
Method Article
Ex Vivo הדמיה של נדידת תאי המוח הקטן אחרי הלידה גרגיר שימוש Confocal Macroscopy
In This Article
Summary
During postnatal cerebellum development, immature granule cells originating from the germinal zone exhibit distinct modalities of migration to reach their final destination and to establish neuronal networks. This protocol describes the preparation of cerebellar slices and the confocal macroscopic approach used to investigate the factors that regulate neuronal migration.
Abstract
במהלך התפתחות לאחר לידה, תאים לא בשלים גרגיר (interneurons מעוררת) תערוכה הגירה משיקה בשכבת גרגרים החיצונית, ולאחר מכן הגירת רדיאלי בשכבה המולקולרית ושכבת תאי פורקינג להגיע שכבת גרגרים הפנימית של קליפת המוח הקטנה. ברירת מחדל בתהליכים נודדים גורמת גם מוות של תאים או מיקום שגוי של תאי העצב, שהוביל לגירעונות בפונקציות המוח הקטן מגוונות. נדידת תאי גרגיר הצנטריפטלי כרוכה במספר מנגנונים, כגון chemotaxis והשפלה מטריקס, כדי להנחות את התאים למיקום הסופי שלהם, אבל הגורמים המווסתים את נדידת תאים בכל שכבת קליפת המוח ידועים רק באופן חלקי. בשיטה שלנו, פרוסות המוח הקטן חריפות ערוכות מP10 חולדות, תאי גרגיר מסומנים עם סמן cytoplasmic ניאון ורקמות בתרבית על קרום מוסיף 4-10 שעות לפני תחילת ניטור של נדידת תאים בזמן אמת על ידי macroscopy confocal על 37 מעלות צלזיוס בתוך הנוכחות של CO 2. במהלך נדידתם בשכבות בקליפת המוח השונות של המוח הקטן, יכולים להיות חשופים תאי גרגיר לאגוניסטים neuropeptide או יריבים, מעכבי פרוטאז, חוסמי של effectors תאיים או חומרים רעילים אפילו כמו אלכוהול או מתיל כספי לחקור תפקידם האפשרי ברגולציה של נדידת נוירונים .
Introduction
במוח הקטן הפיתוח, שמונה סוגים שונים של תאי עצב מיוצרים ברצף בין השבוע השני העוברי והשבוע לאחר הלידה השנייה במכרסמים 1. מקורו בתחילה מ, אזור נבטי עיקרי, תאים לא בשלים גרגיר (GC) הם נוירונים האחרונים שהופקו מהשכבה החיצונית הגרעינית (EGL), אזור נבטי משנית 2. במהלך שלושת השבועות לאחר הלידה הראשון, קליפת המוח הקטנה הוא מבנה foliated המאורגן בארבע שכבות כוללים EGL, השכבה המולקולרית (ML), שכבת פורקינג 'התא (PCL) והשכבה הפנימית גרגירים (IGL) (איור 1). באמצעות הגירה הצנטריפטלי, GCS לא בוגר, interneurons glutamatergic, להגיע IGL בתוך כ 2 ימים. בשבוע שלאחר הלידה השלישי, EGL נעלם וIGL מהווה מה שנקרא שכבת גרגרים (GL) במוח הקטן למבוגר. בGL, GCS לקבל תשומות הסינפטי מעוררות מסיבי אזובי ותאי מברשת חד קוטביים, ותשומות הסינפטי מעכבות מהאקסונים של תאי Golgi. בML, האקסונים GC לעשות סינפסות מעוררות עם נוירונים GABAergic כוללים תאי פורקינג ', תאי סל, תאים דמויים כוכב, ותאי Golgi 2.
תצפית בזמן אמת של תנועת תא במוח קטנה פרוסות חריפות המתקבלות ממכרסמים לאחר לידה מוקדמות מוכיחה כי GCS לשנות את צורתם במקביל לשינויים באופנות והמהירות של הגירה במסלולם במוח הקטנה הקליפה 3. במהלך שבועיים שלאחר לידה ראשון, מבשרי GC להתרבות באופן פעיל בחלק העליון של EGL. בחלק האמצעי של EGL, GCS postmitotic להעביר בעקיפין בכיוון של התהליך הגדול יותר שלהם. בגבול EGL-ML, GCS להאט את תנועתם, התאים מתחילים להיכנס לתהליך יורד אנכי קצר לML. בML, יש לי GCS גוף מוארך אנכי תא, תהליך נגרר דק ותהליך מוביל משופע יותר, ולהעביר רדיאלית לאורך סיבי גליה ברגמן. בתוך הPCL, GCS לעצור התנועה שלהם, אבל אחרי שלב ממושך נייח (2 שעות), שהם חוצים את גבול PCL-IGL. בIGL, GCS להעביר לתחתית השכבה בהעדר תמיכת סיבי גליה. ברגע שהעצות של התהליך המוביל להתקרב לגבול עניין IGL-לבן (WM), GCS איטי ולעצור את תנועתם. סעיפים רוחביים של המוח הקטן עדיפים ללימודי הגירה משיקים בEGL בעוד פרוסות sagittal מוקדשות לרדיאלי הגירה בML, PCL וIGL. כמה גורמי רגולציה של תנועות GC כוללים נוירו (למשל, סומטוסטטין, PACAP) זוהה עד כה, אך המנגנונים שלמים המעורבים בשליטת המרחב ובזמן של הגירת GC בכל שכבת קליפת המוח עדיין 1,4,5,6 במידה רבה לא ידועים.
הגירת GC נחקרה במהלך 20 השנים האחרונות באמצעות וידאו ומיקרוסקופיה confocal באמצעות תאורת אור או מועברת לתאים מבודדים בתרבית או גילוי הקרינה לACUפרוסות המוח הקטן te. DiI תחילה lipophilic, ולאחרונה צבעים "Tracker תא" וחלבוני ניאון הביע תא שמשו למיקרוסקופיה confocal או שני פוטונים 7,8. ניסויים מוצלחים תלויים במספר ההליכים הספציפיים שהופכים את הפרוטוקול פשוט אך לא קל. בפרט, פרוסות חריפות צריכה להיות התייצבו במהלך תצפיות בדרך כלל עם רשת רשת ניילון תוצרת בית 9. עוצמת הארת אור צריך להיות נמוך ככל האפשר כדי למנוע phototoxicity וphotobleaching כפי שהוצע על ידי גישת מיקרוסקופ confocal סריקה מרובות. בנוסף, טמפרטורה ו- CO 2 פרמטרים סביבתיים מרכזיים מאז חוסר יציבות עלולה להשפיע על נדידת נוירונים. כדי להקל ולשפר את הליכי הניסוי, פיתחנו פרוטוקול macroscopy confocal שמגביל תנועות פרוסה, מבטיח פרמטרים סביבתיים קבועים, מפחית photobleaching, מגביר שדה הראייה (על סדר milimeters) וconsequently מספר התאים (עשרות) שניתן לעקוב באמצעות ניתוח תמונה. לפיכך, 180 מיקרומטר פרוסות עבות בתרבית במוסיף קרום, ו6-גם צלחות מועברות ישירות תחת מטרה ממונעת 2X של macroscope confocal המסחרי מצוידים בדגירה גדולה קאמרית, מערכת בקרת רטט טמפרטורה ו- CO 2 בקרים ו. זמן מעידות וZ- ערימות מבוצעות לאחר מכן על פני כמה שעות וכלים תרופתיים או ניתן להוסיף מולקולות ביו או נמסרו במדיום הדגירה. שיטה זו יכולה גם להיות מותאמת ללמוד ההגירה של הסוגים של תאי עצב במוח הקטן או המוח בשלבי התפתחותיים שונים האחרים.
Protocol
בעלי חיים (זכר או נקבת חולדות Wistar) נולדו וגדל במתקן בעלי חיים מוכר (אישור B.76-451-04), על פי המדריך הצרפתי לטיפול ושימוש בחיות מעבדה. ניסויים שנערכו תחת פיקוחו של חוקרים מורשים (MB, DV ו- LG) בהתאם לאירופה מועצת קהילת ההוראה (2010/63 / UE 22 בספטמבר 2010) ומשרד החקלאות הצרפתי.
1. הכנת מדיה וכלים
- בארון בטיחות ביולוגי, להכין BSS של 1x האנק (HBSS) במים סטריליים מ10x פתרון מניות המכיל CaCl 2 (/ L 1.85 g) / MgSO 4 (.9767 גר '/ ליטר) ללא MgCl 2. להוסיף NaHCO 3 (350 מיקרוגרם / מיליליטר) ל1x פתרון של HBSS.
- בארון בטיחות ביולוגי, להוסיף תוספת N2 (מפתרון מניות 100x) ופניצילין (100 יחידות / מיליליטר) -streptomycin פתרון לבינוני נשר שונה (DMEM) התערובת המזינה של Dulbecco (מ"ג 0.1 / מיליליטר)ture F-12 (1: 1).
- בתנאים סטריליים, להכין aliquot (25 μl, 2 מ"מ) של תא cytoplasmic ניאון הצבע נקרא Tracker הירוק בDMSO (1.075 מיליליטר ל1 מ"ג). לדלל aliquot אחד ב 5 מיליליטר של DMEM בצינור חרוטי 15 מיליליטר.
- הכן קרח דלי מלא כדי לשמור על תקשורת ב 4 מעלות צלזיוס.
- לטהר צמרות מעבדה ספסל וכלים עם 70% אתנול.
2. Dissection של המוח קטן מחולדות P10
- במהירות לערוף גורי חולדה (P10) במספריים הפעלה מעוקלים מאחורי אוזניים כדי לקבל את תחילת חוט השדרה.
- בחלק האחורי בצד של הראש הערוף, לעשות חתך קו האמצע של העור מהצוואר לאף עם איריס מספריים הקנס ולהפריד את העור מהגולגולת עם איריס מספריים קנס ו# 3 מלקחיים דומון.
- השתמש במספרי איריס בסדר לעשות בעדינות שני חתכים רוחב מהבסיס באזור מקורי של הגולגולת. הסר את הגולגולת גזור עם שני # 3 מלקחיים. לנתק את החזייהמכל דבקות בגולגולת באמצעות אותו המלקחיים.
- העבר את המוח עם סוף הכפית של מרית לצלחת פטרי (מ"מ Ø 35) המכילה 2 מיליליטר של מדיום HBSS קר כקרח.
- שים את צלחת פטרי המכילה את המוח בצלחת פטרי גדולה יותר (O מ"מ 100) מלאה קרח וההעברה לשלב של סטראו.
- תחת סטראו, לבודד את המוח הקטן מהמוח על ידי dilaceration באמצעות שני # 3 מלקחיים. באופן דומה, להסיר את חוט השדרה השייר וpial-קרום.
- העבר את המוח הקטן בצלחת פטרי מלאה-HBSS חדשה (O 35 מ"מ, 2 מיליליטר) עם הכפית של סוף מרית ולשמור על קרח.
3. הכנת פרוסות המוח הקטן חריפות
- תחת סטראו, לחתוך את המוח הקטן בין vermis והאונה הימנית כפי שצוין על ידי שני ראשי החץ באיור 2 א עם ידית אזמל # סטנדרטי 3 מוצקים ולהב כירורגים # 15.
- שים טיפה אחת של ציאןדבק oacrylate על דיסק דגימת vibratome ולחכות 15-25 שניות לחסל אד ממס רעיל.
- לאסוף את המוח הקטן לחתוך עם סוף הכפית של מרית ולהסיר HBSS העודף במגבות נייר נקיות.
- להביא את המוח הקטן הקרוב לדיסק הדגימה. תקן את הקצה לחתוך לדיסק הדגימה ולחכות 10 שניות.
- הכנס את דיסק הדגימה למגש החיץ עם מניפולטור, ולסובב אותו כל כך הציר הרוחבי של המוח הקטן הוא בניצב לבעל הסכין. תקן את דיסק הדגימה עם מפתח אלן ולמלא בעדינות את מגש החיץ עם מדיום HBSS עד המוח הקטן מכוסה.
- טען קרח כתוש לאמבטיה הקירור.
- נקה את שלוש פעמים הלהב עם 70% אתנול לחסל את כל שמן.
- הכנס את הלהב לתוך מחזיק הסכין ובטוח עם הידוק בורג.
- הנח את זכות קצה להב מאחורי הקצה האחורי (מהתצוגה של המשתמש) של הדגימה ולהגדיר אותה כנקודת התחלה. השתמש קדימה הפקודה להגדיר tהוא נקודת הסיום לאחר הקצה הקדמי של הדגימה.
- בחר חתך מהירות 2.5 וחתך תדירות ב8. עובי זמירה בחר ב 180 מיקרומטר. התחל חתך רקמות.
- להרים כל קטע באמצעות זכוכית רחבה נשא מקוצצת פיפטה פסטר והעברה לHBSS המכיל צלחת פטרי (O 35 מ"מ) שמר על קרח.
- שימוש בשני מלקחיים 5 #, להסיר את קרומי המוח בזהירות מהמוח הקטן כאשר מפריע ללהב. לאסוף מקסימום של 5 פרוסות למוח קטן (איור 2, C).
- הסר את קרומי המוח מפרוסות המוח הקטן בזהירות עם שני # 5 מלקחיים תחת סטראו ולהפריד את האוניות בעדינות לטעינת בדיקה טובה יותר.
4. פלורסנט הכתמה של interneurons החיים
- העבר את פרוסות המוח הקטן עם טפטפת פסטר מקוצץ זכוכית רחבה נשא לצלחת 6-היטב (מקסימום 3 פרוסות / בכל טוב). לשאוב את מדיום HBSS.
- דגירה פרוסות (3 מקסימום) ב 5 מיליליטר של Lפתרון oading של צבע הניאון (10 מיקרומטר).
- כדי להגן מפני אור, לכסות את microplate עם רדיד אלומיניום. שים את זה על שולחן ג'יירו-מרגש ב 35 סל"ד במשך 10 דקות ב RT כדי להקל על תיוג תא.
- העברת פרוסות על הקרום של Transwell להכניס (3.0 מיקרומטר גודל נקבובית; איור 2 ד) עם פיפטה פסטר מקוצצת זכוכית רחבה נשא. לשאוב את מדיום הטעינה עם פיפטה.
- הסר את התוסף ולמלא היטב עם 1.9 מיליליטר של DMEM. החלף את הכנס ולהוסיף DMEM של 100 μl על גבי הפרוסה כדי לכסות את הרקמה.
- מניחים את הצלחת המכילה את התרבות מוסיף בתא החממה (37 מעלות צלזיוס, 5% CO 2) עבור 2 שעות שמספיק להתבונן GCS בML. לשקר רקמות שטוחות כדי לאפשר עיקול על הקרום להוסיף (איור 2E). ודא שפרוסות לא מתייבשות.
5. הדמיה Ex vivo באמצעות Confocal Macroscopy
- העבר את הצלחת ללאמכסה הפלסטיק לחממה צמוד לדוכן של macroscope confocal. מניחים כיסוי זכוכית על להכניס את הצלחת של macroscope. לשמור על הטמפרטורה של החדר ב37.0 מעלות צלזיוס ± 0.5 ° C, ולספק את הפרוסות עם זרימת גז קבועה (95% O 2, 5% CO 2) דרך להוסיף הצלחת כדי לשמור על pH הקבוע. חכה לשעה 2 נוספת לפני ניסוי הזמן לשגות.
- מרחק כדי להמחיש הגירת GC בחתכי הרקמה, להאיר את ההכנה עם אור באורך גל 488 ננומטר באמצעות דיודות לייזר דרך macroscope סריקת לייזר confocal מצויד מטרה יבשה X2 (עבודה: 39 מ"מ, קוטר: 58 מ"מ, NA = 0.234), ולזהות פליטת הקרינה 500-530 ננומטר.
- דק כדי לפתור את התנועה של GCS, לרכוש תמונות עם גורם נוסף זום אופטי של 1.5 עד 2.0. לאסוף תמונות של GNS במישור מוקד אחד או עד 10 מטוסי מוקד שונים לאורך ציר z בכל 30 דקות במשך שעה עד 12.
- בעת צורך, להסיר את כיסוי הזכוכית ולהוסיף כמויות קטנות (1-10 μl) של מפעילים או מעכבים ביולוגיים בDMEM עם פיפטה 10 μl ללמוד את השפיעו על הגירת GC.
מעקב 6. סלולארי
- עבור כל שעה של הסרט, לבצע הקרנת Z- מחסנית באמצעות מצב ecart-הסוג בImageJ. לווסת את הניגוד ואת רמות הבהירות של תמונות הרצופות כדי להקל על זיהוי והמעקב של GCS שכותרת. מפה ידנית בכל עמדה על תמונת ההתייחסות (בזמן t = 0).
- השתמש בתוסף "מעקב ידני" בתפריט החלקיקים לנתח ולקבוע על ידי לחיצה על הנקודה של כל גוף תא הכובד במהלך הזמן לשגות. לייצא את נתוני מעקב הגלם בגיליון אלקטרוני.
- לארגן מחדש את נתוני מעקב גלם המיוצאים מImageJ עם תכנית חכמה תוצרת בית (http://primacen.fr, נכתבה בקוד PHP) המזהות את כל תא ועמדות הקשורות. באמצעות התכנית, calculatדואר המרחק הכולל נסע ומהירות הממוצעת של הגירה לכל תא. לסווג ולהשוות מאפיינים של נדידת תאים בתנאי בקרה וטיפול במסננים מתאימים באמצעות אותה התכנית.
תוצאות
במוח הקטן לאחר הלידה המוקדם, GCS מציג שינויים משמעותיים במצב והמהירות של הגירתם כפי שהם חוצים שכבות בקליפת המוח שונות 1 (איור 1). סעיף זה ממחיש דוגמאות של תוצאות שניתן להשיג על ידי לימוד הגירת GC בסביבה הטבעית שלהם סלולרית. פרוסות רקמת המוח הקטן עכברוש P10 שכותרתו עם צבע פלואורסצנטי ירוק נבחנות תחת macroscope confocal (איור 3 א) ואנו מראים כי GCS להעביר רדיאלית בML עם מהירות של 18 מיקרומטר / שעה ממוצעת (איור 3, C). עד כה, את התפקיד של אינטראקציות / תקשורת בין תאים עצביים וגליה כוללים גורמי הרגולציה והמנגנונים מולקולריים המעורבים בשליטה של נדידת תאים בכל שכבת קליפת המוח הוא במידה רבה לא ידועה. כתוצאה מכך, הבעיה העיקרית היא לזהות נוירו, מוליכים עצביים, neurotrophins ורכיבי מטריצה תאיים שיכול לשחק תפקיד באלה שכבה-ספציפי בקליפת המוחשינויי פיק של המהירות במהלך תהליך הגירתם. פוליפפטיד יותרת המוח adenylate הפעלה-cyclase (PACAP) מזוהה בעיקר בPCL, אלא גם בML וIGL בשבועות לאחר הלידה הראשונים במכרסמים 7,10,11. יישום של PACAP38 (10 -6 M) למדיום התרבות הביא 79% ירידה במהירות של GC בML. לדוגמא, מהירות ההעברה של GCS בML ירד מ 11.9 מיקרומטר / שעה בתנאי שליטה 2.5 מיקרומטר / שעה לאחר מתן PACAP38 (איור 4 א). activator רקמות סוג plasminogen (tPA) הוא חבר של מפל מפרקי החלבונים שמוביל לזילות של רכיבי מטריצה תאית (EM) כגון מולקולות הידבקות תא או laminin 12,13. tPA וplasminogen, מצע של tPA, מתגלים בשכבות בקליפת המוח במהלך ההתפתחות של המוח הקטן 14,15,16 לאחר הלידה. מנהל של PAI-1 (10 -7 מ '), מעכב של אנדוגני tPA, הפחית ב- 78% GCהגירה בML. לדוגמא, GCS מופחת מהירות הגירה בML מ19.2 מיקרומטר / שעה בתנאי שליטה 4.2 מיקרומטר / שעה לאחר תוספת של PAI-1 (איור 4). תוצאות אלו מצביעות כי PACAP מפעילה השפעה מעכבת ישירה על תנועות GC ושפרוטאז סרין tPA מאפשר ההגירה של GCS בML של המוח הקטן עכברוש פיתוח.

איור 1: ייצוג 3D של הגירת GC בקליפת המוח הקטנה שלאחר הלידה. 1-4, הרחבה של תהליכי GC והגירה משיקה בEGL. 5, הגירת רדיאלי בML לאורך סיבי גליה ברגמן. 6, שלב חלוף נייח בPCL. 7, הגירת גליה-עצמאית רדיאלי בIGL. 8, השלמת הגירת GC בIGL GC, תא גרגיר, באדום;. EGL, שכבת גרגרים חיצונית; B, גליה ברגמן, בסגול הכהה; G, תא Golgi, בצהוב; CF, סיבי טיפוס, בכחול; ז, תאי גרגיר postmigratory, באור הירוק; IGL, שכבת גרגרים פנימית; MFT, סיבי אזובי מסוף, בצבע ירוק כהה, ML, שכבה מולקולרית; P, תא פורקינג ', באור סגולה; שכבת PCL, פורקינג' תא. נתון זה שונה מ5.

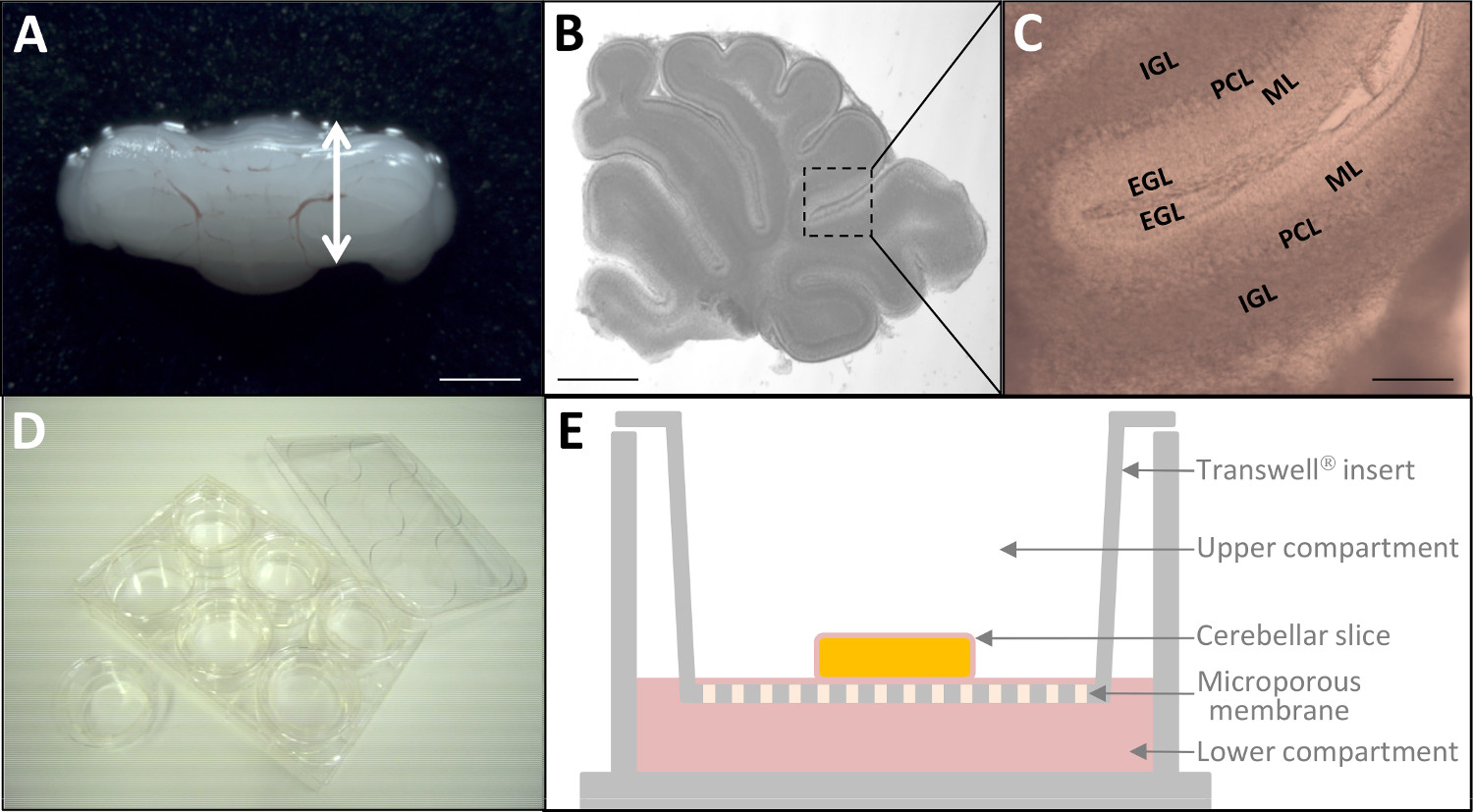
איור 2: תרבות vivo לשעבר של פרוסות המוח הקטן P10 () המוח הקטן גזור מן החולדה P10.. בר סולם = 6 מ"מ. מיקרוסקופ (B) של חיים 180 פרוסה המוח הקטן מיקרומטר עבה דרך stereomicroscopy. בר סולם = 3 מ"מ. (ג) בהגדלה גבוהה יותר, ארבע השכבות בקליפת המוח (EGL, ML, PCL, IGL) של המוח הקטן הן כבר להבחין. בר סולם = 1 מ"מ. (ד) לאחר תיוג ניאון, פרוסות רקמה מונחים על התרבות מוסיף (קוטר 24 מ"מ) לתוך צלחת 6-גם. ייצוג סכמטי של הכנס תרבות עם קרום פוליאסטר תרבית רקמה שטופל (E). אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}

איור 3: הגירה דינמית של GCS בשכבות בקליפת המוח של המוח הקטן P10 העכברוש (א) להציג Macroconfocal (XYZ, הקרנת 2D) של פרוסת המוח הקטן עכברוש P10 בי GCS מסומנים עם צבע פלואורסצנטי cytoplasmic ירוק.. בר סולם = 75 מיקרומטר. זמן לשגות הדמיה מראה GC (ב)תנועות בML ידי macroscopy confocal במשך 4 שעות בתנאי שליטה. כוכבית (*) סמל מסמן את סומה GC. זמן שחלף (בדקות) מצויינים בתחתית כל photomicrograph. בר סולם = 10 מיקרומטר. (ג) שינויים עוקבים במרחק שעובר סומה GC.

איור 4:. אפקט של מעכב פרוטאז neuropeptide והגירת GC GC () היה במעקב בML ידי macroscopy confocal לשעה 2 בתנאי שליטה ולאחר מכן לשעה 2 בנוכחות פוליפפטיד adenylate יותרת המוח ההפעלה-cyclase (PACAP). GC (ב) היה במעקב בML ידי macroscopy confocal לשעה 2 בתנאי שליטה ולאחר מכן לשעה 2 בנוכחות activator plasminogen מעכב-1 (PAI-1).
Discussion
פרוטוקול זה מתאר את התרבות של פרוסות המוח הקטן עכברוש P10 חריפים במערכת Transwell ותיוג הניאון של GC עם צבע ניאון ירוק ללמוד נדידת תאים במהלך התפתחות לאחר לידה באמצעות macroscopy confocal. פרוטוקול זה מאפשר תצפיות של נדידת תאים לתקופה של עד 12 שעות ובדיקה של התפקידים האפשריים של ויסות גורמים בהגירה כוללים אגוניסטים או יריבים של נוירופפטידים, מעכבי אנזים, מאפנני תא איתות או חומרים רעילים במהלך הניסוי. חור קטן בהוספת הקרום עם קצה פיפטה יש צורך להקל על ממשל של תרכובות במדיום הדגירה. פיפטה קצה מעוגלת תוצרת בית ניתן להשתמש כדי להקל על המשלוח של פתרון.
נושא אחד מחקרי נדידת תאים בחי פרוסות רקמה הוא שהתנועות של הרקמה עצמה יכולה לעשות תא מעקב קשה. ואילו גישות קודמות הציעו לייצב SLI בעדינותCES עם רשת ניילון או שכבה דקה של קולגן זנב החולדה 7,17, יתרון עיקרי של טכנולוגיה זו היא ההעברה הפשוטה וישירה של צלחת תרבות 6 היטב המכילה פרוסות המוח הקטן במוסיף קרום מן החממה CO 2 תחת המטרה של macroscope confocal. בקרי טמפרטורה משולבת ו- CO 2 גם מספקים פרמטרים סביבתיים מתאימים וקבועים חיוניים לתא הגירה 9. לכן, תנאי תרבות נשמרים במהלך תצפיות ותנועות רקמה הם מזעריים מאז פרוסות מחוברות היטב להוספת הקרום. ייצוב הרקמות מאומת על ידי ביצוע העמדה של קצוות פרוסה או תאי פורקינג שצריך להיות קבוע אזכור במהלך רכישה. בנוסף, פרוסות המוח הקטן (בין 12 ל 18) שהופצו ב6 הבארות של הצלחת ניתן לצפות במהירות בפירוט בשלב ממונע וזום אופטי. בשל המרחק הגדול עבודה (X2, 39 מ"מ) של המטרה היבשה, עלית-observation הוא ללא טבילה וממשל של תרכובות במדיום התרבות הוא הרבה יותר קל. לכן, פרמטרים סביבתיים ודמיון תמיכת התרבות בCO 2 באינקובטור וconfocal macroscopy להוביל לשימור מרבי של המדגם הביולוגי.
יתרון נוסף של הפרוטוקול הוא השדה הגדול של תצוגה וכתוצאה מכך המספר הגדול של תאים שניתן לראות בו זמנית. לדוגמא, יש לנו קבענו בעבר כי הצפיפות של GCS ניאון עם הגירת רדיאלי בML הייתה 1,124 ± 138 תאים / מ"מ 2 18. macroscopy confocal (X2, NA = 0.234) בעל רזולוציה לרוחב נמוך יותר בהשוואה למיקרוסקופיה confocal (40X, NA = 1.25), אך תא גופות של GCS ניתן לעקוב בקלות והמהירות הממוצעת של הגירה היא השוואה בין שני גישות טכנולוגיות 7,18 .
מלבד שיפור טכני לרכישות תמונה, האיכות של פרוסות וה רקמותאיכות דואר של תיוג הן נקודות מפתח לניסויים מוצלחים. תמיד לשמור על תקשורת ורקמות על קרח במהלך תהליכי נתיחה, לחסל שמן על vibratome להבים ולא לנצל את פרוסות רקמה במגע עם דבק. סעיפי sagittal והרוחביים מותאמים לרדיאלי והגירה משיקה בהתאמה. השתמש באורכים שונים של דגירה לגילוי נאות בשכבות בקליפת המוח השונות של המוח הקטן. פעמים דגירה ארוכות (עד 8 שעות) נחוצות כדי לזהות את ההגירה של מספר רב של GCS בPCL וIGL. מאז הגירת GC הוא תהליך פיסיולוגי במהלך חלונות מרחב ובזמן ספציפיים, השליטה החיובית היא שהתאים צריכים להעביר כראוי. בפרט, GC ציר רב בML הוא אחד מחוון הבריאות העיקרי לפרוסות המוח הקטן sagittal. להתחלת ניסויים, תצפיות של תנועות GC בML הוא הציע. ואכן, את הצורה של GC עם גוף תא מוארך אנכי צריכה להיחשב כנקודת התייחסות להתחיל ACquisition עם שליטה רצופה (2 שעות) וטיפול (2 שעות) תקופות שניתן לבצע בקלות בML.
צבעי ניאון כמו משפחת Tracker הנייד או חלבוני הניאון באו לידי ביטוי במבנים גנטיים יכולים לשמש כקליעים נותבים ללימודי נדידת תאים. בשל קינטיקה האיטית של הגירת GC (מחסנית 1 בכל 30 דקות), יכולים גם להתבצע ניסויי ססגוניות במצב רציף מאז 4 קורות לייזרים (405, 488, 532 ו633 ננומטר) זמינים במערכת. בהתחשב הגירת רדיאלי צנטריפטלי וצנטריפוגלי, מעקב של interneurons אחרת גם יכול להתממש 18. בפרט, יכולים להיות מקומיים סוגי תאים רבים פחות בקלות רבה יותר עם שדה גדול של השקפה. לבסוף, פרוטוקול זה יכול לשמש כדי לחקור נדידת תאים בשלבים אחרים של פיתוח המוח הקטן, אלא גם אזורים אחרים במוח.
Disclosures
יש המחברים אין לחשוף.
Acknowledgements
עבודה זו נתמכה על ידי המכון למחקר והחדשנות בביו-רפואה (IRIB), פלטפורמת ההדמיה הנייד של נורמנדי (PRIMACEN), INSERM, IBiSA, אוניברסיטת רואן, האירופי לפיתוח אזורי הקרן (ERDF - PeReNE, Interreg 4A), LARC-Neurosciences הרשת ואזור Haute-Normandie.
Materials
| Name | Company | Catalog Number | Comments |
| Dulbecco’s modified eagle medium (DMEM) nutrient mixture F-12 | Sigma-Aldrich | D8437 | |
| Hank's balanced salt solution 10x | Sigma-Aldrich | H1641 | |
| PACAP38 | INRS, Canada | Bourgault et al., 200919 | |
| PAI-1 | Calbiochem | 528208 | |
| N-2 supplement | Fisher Scientific / Gibco/ invitrogen | O973 | |
| Cyanoacrylate glue | Loctite | ||
| Cell Tracker Green CMFDA | Invitrogen | C2925 | |
| Polyester Transwell-Clear inserts | Corning | 3452 | |
| Penicillin-Streptomycin | Sigma-Aldrich | P0781 | |
| 6-well cell culture cluster | Corning | 3516 | |
| DMSO | Fisher Scientific | BP231-100 | |
| Tissue culture dish 35 mm diameter | BD Falcon | 353004 | |
| Tissue culture dish 100 mm diameter | Thermo SCIENTIFIC | 130182 | |
| Polypropylen tube (15 ml) | BD Falcon | 352096 | |
| Ethanol 70% | Fisher Chemical | E/0800DF/21 | |
| Biological safety cabinet fume hood | Thermo Scientific | MSC9 Class II A2 | |
| Adjustable-volume pipette (0.5-10 µL) | Eppendorf | 4910 000.018 | |
| Gyro-rocker, SSL3 | Stuart | ||
| CO2 incubator, Hera Cell 150 | Thermo Scientific | ||
| Vibrating blade microtome, VT1000S | Leica Microsystems | ||
| Confocal macroscope, TCS LSI | Leica Microsystems | ||
| Temperature controller | PeCon | ||
| CO2-controller | PeCon | ||
| Stereomicroscope, M205 C | Leica Microsystems | ||
| Operating scissors, curved, blunt/blunt | Medicon | 03.03.17 | |
| Hardened fine iris scissors, straight, sharp/sharp | FST | 149090-11 | |
| Dumont #3 and #5 forceps | FST | 11293-00 and 11252-20 | |
| Vibratome injector blades/single edge | Leica Microsystems | 39053250 | |
| Standard scalpel handle #3 solid | FST | 10003-12 | |
| Surgical blade #15 | Swann-Morton | 205 |
References
- Komuro, Y., Kumada, T., Ohno, N., Foote, K. D., Komuro, H. Migration in the Cerebellum. Cellular Migration and Formation of Neuronal Connections: Comprehensive Developmental Neuroscience. 2, 281-297 (2013).
- Altman, J., Bayer, S. A. Development of the cerebellar system in relation to its evolution, structure, and functions. , CRC Press. Boca Raton, FL. (1997).
- Komuro, H., Rakic, P. Distinct modes of neuronal migration in different domains of developing cerebellar cortex. Journal of Neuroscience. 18 (4), 1478-1490 (1998).
- Komuro, H., Yacubova, E. Recent advances in cerebellar granule cell migration. Cellular and Molecular Life Sciences. 60 (6), 1084-1098 (2003).
- Fahrion, J. K., et al. Rescue of neuronal migration deficits in a mouse model of fetal Minamata disease by increasing neuronal Ca2+ spike frequency. Proceedings of National Academy of Sciences USA. 109 (13), 5057-5062 (2012).
- Raoult, E., et al. Pituitary adenylate cyclase-activating polypeptide (PACAP) stimulates the expression and the release of tissue plasminogen activator (tPA) in neuronal cells: involvement of tPA in the neuroprotective effect of PACAP. Journal of Neurochemistry. 119 (5), 10-1111 (2011).
- Cameron, D. B., et al. Cerebellar cortical-layer-specific control of neuronal migration by pituitary adenylate cyclase-activating polypeptide. Neuroscience. 146 (2), 697-712 (2007).
- Renaud, J., et al. Plexin-A2 and its ligand, Sema6A, control nucleus-centrosome coupling in migrating granule cells. Nature Neuroscience. 4, 440-449 (2008).
- Komuro, H., Rakic, P. Dynamics of granule cell migration: a confocal microscopic study in acute cerebellar slice preparations. Journal of Neuroscience. 15 (2), 1110-1120 (1995).
- Nielsen, H. S., Hannibal, J., Fahrenkrug, J. Expression of pituitary adenylate cyclase activating polypeptide (PACAP) in the postnatal and adult rat cerebellar cortex. Neuroreport. 9 (11), 2639-2642 (1998).
- Hannibal, J. Pituitary adenylate cyclase-activating peptide in the rat central nervous system: an immunohistochemical and in situ hybridization study. Journal of Comparative Neurology. 453 (4), 389-417 (2002).
- Garcia-Rocha, M., Avila, J., Armas-Portela, R. Tissue-type plasminogen activator (tPA) is the main plasminogen activator associated with isolated rat nerve growth cones. Neuroscience Letters. 180 (2), 123-126 (1994).
- Ware, J. H., DiBenedetto, A. J., Pittman, R. N. Localization of tissue plasminogen activator mRNA in the developing rat cerebellum and effects of inhibiting tissue plasminogen activator on granule cell migration. Journal of Neurobiology. 28 (1), 9-22 (1995).
- Friedman, G. C., Seeds, N. W. Tissue plasminogen activator mRNA expression in granule neurons coincides with their migration in the developing cerebellum. Journal of Comparative Neurology. 360 (4), 658-670 (1995).
- Seeds, N. W., Siconolfi, L. B., Haffke, S. P. Neuronal extracellular proteases facilitate cell migration, axonal growth, and pathfinding. Cell and Tissue Research. 290 (2), 367-370 (1997).
- Basham, M. E., Seeds, N. W. Plasminogen expression in the neonatal and adult mouse brain. Journal of Neurochemistry. 77 (1), 318-325 (2001).
- Bourgault, S., et al. Molecular and conformational determinants of pituitary adenylate cyclase-activating polypeptide (PACAP) for activation of the PAC1 receptor. Journal of Medical Chemistry. 52 (10), 3308-3316 (2009).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved