquistate dall'Università del Kentucky per generare l'aerosol 3R4F CS (600 μg di aerosol TPM / L di particolato totale) per una concentrazione di esposizione target equivalente a 28 μg di nicotina / L.CS fumo dalle sigarette 3R4F è stato prodotto utilizzando macchine per fumatori rotative a 30 porte secondo Phillips et al.27 . Sulla base del protocollo sul fumo intensivo di Health Canada, 55 ml di volume di soffio, un soffio ogni 30 s e il blocco del 100% dei fori di ventilazione sono stati applicati alle sigarette 3R4F per fornire un'esposizione sufficiente. Tutti i dettagli aggiuntivi sul disegno dello studio sono stati riportati in precedenza27,28.
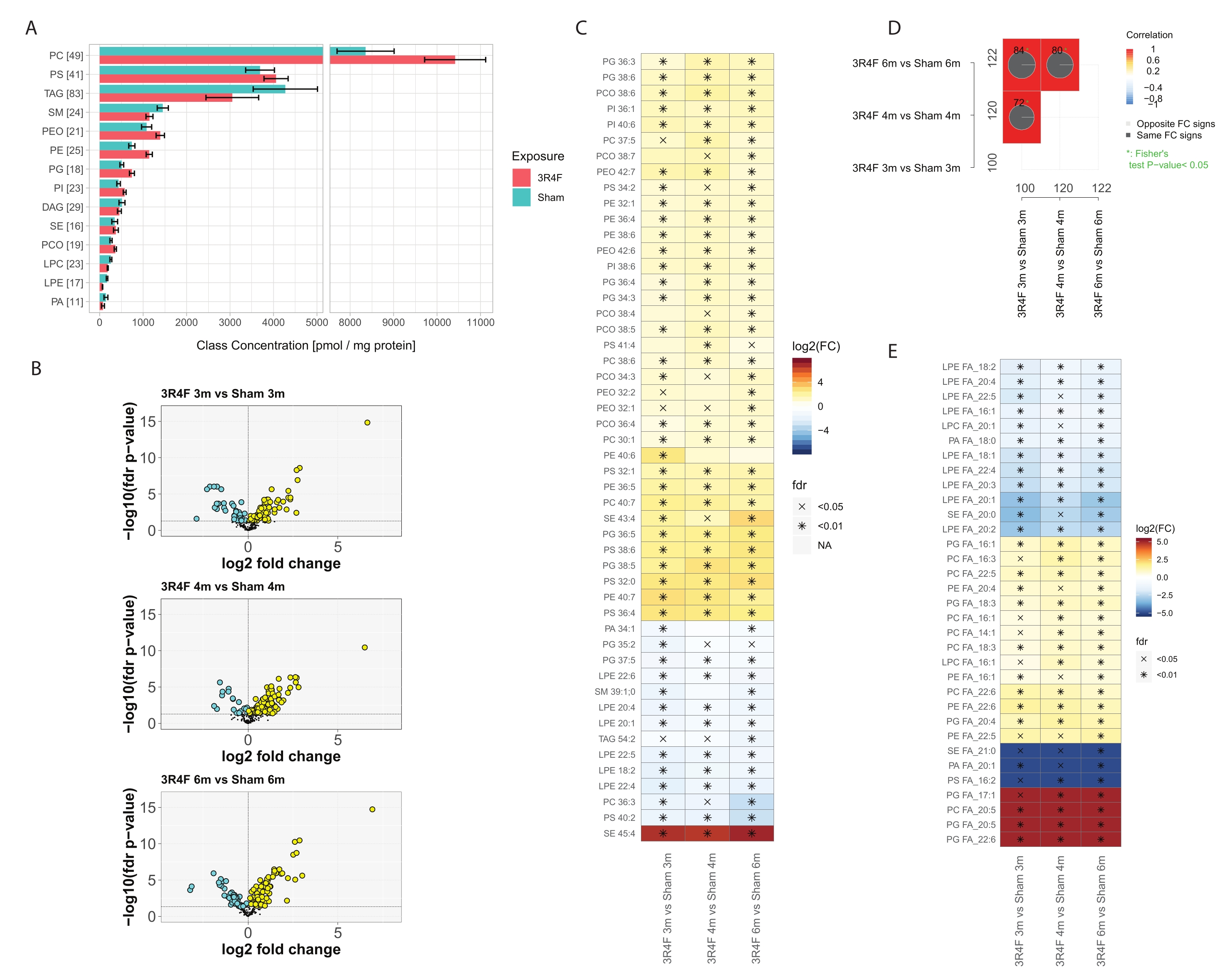
Nel complesso, ~ 400 specie di lipidi molecolari delle 14 classi lipidiche più abbondanti sono state identificate e quantificate (Figura 3). La concentrazione lipidica totale normalizzata per classe era leggermente elevata per le classi lipidiche PE, PEO, PC, PCO, PI e PG, e una certa downregulation è stata osservata per i lipidi SM, LPE e PA (Figura 3A), mentre nessuna differenza poteva essere vista per TAG e PS. È interessante notare che livelli elevati di lipidi plasmalogeni PCO e PEO nel gruppo CS sono stati riportati nelle cellule infiammatorie29 . Una delle funzioni intracellulari dei lipidi plasmalogeni è l'attività antiossidante. Sotto esposizione a specie reattive dell'ossigeno (ROS) e azoto (RNS), è stata riportata l'ossidazione selettiva dei plasmalogeni rispetto ai fosfolipidi diacilici30. Il legame enil-etere nella struttura chimica dei plasmalogeni è sensibile all'attacco radicale dello stress ossidativo31. I principali prodotti dell'attacco radicale sono gli acidi eicosatetraenoici idrossilati, i fosfolipidi 2-monoacilglicerolo, il pentadecanolo, gli acidi formici, α-idrossialdeidi di varie lunghezze di catena, l'1-formil-2-arachidonoil glicerofosfolipidi e i lisofosfolipidi.
Questi risultati mostrano che, tra le caratteristiche molecolari basate sulla formula molecolare totale, 100-120 composti sono stati significativamente influenzati dall'esposizione a CS (Figura 3D). La deconvoluzione dettagliata delle informazioni di frammentazione MS2 e la normalizzazione delle intensità del segnale del frammento hanno permesso la definizione approssimativa della quantità totale di ciascun acido grasso coniugato (FA) rappresentato in ciascuna classe lipidica (Figura 3E) e il confronto di questi dati tra gruppi esposti e gruppi di controllo. È stata osservata una chiara diminuzione sia dei FA completamente saturi che di quelli con solo poche insaturazioni, soprattutto nel contesto dei lisosolipidi. Al contrario, gli acidi grassi polinsaturi (PUFA) nella composizione delle classi di fosfolipidi PC, PE e PG erano elevati nel gruppo CS per tutti i punti temporali. I casi estremi di PC e PG coniugati con acido eicosapentanoico (EPA) e docosaesaenoico (DHA) sono stati rilevati esclusivamente nel gruppo di esposizione 3R4F CS (Figura 3E, colore rosso).

Figura 1: Layout delle piastre per la distribuzione del campione nel saggio di Bradford. I campioni bianchi e standard sono posti in triplice copia nelle colonne 1-3; I campioni sconosciuti sono inseriti in duplicato nelle colonne 4-11. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 2: Disposizione della piastra della micropiastra a 96 pozzetti per l'analisi della spettrometria di massa shotgun. La distribuzione dei campioni bianchi, standard, sconosciuti e QC per l'acquisizione in modalità di ionizzazione positiva è mappata sul lato sinistro della piastra (colonne 1-6); Tutti i campioni per la modalità di ionizzazione negativa sono posizionati sulla destra (colonne 7-12). Abbreviazioni: pos = positivo; QC = controllo qualità; neg = negativo; TB = vuoto totale. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}

Figura 3: Analisi lipidomica shotgun di polmoni di topo esposti al fumo di sigaretta. I topi Apoe-/− sono stati esposti al CS dalla sigaretta di riferimento 3R4F o all'aria fresca (sham). (A) Concentrazioni della classe lipidica e numero di specie lipidiche per classe. Per ciascuna classe, il numero di specie lipidiche quantificate è indicato tra parentesi. Intervalli di confidenza medi e del 95% per ciascuna classe lipidica, separatamente per ciascun tipo di esposizione (aggregati su tre punti temporali). (B) Grafici del vulcano che mostrano la dimensione dell'effetto (variazione log2 volte) e la significatività (-log10 FDR-adjusted p value) delle specie lipidiche quantificate per il confronto 3R4F CS rispetto a sham nei tre punti temporali. I lipidi significativamente elevati sono contrassegnati in giallo; i lipidi significativamente diminuiti sono marcati in ciano (valore p aggiustato per FDR < 0,05). (C) Abbondanza differenziale delle 25 specie lipidiche con le maggiori variazioni assolute medie di piega. Le variazioni di piega di Log2 per il confronto 3R4F CS rispetto a quello fittizio sono codificate a colori e la significatività statistica è indicata: *: valore p aggiustato per FDR < 0,01; X: valore p corretto per FDR < 0,05. (D) Grafici di confronto. I coefficienti di correlazione per i confronti di fold-change sono codificati a colori e viene indicato il numero di lipidi condivisi differenzialmente abbondanti (numeri totali ai margini). I grafici a torta mostrano la percentuale di lipidi condivisi differenzialmente abbondanti con la stessa direzione di cambiamento (segno FC). L'asterisco indica una significativa sovrapposizione dei lipidi differenzialmente abbondanti. (E) Abbondanza differenziale di FA coniugati per classe lipidica. Mappa di calore come nel pannello C per gli acidi grassi coniugati con differenze significative di abbondanza tra 3R4F e esposizione fittizia in tutti e tre i punti temporali (valore p aggiustato per FDR < 0,05). La quantità di ciascun acido grasso per classe lipidica è stata stimata in base alle intensità MS2 dei frammenti di acido grasso. Abbreviazioni: CS = fumo di sigaretta; FDR = tasso di false scoperte; FC = cambio di piega; FA = acidi grassi; PC = fosfatidilcolina; PS = fosfatidilserina; TAG = triacilglicerolo; SM = sfingomielina; PEO = plasmalogen fosfatidiletanolammina; PE = fosfatidiletanolammina; PG = fosfatidil glicerolo; PI = fosfatidilinositolo; DAG = diacilglicerolo; SE = esteri sterolo/colesterile; PCO = plasmalogen fosfatidilcolina; LPC = lyso fosfatidilcolina; LPE = lyso fosfatidiletanolammina; PA = acido fosfatidico. Fare clic qui per visualizzare una versione ingrandita di questa figura.
{kind=link}
| Concentrazione finale BSA (mg/ml) | Soluzione di BSA a 2 mg/ml (μL) | Tampone bicarbonato di ammonio (μL) |
| 1 | 50 | 50 |
| 0.75 | 37.5 | 62.5 |
| 0.5 | 25 | 75 |
| 0.25 | 12.5 | 87.5 |
| 0.125 | 12.5 | 187.5 |
| 0.0625 | 50 di soluzione 0,125 mg/mL | 50 |
| 0 | 0 | 100 |
Tabella 1: Schema di diluizione dello standard interno BSA per la curva di taratura per il saggio di Bradford. Abbreviazione: BSA = albumina sierica bovina.
Informazioni supplementari: File MFQL utilizzati per l'identificazione dei lipidi LipidXplorer. Il pacchetto .zip è costituito dai singoli file MFQL richiesti, ciascuno denominato seguendo questo modello: __MS2.mfql (ad esempio, PE_neg_MS2.mfql). I nomi dei file per l'identificazione standard interna etichettata includono il tag D7 (D9) nella (ad esempio, PED7_neg_MS2.mfql). Questi file MFQL vengono utilizzati per verificare la quantità di deuterio nello standard interno. Clicca qui per scaricare questo file.
Discussione
Sebbene i progressi nella SM abbiano prodotto una varietà di metodi per monitorare accuratamente molte specie lipidiche, il precedente profilo lipidico di vari tessuti di mammifero non ha mostrato risultati coerenti e, di conseguenza, le particolari funzioni tessuto-specifiche dei lipidi rimangono poco chiare. Rispetto all'analisi funzionale delle proteine, per la quale sono disponibili approcci più robusti per eliminare l'espressione di particolari composti, la maggior parte dei lipidi non può essere selettivamente disattivata o sovraespressa nei tessuti, rendendo difficile la valutazione funzionale dei lipidi. La profilazione avanzata delle concentrazioni di lipidi tissutali può fornire un approccio alternativo per identificare le associazioni tra lipidi circolanti e malattie umane.
Nel valutare metodi lipidomici completi in grado di coprire qualitativamente e quantitativamente il profilo lipidico endogeno di qualsiasi tessuto murino, abbiamo dato la preferenza al metodo lipidomica shotgun. In generale, sono possibili due tipi opposti di analisi del campione: o uno screening completamente non mirato dei lipidi utilizzando la separazione basata sulla cromatografia liquida (LC) con ulteriore rilevamento spettrometrico di massa per rivelare la complessità lipidica totale del campione, o approcci mirati, che consentono per lo più una quantificazione molto precisa di specifici lipidi di interesse. Al contrario, la caratteristica forte del flusso di lavoro della lipidomica shotgun presentato è la rapida copertura completa di centinaia di lipidi endogeni da classi lipidiche predefinite, che possono ancora essere eseguite in modo semi-quantitativo robusto.
Le efficienze di ionizzazione delle diverse classi lipidiche sono dipendenti dalla struttura e possono variare drasticamente a seconda delle diverse condizioni di ionizzazione sperimentale. A differenza dei metodi di separazione basati su LC, l'analisi shotgun riduce al minimo queste differenze grazie all'infusione diretta simultanea dell'intero estratto lipidico nelle stesse condizioni di ionizzazione nello strumento MS. L'uso di analoghi lipidici marcati isotopicamente o standard strutturalmente simili non endogeni consente la semi-quantificazione di tutte le classi lipidiche. La lipidomica shotgun fornisce una bassa variabilità inter- e intra-run durante l'analisi della SM; Di conseguenza, questo metodo produce coefficienti di variazione21 inferiori rispetto ai metodi basati sulla cromatografia liquida non mirata, che richiedono più standard per una quantificazione adeguata. È importante sottolineare che, sebbene nel metodo corrente non venga utilizzata alcuna curva di calibrazione esterna, il metodo è ancora considerato pienamente quantitativo32.
Un livello di standard interno marcato (o standard non etichettato che non è espresso in modo endogeno) per classe lipidica è sufficiente per la quantificazione della maggior parte dei lipidi. Solo poche pubblicazioni hanno riportato una convalida parziale del metodo per un metodo di lipidomica shotgun. Ad esempio, in Gryzbek et al.17 e Surma et al.21, sono state preparate curve di calibrazione invertite utilizzando standard interni e una quantità fissa di matrice di campionamento. La linearità è stata valutata dalla regressione lineare delle quantità lipidiche log-trasformate e delle loro intensità e riportata rispettivamente come R2 e pendenza. Il limite di rilevamento (LOD) e il limite di quantificazione (LOQ) sono stati determinati mediante regressione lineare ponderata basata su un rapporto segnale-rumore di 3 per LOD e 10 per LOQ. Per la maggior parte delle classi lipidiche, il LOQ è stato definito tra 2-9,8 pmol per il tessuto adiposo e 0,05-5μM nel plasma. In entrambi i casi, sono stati utilizzati singoli standard interni non endogeni per classe per ricavare stime per tutti i lipidi della classe. Tuttavia, in questo lavoro, non forniamo LOD / LOQ a causa di diverse preoccupazioni: la matrice endogena non è priva di composti e la matrice surrogata per i tessuti non è disponibile - con questo, il picco di piccole quantità note di standard non è possibile. Non eseguiamo un classico approccio di quantificazione mirata con l'uso di una serie di curve di calibrazione di un particolare composto normalizzato dal suo identico standard isotopicamente marcato a causa dell'inesistenza di standard puri e della disponibilità molto limitata di lipidi marcati isotopicamente. I rivelatori Orbitrap eseguono automaticamente la conversione del segnale transitorio applicando una trasformazione di Fourier, e alcuni segnali sono già sostituiti - di conseguenza, l'intervallo di concentrazione inferiore sarà solo lineare fino a qualche segnale minimo, al di sotto del quale la molecola non sarà più rilevabile. I valori segnale-rumore del software Xcalibur dipendono dal rapporto m/z della molecola; Di conseguenza, i composti di ciascuna classe lipidica, contenenti diverse combinazioni di acidi grassi, avranno valori di rumore diversi. Inoltre, i valori di LOQ/LOQ sono strettamente legati al tipo di matrice e, quando la quantificazione del lipidoma viene effettuata in diversi tessuti di roditori, dovrebbe riflettersi nella valutazione dei LOQ per ciascun tipo di tessuto individualmente.
Infatti, il metodo offre un elevato intervallo di quantificazione dinamica lineare fino a quattro ordini di grandezza33 e un'ottima sensibilità per coprire i più importanti lipidi strutturali endogeni, che possono essere ulteriormente aumentati da miglioramenti tecnici nell'acquisizione della SM32. I coefficienti di variazione delle concentrazioni lipidiche medie erano per lo più inferiori al 15%, il che significa che la lipidomica del fucile è conforme ai requisiti della Food and Drug Administration (FDA) come metodo da prendere in considerazione per gli studi di buona pratica di laboratorio (GLP) e buona pratica clinica (GCLP)34.
Tuttavia, va notato che, a causa della loro diversa polarità, alcune classi lipidiche diventano molto più influenzate dal contributo di specifici FA coniugati. Ciò porta alla distorsione nella risposta di intensità in miscele equimolari contenenti un'ampia gamma di FA coniugati, con conseguente bias di quantificazione, come evidenziato da Koivusalo et al.35 per le classi di fosfolipidi. Da notare, questi autori hanno presentato dati per una vasta gamma di FA, da 24-48 lunghezza della catena, che probabilmente non riflette la situazione in un campione biologico reale. La risposta per le specie polinsaturi è stata superiore del 40% rispetto alle specie completamente sature, ma questo effetto è stato osservato solo per concentrazioni più elevate. Quando la miscela è stata progressivamente diluita, l'effetto dell'insaturazione è gradualmente diminuito e praticamente scomparso a 0,1 pmol/μL per specie. Inoltre, tutte le misurazioni sono state effettuate su strumenti a trappola ionica e tripli quadrupoli e non su apparecchiature Q-Exactive.
Un altro vantaggio del flusso di lavoro presentato è la sua flessibilità tecnica, che consente l'adattamento a requisiti specifici del progetto. Ad esempio, qualsiasi protocollo di estrazione lipidica - come i metodi modificati Bligh e Dyer36, metil terz-butil etere 37 o butanolo-metanolo38 - può essere utilizzato per preparare un estratto lipidico totale prima dell'analisi MS. Il principale limite dell'estrazione cloroformio-metanolo è che la fase inferiore contiene la frazione lipidica, il che complica il lavoro di routine e soprattutto l'automazione; Inoltre, deve essere considerata la tossicità del cloroformio. L'estrazione con terz-butil etere è ampiamente utilizzata per l'analisi lipidica di campioni di plasma37 ed è stata proposta una versione automatizzata21. In questo caso, abbiamo scelto il metodo BUME perché fornisce recuperi ancora migliori per le classi lipidiche PG, PI, PA e PS38, un minore consumo di solvente e la possibilità di automazione39, mentre le concentrazioni complessive quantificate per i campioni di tessuto estratti con tutti e tre i metodi erano comparabili. Inoltre, mentre l'estrazione del campione è stata eseguita manualmente nel lavoro corrente, è anche possibile ottenere risultati riproducibili e precisi da grandi set di campioni mediante la preparazione automatica del campione e l'estrazione dei lipidi in un formato a 96 pozzetti40,41. Ciò consente di implementare l'analisi lipidomica in studi clinici e tossicologici su larga scala.
Nel lavoro attuale, abbiamo eseguito l'acquisizione MS di modi positivi e negativi separatamente senza commutazione di polarità come descritto da Schuhmann et al.42. La stabilità del segnale nanomate è migliore in modalità negativa per una soluzione leggermente meno concentrata rispetto alla modalità positiva. Inoltre, abbiamo sviluppato una procedura completamente tracciabile con il software di conversione da file .raw a mzML, che fornisce i valori da specificare nel software LipidXplorer - con questo, non sono necessari calcoli manuali della pendenza della risoluzione. Abbiamo anche applicato diverse sostituzioni di impostazione del rumore perché, in modalità positiva, i livelli di rumore degli spettri sono più alti rispetto alla modalità negativa. Tutti i passaggi sono stati ottimizzati per analisi di routine tracciabili e ad alto rendimento.
Per l'identificazione, l'analisi lipidomica shotgun può sfruttare il comportamento unico di diverse classi lipidiche, che formano addotti unici in diverse modalità di polarità. In questo metodo, le specie di PC e PE con la stessa massa molecolare che si sovrappongono nel modo di ionizzazione positiva possono essere completamente separate nella modalità di ionizzazione negativa, poiché il PC forma addotti acetato o formiato e il PE viene ionizzato in forma deprotonata. Inoltre, la deconvoluzione strutturale (parziale) è possibile per il metodo utilizzando non solo la formula molecolare ma anche la struttura degli acidi grassi sfusi. Ad esempio, l'identificazione FA sul livello del carbonio totale e del conteggio totale dell'insaturazione funziona per tutti i fosfolipidi, DAG, TAG e liso-fosfolipidi. La quantificazione bottom-up di ciascuna forma isomerica può essere parzialmente eseguita per alcune delle classi di fosfolipidi43 , ma è molto più complessa per DAG e TAG a causa della risposta ineguale del segnale di diversi FA negli spettri MS2.
È altresì importante sottolineare la necessità di attuare adeguate procedure di controllo della qualità, pienamente in linea con le recenti iniziative in questo settore44. Poiché desideriamo garantire un'adeguata tracciabilità e riproducibilità dei dati tra e all'interno del laboratorio, sono state adottate una serie di misure come la corretta randomizzazione dei campioni per tutte le fasi dell'analisi, il lavoro con miscele standard certificate dal fornitore, l'inclusione di campioni di controllo qualità, procedure per verificare l'accettazione o il rifiuto dei lotti e la creazione di un database interno per monitorare le prestazioni del controllo qualità a lungo termine. Coerente con queste iniziative è anche la necessità di un metodo standardizzato per affrontare la stabilità del campione. In generale, la maggior parte dei lipidi strutturali non è influenzata dall'ossidazione lipidica, che è più rilevante per le ossipiline, i lipidi ossidati e gli acidi grassi polinsaturi che sono, quindi, molto più sensibili alle condizioni di conservazione e manipolazione. Tuttavia, la corretta valutazione della stabilità del campione è ancora un compito tecnicamente impegnativo.
Tuttavia, questo protocollo ha una limitazione quando si tratta di determinare i livelli submolecolari dei composti. Considerando che non vi è separazione dell'estratto totale, tutte le isoforme di lipidi con la stessa massa molecolare ma diverse composizioni di acidi grassi sono fuse nell'analisi MS. Per la maggior parte delle classi, è possibile ottenere una deconvoluzione parziale della struttura utilizzando i rapporti di frammentazione dei frammenti di acidi grassi residui in MS2. Tuttavia, la quantificazione indipendente di ciascuna isoforma rimane un compito particolarmente impegnativo a causa delle grandi differenze nei comportamenti di frammentazione delle diverse isoforme e del fatto che gli standard chimici puri non sono disponibili in una varietà sufficientemente ampia per definire i valori di compensazione. Un'altra limitazione è che il processo ESI genera inevitabilmente artefatti, con conseguente generazione artificiale di picchi per alcuni lipidi come DAG, Pas e FA, che possono portare a una quantificazione errata.
Successivamente, riassumiamo le parti più critiche del protocollo in base alla nostra esperienza. Il primo è legato al fatto che ogni tipo di tessuto di topo ha un profilo lipidico unico sia in termini di quantità lipidica che di rapporti di classe. Con questo, le quantità iniziali del tessuto basate sul contenuto proteico totale prima dell'estrazione devono essere attentamente determinate al fine di non saturare il segnale MS e non lasciare l'intervallo di quantificazione dinamica a causa dell'aggregazione lipidica ad alte concentrazioni33 o - all'estremo opposto - per fornire un segnale MS sufficiente a coprire i principali composti lipidici per ciascuna classe lipidica.
Il secondo aspetto critico è quello di garantire un corretto allineamento della posizione dell'uscita del chip nano-sorgente di infusione diretta con il capillare di trasferimento dello spettrometro di massa. Considerando che la calibrazione completa dello spettrometro di massa in entrambe le modalità viene eseguita settimanalmente, lo scambio tra la sorgente di calibrazione e la configurazione del chip nano-sorgente può essere la ragione di una drammatica variabilità nell'intensità del segnale a causa del disallineamento durante l'installazione.
Un'altra parte critica del protocollo è l'attenta gestione della miscela standard interna. Poiché questa miscela contiene una quantità significativa di diclorometano, una volta aperta, dovrebbe essere consumata rapidamente per evitare una lunga conservazione e molteplici usi che portano all'evaporazione e al cambiamento di concentrazione artificiale. Inoltre, è importante una manipolazione costante della miscela standard dopo la rimozione dallo stoccaggio a -20 °C, poiché le differenze di temperatura possono portare a incoerenze di volume durante il pipettaggio con pipette a cuscino d'aria. Un'opzione sarebbe quella di sostituire il diclorometano nel tampone di risospensione standard con metanolo puro, che potrebbe migliorare la praticità di manipolazione ma potrebbe influire negativamente sulla solubilità di alcune classi lipidiche e, quindi, ridurre l'accuratezza della quantificazione per queste classi lipidiche.
L'ultima parte critica è l'elaborazione dei dati. Il flusso di lavoro di elaborazione dei dati combina la conversione del software Peak by Peak dal formato .raw al formato .mzML, applicando la media di scansione MS2 e la filtrazione del rumore MS1 e MS2, nonché il prelievo dei picchi e la compressione dei dati. In alternativa, il software Proteowizard può essere utilizzato anche per la conversione dei dati, ma, in questo caso, diverse impostazioni in LipidXplorer devono essere definite manualmente. Tutta la complessità della lipidomica shotgun è particolarmente concentrata sulla fase di deconvoluzione degli spettri MS1 e MS2 a iniezione diretta. Il software open source LipidXplorer importa gli spettri convertiti dal formato di file mzML in base alla precisione della massa, alla risoluzione di massa e alla pendenza della sua modifica con l'aumentare di m / z. Il software unisce più singoli spettri MS e MS/MS acquisiti all'interno di un'analisi. Successivamente, allinea i singoli picchi all'interno di diverse esecuzioni di campioni e, in ogni gruppo di picchi allineati, sostituisce le loro masse con la loro massa media ponderata per la singola intensità, mentre le loro abbondanze in ciascun file di dati rimangono intatte. Le masse rappresentative dei cluster di picco allineati e le singole intensità di picco sono memorizzate in un database di scansione master. Il database di scansione master contiene tutti gli spettri MS1 e MS2 generati per tutti i campioni nel batch e può essere deconvoluto alle identificazioni lipidiche mediante query scritte nel linguaggio di query di frammentazione molecolare (MFQL).
Nel complesso, il metodo copre l'identificazione dei lipidi DAG, TAG e SE basati sulla modalità positiva e lipidi PC, PE, PS, PI, PA, PG, SM, LPC e LPE basati sull'acquisizione in modalità negativa. Durante l'identificazione dei lipidi, viene eseguita la correzione isotopica per MS1 e MS2 e le intensità regolate sono riportate nel file di output .xlsx. In alternativa, sono disponibili diversi altri software per elaborare i dati del fucile, come ALEX45 e LipidHunter46.
Gli acidi grassi - i principali componenti della membrana nucleare e cellulare - sono immagazzinati sotto forma di fosfolipidi per un'ulteriore conversione in molecole bioattive. Possono essere convertiti dagli enzimi lisofosfolipidi aciltransferasi in lisofosfolipidi attraverso la via Lands47. L'enzima LPCAT3, ad esempio, è noto per mostrare un'elevata specificità per incorporare AA negli intermedi lisofosfatidilcolina e lisofosfatidilserina. L'espressione relativamente elevata di questi enzimi è stata riportata all'interno delle cellule infiammatorie, come i macrofagi alveolari e le cellule epiteliali bronchiali47, portando al rilascio di proporzioni relative maggiori di fosfolipidi contenenti AA in quelle cellule. Tuttavia, in seguito alla loro liberazione dai fosfolipidi di membrana da parte della fosfolipasi A2, la conversione dei PUFA in vari tipi di mediatori lipidici è catalizzata da molti enzimi48. Inoltre, questi PUFA possono diventare il substrato per la produzione di prostaglandine pro-infiammatorie e anti-infiammatorie attraverso l'azione degli enzimi cicloossigenasi (COX)-1 e COX-247. I fosfolipidi sono una delle fonti di mediatori lipidici rilasciati in particolari luoghi in cui questi mediatori sono tenuti a dimostrare i loro effetti biologici.
Il polmone è un organo complesso che comprende più tipi di cellule, ognuna delle quali svolge ruoli sovrapposti e di nicchia nel facilitare il normale sviluppo e la funzione polmonare. Sono stati condotti solo pochi studi per isolare e profilare diversi tipi di cellule nel topo o nel polmone umano (ad esempio, cellule alveolari di tipo 2)49,50; Altri tipi principali di cellule polmonari non sono stati caratterizzati. Un altro interessante studio51 è stato condotto per isolare ed eseguire analisi lipidiche di cellule immunitarie endoteliali, epiteliali, mesenchimali e miste nel polmone del topo. È stato osservato che la concentrazione di PUFA incorporati in PCO e PG era arricchita nelle cellule immunitarie. Considerando il fatto che la CS induce un aumento delle cellule immunitarie all'interno del polmone (4-5 volte), come valutato dal lavaggio broncoalveolare (BAL)52, i cambiamenti lipidici totali osservati in questo studio potrebbero essere spiegati da un aumento cumulativo del reclutamento delle cellule immunitarie nel polmone del topo.
In conclusione, la distorsione osservata degli acidi grassi monoinsaturi e dei PUFA incorporati nei fosfolipidi può riflettere l'eccesso di alcuni FA all'interno della cellula e/o costituire una risorsa intracellulare di precursori dell'ossilpina che sono sovraprodotti sotto stress ossidativo e condizioni infiammatorie. Tuttavia, ulteriori dati su EPA libero, DHA e altre ossilipine sono essenziali per chiarire questo punto e non rientrano nell'ambito dell'attuale applicazione del metodo.
Divulgazioni
Tutti gli autori sono dipendenti di Philip Morris International. Philip Morris International è l'unica fonte di finanziamento e sponsor di questo progetto.
Riconoscimenti
Gli autori desiderano ringraziare il team di studio e in particolare riconoscere l'assistenza tecnica e il supporto dei team di bioricerca e aerosol presso PMI R & D, Philip Morris International Research Laboratories Pte. Ltd., Singapore, e PMI R&D, Philip Morris Products S.A., Neuchâtel, Svizzera. Gli autori ringraziano Sam Ansari per aver gestito il biobanking e riconoscono il sostegno di Sindhoora Bhargavi Gopala Reddy per la redazione di una bozza del manoscritto.
Materiali
| Name | Company | Catalog Number | Comments |
| 1, 5, and 2 mL self-lock tubes | Eppendorf | 30120086, 30120094 | |
| 3 mm stainless still beads | Qiagen | 69997 | |
| 4.1 µm nozzle chip | Advion | HD-D-384 | |
| Acetic acid | Sigma Aldrich | 45754-100ML-F | |
| Ammonium acetate | Honeywell | 14267-25G | |
| Ammonium bicarbonate | Sigma Aldrich | 09830-500G | |
| Bovine serum albumin standard, 2 mg/mL | Thermo Scientific | 23209 | |
| Butanol | Honeywell | 33065-2.5L | |
| Chloroform | Sigma Aldrich | 650498-1L | |
| Dichloromethane | Honeywell | 34856-1L | |
| Ethyl acetate | Honeywell | 33211 | |
| Greiner CELLSTAR 96 well plates | Sigma | M9686 | |
| Heptane | Sigma Aldrich | 34873-2.5L | |
| Isopropanol | Fisher Scientific | A461 | |
| Methanol | Fisher Scientific | A456 | |
| Mouse pooled plasma | BioIvt | ||
| Mouse SPLASH standard | Avanti Polar Lipids | 330710X | Internal standard |
| Nunc 96-flat bottom well transparent plates | VWR | 62409-068 | |
| Plastic spatula | Sigma | Z560049-300EA | |
| Quick Start Bradford 1x Dye reagent | BioRad | 5000205 | |
| Serum diluent | Sigma Aldrich | D5197 | |
| Equipment/software | |||
| CryoPrep CP02 impactor instrument | Covaris | Magnetic hammer | |
| Centrifuge 5427R | Eppendorf | . | Centrifuge |
| ChipSoft 8.3 | Advion Biosciences | . | Software to set up method and acquisition on the Triversa nanomate robot |
| LipidXplorer 1.2.8.1 | N/A | . | Software to identify lipids |
| Peak By Peak | SpectroSwiss | . | Software to convert .raw data from MS to .mzml format |
| ProteoWizard | ProteoWizard | . | Alternative (open source) software to convert .raw data from MS to .mzml format |
| Q-Exactive MS | Thermo Fisher | . | High resolution orbitrap mass spectrometer |
| Qiagen Tissue Lyser II | Qiagen | . | Tissue lyser |
| SpeedVac SPD140DDA | Thermo Fisher | . | Vacuum concentrator |
| Tecan Infinite M nano plus | Tecan | . | Plate reader |
| ThermoMixer C | Eppendorf | . | Thermomixer |
| TriVersa Nanomate | Advion Biosciences | . | Direct infusion nano-source |
| Xcalibur 4.3 | Thermo Scientific | . | Software to set up method and acquisition on the Q-Exactive MS |
Riferimenti
- Salehi, N., Janjani, P., Tadbiri, H., Rozbahani, M., Jalilian, M. Effect of cigarette smoking on coronary arteries and pattern and severity of coronary artery disease: A review. Journal of International Medical Research. 49 (12), 3000605211059893 (2021).
- Churg, A., Sin, D. D., Wright, J. L. Everything prevents emphysema: Are animal models of cigarette smoke-induced chronic obstructive pulmonary disease any use. American Journal of Respiratory Cell and Molecular Biology. 45 (6), 1111-1115 (2011).
- Lee, G., Walser, T. C., Dubinett, S. M. Chronic inflammation, chronic obstructive pulmonary disease, and lung cancer. Current Opinion in Pulmonary Medicine. 15 (4), 303-307 (2009).
- Titz, B., et al. Effects of cigarette smoke, cessation, and switching to two heat-not-burn tobacco products on lung lipid metabolism in C57BL/6 and Apoe-/- mice-An integrative systems toxicology analysis. Toxicological Sciences. 149 (2), 441-457 (2016).
- Morissette, M. C., Shen, P., Thayaparan, D., Stampfli, M. R. Disruption of pulmonary lipid homeostasis drives cigarette smoke-induced lung inflammation in mice. European Respiratory Journal. 46 (5), 1451-1460 (2015).
- Jubinville, E., et al. Interplay between cigarette smoking and pulmonary reverse lipid transport. European Respiratory Journal. 50 (3), 1700681 (2017).
- Han, X., Gross, R. W. Shotgun lipidomics: Electrospray ionization mass spectrometric analysis and quantitation of cellular lipidomes directly from crude extracts of biological samples. Mass Spectrometry Reviews. 24 (3), 367-412 (2005).
- Titz, B., et al. Multi-omics systems toxicology study of mouse lung assessing the effects of aerosols from two heat-not-burn tobacco products and cigarette smoke. Computational and Structural Biotechnology Journal. 18, 1056-1073 (2020).
- Hartung, T., et al. Systems toxicology: Real world applications and opportunities. Chemical Research in Toxicology. 30 (4), 870-882 (2017).
- Talikka, M., et al., Lyubimov, A., et al. Systems Toxicology. Encyclopedia of Drug Metabolism and Interactions. , (2016).
- Ejsing, C. S., et al. Global analysis of the yeast lipidome by quantitative shotgun mass spectrometry. Proceedings of the National Academy of Sciences of the United States of America. 106 (7), 2136-2141 (2009).
- Papan, C., et al. Systematic screening for novel lipids by shotgun lipidomics. Analytical Chemistry. 86 (5), 2703-2710 (2014).
- Carvalho, M., et al. Effects of diet and development on the Drosophila lipidome. Molecular Systems Biology. 8, 600 (2012).
- Strittmatter, N., et al. Shotgun lipidomic profiling of the NCI60 cell line panel using rapid evaporative ionization mass spectrometry. Analytical Chemistry. 88 (15), 7507-7514 (2016).
- Lydic, T. A., et al. Rapid and comprehensive 'shotgun' lipidome profiling of colorectal cancer cell derived exosomes. Methods. 87, 83-95 (2015).
- Sampaio, J. L., et al. Membrane lipidome of an epithelial cell line. Proceedings of the National Academy of Sciences of the United States of America. 108 (5), 1903-1907 (2011).
- Grzybek, M., et al. Comprehensive and quantitative analysis of white and brown adipose tissue by shotgun lipidomics. Molecular Metabolism. 22, 12-20 (2019).
- Stegemann, C., et al. Comparative lipidomics profiling of human atherosclerotic plaques. Circulation: Cardiovascular Genetics. 4 (3), 232-242 (2011).
- Tajima, Y., et al. Lipidomic analysis of brain tissues and plasma in a mouse model expressing mutated human amyloid precursor protein/tau for Alzheimer's disease. Lipids in Health and Disease. 12, 68 (2013).
- Gross, R. W., Han, X. Shotgun lipidomics of neutral lipids as an enabling technology for elucidation of lipid-related diseases. American Journal of Physiology: Endocrinology and Metabolism. 297 (2), 297-303 (2009).
- Surma, M. A., et al. An automated shotgun lipidomics platform for high throughput, comprehensive, and quantitative analysis of blood plasma intact lipids. European Journal of Lipid Science and Technology. 117 (10), 1540-1549 (2015).
- Heiskanen, L. A., Suoniemi, M., Ta, H. X., Tarasov, K., Ekroos, K. Long-term performance and stability of molecular shotgun lipidomic analysis of human plasma samples. Analytical Chemistry. 85 (18), 8757-8763 (2013).
- Zhang, S., Van Pelt, C. K. Chip-based nanoelectrospray mass spectrometry for protein characterization. Expert Review of Proteomics. 1 (4), 449-468 (2004).
- Surma, M. A., et al. Mouse lipidomics reveals inherent flexibility of a mammalian lipidome. Scientific Reports. 11 (1), 19364 (2021).
- Herzog, R., Schwudke, D., Shevchenko, A. LipidXplorer: Software for quantitative shotgun lipidomics compatible with multiple mass spectrometry platforms. Current Protocols in Bioinformatics. 43, 11-30 (2013).
- R: A language and environment for statistical computing. R: Foundation for Statistical Computing Available from: https://www.R-project.org/ (2018)
- Phillips, B., et al. A 7-month cigarette smoke inhalation study in C57BL/6 mice demonstrates reduced lung inflammation and emphysema following smoking cessation or aerosol exposure from a prototypic modified risk tobacco product. Food and Chemical Toxicology. 80, 328-345 (2015).
- Phillips, B., et al. A six-month systems toxicology inhalation/cessation study in ApoE(-/-) mice to investigate cardiovascular and respiratory exposure effects of modified risk tobacco products, CHTP 1.2 and THS 2.2, compared with conventional cigarettes. Food and Chemical Toxicology. 126, 113-141 (2019).
- Zemski Berry, K. A., Murphy, R. C. Electrospray ionization tandem mass spectrometry of glycerophosphoethanolamine plasmalogen phospholipids. Journal of the American Society for Mass Spectrometry. 15 (10), 1499-1508 (2004).
- Engelmann, B. Plasmalogens: Targets for oxidants and major lipophilic antioxidants. Biochemical Society Transactions. 32, 147-150 (2004).
- Nagan, N., Zoeller, R. A. Plasmalogens: Biosynthesis and functions. Progress in Lipid Research. 40 (3), 199-229 (2001).
- Southam, A. D., Weber, R. J., Engel, J., Jones, M. R., Viant, M. R. A complete workflow for high-resolution spectral-stitching nanoelectrospray direct-infusion mass-spectrometry-based metabolomics and lipidomics. Nature Protocols. 12 (2), 310-328 (2016).
- Yang, K., Han, X. Accurate quantification of lipid species by electrospray ionization mass spectrometry - Meet a key challenge in lipidomics. Metabolites. 1 (1), 21-40 (2011).
- Zullig, T., Trotzmuller, M., Kofeler, H. C. Lipidomics from sample preparation to data analysis: a primer. Analytical and Bioanalytical Chemistry. 412 (10), 2191-2209 (2020).
- Koivusalo, M., Haimi, P., Heikinheimo, L., Kostiainen, R., Somerharju, P. Quantitative determination of phospholipid compositions by ESI-MS: Effects of acyl chain length, unsaturation, and lipid concentration on instrument response. Journal of Lipid Research. 42 (4), 663-672 (2001).
- Bligh, E. G., Dyer, W. J. A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology. 37 (8), 911-917 (1959).
- Matyash, V., Liebisch, G., Kurzchalia, T. V., Shevchenko, A., Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. Journal of Lipid Research. 49 (5), 1137-1146 (2008).
- Lofgren, L., Forsberg, G. B., Stahlman, M. The BUME method: A new rapid and simple chloroform-free method for total lipid extraction of animal tissue. Scientific Reports. 6, 27688 (2016).
- Lofgren, L., et al. The BUME method: A novel automated chloroform-free 96-well total lipid extraction method for blood plasma. Journal of Lipid Research. 53 (8), 1690-1700 (2012).
- Jung, H. R., et al. High throughput quantitative molecular lipidomics. Biochimica et Biophysica Acta. 1811 (11), 925-934 (2011).
- Lavrynenko, O., et al. Ceramide ratios are affected by cigarette smoke but not heat-not-burn or e-vapor aerosols across four independent mouse studies. Life Sciences. 263, 118753 (2020).
- Schuhmann, K., et al. Shotgun lipidomics on a LTQ Orbitrap mass spectrometer by successive switching between acquisition polarity modes. Journal of Mass Spectrometry. 47 (1), 96-104 (2012).
- Schuhmann, K., et al. Quantitative fragmentation model for bottom-up shotgun lipidomics. Analytical Chemistry. 91 (18), 12085-12093 (2019).
- Lipidomics Standards Initiative Consortium. Lipidomics needs more standardization. Nature Metabolism. 1 (8), 745-747 (2019).
- Husen, P., et al. Analysis of lipid experiments (ALEX): A software framework for analysis of high-resolution shotgun lipidomics data. PloS One. 8 (11), 79736 (2013).
- Ni, Z., Angelidou, G., Lange, M., Hoffmann, R., Fedorova, M. LipidHunter identifies phospholipids by high-throughput processing of LC-MS and shotgun lipidomics datasets. Analytical Chemistry. 89 (17), 8800-8807 (2017).
- Zemski Berry, K. A., Murphy, R. C., Kosmider, B., Mason, R. J. Lipidomic characterization and localization of phospholipids in the human lung. Journal of Lipid Research. 58 (5), 926-933 (2017).
- Ghosh, M., Tucker, D. E., Burchett, S. A., Leslie, C. C. Properties of the Group IV phospholipase A2 family. Progress in Lipid Research. 45 (6), 487-510 (2006).
- Besnard, V., et al. Deletion of Scap in alveolar type II cells influences lung lipid homeostasis and identifies a compensatory role for pulmonary lipofibroblasts. Journal of Biological Chemistry. 284 (6), 4018-4030 (2009).
- Plantier, L., et al. Activation of sterol-response element-binding proteins (SREBP) in alveolar type II cells enhances lipogenesis causing pulmonary lipotoxicity. Journal of Biological Chemistry. 287 (13), 10099-10114 (2012).
- Kyle, J. E., et al. Cell type-resolved human lung lipidome reveals cellular cooperation in lung function. Scientific Reports. 8 (1), 13455 (2018).
- Lugg, S. T., Scott, A., Parekh, D., Naidu, B., Thickett, D. R. Cigarette smoke exposure and alveolar macrophages: mechanisms for lung disease. Thorax. 77 (1), 94-101 (2022).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneEsplora altri articoli
This article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati