
Zum Anzeigen dieser Inhalte ist ein JoVE-Abonnement erforderlich. Melden Sie sich an oder starten Sie Ihre kostenlose Testversion.
Method Article
Laser-Capture Microdissection RNA-Sequenzierung für räumliche und zeitliche gewebespezifische Genexpressionsanalyse in Pflanzen
In diesem Artikel
Zusammenfassung
Hier wird ein Protokoll zur Laser-Capture-Mikrodissektion (LCM) von Pflanzengeweben vorgestellt. LCM ist eine mikroskopische Technik zur kontaminationsfreien Isolierung von Gewebebereichen. Das Verfahren umfasst Gewebefixierung, Paraffineinbettung, Schnitt, LCM und RNA-Extraktion. RNA wird in der nachgeschalteten gewebespezifischen, zeitlich aufgelösten Analyse von Transkriptomen eingesetzt.
Zusammenfassung
Die Entwicklung eines komplexen mehrzelligen Organismus wird von unterschiedlichen Zelltypen bestimmt, die unterschiedliche Transkriptionsprofile aufweisen. Um transkriptionelle regulatorische Netzwerke zu identifizieren, die Entwicklungsprozesse steuern, ist es notwendig, die räumlichen und zeitlichen Genexpressionsprofile dieser einzelnen Zelltypen zu messen. Daher ist die Einsicht in die räumlich-zeitliche Kontrolle der Genexpression unerlässlich, um zu verstehen, wie biologische und Entwicklungsprozesse reguliert werden. Hier beschreiben wir eine Laser-Capture-Mikrodissektion (LCM) Methode, um eine kleine Anzahl von Zellen aus drei Gersten-Embryo-Organen während eines Zeitverlaufs während der Keimung zu isolieren, gefolgt von Transkriptprofilierung. Die Methode besteht aus Gewebefixierung, Gewebeverarbeitung, Paraffineinbettung, Schnitt, LCM und RNA-Extraktion, gefolgt von Echtzeit-PCR oder RNA-Seq. Diese Methode hat es uns ermöglicht, räumliche und zeitliche Profile von Samenorgan-Transkriptomen aus unterschiedlicher Anzahl von Zellen (Zehn bis Hunderte) zu erhalten, was eine viel größere Gewebespezifität als typische Massengewebeanalysen bietet. Aus diesen Daten konnten wir transkriptionelle regulatorische Netzwerke definieren und vergleichen sowie kandidatenregulatorische Transkriptionsfaktoren für einzelne Gewebe vorhersagen. Die Methode sollte auf andere Pflanzengewebe mit minimaler Optimierung anwendbar sein.
Einleitung
Pflanzenentwicklung und -wachstum beinhalten die koordinierte Wirkung transkriptionsregulatorischer Netzwerke innerhalb verschiedener Zellen, die in einer komplexen zellulären Umgebung existieren. Um die Aktivität dieser regulatorischen Netzwerke zu verstehen, benötigen wir das Wissen über die räumliche und zeitliche Genexpression innerhalb verschiedener Zelltypen über Entwicklungsstadien hinweg. Aufgrund der technischen Herausforderung, eine kleine Anzahl von Zellen zu isolieren und zu analysieren, werden jedoch häufiger Analysen der Genexpression in ganzen Organen oder Massengewebeproben durchgeführt. Die Methode, die wir hier beschreiben, hat es ermöglicht, eine räumliche und zeitliche gewebespezifische Transkriptomanalyse durch Kopplung von LCM mit RNA-seq zu erhalten.
LCM wurde vor zwei Jahrzehnten von Emmert-Buck und Kollegen1entwickelt. Die Technik ermöglichte es den Forschern, Einzelne Zellen oder Zellcluster mithilfe direkter mikroskopischer Visualisierung und Manipulation mit einem schmalen Strahllaser1präzise aus ihrer Umgebung zu isolieren. Seitdem ist die Methode weit verbreitet in der Krebsbiologie und Pathologie2,3. Viele Pflanzenforschungsgruppen haben auch LCM für die Verwendung mit verschiedenen Pflanzenarten und verschiedenen Gewebetypen4,5,6,7,8,9,,10,11angepasst. In jüngster Zeit haben mehrere Papiere auch LCM auf Eudikot und Monocot Samen verwendet, um Embryonen, Endospermen und andere Samenstrukturen während der Samenentwicklung und Keimung10,12,13zu untersuchen. Die meisten anderen häufig verwendeten einzelligen Isolationsmethoden wie Mikropipettieren, Zellsortierung, magnetische Trennung und mikrofluidische Plattformen hängen von der enzymatischen Verdauung oder mechanischen Homogenisierung ab, um Zellen zu dissoziieren. Dies kann die Genexpression stören und technische Artefakte einführen, die die Dateninterpretation verwirren14,15. Diese Methoden erfordern auch Vorkenntnisse über Markergene für jeden Zelltyp, um die dissoziierten Zellen mit ihrer räumlichen Position und ihrem wahren Zelltyp in Beziehung zu setzen. Eine weitere Gruppe von Techniken hängt von der Affinitäts-basierten Isolierung subzellulärer Strukturen anstelle von ganzen Zellen ab, z.B. INTACT (Isolation of Nuclei Tagged in Cell Types) und TRAP (Translating Ribosome Affinity Purification)16,17. Die Affinitätskennzeichnung und -reinigung von Kernen oder Ribosomen ist jedoch bei Pflanzenarten, die keine gut etablierten Transformationsprotokolle haben, technisch eine Herausforderung. LCM nutzt die schnelle Gewebefixierung, um Transkriptionspiegel und konventionelle histologische Identifikation durch direkte Visualisierung von Zellen innerhalb ihres normalen Gewebe-Organ-Kontextes zu erhalten, wodurch diskrete Zellen in kurzer Zeit isoliert werden können18,19.
Das hier vorgestellte Protokoll ist eine optimierte Methode zur Isolierung bestimmter Zellen oder Zelltypen aus den Gewebeabschnitten von Getreidesamen, die auf die meisten Zellen angewendet werden kann, die histologisch identifiziert werden können. LCM bietet eine kontaktfreie Methode der Zellisolierung, die die Kontamination erheblich reduziert und die Integrität der zurückgewonnenen RNA erhöht. Darüber hinaus veranschaulicht die Methode die Leistungsfähigkeit von LCM in groß angelegten genomweiten Studien, beginnend mit kleinen Mengen biologischer Materialien. Wir beschreiben auch die lineare Amplifikation von RNA zur Erzeugung von ausreichendem Inputmaterial für nachgeschaltete Transkript-/Transkriptomanalysen.
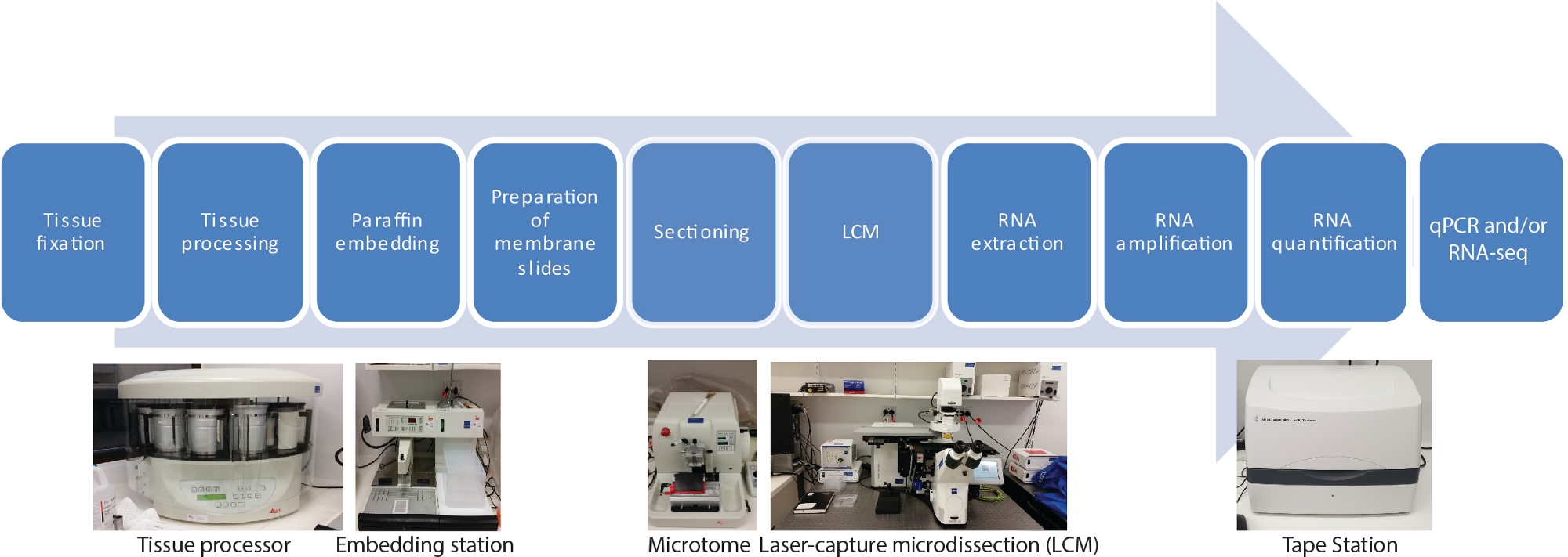
Es gibt zehn Hauptschritte in diesem LCM-RNA-Seq-Protokoll für räumliche und zeitliche gewebespezifische Transkriptome, einschließlich Fixierung von Gewebeproben, Dehydrierung, Paraffinininfiltration, Einbettung, Schnitt, LCM, RNA-Extraktion, RNA-Verstärkung, RNA-Quantifizierung und qRT-PCR und/oder RNA-seq (Abbildung 1).

Abbildung 1: Flussdiagramm von LCM, gefolgt von RNA-seq oder qRT-PCR. LCM ist eine räumlich präzise und berührungslose Technik, um Zellen aus festen Gewebeabschnitten mittels eines Laserstrahls unter mikroskopischer Visualisierung zu sammeln. Der Prozess beginnt mit der Fixierung von Gewebeproben, gefolgt von einer Dehydrierung mit einer Gradientenreihe von Ethanol und Xylol und endet mit Paraffinininfiltration. Der Prozess kann mit hilfe eines Tissueprozessors vollautomatisiert werden. Sobald das Gewebe mit Paraffin infiltriert ist, wird es mit einer Einbettstation in eine Form mit geschmolzenem Paraffin eingebettet. Das Schneiden erfolgt mit Mikrotome auf die gewünschte Dicke eingestellt. Dias werden vorbereitet und LCM unmittelbar vor der RNA aus gefangenen Zellen extrahiert. Auf die RNA-Extraktion folgen direkt zwei Runden RNA-Verstärkung vor qRT-PCR und/oder RNA-seq. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Protokoll
Da das Endprodukt RNA ist, achten Sie darauf, die Arbeit mit RNases nicht zu kontaminieren. Das Tragen von Handschuhen ist ein Muss. Verwenden Sie Diethylpyrocarbonat (DEPC) -behandeltes Wasser, Puffer usw. Autoklav-Puffer und backen Glaswaren vor der Verwendung.
1. Gewebefixierung
- Vorbereitung Fixierung der Wahl je nach Art und Gewebetypen; für Gerstensaatgut verwenden Sie Farmer es fixative (75% Ethanol, 25% Eisessigsäure (v/v)).
- Kühlen Sie das Fixativ auf Eis vor der Ernte von Geweben.
- Sammeln Sie das Pflanzenmaterial von Interesse und sezieren Sie es bei Bedarf in Stücke von geeigneter Größe, um in die ausgewählte Einbettform zu passen. Für Gerstensamen, schneiden Sie den Samen in die Hälfte längs, um das Eindringen der fixativen Lösung zu helfen und in die Einbettform zu passen.
- Tauchen Sie das Gewebe in mindestens 10x Volumen eiskalt fixativ. Für Gerstensamen, tauchen Sie den Samen in die Hälfte in die Fixierung geschnitten.
- Verwenden Sie Vakuuminfiltration, um die Penetration von Fixativ zu beschleunigen. Das Gewebe sollte nach der Vakuuminfiltration des Fixiativs sinken. Für Gerstensamen 30 min Vakuuminfiltration verwenden.
- Ersetzen Sie das Fixativ und inkubieren Sie bei 4 °C, damit das Fixativ vollständig in das Gewebe eindringen kann. Für Gerstensamen die Proben über Nacht inkubieren (ca. 12-16 h).
HINWEIS: Dünnere oder kleine Gewebe benötigen aufgrund der höheren Diffusionsrate des Fixativs in das Gewebe eine kürzere Fixierungszeit. - Entfernen Sie das Gewebe aus dem Fixierungsmittel und übertragen Sie das Gewebe in Kassetten und beginnen Sie dann mit der Gewebeverarbeitung.
HINWEIS: Kleines oder zerbrechliches Gewebe, wie z. B. Blattgewebe, kann zur Fixierung in Kassetten gelegt werden, um sicherzustellen, dass es während der Fixierung nicht beschädigt wird. Biopsiebeutel, Pads oder Wraps könnten verwendet werden, um das Gewebe während der Gewebefixierung und der Gewebeverarbeitungsschritte sicher in den Kassetten zu halten.
2. Gewebeverarbeitung
- Verwenden Sie einen automatisierten Gewebeprozessor in Schritt 2 mit mindestens 10 Lösungskammern und 2 beheizten Paraffinkammern (siehe Materialtabelle).
- Prüfen Sie, ob in jeder Kammer ausreichende Lösungsmengen vorhanden sind; Lösungen nach allen wenigen Verwendungen des Gewebeprozessors zu ersetzen.
- Kassetten mit Gewebe in den Metallkorb legen. Befestigen Sie den Metallkorb an seinem Halter über Kammer 1. Der Halter dreht und "dunk und dip" die Kassetten in die Kammern, nach dem vorgesehenen Programm.
- Stellen Sie das Programm ein, indem Sie Knopfklicks auf die Bedienfelder des Tissueprozessors ausführen, bei denen die Dauer für jede Kammer eingestellt wird.
- Drücken Sie die Schaltfläche "Starten", um das Verarbeitungsprogramm zu starten. Das folgende Programm ist für Gerstensamen konzipiert und läuft über Nacht (ca. 18 h)
- Führen Sie die Austrocknung durch Eintauchen der Kassette für jeweils 1 h 30 min in der Gradientenserie von Ethanol (75%, 85%, 100%, 100% und 100% (v/v) Ethanol).
- Clearing mit Ethanol: Xylol-Gradient für 1 h 30 min jeweils um 75:25, 50:50, 25:75 (Ethanol: Xylol %, v/v). Dann tauchen Sie die Kassette für 1 h 30 min von 100% Xylol dann 100% Xylol.
- Paraffinininfiltration bei 55-60 °C für 1 h 30 min zweimal in geschmolzenem Paraffin durchführen.
HINWEIS: Die Temperatur der Paraffinheizungskammern kann auf der Rückseite des Gewebeprozessors eingestellt werden.
- Am nächsten Morgen entfernen Sie die Kassetten aus dem Gewebeprozessor und fahren Sie mit der Paraffineinbettung fort.
HINWEIS: Die Programmzeit kann zwischen verschiedenen Gewebetypen variieren. Vakuum und/oder Rührung können während der Gewebeverarbeitung verwendet werden, um die Infiltration ausgewählter Lösungen zu beschleunigen, indem sie"V" und/oder "Agitationstasten" auf dem Bedienfeld des Gewebeprozessors drücken.
3. Paraffin-Einbettung
- Verwenden Sie in diesem Schritt eine Einbettungsmaschine (siehe Materialtabelle).
- Stellen Sie vor, dass die Einbettungsmaschine mindestens ein paar Stunden vor der Einbettung einschaltet, damit das Paraffin in den Reservoirs vollständig schmelzen kann.
- Schalten Sie die kalte Platte vor dem Start ein.
- Einbetten von Proben in Formen, indem Sie die Proben mit feinen Zangen in Position halten und geschmolzenes Paraffin in die Form abgeben. Sicherstellen der ordnungsgemäßen Orientierung der Proben für jeden Versuchszweck. Bei Gerstensamen die Saat längs an der Schnittrichtung orientieren, um Längsschnitte zu erhalten.
HINWEIS: Einbettformen gibt es in verschiedenen Größen. Wählen Sie eine geeignete Größe aus, damit die Probe richtig positioniert und eingebettet werden kann. Die Ausrichtung der Proben sollte je nach Versuchsbedarf berücksichtigt werden. Wenn Längsschnitte erforderlich sind, sollte die Probe längs an der Schnittrichtung orientiert sein, während bei Querabschnitten die Probe parallel zur Schnittrichtung ausgerichtet werden sollte. - Legen Sie eine saubere Kassette auf die Form und stellen Sie sicher, dass ausreichend Paraffin die gesamte Kassette vollständig abdeckt, um die Probe auf der Kassette zu halten.
- Legen Sie die Form auf die kalte Platte und lassen Sie das Paraffin vollständig einstellen (10-20 min), bevor Sie den Block aus der Form lösen.
- Fahren Sie mit dem Schnitt fort oder übertragen Sie die Blöcke zur Lagerung auf 4 °C.
HINWEIS: Das Protokoll kann hier angehalten werden. Die eingebetteten Blöcke können bis zu drei Monate bei 4 °C gelagert werden.
4. Herstellung von Polyethylennaphthaat (PEN) Membranschlitten
- Submerge PEN Membranschlitten in RNase Deaktivierungslösung für 3 s gefolgt von zwei kurzen Wäbungen in DEPC-behandeltem Wasser, um RNasen auf den Dias zu entfernen. Trocknen Sie die Dias in einem 37 °C Inkubator, um die übrig gebliebene Lösung zu entfernen.
- UV-Behandlung der Dias mit einer UV-Lampe in einem laminaren Durchflussschrank für 30 min, um die hydrophilen Eigenschaften für eine verbesserte Paraffinhaftung zu verbessern.
5. Schnitt
- Verwenden Sie ein Mikrotom im Schnittschritt (siehe Materialtabelle).
- Legen Sie eine neue Klinge in den Messerhalter, und halten Sie immer Messerschutz, wenn nicht aktiv schneiden
VORSICHT: Mikrotomklingen sind extrem scharf und können erheblichen Schaden anrichten, wenn sie unsachgemäß behandelt werden.
HINWEIS: Es gibt zwei Verriegelungsmechanismen am Mikrotom, einer befindet sich an der Seite der Maschine und der andere am Griff des Rades. Beide sind zu engagieren, wenn sie nicht aktiv geschnitten werden. - Stellen Sie den Messerblock so an, dass er so nah wie möglich an der Probe ist, ohne sich zu berühren. Stellen Sie sicher, dass der Mikrotomarm nie in vollen Kontakt mit dem Messerblock kommt, da dies katastrophale strukturelle Schäden am Mikrotom verursacht.
- Schalten Sie die kalte Platte vor dem Start ein. Halten Sie Paraffinblöcke vor dem Schnitt auf der kalten Platte auf und kühlen Sie die Blöcke bei Bedarf während des Schnitts, um eine Enthärtung der Blöcke zu verhindern.
- Füllen Sie das Wasserbad vor dem Start mit DEPC-behandeltem Wasser und erhitzen Sie es auf 42 °C.
- Schneiden Sie Blöcke auf die gewünschte Tiefe (wo der Abschnitt, den Sie interessieren) und Abschnitt Paraffinblöcke mit der gewünschten Dicke (6-10 m) mit dem Mikrotome; ein gut geschnittener Block bildet ein "Band" am Rand der Klinge. Für Gerstensamen, Abschnitt mit 8 m Dicke.
- Tragen Sie die Bänder vorsichtig mit feinem Pinsel oder feinen Zangen vom Mikrotom ins Wasserbad, um sicherzustellen, dass das Band flach auf der Wasseroberfläche ist.
- Halten Sie eine Rutsche in einem 45°-Winkel, mit einer Aufwärtsbewegung, heben Sie ein Band aus dem Wasser auf die Rutsche und entfernen Sie vorsichtig überschüssiges Wasser mit einem fusselfreien Gewebe.
- Trockene Schlitten für 30 min bei 37 °C, um das verbleibende Wasser unter dem Paraffin zu eliminieren.
- Fahren Sie mit der Paraffinentfernung fort oder lagern Sie sie bei 4 °C in einer geschlossenen Box unter Dehydrierungsbedingungen (innerhalb weniger Tage zu verwenden).
- Entfernen Sie Paraffin, indem Sie die Dias 3x für je 20 s in Xylol waschen, gefolgt von 2x Waschungen von 30 s in 100% (v/v) Ethanol und 2x Waschungen von 30 s in 70% (v/v) Ethanol.
- Fahren Sie sofort mit der Laser-Capture-Mikrodissektion fort, nachdem Paraffin entfernt wurde.
HINWEIS: Cryosectioning ist eine alternative Methode, die erfolgreich mit LCM gekoppelt wurde. Die Probenvorbereitung für die Kryosektion wird sich unterscheiden.
6. Laser-Capture-Mikrodissektion
- Verwenden Sie ein Laser-Capture-Mikrodissektionsmikroskop (siehe Tabelle der Materialien), um Zellen aus deparaffinierten und getrockneten Gewebeabschnitten zu mikrosektieren.
- Laden Sie Folien auf die drei verfügbaren Steckplätze.
- Verwenden Sie die speziellen Klebekappen von Sammelrohren, um die gefangenen Proben zu sammeln. Die Erfassung ohne Flüssigkeit ("trockene" Sammlung) minimiert die RNase-Aktivität. Laden Sie die Sammelrohre in die verfügbaren Schlitze.
- Verschieben Sie die Bühne, um den Bereich der Zutaste zu finden, die geschnitten werden muss. Dies kann mit der Maus oder dem Joystick des LCM-Geräts oder den Pfeiltasten auf der Tastatur erfolgen.
- Um die Schnittgeschwindigkeit zu optimieren, Schneiden Energie und Fokus, Laserdruck katapultieren (LPC) Energie und Fokus, schneiden Sie zuerst auf einem leeren Segment frei von Gewebe auf dem Membranschlitten. Für Gerstensamen, Schnittgeschwindigkeit = 18, CutEnergy = 52 CutFocus = 63, LPCEnergy = 78, LPC Fokus = 61 bei 10-facher Vergrößerung.
HINWEIS: Schneidfokus und Energie müssen für verschiedene Dias, verschiedene Gewebe und erfasste Flächen angepasst werden, aber allgemeine Regeln sind, dass die Katapultleistung höher ist als die Schneidleistung und der Laser für das Katapultieren defokussiert werden muss. Je höher die Vergrößerung der Objektivlinse, desto geringer der Fokus des Lasers und desto höher die Energie. - Verwenden Sie die Zeichnungswerkzeuge, um Zellen auszuwählen, indem Sie den Interessenbereich umreißen.
- Wählen Sie die RoboLPC-Funktion aus der Funktionssymbolleiste aus, um Zellen basierend auf den optimierten Parametern in die Klebekappen zu katapultieren, die durch Schneiden auf einem schwarzen Segment oben erhalten werden.
HINWEIS: Die LCM-Parameter variieren zwischen Gewebetypen sowie Der Dicke des Abschnitts, der Gewebehärte und der Objektivlinsen. Daher ist es am besten, jeden Schlitten auf einer einfachen Membranfläche ohne Gewebeprobe zu optimieren, bevor die eigentliche Probe geschnitten wird. - Verwenden Sie die Flag-Werkzeuge, um Bereiche von Interesse zu markieren, um sie sofort zu finden, indem Sie dieses Flag aus der Elementliste auswählen.
- Überprüfen Sie mit der Schaltfläche "CapCheck", um die Klebekappe zu überprüfen, um zu bestätigen, dass Proben erfasst wurden. Typischerweise ist LCM von 10-15 Abschnitten (ca. 200 Zellen) pro Kappe für die RNA-Extraktion erforderlich.
- Halten Sie die gefangenen Proben auf Eis. Gehen Sie sofort für die RNA-Extraktion vor, um einen RNA-Abbau zu vermeiden.
HINWEIS: Einige LCM-Mikroskope sind mit fluoreszierendem Licht ausgestattet, das die Erfassung von Zellen ermöglicht, die mit Fluoreszenzmarkern gekennzeichnet sind.
7. RNA-Extraktion
- Verwenden Sie ein SET für die RNA-Extraktion nach LCM mit niedrigem Input (siehe Tabelle der Materialien). Solche Kits wurden entwickelt, um qualitativ hochwertige Gesamt-RNA konsequent aus weniger als zehn Zellen zurückzugewinnen.
- Isolieren Sie die gesamte RNA aus den erfassten Zelltypen gemäß den Anweisungen des Herstellers, einschließlich der on-column DNase-Behandlung.
HINWEIS: Der erste Schritt der RNA-Extraktion, bei dem die Röhre invertiert und geflickt wird, ist entscheidend, um sicherzustellen, dass die gefangenen Proben auf dem Deckel in Kontakt mit dem hinzugefügten Extraktionspuffer sind.
8. RNA-Verstärkung
- Verwenden Sie ein Antisense RNA (aRNA) Amplifikationskit (siehe Tabelle der Materialien) zur aRNA-Verstärkung aus der RNA, die durch In-vitro-Transkription extrahiert wird, um eine ausreichende aRNA für die RNA-Seq-Bibliothekssynthese zu erzeugen.
- Führen Sie zwei Verstärkerrunden mit dem aRNA-Verstärkungskit gemäß den Anweisungen des Herstellers durch.
HINWEIS: Es ist wichtig, den Thermocycler und Deckel auf die vom Hersteller des Kits angewiesene Temperatur (siehe Tabelle der Materialien)vorzuheizen. Ein alternativer Ansatz anstelle der RNA-Verstärkung besteht darin, ein Vorbereitungskit für die Bibliothek mit niedrigem Eingang zu verwenden, um die Bibliothek direkt aus extrahierter RNA zu synthetisieren.
9. RNA-Quantifizierung
- Quantifizieren und qualifizieren Sie aRNA mit einem automatisierten Elektrophoresesystem (siehe Tabelle der Materialien).
HINWEIS: Ein automatisiertes Elektrophoresesystem wird bevorzugt, da es weniger Proben (1-2 l) benötigt und ein gelähnliches Bild und Elektropherogramm für jede einzelne Probe liefert.
10. qRT-PCR und/oder RNA-seq
- Synthetisieren Sie cDNA aus der aRNA für qRT-PCR oder um RNA-seq-Bibliotheken mit Standard-RNA-Seq-Bibliothekskits zu erstellen.
Ergebnisse
Mit unserem LCM RNA-seq-Protokoll10haben wir räumliche und zeitliche gewebespezifische Transkriptome aus Gerstensamen während der Keimung erzeugt. Die Studie wurde durchgeführt, indem LCM RNA-seq auf eine kleine Anzahl von Zellen aus drei Embryoorganen (Plumule, Radicle Tip, Scutellum) alle 8 h über einen 48 h-Zeitverlauf während der Keimung (0-48 h, 7 Zeitpunkte) angewendet wurde (Abbildung 2A,B).
Diskussion
Viele gewebespezifische Genexpressionsstudien wurden durch die Handsektion von Proben begrenzt, was zeitaufwändig, arbeitsintensiv ist, ein hohes Kontaminationsrisiko hat und nur Proben verwenden kann, die ein menschlicher Aktivist ausreichend geschickt ernten kann. LCM ist eine präzise und berührungslose Technik, um Zellen aus festen Gewebeabschnitten mithilfe eines mechanisch betriebenen Laserstrahls unter mikroskopischer Visualisierung zu sammeln.
Eine gute Probenvorbereitung ist für LC...
Offenlegungen
Die Autoren haben nichts zu verraten.
Danksagungen
Diese Arbeit wurde vom Australian Research Council Centre of Excellence in Plant Energy Biology (CE140100008) an JW unterstützt. M.G.L wurde von einem Startstipendium der La Trobe University unterstützt. Wir danken der La Trobe Genomics Platform für ihre Unterstützung bei der Sequenzierung und Datenanalyse mit hohem Durchsatz. Wir danken Associate Professor Matthew Tucker für die fachkundige Beratung bei der Einrichtung von LCM in unserem Labor.
Materialien
| Name | Company | Catalog Number | Comments |
| Acetic acid 100 % ACS/R. | AnalaR NORMAPUR (BioStrategies) | VWRC20104.323 | |
| AdhesiveCap 200 opaque | Zeiss | 415190-9181-000 | |
| Clear base moulds 8 X 10 | Leica | 3803015 | |
| Diethyl pyrocarbonate | Sigma-Aldrich | 40718-25ML | |
| High Sensitivity RNA ScreenTape | Agilent | 5067-5579 | |
| Lowprofile disp.blades DB80LS | Leica | 14035843489 | |
| MembraneSlide 1.0 PEN | Zeiss | 415190-9041-000 | |
| MessageAmp II aRNA Amplification Kit | Ambion (ThermoFisher) | AMB17515 | |
| On-Column DNase I Digestion Set | Sigma-Aldrich | DNASE70 | |
| Ovation RNA-Seq System V2 | NuGen (Integrated Science) | 7102-08 | |
| Paraffin (Surgipath Paraplast) | Leica | 39601006 | |
| PicoPure RNA Isolation Kit | ABI (ThermoFisher) | KIT0214 | |
| RNaseZap RNase Decontamination Solution | Ambion (ThermoFisher) | AM9780 | |
| Xylene | AnalaR NORMAPUR (BioStrategies) | VWRC28975.360 | |
| Leica Benchtop Tissue Processor | Leica Biosystems | TP1020 | |
| Leica Heated Paraffin Embedding Module | Leica Biosystems | EG1150H | |
| Leica Cold Plate | Leica Biosystems | EG1150C | |
| Safemate Class 2 Biological Safety Cabinets | LAF Technologies Pty Ltd | Safemate 1.5 | |
| Leica Fully Automated Rotary Microtome | Leica Biosystems | RM2265 | with PALMRobo v 4.6 software |
| Zeiss PALM MicroBeam LCM system | Zeiss miscroscopy | ||
| TapeStation | Agilent | TapeStation 2200 |
Referenzen
- Emmert-Buck, M. R., et al. Laser capture microdissection. Science. 274 (5289), 998-1001 (1996).
- Alevizos, I., et al. Oral cancer in vivo gene expression profiling assisted by laser capture microdissection and microarray analysis. Oncogene. 20 (43), 6196-6204 (2001).
- Cong, P., et al. In situ localization of follicular lymphoma: description and analysis by laser capture microdissection. Blood, The Journal of the American Society of Hematology. 99 (9), 3376-3382 (2002).
- Blokhina, O., et al. Laser capture microdissection protocol for xylem tissues of woody plants. Frontiers in Plant Science. 7, 1965 (2017).
- Casson, S., Spencer, M., Walker, K., Lindsey, K. Laser capture microdissection for the analysis of gene expression during embryogenesis of Arabidopsis. The Plant Journal. 42 (1), 111-123 (2005).
- Chen, Z., et al. LCM-seq reveals the crucial role of LsSOC1 in heat-promoted bolting of lettuce (Lactuca sativa L.). The Plant Journal. 95 (3), 516-528 (2018).
- Jiao, Y., et al. A transcriptome atlas of rice cell types uncovers cellular, functional and developmental hierarchies. Nature Genetics. 41 (2), 258-263 (2009).
- Kivivirta, K., et al. A protocol for laser microdissection (LMD) followed by transcriptome analysis of plant reproductive tissue in phylogenetically distant angiosperms. Plant Methods. 15 (1), 1-11 (2019).
- Li, P., et al. The developmental dynamics of the maize leaf transcriptome. Nature Genetics. 42 (12), 1060-1067 (2010).
- Liew, L. C., et al. Temporal tissue-specific regulation of transcriptomes during barley (Hordeum vulgare) seed germination. The Plant Journal. 101 (3), 700-715 (2020).
- Matas, A. J., et al. Tissue-and cell-type specific transcriptome profiling of expanding tomato fruit provides insights into metabolic and regulatory specialization and cuticle formation. The Plant Cell. 23 (11), 3893-3910 (2011).
- Sakai, K., et al. Combining laser-assisted microdissection (LAM) and RNA-seq allows to perform a comprehensive transcriptomic analysis of epidermal cells of Arabidopsis embryo. Plant Methods. 14 (1), 10 (2018).
- Zhan, J., et al. RNA Sequencing of Laser-Capture Microdissected compartments of the maize kernel identifies regulatory modules associated with endosperm cell differentiation. The Plant Cell. 27 (3), 513-531 (2015).
- Hwang, B., Lee, J. H., Bang, D. Single-cell RNA sequencing technologies and bioinformatics pipelines. Experimental & Molecular Medicine. 50 (8), 1-14 (2018).
- Zeb, Q., Wang, C., Shafiq, S., Liu, L. . Single-Cell Omics. , 101-135 (2019).
- Deal, R. B., Henikoff, S. The INTACT method for cell type-specific gene expression and chromatin profiling in Arabidopsis thaliana. Nature Protocols. 6 (1), 56 (2011).
- Heiman, M., Kulicke, R., Fenster, R. J., Greengard, P., Heintz, N. Cell type-specific mRNA purification by translating ribosome affinity purification (TRAP). Nature Protocols. 9 (6), 1282 (2014).
- Bevilacqua, C., Ducos, B. Laser microdissection: A powerful tool for genomics at cell level. Molecular Aspects of Medicine. 59, 5-27 (2018).
- Nelson, T., Tausta, S. L., Gandotra, N., Liu, T. Laser microdissection of plant tissue: what you see is what you get. Annual Reviews in Plant Biology. 57, 181-201 (2006).
- Day, R. C., Grossniklaus, U., Macknight, R. C. Be more specific! Laser-assisted microdissection of plant cells. Trends in Plant Science. 10 (8), 397-406 (2005).
- Takahashi, H., et al. A method for obtaining high quality RNA from paraffin sections of plant tissues by laser microdissection. Journal of Plant Research. 123 (6), 807-813 (2010).
- Schroeder, A., et al. The RIN: an RNA integrity number for assigning integrity values to RNA measurements. BMC Molecular Biology. 7 (1), 3 (2006).
- Ferreira, E. N., et al. Linear mRNA amplification approach for RNAseq from limited amount of RNA. Gene. 564 (2), 220-227 (2015).
- Schneider, J., et al. Systematic analysis of T7 RNA polymerase based in vitro linear RNA amplification for use in microarray experiments. BMC Genomics. 5 (1), 29 (2004).
- Shanker, S., et al. Evaluation of commercially available RNA amplification kits for RNA sequencing using very low input amounts of total RNA. Journal of Biomolecular Techniques. 26 (1), 4 (2015).
- Bhattacherjee, V., et al. Laser capture microdissection of fluorescently labeled embryonic cranial neural crest cells. Genesis. 39 (1), 58-64 (2004).
- Clément-Ziza, M., Munnich, A., Lyonnet, S., Jaubert, F., Besmond, C. Stabilization of RNA during laser capture microdissection by performing experiments under argon atmosphere or using ethanol as a solvent in staining solutions. RNA. 14 (12), 2698-2704 (2008).
- Blokhina, O., et al. Parenchymal Cells Contribute to Lignification of Tracheids in Developing Xylem of Norway Spruce. Plant Physiology. 181 (4), 1552-1572 (2019).
- Schad, M., Lipton, M. S., Giavalisco, P., Smith, R. D., Kehr, J. Evaluation of two-dimensional electrophoresis and liquid chromatography-tandem mass spectrometry for tissue-specific protein profiling of laser-microdissected plant samples. Electrophoresis. 26 (14), 2729-2738 (2005).
- Schad, M., Mungur, R., Fiehn, O., Kehr, J. Metabolic profiling of laser microdissected vascular bundles of Arabidopsis thaliana. Plant Methods. 1 (1), 2 (2005).
- Latrasse, D., et al. The quest for epigenetic regulation underlying unisexual flower development in Cucumis melo. Epigenetics & Chromatin. 10 (1), 22 (2017).
- Turco, G. M., et al. DNA methylation and gene expression regulation associated with vascularization in Sorghum bicolor. The New Phytologist. 214 (3), 1213-1229 (2017).
- Gomez, S. K., Harrison, M. J. Laser microdissection and its application to analyze gene expression in arbuscular mycorrhizal symbiosis. Pest Management Science: Formerly Pesticide Science. 65 (5), 504-511 (2009).
- Roux, B., et al. An integrated analysis of plant and bacterial gene expression in symbiotic root nodules using laser-capture microdissection coupled to RNA sequencing. The Plant Journal. 77 (6), 817-837 (2014).
- Tang, W., Coughlan, S., Crane, E., Beatty, M., Duvick, J. The application of laser microdissection to in planta gene expression profiling of the maize anthracnose stalk rot fungus Colletotrichum graminicola. Molecular Plant-Microbe Interactions. 19 (11), 1240-1250 (2006).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenWeitere Artikel entdecken
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten