
Method Article
Ex Vivo Imaging de Postnatal cerebelosa gránulo migración celular utilizando confocal Macroscopía
En este artículo
Resumen
During postnatal cerebellum development, immature granule cells originating from the germinal zone exhibit distinct modalities of migration to reach their final destination and to establish neuronal networks. This protocol describes the preparation of cerebellar slices and the confocal macroscopic approach used to investigate the factors that regulate neuronal migration.
Resumen
Durante el desarrollo postnatal, las células granulares inmaduros (interneuronas excitatorias) exhiben la migración tangencial en la capa granular externa, y la migración luego radial en la capa molecular y la capa de células de Purkinje para llegar a la capa granular interna de la corteza cerebelosa. Por defecto en los procesos migratorios induce la muerte celular, ya sea o extravío de las neuronas, lo que lleva a los déficits en diversas funciones del cerebelo. La migración de células granulares centrípeta implica varios mecanismos, tales como la quimiotaxis y la degradación de la matriz extracelular, para guiar a las células hacia su posición final, pero los factores que regulan la migración de células en cada capa cortical son sólo parcialmente conocida. En nuestro método, rebanadas cerebelosa aguda, se preparan a partir P10 ratas, las células granulares están etiquetados con un marcador fluorescente citoplásmica y los tejidos se cultivan en insertos de membrana de 4 a 10 hr antes de iniciar la monitorización en tiempo real de la migración celular por macroscopía confocal a 37 ° C en elpresencia de CO 2. Durante su migración en las diferentes capas corticales del cerebelo, células granulares pueden estar expuestos a los agonistas o antagonistas de neuropéptido, inhibidores de proteasa, bloqueadores de los efectores intracelulares o sustancias incluso tóxicos tales como alcohol o metilmercurio para investigar su posible papel en la regulación de la migración neuronal .
Introducción
En el cerebelo en desarrollo, ocho tipos diferentes de neuronas se producen secuencialmente entre la segunda semana embrionario y la segunda semana postnatal en los roedores 1. Originario inicialmente a partir de, una zona germinal primaria, las células granulares inmaduros (GC) son los últimos neuronas para ser producidos a partir de la capa externa granular (EGL), una zona germinal secundaria 2. Durante las tres primeras semanas después del parto, la corteza cerebelosa es una estructura foliada organizado en cuatro capas que incluyen la EGL, la capa molecular (ML), la capa de células de Purkinje (PCL) y la capa granular interna (IGL) (Figura 1). A través de la migración centrípeta, inmadura GC, interneuronas glutamatérgicas, alcanzar el IGL dentro de aproximadamente 2 días. Por la tercera semana después del parto, la EGL desaparece y el IGL constituye lo que se llama la capa granular (GL) en el cerebelo adulto. En el GL, GC recibir entradas sinápticas excitatorias de fibras musgosas y células cepillo unipolares, yentradas sinápticas inhibitorias de axones de las células de Golgi. En el ML, los axones GC hacen sinapsis excitatorias con las neuronas GABAérgicas incluyendo las células de Purkinje, células cesta, las células estrelladas y las células de Golgi 2.
Observación en tiempo real del movimiento de las células en rodajas cerebelosa aguda, obtenidos a partir de roedores postnatales tempranas demuestra que GCs modifican su forma concomitante con los cambios en la modalidad y la velocidad de migración durante su ruta en la corteza cerebelosa 3. Durante las dos primeras semanas después del parto, precursores GC proliferan activamente en la parte superior de la EGL. En la parte media de la EGL, postmitotic GCs migran tangencialmente en la dirección de su proceso más grande. En la frontera EGL-ML, GC ralentizar su movimiento, las células comienzan a entrar en un proceso descendente vertical corta en el ML. En el ML, GC tiene un cuerpo celular alargado verticalmente, un proceso posterior delgado y un proceso que lleve más voluminoso, y migran radialmente a lo largo de las fibras gliales Bergmann. En elPCL, GC detener su movimiento, pero después de una fase estacionaria prolongado (2 horas), que cruzan la frontera PCL-IGL. En la IGL, GCs migran hacia la parte inferior de la capa en la ausencia de soporte de fibra glial. Una vez que las puntas del proceso que conduce acercarse a la frontera importa IGL-blanca (WM), GC lenta y detener su movimiento. Se prefieren las secciones transversales del cerebelo para los estudios de migración tangencial en el EGL mientras rebanadas sagital se dedican a la migración radial en el ML, PCL y IGL. Algunos factores de regulación de los movimientos de GC incluidos los neuropéptidos (por ejemplo, la somatostatina, PACAP) han sido identificados hasta el momento, pero los mecanismos completos implicados en el control espacio-temporal de la migración GC en cada capa cortical siguen siendo 1,4,5,6 gran parte desconocido.
Migración GC se ha estudiado durante los últimos 20 años a través de video y microscopía confocal utilizando iluminación de luz transmitida ya sea por células cultivadas aisladas o detección de fluorescencia para acurebanadas del cerebelo TE. DiI Inicialmente lipófilos, y más recientemente colorantes "Rastreo de la célula" y proteínas fluorescentes de células-expresado se utilizaron para la microscopía confocal o de dos fotones 7,8. Experimentos exitosos dependen de una serie de procedimientos específicos que hacen que el protocolo simple, pero no fácil. En particular, las rebanadas agudas tienen que ser estabilizado durante las observaciones en general, con una red de malla de nylon casera 9. La intensidad de iluminación de luz tiene que ser lo más bajo posible para evitar la fototoxicidad y photobleaching tal como se propone en los escaneos multipunto enfoque microscopio confocal. Además, la temperatura y CO 2 son parámetros ambientales clave desde la inestabilidad puedan afectar a la migración neuronal. Para facilitar y perfeccionar los procedimientos experimentales, hemos desarrollado un protocolo macroscopía confocal que limita los movimientos rebanada, asegura parámetros ambientales constantes, reduce fotoblanqueo, campo de visión aumenta (del orden de milímetros) y consequtemente el número de células (decenas) que pueden ser rastreados a través de análisis de imágenes. Por lo tanto, 180 micras rodajas gruesas se cultivan en insertos de membrana, y placas de 6 pocillos se transfieren directamente bajo un objetivo motorizado 2X de un macroscopio confocal comercial equipado con una gran cámara de incubación, la temperatura y el CO 2 controladores y un sistema de control de vibraciones. Time-lapsos y z-pilas se realizaron a continuación durante varias horas y herramientas farmacológicas o moléculas bioactivas se pueden añadir o entregados en el medio de incubación. Este método también podría ser adaptado para estudiar la migración de los otros tipos de neuronas en el cerebelo o cerebro en diferentes etapas de desarrollo.
Protocolo
Animales (hombre o ratas Wistar hembras) nacieron y se criaron en un animalario acreditado (aprobación B.76-451-04), según el libro de francés para el cuidado y uso de animales de laboratorio. Los experimentos se llevaron a cabo bajo la supervisión de investigadores autorizados (MB, DV y LG), de conformidad con la Directiva Europea Consejo de la Comunidad (2010/63 / UE de 22 de septiembre de 2010) y el Ministerio de Agricultura francés.
1. Preparación de Medios y Herramientas
- En una cabina de seguridad biológica, preparar BSS de Hank 1x (HBSS) en agua estéril de 10x solución madre que contiene CaCl2 (1,85 g / l) / MgSO4 (0,9767 g / l), sin MgCl2. Añadir NaHCO3 (350 g / ml) a 1x solución de HBSS.
- En una cabina de seguridad biológica, añadir suplemento N2 (de una solución madre 100x) y penicilina (100 unidades / ml) -streptomycin (0,1 mg / ml) solución a medio Eagle modificado (DMEM) mezcla de nutrientes de Dulbeccotura F-12 (1: 1).
- En condiciones estériles, preparar una alícuota (25 l, 2 mM) de la citoplásmico colorante fluorescente llamado Cell Rastreador verde en DMSO (1,075 ml para 1 mg). Diluir una alícuota en 5 ml de DMEM en un tubo cónico de 15 ml.
- Preparar lleno cubo de hielo para mantener los medios de comunicación a 4 ° C.
- Descontaminar mesas de trabajo de laboratorio y herramientas con etanol al 70%.
2. Disección de cerebelo de ratas P10
- Rápidamente decapitar a crías de rata (P10) con tijeras curvas de funcionamiento detrás de las orejas con el fin de obtener el comienzo de la médula espinal.
- En la parte posterior del lado de la cabeza decapitada, que la línea media de la incisión de la piel desde el cuello hasta la nariz con iris tijeras finas y se separa la piel del cráneo con iris finas tijeras y Dumont # 3 fórceps.
- Con unas tijeras finas iris para hacer delicadamente dos incisiones laterales desde la base hasta la región rostral del cráneo. Retire el cráneo disecado con dos # 3 fórceps. Separe el sujetadordesde cualquier adherencia con el cráneo con las mismas pinzas.
- Transferir el cerebro con el extremo cuchara de una espátula para placa de Petri (Ø 35 mm) que contiene 2 ml de medio HBSS enfriado con hielo.
- Coloque la placa de Petri que contiene el cerebro en un plato más grande Petri (Ø 100 mm), lleno de hielo y traslado a la etapa del microscopio estereoscópico.
- Bajo un microscopio estereoscópico, aislar el cerebelo del cerebro por dilaceración utilizando dos # 3 fórceps. Del mismo modo, eliminar la médula espinal residual y el pial-membrana.
- Transferir el cerebelo en una nueva placa de Petri llena de HBSS (Ø 35 mm, 2 ml) con el extremo cuchara de una espátula y mantener en hielo.
3. Preparación de rebanadas cerebelosa aguda
- Bajo el microscopio estereoscópico, cortar el cerebelo entre los vermis y el hemisferio derecho como se indica por las dos cabezas de flecha en la figura 2A con un estándar de mango de bisturí # 3 sólido y una cuchilla quirúrgica # 15.
- Ponga una gota de cianpegamento oacrylate en la platina vibratome y esperar 15-25 segundos para eliminar los vapores de solventes tóxicos.
- Recoger el cerebelo corte con el extremo cuchara de una espátula y retirar el exceso de HBSS con toallas de papel limpias.
- Traiga el cerebelo cerca del platina. Fijar el borde del corte de la platina y esperar 10 seg.
- Inserte el disco en la bandeja de muestra de tampón con el manipulador, y girarla de manera que el eje transversal del cerebelo es perpendicular al soporte de la cuchilla. Fijar la platina con una llave Allen y rellene suavemente el baño tampón con medio HBSS hasta el cerebelo está cubierto.
- Cargar hielo triturado en el baño de enfriamiento.
- Limpiar la cuchilla tres veces con etanol al 70% para eliminar cualquier aceite.
- Inserte la hoja en el soporte de la cuchilla y asegure con el tornillo de fijación.
- Coloque el filo de la cuchilla derecha detrás del borde posterior (a la vista del usuario) de la muestra y definirlo como un punto de partida. Utilice adelante comandar definir tque el punto final después de que el borde frontal de la muestra.
- Seleccione seccionar velocidad a 2,5 y seccionar frecuencia a 8. Seleccione el grosor de recorte a 180 micras. Iniciar seccionamiento de tejidos.
- Recoge cada sección usando un vaso de gran calibre truncado pipeta Pasteur y traslado al HBSS que contienen placa de Petri (Ø 35 mm) se mantuvieron en hielo.
- El uso de dos # 5 fórceps, eliminar las meninges cuidadosamente desde el cerebelo cuando interferir con la cuchilla. Reunir un máximo de 5 rebanadas por cerebelo (Figura 2B, C).
- Retire las meninges de las rebanadas del cerebelo cuidadosamente con dos # 5 pinzas bajo el microscopio estereoscópico y separar los lóbulos suavemente para una mejor carga de la sonda.
4. fluorescente tinción de interneuronas de Vida
- Traslado rebanadas del cerebelo con una pipeta Pasteur de vidrio de gran calibre truncado a una placa de 6 pocillos (máx 3 rebanadas / por pocillo). Aspirar el medio HBSS.
- Incubar las rebanadas (3 como máximo) en 5 ml de loading solución del colorante fluorescente (10 M).
- Para protegerlo de la luz, cubrir la microplaca con papel de aluminio. Póngalo en una mesa de giro-en movimiento a 35 rpm durante 10 minutos a temperatura ambiente para facilitar el etiquetado celular.
- Rodajas de transferencia sobre la membrana de un Transwell Insert (3,0 micras de tamaño de poro; Figura 2D) con una pipeta Pasteur de vidrio de gran calibre truncado. Aspirar el medio de carga con pipeta.
- Retire el inserto y llenar el pozo con 1,9 ml de DMEM. Reemplazar el inserto y añadir 100 l de DMEM en la parte superior de la rebanada para cubrir el tejido.
- Coloque la placa que contiene los insertos de cultivo en la cámara de la incubadora (37 ° C, 5% de CO 2) durante 2 horas, que es suficiente para observar los GC en el ML. Acuéstese tejidos planos para permitir la unión en la membrana de inserción (Figura 2E). Asegúrese de que las rebanadas no están secando.
5. Ex vivo Imaging través confocal Macroscopía
- Transferir el plato sinla tapa de plástico en una incubadora unido al soporte de un macroscopio confocal. Coloque una cubierta de vidrio en la placa de inserción del macroscopio. Mantenga la temperatura de la cámara de 37.0 ° C ± 0,5 ° C, y suministramos las rebanadas con un flujo constante de gas (95% O 2, 5% de CO 2) a través de la inserción de la placa para mantener el pH constante. Espere 2 horas más antes de experimento de lapso de tiempo.
- Para visualizar la distancia de migración de GC en los cortes de tejido, iluminar la preparación con una luz de longitud de onda de 488 nm por medio de un diodo láser a través de un macroscopio láser confocal de barrido equipado con un objetivo seco X2 (de trabajo: 39 mm, diámetro: 58 mm, NA = 0,234), y detectar la emisión de fluorescencia desde 500 hasta 530 nm.
- Para resolver finamente el movimiento de los GC, adquirir imágenes con un factor de zoom óptico adicional de 1,5 a 2,0. Recoge imágenes de BN en un solo plano focal o hasta 10 planos focales diferentes a lo largo del eje z cada 30 min durante un máximo de 12 horas.
- Cuando es necesario, retire la cubierta de cristal y añadir pequeños volúmenes (1-10 mu l) de activadores biológicos o inhibidores en DMEM con una pipeta de 10 l para estudiar su efecto sobre la migración de GC.
Seguimiento 6. celular
- Para cada momento de la película, realice la proyección z-pila a través del modo de tipo ecart en ImageJ. Modular el contraste y los niveles de brillo de las imágenes sucesivas para facilitar la identificación y el seguimiento de los GC etiquetados. Mapa manualmente cada posición en la instantánea de referencia (en t = 0).
- Utilice el plugin "seguimiento manual" en el menú de partículas Analizar y determinar haciendo clic en el punto de cada cuerpo celular gravedad durante lapsos de tiempo. Exportar los datos de seguimiento de primas en una hoja de cálculo.
- Reorganizar los datos de seguimiento primas exportadas desde ImageJ con un programa inteligente de fabricación casera (http://primacen.fr, escrito en código PHP) que identifican a cada célula y cargos asociados. Usando el programa, calculate la distancia total recorrida y la velocidad media de la migración para cada celda. Clasificar y comparar las características de la migración celular en condiciones de control y de tratamiento bajo filtros adecuados utilizando el mismo programa.
Resultados
A principios del cerebelo posnatal, GC presentan cambios significativos en su modo y la velocidad de la migración, ya que se cruzan diferentes capas corticales 1 (Figura 1). Esta sección ilustra ejemplos de los resultados que se pueden obtener mediante el estudio de la migración GC en su medio celular natural. Rata P10 cortes de tejido cerebeloso marcados con un colorante fluorescente verde se examinan bajo un macroscopio confocal (Figura 3A) y nos muestran que los GC migrar radialmente en la ML con una velocidad media de 18 m / h (Figura 3B, C). Hasta la fecha, el papel de las interacciones / comunicaciones entre las células neuronales y gliales incluyendo los factores de regulación y mecanismos moleculares implicados en el control de la migración de células en cada capa cortical son en gran parte desconocidos. En consecuencia, la cuestión principal es identificar los neuropéptidos, neurotransmisores, las neurotrofinas y los componentes de la matriz extracelular que podrían desempeñar un papel en estos cortical capa específicacambios fic de la velocidad durante su proceso de migración. Polipéptido activador de la adenilato ciclasa pituitaria (PACAP) se detecta principalmente en el PCL, sino también en el ML y el IGL durante las dos primeras semanas después del parto en roedores 7,10,11. Aplicación de PACAP38 (10 -6 M) al medio de cultivo resultó en una disminución de velocidad 79% de la GC en el ML. Por ejemplo, la velocidad de migración de GCs en el ML se redujo de 11,9 m / h en condiciones de control a 2,5 micras / hr después de la administración de PACAP38 (Figura 4A). Activador del plasminógeno de tipo tisular (tPA) es un miembro de la cascada proteolítica que conduce a la degradación de los componentes de la matriz extracelular (EM) tales como moléculas de adhesión celular o laminina 12,13. tPA y del plasminógeno, un sustrato de tPA, se detectan en las capas corticales durante el desarrollo del cerebelo postnatal 14,15,16. La administración de PAI-1 (10 -7 M), un inhibidor endógeno de tPA, reduce en 78% la GCmigración en el ML. Por ejemplo, GCs reduce la velocidad de migración en el ML de 19,2 m / h en condiciones de control a 4,2 micras / hr después de la adición de PAI-1 (Figura 4B). Estos resultados indican que PACAP ejerce un efecto inhibidor directo sobre los movimientos de GC y que la serina proteasa tPA facilita la migración de GCs en el ML del cerebelo de rata en desarrollo.

Figura 1: Representación 3D de la migración de GC en la corteza cerebelosa postnatal. 1-4, extensión de los procesos de GC y la migración tangencial en el EGL 5., La migración radial en el ML largo de las fibras gliales Bergmann. 6, fase estacionaria transitoria en el PCL. 7, la migración glial-independiente radial en el IGL. 8, Finalización de GC migración en el IGL GC, de células granulares, en rojo;. EGL, Capa granular externa; B, Bergmann células gliales, de color morado oscuro; G, células de Golgi, en amarillo; cf, fibras de escalada, en azul; g, células granulares postmigratory, en luz verde; IGL, capa granular interna; MFT, fibra de musgo terminal, en verde oscuro; ML, capa molecular; P, de células de Purkinje, en morado claro; capa de células PCL, Purkinje. Esta cifra ha sido modificado desde 5.

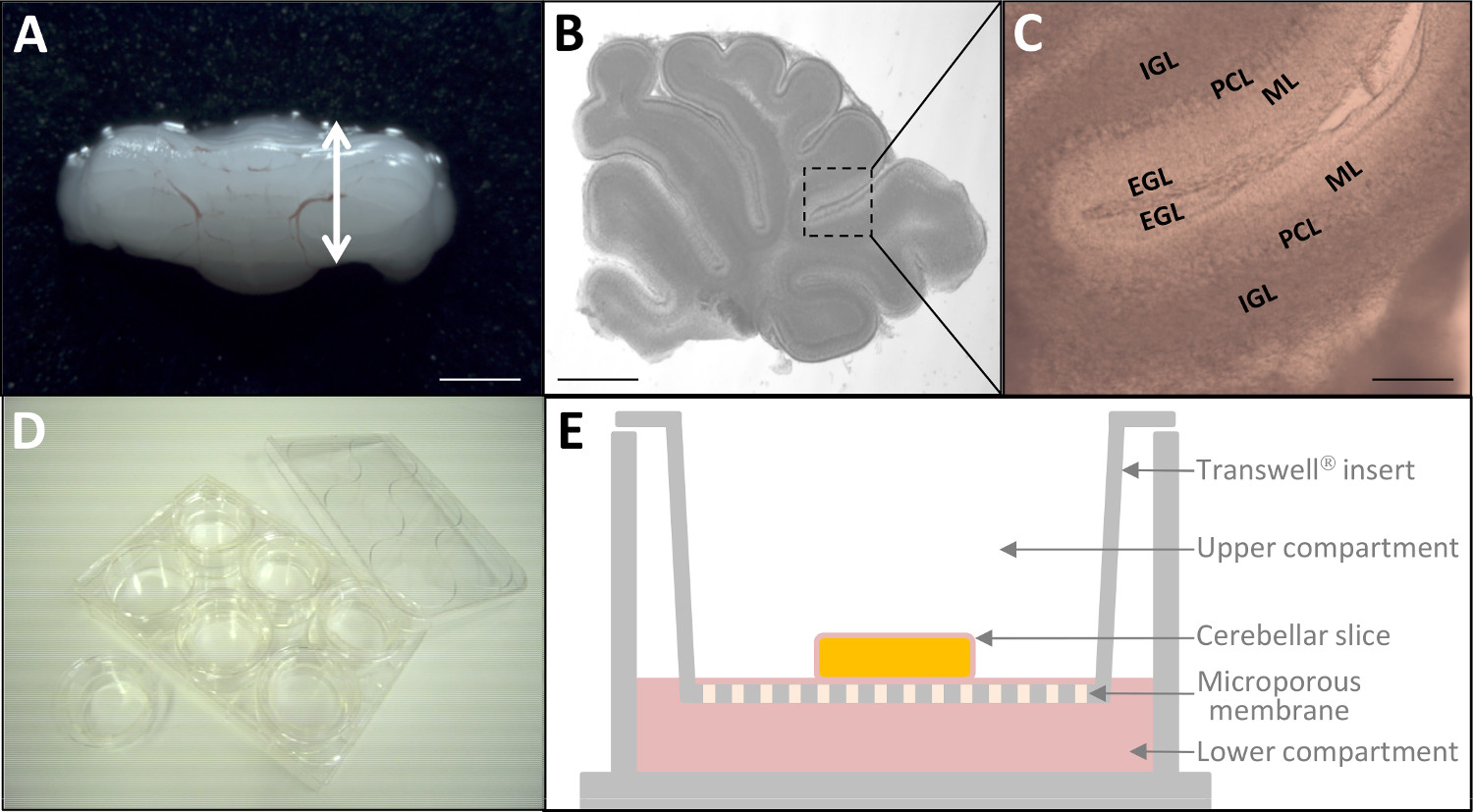
Figura 2: la cultura ex vivo de las rebanadas del cerebelo P10 (A) cerebelo disecado de rata P10.. La barra de escala = 6 mm. (B) Micrografía de vivir 180 micras de espesor rebanada cerebelosa a través de microscopía estereoscópica. La barra de escala = 3 mm. (Gatoun aumento mayor, las cuatro capas corticales (EGL, ML, PCL, IGL) del cerebelo ya son distinguibles. La barra de escala = 1 mm. (D) Después del marcaje fluorescente, cortes de tejido se colocan en insertos de cultivo (24 mm de diámetro) en una placa de 6 pocillos. (E) Representación esquemática de un inserto de la cultura con la membrana de poliéster de cultivo de tejidos tratados. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: la migración dinámica de GC en las capas corticales del cerebelo P10 rata (A) vista Macroconfocal (xyz, proyección 2D) de una rebanada del cerebelo de rata P10 en el que los GC se marcan con un colorante fluorescente citoplásmica verde.. La barra de escala = 75 micras. (B) Time-lapse muestra GCmovimientos en la ML por macroscopía confocal de 4 horas en condiciones de control. Asterisco (*) símbolo marca el soma GC. El tiempo transcurrido (en min) se indica en la parte inferior de cada fotomicrografía. La barra de escala = 10 micras. (C) los cambios secuenciales en la distancia recorrida por GC soma.

Figura 4:. Efecto de neuropéptido y proteasa inhibidor de la migración GC (A) GC fue rastreado en el ML por macroscopía confocal durante 2 horas en condiciones de control y luego durante 2 horas en presencia de polipéptido de adenilato-ciclasa pituitaria activación (PACAP). (B) GC fue localizado en el ML by macroscopía confocal durante 2 horas en condiciones de control y luego durante 2 horas en presencia del inhibidor del activador del plasminógeno-1 (PAI-1).
Discusión
Este protocolo describe la cultura de rata P10 rebanadas cerebelosa aguda en el sistema Transwell y el marcaje fluorescente de GC con un colorante fluorescente verde para estudiar la migración celular durante el desarrollo postnatal a través de macroscopía confocal. Este protocolo permite observaciones de la migración de células durante un periodo de hasta 12 horas y la prueba de las posibles funciones de los factores en la migración, incluyendo agonistas o antagonistas de los neuropéptidos, inhibidores de enzimas, moduladores de señalización celular o sustancias tóxicas durante el experimento regular. Un pequeño orificio en el inserto de membrana con una punta de pipeta es necesario para facilitar la administración de los compuestos en el medio de incubación. Una pipeta de punta curvada hecha en casa se puede utilizar para facilitar la entrega de la solución.
Una cuestión de los estudios de migración celular en cortes de tejido vivo es que los movimientos del propio tejido pueden hacer difícil el seguimiento de la célula. Considerando lo han propuesto enfoques anteriores para estabilizar suavemente slices con un nylon de malla o una fina capa de colágeno de cola de rata 7,17, una de las principales ventajas de esta tecnología es la transferencia simple y directa de una placa de cultivo de 6 pocillos que contiene las rebanadas del cerebelo en insertos de membrana de la incubadora de CO 2 en el marco del objetivo de un macroscopio confocal. Controladores de temperatura integrado y CO 2 también proporcionan los parámetros ambientales adecuadas y constantes esenciales para la migración celular 9. Por lo tanto, las condiciones de cultivo se mantienen durante las observaciones y los movimientos de tejido se reducen al mínimo ya que las rebanadas están bien unidos a la pieza de inserción de membrana. Estabilización del tejido se verifica siguiendo la posición de los bordes rebanada o células de Purkinje que deben ser corregidos referencias durante la adquisición. Además, las rebanadas del cerebelo (entre 12 y 18) distribuidas en los 6 pocillos de la placa se pueden observar rápidamente en detalle con una platina motorizada y un zoom óptico. Debido a la gran distancia de trabajo (X2, 39 mm) del objetivo seco, epi-observación está libre de inmersión y la administración de compuestos en el medio de cultivo es mucho más fácil. Por lo tanto, los parámetros ambientales y similitudes de apoyo en la cultura de CO 2 incubadora y confocal macroscopía conducen a la máxima conservación de la muestra biológica.
Otra ventaja del protocolo es el gran campo de visión y por consiguiente el gran número de células que se pueden observar de forma simultánea. Por ejemplo, se ha determinado previamente que la densidad de los GC fluorescentes con la migración radial en la ML fue 1.124 ± 138 células / mm 2 18. Macroscopía confocal (X2, NA = 0,234) tiene una resolución lateral inferior en comparación con la microscopía confocal (40X, NA = 1,25), pero la célula cuerpos de los GC pueden ser rastreados con facilidad y la velocidad media de la migración es comparable entre los dos enfoques tecnológicos 7,18 .
Además de la mejora técnica de las adquisiciones de imágenes, la calidad de los cortes de tejido y XXe calidad de etiquetado son puntos clave para los experimentos exitosos. Siempre mantenga los medios de comunicación y los tejidos en hielo durante los procesos de disección, eliminar aceite en vibratome cuchillas y no utilizan rebanadas de tejido en contacto con pegamento. Secciones sagitales y transversales se adaptan a radial y la migración tangencial respectivamente. Utilizar diferentes longitudes de incubación para la detección apropiada en las diferentes capas corticales del cerebelo. Tiempos de incubación largos (hasta 8 horas) son necesarios para detectar la migración de numerosos GC en el PCL y el IGL. Dado que la migración GC es un proceso fisiológico durante las ventanas de espacio-temporales específicos, el control positivo es que las células tienen que migrar correctamente. En particular, numerosos GC cabezal en el ML es uno de los indicadores de salud principal de rebanadas sagital del cerebelo. Para comenzar los experimentos, se sugiere la observación de los movimientos de GC en el ML. De hecho, la forma de GC con cuerpo celular alargado verticalmente debe ser considerado como un punto de referencia para comenzar acquisition con control de sucesivos (2 horas) y tratamiento (2 hr) períodos que se pueden realizar fácilmente en el ML.
Los colorantes fluorescentes como la familia Cell Tracker o proteínas fluorescentes expresados a través de construcciones genéticas pueden ser utilizados como trazadores para los estudios de migración celular. Debido a la lenta cinética de la migración GC (1 pila cada 30 minutos), los experimentos multicolor también se pueden realizar en el modo secuencial desde 4 lasers vigas (405, 488, 532 y 633 nm) están disponibles en el sistema. Teniendo en cuenta la migración radial centrípeta y centrífuga, el seguimiento de otras interneuronas también se pueden realizar 18. En particular, menos numerosos tipos de células pueden ser más fácilmente localizadas con un gran campo de visión. Finalmente, este protocolo se puede utilizar para estudiar la migración de células en otras etapas del desarrollo del cerebelo, sino también a otras áreas del cerebro.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Este trabajo fue apoyado por el Instituto para la Investigación y la Innovación en Biomedicina (IRIB), la Plataforma de imágenes de células de Normandía (PRIMACEN), Inserm, IBISA, la Universidad de Rouen, el Fondo Europeo de Desarrollo Regional (FEDER - Perené, Interreg 4A), la-Neurociencias LARC Red y la región de Haute-Normandie.
Materiales
| Name | Company | Catalog Number | Comments |
| Dulbecco’s modified eagle medium (DMEM) nutrient mixture F-12 | Sigma-Aldrich | D8437 | |
| Hank's balanced salt solution 10x | Sigma-Aldrich | H1641 | |
| PACAP38 | INRS, Canada | Bourgault et al., 200919 | |
| PAI-1 | Calbiochem | 528208 | |
| N-2 supplement | Fisher Scientific / Gibco/ invitrogen | O973 | |
| Cyanoacrylate glue | Loctite | ||
| Cell Tracker Green CMFDA | Invitrogen | C2925 | |
| Polyester Transwell-Clear inserts | Corning | 3452 | |
| Penicillin-Streptomycin | Sigma-Aldrich | P0781 | |
| 6-well cell culture cluster | Corning | 3516 | |
| DMSO | Fisher Scientific | BP231-100 | |
| Tissue culture dish 35 mm diameter | BD Falcon | 353004 | |
| Tissue culture dish 100 mm diameter | Thermo SCIENTIFIC | 130182 | |
| Polypropylen tube (15 ml) | BD Falcon | 352096 | |
| Ethanol 70% | Fisher Chemical | E/0800DF/21 | |
| Biological safety cabinet fume hood | Thermo Scientific | MSC9 Class II A2 | |
| Adjustable-volume pipette (0.5-10 µL) | Eppendorf | 4910 000.018 | |
| Gyro-rocker, SSL3 | Stuart | ||
| CO2 incubator, Hera Cell 150 | Thermo Scientific | ||
| Vibrating blade microtome, VT1000S | Leica Microsystems | ||
| Confocal macroscope, TCS LSI | Leica Microsystems | ||
| Temperature controller | PeCon | ||
| CO2-controller | PeCon | ||
| Stereomicroscope, M205 C | Leica Microsystems | ||
| Operating scissors, curved, blunt/blunt | Medicon | 03.03.17 | |
| Hardened fine iris scissors, straight, sharp/sharp | FST | 149090-11 | |
| Dumont #3 and #5 forceps | FST | 11293-00 and 11252-20 | |
| Vibratome injector blades/single edge | Leica Microsystems | 39053250 | |
| Standard scalpel handle #3 solid | FST | 10003-12 | |
| Surgical blade #15 | Swann-Morton | 205 |
Referencias
- Komuro, Y., Kumada, T., Ohno, N., Foote, K. D., Komuro, H. Migration in the Cerebellum. Cellular Migration and Formation of Neuronal Connections: Comprehensive Developmental Neuroscience. 2, 281-297 (2013).
- Altman, J., Bayer, S. A. . Development of the cerebellar system in relation to its evolution, structure, and functions. , (1997).
- Komuro, H., Rakic, P. Distinct modes of neuronal migration in different domains of developing cerebellar cortex. Journal of Neuroscience. 18 (4), 1478-1490 (1998).
- Komuro, H., Yacubova, E. Recent advances in cerebellar granule cell migration. Cellular and Molecular Life Sciences. 60 (6), 1084-1098 (2003).
- Fahrion, J. K., et al. Rescue of neuronal migration deficits in a mouse model of fetal Minamata disease by increasing neuronal Ca2+ spike frequency. Proceedings of National Academy of Sciences USA. 109 (13), 5057-5062 (2012).
- Raoult, E., et al. Pituitary adenylate cyclase-activating polypeptide (PACAP) stimulates the expression and the release of tissue plasminogen activator (tPA) in neuronal cells: involvement of tPA in the neuroprotective effect of PACAP. Journal of Neurochemistry. 119 (5), 10-1111 (2011).
- Cameron, D. B., et al. Cerebellar cortical-layer-specific control of neuronal migration by pituitary adenylate cyclase-activating polypeptide. Neuroscience. 146 (2), 697-712 (2007).
- Renaud, J., et al. Plexin-A2 and its ligand, Sema6A, control nucleus-centrosome coupling in migrating granule cells. Nature Neuroscience. 4, 440-449 (2008).
- Komuro, H., Rakic, P. Dynamics of granule cell migration: a confocal microscopic study in acute cerebellar slice preparations. Journal of Neuroscience. 15 (2), 1110-1120 (1995).
- Nielsen, H. S., Hannibal, J., Fahrenkrug, J. Expression of pituitary adenylate cyclase activating polypeptide (PACAP) in the postnatal and adult rat cerebellar cortex. Neuroreport. 9 (11), 2639-2642 (1998).
- Hannibal, J. Pituitary adenylate cyclase-activating peptide in the rat central nervous system: an immunohistochemical and in situ hybridization study. Journal of Comparative Neurology. 453 (4), 389-417 (2002).
- Garcia-Rocha, M., Avila, J., Armas-Portela, R. Tissue-type plasminogen activator (tPA) is the main plasminogen activator associated with isolated rat nerve growth cones. Neuroscience Letters. 180 (2), 123-126 (1994).
- Ware, J. H., DiBenedetto, A. J., Pittman, R. N. Localization of tissue plasminogen activator mRNA in the developing rat cerebellum and effects of inhibiting tissue plasminogen activator on granule cell migration. Journal of Neurobiology. 28 (1), 9-22 (1995).
- Friedman, G. C., Seeds, N. W. Tissue plasminogen activator mRNA expression in granule neurons coincides with their migration in the developing cerebellum. Journal of Comparative Neurology. 360 (4), 658-670 (1995).
- Seeds, N. W., Siconolfi, L. B., Haffke, S. P. Neuronal extracellular proteases facilitate cell migration, axonal growth, and pathfinding. Cell and Tissue Research. 290 (2), 367-370 (1997).
- Basham, M. E., Seeds, N. W. Plasminogen expression in the neonatal and adult mouse brain. Journal of Neurochemistry. 77 (1), 318-325 (2001).
- Bourgault, S., et al. Molecular and conformational determinants of pituitary adenylate cyclase-activating polypeptide (PACAP) for activation of the PAC1 receptor. Journal of Medical Chemistry. 52 (10), 3308-3316 (2009).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados