
Method Article
Ex-vivo-Bildgebung von Postnatale Kleinhirn-Körnerzellmigration mittels konfokaler Makros
In diesem Artikel
Zusammenfassung
During postnatal cerebellum development, immature granule cells originating from the germinal zone exhibit distinct modalities of migration to reach their final destination and to establish neuronal networks. This protocol describes the preparation of cerebellar slices and the confocal macroscopic approach used to investigate the factors that regulate neuronal migration.
Zusammenfassung
Während der postnatalen Entwicklung, unreifen Körnerzellen (exzitatorischen Interneuronen) zeigen tangentiale Migration in der äußeren Körnerschicht, und dann radial Migration in der Molekularschicht und der Purkinje-Zellschicht, um die innere Körnerschicht der Kleinhirnrinde zu erreichen. Standard in Migrationsprozesse induziert entweder Zelltod oder falsche Platzierung der Neuronen, was zu Defiziten in diverse Kleinhirnfunktionen. Zentripetale Körnerzellmigration umfasst mehrere Mechanismen, wie Chemotaxis und extrazellulären Matrixabbau, um die Zellen in Richtung ihrer endgültigen Position zu führen, aber die Faktoren, die die Zellmigration in jeder Kortikalschicht regulieren sind nur teilweise bekannt. In unserem Verfahren werden akute zerebelläre Scheiben von P10-Ratten werden Körnerzellen mit einem fluoreszierenden Marker markiert zytoplasmatischen und Gewebe kultiviert werden auf Membraneinsätzen 4-10 h, bevor eine Echtzeitüberwachung der Zellmigration durch konfokale makroskopisch bei 37 ° C in demVorhandensein von CO 2. Während ihrer Wanderung in den verschiedenen kortikalen Schichten des Kleinhirns können Körnerzellen zu Neuropeptid-Agonisten oder -Antagonisten, Proteaseinhibitoren, Blocker von intrazellulären Effektoren oder sogar toxische Substanzen, wie Alkohol oder Methyl ausgesetzt werden, um ihre mögliche Rolle bei der Regulation der neuronalen Migration zu untersuchen .
Einleitung
In sich entwickelnden Kleinhirn sind acht verschiedene Typen von Nervenzellen nacheinander zwischen der zweiten Woche der embryonalen und der zweiten Lebenswoche bei Nagern 1 hergestellt. Ursprung zunächst aus, eine primäre Keimzone, unreife Körnerzellen (GC) werden die letzten Neuronen vom externen Körnerschicht (EGL), einer sekundären Keimzone 2 erzeugt werden. In den ersten drei Wochen nach der Geburt, die Kleinhirnrinde ist eine blättrige Struktur in vier Schichten einschließlich der EGL, der Molekularschicht (ML) organisiert, die Purkinje Zellschicht (PCL) und die innere Körnerschicht (IGL) (Abbildung 1). Durch zentripetale Migration, unreife GCs, glutamatergen Inter, erreichen die IGL innerhalb von ca. 2 Tagen. Von der dritten Lebenswoche, verschwindet die EGL und IGL bildet die sogenannte granulare Schicht (GL) in der Erwachsenen-Cerebellum. In der GL, GCs empfangen exzitatorische synaptische Eingänge von Moosfasern und unipolar Bürstenzellen undhemmende synaptische Eingänge von Golgi Zelle Axone. In der ML, GC Axone bilden Synapsen mit GABAergen Neuronen einschließlich Purkinje-Zellen, Korbzellen, Sternzellen und Golgi-Zellen 2.
Echtzeit-Beobachtung der Bewegung der Zellen in akuten zerebelläre Scheiben von frühen postnatalen Nagetieren zeigt, dass GCs ihre Form ändern, die gleichzeitig mit Änderungen in der Modalität und Migrationsgeschwindigkeit während der Route in der Kleinhirnrinde 3. Während der ersten zwei Wochen nach der Geburt, GC Vorläufer aktiv vermehren sich an der Spitze der EGL. Im mittleren Teil der EGL, postmitotischen GCs tangential wandern in Richtung der größeren Prozesses. Bei der EGL-ML Grenze, GCs verlangsamen ihre Bewegung, beginnen die Zellen, um eine kurze vertikale Steigvorgang in die ML eingeben. In der ML, GCs eine vertikal verlängerte Zellkörper, eine dünne Hinter Verfahren und eine voluminösere führenden Prozess, und radial wandern entlang der Bergmann Gliazellen Fasern. ImPCL, GCs stoppen ihre Bewegung aber nach einer längeren stationären Phase (2 h), die PCL-IGL Grenze zu überqueren sie. Im IGL, GCs wandern in Richtung der Unterseite der Schicht in Abwesenheit von Gliazellen Faserträger. Sobald die Spitzen der führenden Prozessansatz der IGL-weißen Substanz (WM) Grenze, GCs langsam und ihre Bewegung zu stoppen. Querschnitten des Kleinhirns für tangential Migrationsstudien in der EGL bevorzugt, während sagittal sind bemüht, die Migration in der ML, PCL und IGL radial. Einige regulatorischen Faktoren von GC Bewegungen einschließlich Neuropeptide (zB Somatostatin, PACAP) wurden bisher identifiziert, aber die vollständige Mechanismen in der raum-zeitlichen Steuerung der GC Migration in jedem Rindenschicht beteiligt sind noch weitgehend unbekannt 1,4,5,6.
GC Migration hat sich in den letzten 20 Jahren durch Video- und konfokale Mikroskopie entweder Durchlichtbeleuchtung für isolierte kultivierte Zellen oder Fluoreszenzdetektion für acu suchtte Kleinhirnschnitten. Zunächst lipophilen DiI und neuerdings "Zell Tracker" Farbstoffe und Zell-exprimiert fluoreszierende Proteine wurden für die konfokale oder Zwei-Photonen-Mikroskopie 7,8 eingesetzt. Erfolgreiche Experimente abhängig von einer Reihe von spezifischen Prozeduren, die das Protokoll einfach, aber nicht einfach. Insbesondere haben akute Scheiben während Beobachtungen im allgemeinen mit einer hausgemachten Nylonmaschennetz 9 stabilisiert sein. Die Intensität der Lichtbeleuchtung so gering wie möglich, Phototoxizität und Photobleaching vermeiden, indem Mehr konfokalen Mikroskop Ansatz vorgeschlagen wird. Darüber hinaus, Temperatur und CO 2 sind die wichtigsten Umweltparameter seit Instabilität kann neuronalen Migration beeinflussen. Zu erleichtern und zu verfeinern die experimentellen Verfahren, haben wir ein Protokoll, das konfokale Makros Scheibe Bewegungen begrenzt, sorgt für eine konstante Umgebungsparameter, verringert Photobleichen, erhöht Gesichtsfeld (in der Größenordnung von Millimetern) und consequ entwickeltständig die Anzahl der Zellen (Dutzenden), die durch Bildanalyse verfolgt werden können. Somit sind 180 & mgr; m dicke Scheiben auf Membraneinsätze kultiviert und 6-well Platten werden direkt unter einem 2X motorisierten Objektiv eines kommerziellen konfokalen Makroskops mit großem Inkubationskammer, Temperatur und CO 2 Controllern und einem Vibrationskontrollsystem ausgestattet war. Zeitlücken und z-Stapel werden dann über mehrere Stunden und pharmakologische Werkzeuge durchgeführt oder bioaktive Moleküle können hinzugefügt oder in das Inkubationsmedium abgegeben wird. Dieses Verfahren könnte auch angepasst ist, um die Wanderung der anderen Typen von Neuronen im Kleinhirn oder Cerebrum in verschiedenen Entwicklungsstadien zu untersuchen.
Protokoll
Tiere (männliche oder weibliche Wistar-Ratten) geboren wurden und in einem akkreditierten Tierhaltung (Genehmigung B.76-451-04) gezüchtet, nach der Französisch Führer für die Pflege und Verwendung von Labortieren. Die Experimente wurden unter der Aufsicht von autorisierten Ermittler (MB, DV und LG) in Übereinstimmung mit der europäischen Richtlinie Community Council (2010/63 / UE des 22. September 2010) und die Französisch Ministerium für Landwirtschaft durchgeführt.
1. Vorbereitung der Medien und Werkzeuge
- In einer biologischen Sicherheitswerkbank, bereiten 1x Hanks BSS (HBSS) in sterilem Wasser von 10x Stammlösung, die CaCl 2 (1,85 g / l) / MgSO 4 (0,9767 g / l) ohne MgCl 2. In NaHCO 3 (350 ug / ml) zur Lösung von HBSS 1x.
- In einer biologischen Sicherheitswerkbank, fügen N2-Ergänzung (von einem 100x Stocklösung) und Penicillin (100 Einheiten / ml) -streptomycin (0,1 mg / ml) Lösung Dulbeccos modifiziertem Eagle Medium (DMEM) Nährstoff-Mixtur F-12 (1: 1).
- Unter sterilen Bedingungen, Herstellung eines Aliquots (25 ul, 2 mM) der zytoplasmatischen Fluoreszenzfarbstoff als Zell Tracker Green in DMSO (1,075 ml 1 mg). Verdünnter einem Aliquot in 5 ml DMEM in einer 15 ml konischen Röhrchen.
- Bereiten gefüllt Eiskübel Medien bei 4 ° C zu halten.
- Dekontaminieren Labortischplatten und Werkzeuge mit 70% Ethanol.
2. Dissection von Kleinhirn von P10 Ratten
- Schnell zu enthaupten Rattenjungen (P10) mit gebogenen Betriebs Schere hinter den Ohren, um den Beginn des Rückenmarks zu erhalten.
- An der Rückseite des abgeschlagenen Kopf, stellen Mittellinienschnitt der Haut vom Hals bis zur Nase mit feinen Iris Schere und trennen die Haut vom Schädel mit feinen Iris Schere und Dumont # 3 Pinzette.
- Verwenden Sie feinen Iris Schere, um zart zu machen zwei seitliche Schnitte von der Basis zum rostralen Bereich des Schädels. Entfernen Sie die seziert Schädel mit zwei # 3 Pinzette. Nehmen Sie den BHin von jeder Haftung mit dem Totenkopf mit den gleichen Zangen.
- Übertragen Sie das Gehirn mit dem Löffel Ende einer Spatel Petrischale (Ø 35 mm) mit 2 ml eiskaltem HBSS Medium.
- Legen Sie die Petrischale mit dem Gehirn in einem größeren Petrischale (Ø 100 mm) mit Eis und Übertragung auf die Bühne des Stereomikroskops.
- Unter einem Stereomikroskop, zu isolieren, das Kleinhirn aus dem Gehirn von Krümmung mit zwei # 3 Pinzette. Ebenso entfernen Sie die Restrückenmark und die Pia-Membran.
- Übertragen Sie das Kleinhirn in einem neuen HBSS gefüllte Petrischale (Ø 35 mm, 2 ml) mit dem Löffel Ende einer Spachtel und auf Eis halten.
3. Herstellung von Acute Cerebellar Slices
- Unter dem Stereomikroskop, schneiden Sie das Kleinhirn zwischen den Wurm und der rechten Hemisphäre, wie durch die zwei Köpfe Pfeil in 2A mit einem Standard-Skalpellgriff # 3 festen und einem # 15 Skalpell.
- Geben Sie einen Tropfen Cyanoacrylate Kleber auf der Vibratom Objektplatte und warten Sie 15-25 Sekunden, um giftige Lösemitteldämpfen zu beseitigen.
- Sammeln Sie die geschnittenen Kleinhirn mit dem Löffel Ende einer Spachtel und überschüssiges HBSS mit sauberen Papiertüchern.
- Bringen Sie das Kleinhirn in der Nähe der Objektplatte. Befestigen Sie die Schnittkante an die Objektplatte und warten Sie 10 sec.
- Legen Sie die Objektplatte in die Pufferwanne mit dem Manipulator, und drehen Sie es, so dass die Querachse des Kleinhirns ist senkrecht zu der Messerhalter. Befestigen Sie den Probenteller mit einem Inbusschlüssel und füllen sanft Pufferwanne mit HBSS Medium, bis das Kleinhirn bedeckt.
- Last zerstoßenem Eis in das Kühlbad.
- Die Schaufel dreimal mit 70% Ethanol gereinigt werden, um jegliches Öl zu beseitigen.
- Legen Sie die Klinge in den Messerhalter einsetzen und mit Klemmschraube.
- Setzen Sie die Schneide direkt hinter der Hinterkante (aus der Sicht des Benutzers) der Probe und definieren es als Ausgangspunkt. Verwenden Sie nach vorne zu befehlen, zu definieren ter Endpunkt, nachdem die vordere Kante des Objektes.
- Wählen Schneidegeschwindigkeit bei 2,5 und Schnittfrequenz am 8. Wählen Trimmdicke bei 180 & mgr; m. Starten Gewebe Schnitte.
- Nehmen Sie jeden Abschnitt mit einem Weit Bohrung Glaspasteurpipette abgeschnitten und Transfer zum HBSS enthaltenden Petrischale (Ø 35 mm) auf Eis gehalten.
- Mit Hilfe von zwei # 5 Zange, entfernen Sie die Hirnhäute vorsichtig aus dem Kleinhirn, wenn Eingriffe in die Klinge. Sammeln maximal 5 Scheiben pro Cerebellum (Abbildung 2B, C).
- Entfernen Sie die Hirnhäute von den Kleinhirnscheiben sorgfältig mit zwei # 5 Zange unter dem Stereomikroskop und trennen Sie die Läppchen sanft für bessere Tastkopfbelastung.
4. Fluoreszenzfärbung Wohninterneuronen
- Übertragen Sie Kleinhirnschnitten mit einem Breitbohrungsglas abgeschnitten Pasteurpipette auf eine Platte mit 6 Vertiefungen (max 3 Scheiben / pro Vertiefung). Saugen Sie das HBSS Medium.
- Scheiben (3 max) in 5 ml L inkubierenoading Lösung des Fluoreszenzfarbstoffs (10 uM).
- Den Inhalt vor Licht zu schützen, decken Sie die Mikrotiterplatte mit Aluminiumfolie. Legte es auf eine Kreiselbewegungstisch bei 35 UpM für 10 min bei RT zu Markierung von Zellen zu erleichtern.
- Transfer Scheiben auf der Membran eines Transwell einfügen (3,0 & mgr; m Porengröße; 2D) mit einer großen Bohrung Glaspasteurpipette abgeschnitten. Saugen Sie das Lade Medium mit Pipette.
- Entfernen Sie den Einsatz und füllen Sie die gut mit 1,9 ml DMEM. Ersetzen Sie den Einsatz und fügen Sie 100 ul DMEM auf der Oberseite der Scheibe, um das Gewebe zu bedecken.
- Die Platte, die die Kultureinsätze in der Inkubationskammer (37 ° C, 5% CO 2) 2 Stunden lang, die ausreicht, um GCs bei der ML zu beobachten ist. Gewebe liegen flach, um die Befestigung an dem Einsatz-Membran (2E) zu ermöglichen. Stellen Sie sicher, dass Scheiben sind nicht trocknen.
5. Ex-vivo-Imaging Durch konfokale Makros
- Übertragen Sie die Platte ohnedie Kunststoffdeckel in einen Inkubator in den Stand eines konfokalen Makroskop angebracht. Legen Sie ein Deckglas auf dem Platteneinsatz des Makroskop. Halten der Temperatur der Kammer bei 37,0 ° C ± 0,5 ° C, und liefern die Scheiben mit konstanter Gasstrom (95% O 2, 5% CO 2) durch den Platteneinsatz, um den pH konstant zu halten. Warten Sie für 2 weitere Stunden vor Zeitraffer-Experiment.
- GC-Migration in den Gewebeschnitten sichtbar zu machen, beleuchten die Präparation mit einem 488 nm Wellenlängenlicht mit Hilfe einer Laserdiode durch ein konfokales Laser-Scanning Makroskops mit X2 Trockenobjektiv ausgestattet (Arbeitsabstand: 39 mm, Durchmesser: 58 mm, NA = 0,234) und erkennt die Fluoreszenzemission von 500 bis 530 nm.
- Um die Bewegung der GCs fein zu beheben, Bilder zu erfassen mit einem zusätzlichen optischen Zoomfaktor von 1,5 bis 2,0. Sammeln Bilder GNS in einer einzigen Brennebene oder bis zu 10 verschiedenen Brennebenen entlang der z-Achse alle 30 min bis zu 12 h.
- Wenn nötig, entfernen Sie die Glasabdeckung und fügen Sie kleine Mengen (1-10 & mgr; l) von biologischen Aktivatoren oder Inhibitoren in DMEM mit 10 ul-Pipette, um ihre Auswirkungen auf die GC-Migration zu studieren.
6. Handy-Tracking
- Für jede Zeit des Films, führen z-Stapel Projektion durch den Ecart-type-Modus in ImageJ. Modulieren der Kontrast und die Helligkeit der aufeinanderfolgenden Bilder, um die Identifikation und die Verfolgung von markierten GCs zu erleichtern. Karte manuell jede Position auf dem Referenzbild (bei t = 0).
- Verwenden Sie die "Manual tracking" Plugin in der Analyse Particle Menü und bestimmen, indem Sie auf die Schwerpunkt jeder Zelle Körper während der Zeitraffer. Exportieren Sie die raw-Tracking-Daten in einer Tabellenkalkulation.
- Reorganisieren die exportierten raw Tracking-Daten von ImageJ mit einem Smart-Home-made-Programm (http://primacen.fr, in PHP-Code geschrieben), dass jede Zelle und der zugehörigen Positionen zu identifizieren. Mit dem Programm, calculate die Gesamtstrecke und die durchschnittliche Geschwindigkeit der Migration für jede Zelle. Klassifizieren und vergleichen Eigenschaften der Zellmigration in Kontroll- und Behandlungsbedingungen unter geeigneten Filter mit dem gleichen Programm.
Ergebnisse
In der frühen postnatalen Cerebellum GCs signifikante Änderungen ihrer Betriebsart und der Migration als sie sich kreuzen verschiedenen kortikalen Schichten 1 (Abbildung 1). In diesem Abschnitt werden Beispiele für Ergebnisse, die durch das Studium GC Migration in ihrer natürlichen zellulären Milieu erhalten werden können. P10 Rattenkleinhirn-Gewebeschnitte mit einem grünen Fluoreszenzfarbstoff markiert sind unter einem konfokalen Makroskop (3A) untersucht, und wir zeigen, dass GCs radial wandern im ML mit einer Durchschnittsgeschwindigkeit von 18 & mgr; m / h (3B, C). Bisher hat sich die Rolle der Wechselwirkung / Kommunikation zwischen neuronalen und glialen Zellen einschließlich der regulatorischen Faktoren und molekularen Mechanismen bei der Steuerung der Zellmigration in jeder Rindenschicht beteiligt sind weitgehend unbekannt. Folglich ist das Hauptproblem zu Neuropeptide, Neurotransmitter, Neurotrophine und extrazellulären Matrixkomponenten, die in diesen Rindenschicht-Spezi eine Rolle spielen könnte identifizierenfic Änderungen der Geschwindigkeit während des Migrationsprozesses. Hypophysen-Adenylat-Cyclase-aktivierendes Polypeptid (PACAP) wird hauptsächlich in der PCL erfasst, sondern auch bei der ML und IGL während der ersten zwei Lebenswochen in Nagetieren 7,10,11. Anwendung von PACAP38 (10 -6 M) zu dem Kulturmedium führte zu einer 79% Verringerung der Geschwindigkeit GC in der ML. Zum Beispiel kann die Migrationsgeschwindigkeit des GCs bei der ML sank von 11,9 & mgr; m / h in Steuerbedingungen bis 2,5 um / h nach der Verabreichung von PACAP38 (4A). Gewebe-Plasminogen-Aktivator (tPA) ist ein Mitglied der proteolytischen Kaskade, die dem Abbau der extrazellulären Matrix (EM) Komponenten führt, wie Zelladhäsionsmoleküle oder Laminin 12,13. tPA und Plasminogen, einem Substrat von tPA werden in kortikalen Schichten während der Entwicklung des postnatalen Cerebellum 14,15,16 detektiert. Verabreichung von PAI-1 (10 -7 M), einem Inhibitor der endogenen tPA, um 78% der GC vermindertemMigration in der ML. Beispielsweise erniedrigte GCs Migrationsgeschwindigkeit in der ML von 19,2 & mgr; m / h in Steuerbedingungen auf 4,2 um / h nach der Zugabe von PAI-1 (Abbildung 4B). Diese Ergebnisse zeigen, dass PACAP übt eine direkte inhibitorische Wirkung auf die GC Bewegungen und daß die Serinprotease tPA erleichtert die Wanderung von GCs bei der ML des entwickelnden Rattenkleinhirn.

Abbildung 1: 3D-Darstellung der GC-Migration in der postnatalen Kleinhirnrinde. 1-4, Erweiterung des GC Prozesse und tangentiale Migration in der EGL. 5, Radial Migration im ML entlang Bergmann Gliazellen Fasern. 6, Transient stationäre Phase in der PCL. 7, Glial unabhängige Radial Migration in der IGL. 8 Fertigstellung von GC Migration in der IGL GC, Körnerzellen, in rot;. EGL, Externen Körnerschicht; B, Bergmann Gliazellen, in dunkelviolett; G, Golgi Zelle, in gelb, cf, Kletterfasern, in blau; g, postmigratory Körnerzellen, in hellgrün; IGL, internen Körnerschicht; mft, Moosfaser Terminal, in dunkelgrün; ML, molekularen Schicht; P, Purkinje Zelle, in Licht lila, PCL, Purkinje Zellschicht. Diese Zahl hat sich von 5 modifiziert.

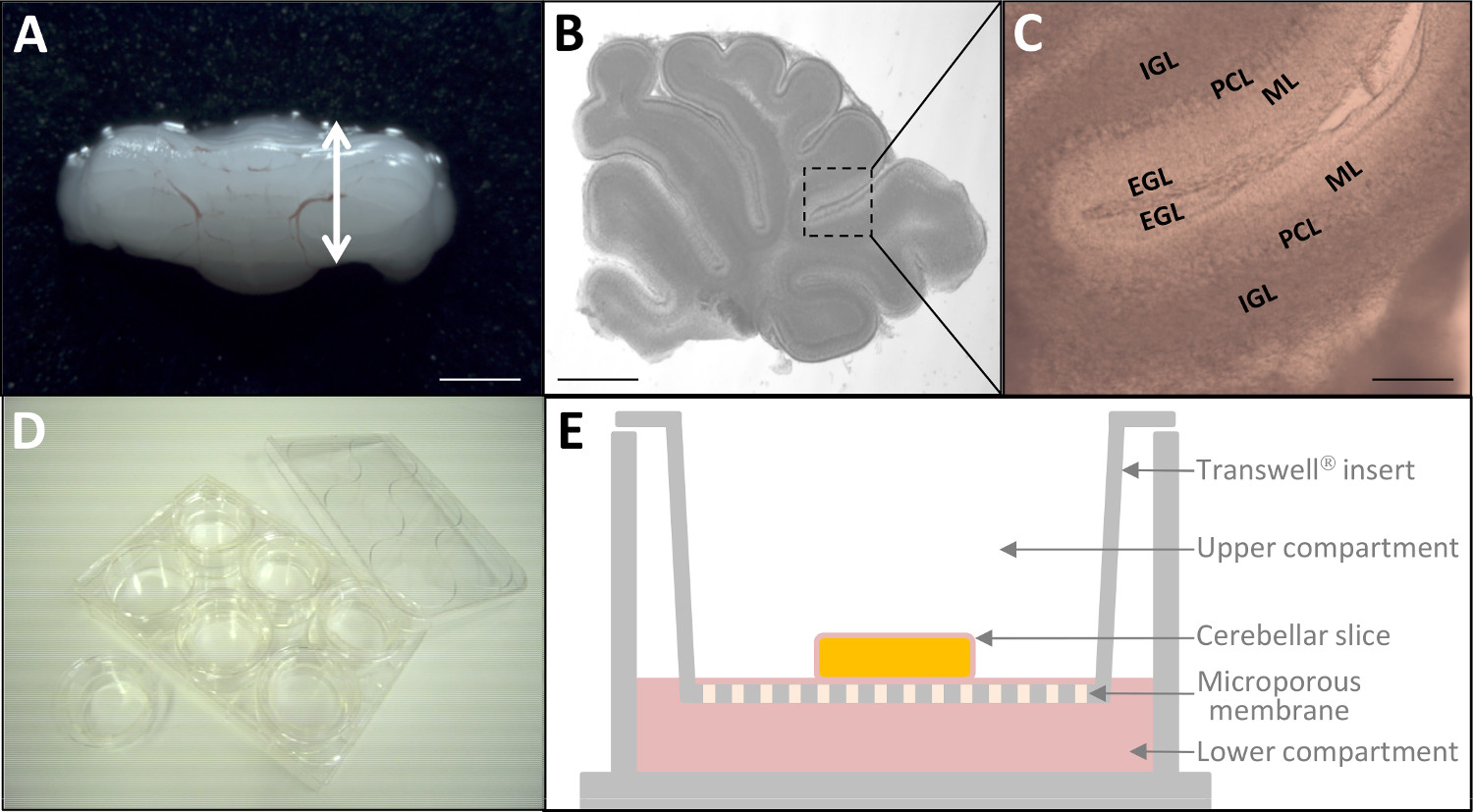
Abbildung 2: Ex-vivo-Kultur von P10 Kleinhirnscheiben (A) seziert Kleinhirn von Ratten-P10.. Maßstabsbalken = 6 mm. (B) Mikroskopische Aufnahme leben 180 um dicke Kleinhirn Scheibe durch Stereomikroskopie. Maßstabsleiste = 3 mm. (C)ein höherer Vergrößerung werden die vier kortikalen Schichten (EGL, ML, PCL, IGL) des Kleinhirns sind bereits erkennbar. Maßstabsbalken = 1 mm. (D) Nach Fluoreszenzmarkierung, sind Gewebeschnitte auf Kultureinsätze (24 mm Durchmesser) in einer 6-Well Platte gelegt. (E) Schematische Darstellung eines Kultureinsatz mit Gewebekultur-behandelte Polyestermembran. Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
{kind=link}

Abbildung 3: Dynamische Migration von GCs in den kortikalen Schichten des Ratten P10 Kleinhirn (A) Macroconfocal Ansicht (xyz, 2D-Projektion) eines P10 Ratte Kleinhirnscheibe, in der GCs sind mit einem grünen zytoplasmatischen Fluoreszenzfarbstoff markiert.. Balken = 75 & mgr; m. (B) Zeitraffer-Bildgebung, die GCBewegungen in der ML durch konfokale Makros 4 h in Kontrollbedingungen. Stern (*) Symbol markiert den GC soma. Abgelaufene Zeit (in min) ist auf dem Boden jeder Mikroaufnahme dargestellt. Maßstabsbalken = 10 & mgr; m. (C) Sequential Abstandsänderungen durch GC Soma bereist.

Abb. 4: Wirkung von Neuropeptid und Protease-Inhibitor der Migration GC (A) GC wurde in der ML durch konfokale makroskopische 2 Stunden in Kontrollbedingungen und verfolgt dann für 2 h in Gegenwart von Hypophysen-Adenylat-Cyclase-aktivierendes Polypeptid (PACAP). (B) GC wurde in der ML durch konfokale makroskopische 2 Stunden in Kontrollbedingungen nachgeführt und dann für 2 h in Gegenwart von Plasminogen-Aktivator-Inhibitor-1 (PAI-1).
Diskussion
Dieses Protokoll beschreibt die Kultur der akuten P10 Rattenkleinhirn-Scheiben in der Transwell-System und der Fluoreszenzmarkierung von GC mit einem grünen Fluoreszenzfarbstoff an Zellmigration während der postnatalen Entwicklung durch konfokale Makros studieren. Dieses Protokoll ermöglicht Beobachtungen der Zellmigration für einen Zeitraum bis zu 12 Stunden und Prüfung der möglichen Rollen der Regulierung Faktoren Migration einschließlich Agonisten oder Antagonisten von Neuropeptiden, Enzyminhibitoren, Zellsignalmodulatoren oder giftige Stoffe während des Experiments. Ein kleines Loch in der Membran Einsatz mit einer Pipettenspitze ist notwendig, um die Verabreichung von Verbindungen im Inkubationsmedium zu erleichtern. Ein hausgemachtes gekrümmte Spitze Pipette verwendet, um die Lieferung der Lösung zu erleichtern.
Ein Problem der Zellmigration Studien am lebenden Gewebeschnitt ist, dass die Bewegungen des Gewebes selbst machen kann Zellverfolgung schwierig. Während frühere Ansätze wurden vorgeschlagen, um sanft zu stabilisieren slices mit einer Nylon Netz oder eine dünne Schicht aus Rattenschwanz Kollagen 7,17, ist ein großer Vorteil dieser Technologie die einfache und direkte Übertragung von einer 6-Well-Kulturplatte, die Kleinhirnscheiben auf Membraneinsätze aus dem CO 2 Inkubator im Rahmen des Ziels eines konfokalen Makroskops. Integrierte Temperatur und CO 2-Controller bieten auch wichtig für die Zellwanderung 9 geeignete und konstante Umgebungsparameter. Daher werden Kulturbedingungen während der Beobachtung und Gewebebewegungen gehalten werden minimiert, da Scheiben sind dem Membraneinsatz befestigt. Stabilisierung des Gewebes wird durch die Position des folgenden Scheibenkanten bzw. Purkinje-Zellen, die Referenzen während der Erfassung festgelegt werden sollte verifiziert. Zusätzlich zerebelläre Scheiben (12 bis 18) in den 6 Vertiefungen der Platte verteilt sind, schnell im Detail mit einem motorisierten Objekttisch und einem optischen Zoom zu beachten. Aufgrund der großen Arbeitsabstand (X2, 39 mm) der Trockenobjektiv, epi-observation frei von Tauch- und Verabreichung von Verbindungen in dem Kulturmedium ist viel einfacher. Daher Makrosumweltparameter und Kultur Unterstützung Ähnlichkeiten in CO 2-Inkubator und konfokale maximale Erhaltung der biologischen Probe führen.
Ein weiterer Vorteil des Protokolls ist das große Sichtfeld und damit die große Anzahl von Zellen, die gleichzeitig beobachtet werden können. Zum Beispiel haben wir bereits festgestellt, dass die Dichte der fluoreszierenden GCs mit radialen Wanderung in der ML war 1124 ± 138 Zellen / mm 2 18. Konfokale Makros (X2, NA = 0,234) eine geringere laterale Auflösung im Vergleich zu konfokalen Mikroskopie (40x, NA = 1,25), aber Zellkörper der GCs können leicht verfolgt werden, und die Durchschnittsgeschwindigkeit von Migration ist vergleichbar zwischen den beiden Technologieansätze 7,18 .
Neben technischen Verbesserungen für Bildaufnahmen, die Qualität der Gewebeschnitte und the Qualität der Kennzeichnung sind die Schlüsselpunkte für erfolgreiche Experimente. Halten Sie Medien und Gewebe auf Eis beim Präparieren Prozesse, eliminieren Öl auf Vibratom Schaufeln und Scheiben von Gewebe nicht nutzen, in Kontakt mit Leim. Sagittal und Querschnitte geeignet sind, die jeweils radial und tangential Migration. Verwenden Sie unterschiedliche Längen der Inkubation entsprechenden Nachweis in den verschiedenen kortikalen Schichten des Kleinhirns. Lange Inkubationszeiten (bis zu 8 h) sind notwendig, um die Migration von zahlreichen GCs in der PCL- und IGL detektieren. Da GC Migration ist ein physiologischer Prozess in bestimmten räumlich-zeitlichen Fenstern, ist die positive Kontrolle, dass die Zellen, um richtig zu migrieren. Insbesondere ist zahlreichen Spindel GC in der ML eines der wichtigsten Gesundheitsindikatoren für sagittal Kleinhirnschnitten. Zum Starten Experimenten wird Beobachtung der GC Bewegungen in der ML vorgeschlagen. In der Tat sollte die Form der GC mit vertikal länglichen Zellkörper als Referenzpunkt, um ac starten berücksichtigt werdenErwerb mit aufeinanderfolgenden Steuer (2 h) und Behandlung (2 Stunden) Zeiten, die leicht in der ML ausgeführt werden können.
Fluoreszenzfarbstoffe, wie die Zelle Tracker Familie oder fluoreszierende Proteine durch genetische Konstrukte exprimiert als Tracer für die Zellmigration Studien verwendet werden. Aufgrund der langsamen Kinetik der GC Migration (1 Stapel alle 30 Minuten), mehrfarbige Experimente können auch im sequentiellen Modus seit 4 Laser Strahlen (405, 488, 532 und 633 nm) auf dem System durchgeführt werden sollen. In Anbetracht zentripetalen und zentrifugalen Radial Migration, Verfolgung anderer Inter realisierbar 18 werden. Insbesondere können weniger zahlreichen Zelltypen leichter mit einem großen Sichtfeld zu lokalisieren. Schließlich kann dieses Protokoll verwendet, um die Zellmigration auf anderen Stufen der Kleinhirnentwicklung, sondern auch andere Hirnareale zu untersuchen.
Offenlegungen
Die Autoren haben nichts zu offenbaren.
Danksagungen
Diese Arbeit wurde durch das Institut für Forschung und Innovation in der Biomedizin (IRIB), der Cell Imaging Platform der Normandie (PRIMACEN), Inserm, ibisa, der Universität von Rouen, dem Europäischen Fonds für regionale Entwicklung (EFRE - PERENE, Interreg 4A), die LARC-Netzwerk Neurowissenschaften und der Region Haute-Normandie.
Materialien
| Name | Company | Catalog Number | Comments |
| Dulbecco’s modified eagle medium (DMEM) nutrient mixture F-12 | Sigma-Aldrich | D8437 | |
| Hank's balanced salt solution 10x | Sigma-Aldrich | H1641 | |
| PACAP38 | INRS, Canada | Bourgault et al., 200919 | |
| PAI-1 | Calbiochem | 528208 | |
| N-2 supplement | Fisher Scientific / Gibco/ invitrogen | O973 | |
| Cyanoacrylate glue | Loctite | ||
| Cell Tracker Green CMFDA | Invitrogen | C2925 | |
| Polyester Transwell-Clear inserts | Corning | 3452 | |
| Penicillin-Streptomycin | Sigma-Aldrich | P0781 | |
| 6-well cell culture cluster | Corning | 3516 | |
| DMSO | Fisher Scientific | BP231-100 | |
| Tissue culture dish 35 mm diameter | BD Falcon | 353004 | |
| Tissue culture dish 100 mm diameter | Thermo SCIENTIFIC | 130182 | |
| Polypropylen tube (15 ml) | BD Falcon | 352096 | |
| Ethanol 70% | Fisher Chemical | E/0800DF/21 | |
| Biological safety cabinet fume hood | Thermo Scientific | MSC9 Class II A2 | |
| Adjustable-volume pipette (0.5-10 µL) | Eppendorf | 4910 000.018 | |
| Gyro-rocker, SSL3 | Stuart | ||
| CO2 incubator, Hera Cell 150 | Thermo Scientific | ||
| Vibrating blade microtome, VT1000S | Leica Microsystems | ||
| Confocal macroscope, TCS LSI | Leica Microsystems | ||
| Temperature controller | PeCon | ||
| CO2-controller | PeCon | ||
| Stereomicroscope, M205 C | Leica Microsystems | ||
| Operating scissors, curved, blunt/blunt | Medicon | 03.03.17 | |
| Hardened fine iris scissors, straight, sharp/sharp | FST | 149090-11 | |
| Dumont #3 and #5 forceps | FST | 11293-00 and 11252-20 | |
| Vibratome injector blades/single edge | Leica Microsystems | 39053250 | |
| Standard scalpel handle #3 solid | FST | 10003-12 | |
| Surgical blade #15 | Swann-Morton | 205 |
Referenzen
- Komuro, Y., Kumada, T., Ohno, N., Foote, K. D., Komuro, H. Migration in the Cerebellum. Cellular Migration and Formation of Neuronal Connections: Comprehensive Developmental Neuroscience. 2, 281-297 (2013).
- Altman, J., Bayer, S. A. . Development of the cerebellar system in relation to its evolution, structure, and functions. , (1997).
- Komuro, H., Rakic, P. Distinct modes of neuronal migration in different domains of developing cerebellar cortex. Journal of Neuroscience. 18 (4), 1478-1490 (1998).
- Komuro, H., Yacubova, E. Recent advances in cerebellar granule cell migration. Cellular and Molecular Life Sciences. 60 (6), 1084-1098 (2003).
- Fahrion, J. K., et al. Rescue of neuronal migration deficits in a mouse model of fetal Minamata disease by increasing neuronal Ca2+ spike frequency. Proceedings of National Academy of Sciences USA. 109 (13), 5057-5062 (2012).
- Raoult, E., et al. Pituitary adenylate cyclase-activating polypeptide (PACAP) stimulates the expression and the release of tissue plasminogen activator (tPA) in neuronal cells: involvement of tPA in the neuroprotective effect of PACAP. Journal of Neurochemistry. 119 (5), 10-1111 (2011).
- Cameron, D. B., et al. Cerebellar cortical-layer-specific control of neuronal migration by pituitary adenylate cyclase-activating polypeptide. Neuroscience. 146 (2), 697-712 (2007).
- Renaud, J., et al. Plexin-A2 and its ligand, Sema6A, control nucleus-centrosome coupling in migrating granule cells. Nature Neuroscience. 4, 440-449 (2008).
- Komuro, H., Rakic, P. Dynamics of granule cell migration: a confocal microscopic study in acute cerebellar slice preparations. Journal of Neuroscience. 15 (2), 1110-1120 (1995).
- Nielsen, H. S., Hannibal, J., Fahrenkrug, J. Expression of pituitary adenylate cyclase activating polypeptide (PACAP) in the postnatal and adult rat cerebellar cortex. Neuroreport. 9 (11), 2639-2642 (1998).
- Hannibal, J. Pituitary adenylate cyclase-activating peptide in the rat central nervous system: an immunohistochemical and in situ hybridization study. Journal of Comparative Neurology. 453 (4), 389-417 (2002).
- Garcia-Rocha, M., Avila, J., Armas-Portela, R. Tissue-type plasminogen activator (tPA) is the main plasminogen activator associated with isolated rat nerve growth cones. Neuroscience Letters. 180 (2), 123-126 (1994).
- Ware, J. H., DiBenedetto, A. J., Pittman, R. N. Localization of tissue plasminogen activator mRNA in the developing rat cerebellum and effects of inhibiting tissue plasminogen activator on granule cell migration. Journal of Neurobiology. 28 (1), 9-22 (1995).
- Friedman, G. C., Seeds, N. W. Tissue plasminogen activator mRNA expression in granule neurons coincides with their migration in the developing cerebellum. Journal of Comparative Neurology. 360 (4), 658-670 (1995).
- Seeds, N. W., Siconolfi, L. B., Haffke, S. P. Neuronal extracellular proteases facilitate cell migration, axonal growth, and pathfinding. Cell and Tissue Research. 290 (2), 367-370 (1997).
- Basham, M. E., Seeds, N. W. Plasminogen expression in the neonatal and adult mouse brain. Journal of Neurochemistry. 77 (1), 318-325 (2001).
- Bourgault, S., et al. Molecular and conformational determinants of pituitary adenylate cyclase-activating polypeptide (PACAP) for activation of the PAC1 receptor. Journal of Medical Chemistry. 52 (10), 3308-3316 (2009).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten