
Method Article
Imagen de la producción de interferón gamma in situ en el bazo de ratón después de la infección por Listeria monocytogenes
En este artículo
Resumen
Aquí, describimos un método de imagen confocal simple para visualizar la localización in situ de las células que secretan la citoquina Interferón gamma en órganos linfoides secundarios murinos. Este protocolo se puede extender para la visualización de otras citoquinas en diversos tejidos.
Resumen
Las citoquinas son pequeñas proteínas secretadas por las células, que median las comunicaciones celulares que son cruciales para obtener respuestas inmunitarias eficaces. Una característica de las citoquinas es su pleiotropismo, ya que son producidos por y pueden afectar a una multitud de tipos de células. Como tal, es importante entender no sólo qué células están produciendo citoquinas, sino también en qué entorno lo hacen, con el fin de definir terapias más específicas. Aquí, describimos un método para visualizar la producción de citoquinas in situ después de la infección bacteriana. Esta técnica se basa en imágenes de células productoras de citoquinas en su entorno nativo mediante microscopía confocal. Para ello, las secciones de tejido se tiñen para marcadores de varios tipos de células junto con una mancha de citoquinas. La secreción de citoquinas, clave para este método, se bloquea directamente in vivo antes de cosechar el tejido de interés, lo que permite la detección de la citoquina que se acumula dentro de las células productoras. Las ventajas de este método son múltiples. En primer lugar, se conserva el microambiente en el que se producen las citoquinas, lo que podría informar en última instancia sobre las señales necesarias para la producción de citoquinas y las células afectadas por esas citoquinas. Además, este método da una indicación de la ubicación de la producción de citoquinas in vivo, ya que no se basa en la reestimulación artificial in vitro de las células productoras. Sin embargo, no es posible analizar simultáneamente la señalización descendente de citoquinas en las células que reciben la citoquina. Del mismo modo, las señales de citoquinas observadas corresponden sólo a la ventana de tiempo durante la cual se bloqueó la secreción de citoquinas. Si bien describimos la visualización del interferón de citoquina (IFN) gamma en el bazo después de la infección del ratón por la bacteria intracelular Listeria monocytogenes, este método podría adaptarse potencialmente a la visualización de cualquier citoquina en la mayoría de los órganos.
Introducción
La orquestación de una respuesta inmune eficiente contra un patógeno requiere una integración compleja de las señales mostradas por una variedad de células inmunitarias que a menudo se dispersan entre el organismo. Para comunicarse, estas células producen pequeñas proteínas solubles con múltiples funciones biológicas que actúan como inmunomoduladores llamados citoquinas. Las citoquinas controlan el reclutamiento, activación y proliferación de células y, por lo tanto, se sabe que son actores clave en la promoción de las respuestas inmunitarias1. Las respuestas inmunitarias eficaces requieren que las citoquinas se liberen en un patrón espaciotemporal muy organizado que conecte células específicas para inducir señales específicas. Por lo tanto, es crucial estudiar la producción de citoquinas y su señalización in situ, teniendo en cuenta el microambiente en el que se producen las citoquinas.
Listeria monocytogenes (L. monocytogenes) es una bacteria intracelular Gram-positiva utilizada como modelo principal para estudiar las respuestas inmunitarias a patógenos intracelulares en ratones. Una citoquina, IFN gamma (IFN) se produce rápidamente, dentro de las 24 h después de la infección por L. monocytogenes. Es necesario para el aclaramiento de patógenos, ya que los ratones noqueados para IFN son altamente susceptibles a la infección de L. monocytogenes 2. IFN es pleiotrópico y producido por múltiples células después de la infección3. Mientras que el IFN producido por células asesinas naturales (NK) es necesario para la actividad antibacteriana directa4, IFN de otras fuentes han demostrado tener otras funciones. De hecho, nosotros y otros encontramos recientemente que el IFN producido por las células T CD8+ tiene una función específica en la regulación directa de la diferenciación de células T5,6,7. Como tal, entender qué células producen IFN (y en qué microambiente) es crucial para diseccionar su función.
La técnica más común para estudiar la producción de citoquinas se basa en la tinción de citoquinas intracelulares analizada por la citometría de flujo. Este método permite la detección simultánea de múltiples citoquinas combinadas con marcadores de superficie celular dentro de una sola muestra, proporcionando una herramienta extremadamente útil para estudiar la producción de citoquinas. Sin embargo, el uso de la técnica antes mencionada implica la pérdida de cualquier información espacial. Además, la detección de citoquinas a menudo se basa en la reestimulación in vitro para permitir la detección de citoquinas. Como tal, se analiza la capacidad de una célula dada para producir una citoquina, y no necesariamente se correlaciona con la secreción real de citoquinas in situ. Otros métodos utilizan ratones reportero para los que la expresión de proteína fluorescente se correlaciona con la transcripción de citoquinas y permite la visualización en un niveldeuna sola célula 8. Aunque este método puede rastrear la transcripción de citoquinas in situ, hay un número limitado de ratones de citoquinas-reportero disponibles. Además, la transcripción, la traducción y la secreción a veces pueden estar desvinculadas, y las proteínas fluorescentes tienen una vida media diferente a la citoquina que informan, por lo que este método a veces no es adecuado para la visualización in situ de la citoquina.
Aquí, describimos un método para visualizar la producción in situ de citoquinas mediante microscopía confocal a resolución de una sola célula. Esta técnica permite la visualización de la fuente celular y el nicho circundante dentro del tejido. Este protocolo describe específicamente la visualización de la producción de IFN en el bazo de ratones infectados por L. monocytogenes, centrándose aquí en la producción de IFN por células NK y células CD8+ T específicas de antígenos. Sin embargo, puede extenderse y adaptarse a la caracterización de cualquier producción de citoquinas en el contexto de otras situaciones en las que se producen citoquinas como infecciones, inflamación o enfermedades autoinmunes, siempre y cuando la citoquina dirigida pueda conservarse en células por inhibidor del transporte de proteínas intracelular.
Protocolo
Todos los experimentos con ratones estuvieron de acuerdo con la Ley de Procedimientos Científicos del Reino Unido de 1986.
1. Transferencia adoptiva de células CD8+ T específicas antigénicas en ratones
- Aislar la ovalbumina (OVA) específica CD8+ T células (OTI) que expresan proteína fluorescente verde (OTI-GFP) o proteína fluorescente roja (OTI-RFP) de la suspensión de ganglios linfáticos de ratones transgénicos del receptor de células T9,10 con un ratón Kit de aislamiento de celda CD8+ T según las instrucciones de fabricación. Preparar la suspensión celular rompiendo los ganglios linfáticos utilizando un émbolo de jeringa, como se describió anteriormente11.
- Transfiera células OTI-GFP u OTI-RFP (3 x 106 células) a ratones de tipo salvaje C57BL/6 por inyección intravenosa según lo descrito por Cahalan, et al.12. Use ratones que suelen tener entre 6 y 12 semanas de edad.
NOTA: Este paso es opcional y solo es necesario para el seguimiento de células CD8+ T específicas del antígeno.
2. Listeria monocytogenes Infección
- Ampliar L. monocytogenes modificado genéticamente para expresar OVA (LM-OVA)13 a una fase exponencial de crecimiento en la infusión de corazón de caldo a 37 oC bajo agitación suave hasta que el OD600 alcance 0.08–0.1, como se describió anteriormente en referencia 14.
- Inyectar 100 l (volumen máximo a 200 l) de 0,1–0,5 LD50 LM-OVA diluido en solución salina tamponada de fosfato (PBS) mediante inyección intravenosa utilizando una jeringa de insulina de 29 g, en ratones de tipo salvaje C57BL/6 receptores que lleven células OTI-GFP u OTI-RFP cuando se indique.
NOTA: En nuestras manos, 0.1 x LD50 LM-OVA corresponde a 2 x 104 unidades formadoras de colonias (CFU). L. monocytogenes genéticamente modificado para expresar OVA se utiliza para activar las células OTI CD8+ T previamente transferidas, pero se pueden utilizar otras cepas de L. monocytogenes.
3. Tratamiento con Brefeldina A (BFA) para bloquear la secreción de citoquinas
- Inyectar 250 g de BFA en 200 ml de PBS por vía intraperitoneal 6 h antes del sacrificio del ratón utilizando una jeringa de insulina de 29 G.
NOTA: El BFA liofilizado se resuspende primero en dimetil sulfóxido (DMSO) para preparar stock de concentración de 25 mg/ml. El BFA se diluye en PBS a temperatura ambiente (RT) para evitar la cristalización antes de la inyección. La inhibición de la secreción de citoquinas induce la acumulación de IFN en las células. Esto es crucial para la detección de citoquinas.
4. Cosecha del bazo
- Eutanasia a los ratones con una concentración creciente de CO2 seguida de dislocación cervical.
NOTA: Siga las pautas de la institución local para la eutanasia humana de ratones. - Limpiar el abdomen con 70% de etanol, hacer una incisión con tijeras para hacer un corte de 1-2 cm a través de la piel en el flanco izquierdo del ratón, donde se encuentra el bazo. Haga cuidadosamente una incisión en el peritoneo para exponer el bazo y sacarlo con pinzas. Cosecha el bazo, teniendo cuidado de no apretarlo con fórceps o cortarlo para evitar interrumpir la arquitectura del bazo.
5. Fijación del bazo con paraformaldehído (PFA)
- Preparar la solución fijativa mezclando 3,75 ml de PBS y 3,75 ml de 0,2 M de L-lisina. Añadir 21 mg de m-periodato de sodio y mezclar bien. A continuación, agregue 2,5 mL de 4% PFA y 20 l de 12 N NaOH.
NOTA: Utilice la solución fijativa el mismo día y deseche el exceso. No lo guardes. Este paso de fijación es importante si la muestra contiene proteínas fluorescentes como GFP. No utilice PFA que contenga trazas de metanol, ya que desnaturaliza las proteínas fluorescentes.
PRECAUCION: La PFA es tóxica y debe manipularse con precaución. - Sumerja el bazo en el fijador y fije durante un mínimo de 4 h, típicamente de 16 a 20 h a 4oC bajo agitación suave.
- Deseche la solución fijativa y agregue 5 ml de PBS durante 5 minutos a RT bajo agitación suave.
- Sustituya el PBS por 5 ml de incubación de PBS fresco durante 1 h a 4 oC bajo agitación suave.
- Sustituya el PBS por 5 ml de sacarosa al 30%, incubar durante 12-24 h.
NOTA: Este método ayuda a mantener la morfología del tejido. Después de la incubación con solución de sacarosa, el órgano debe hundirse en la parte inferior del pozo.
6. Congelación y seccionamiento
- Coloque el hielo seco en un recipiente grande y coloque un recipiente más pequeño en su interior que contenga alrededor de 50 ml de metanol puro y unos trozos de hielo seco.
- Seque suavemente el bazo con una toallita sin pelusas.
- Coloque el bazo dentro de un molde base que contenga una gota de compuesto de temperatura de corte óptima (OCT) en la parte inferior. Tenga cuidado de no producir burbujas. Agregue OCT sobre el bazo.
- Con fórceps, deposite el molde base en la superficie del metanol, asegurándose de que no toque el Oct.
- Cuando esté congelado, proceda con el seccionamiento.
NOTA: El bazo congelado se puede mantener a -80 oC durante varios meses. -
Secciona el tejido usando un criomicroto.
- Ajuste la temperatura de la cámara en el criostato a -21 oC. Cortar secciones del espesor deseado (generalmente alrededor de 10 m). Este protocolo funciona con espesores de hasta 30 m.
- Recoja secciones sobre diapositivas de microscopio de vidrio (ver la Tabla de Materiales)e inspeccione visualmente.
NOTA: Las secciones se pueden mantener a -80 oC durante varios meses.
7. Tinción inmunofluorescente
- Permita que la sección llegue a RT.
- Dibuje un círculo con un bloqueador de líquidos (por ejemplo, pluma PAP) alrededor de la sección de tejido. Dibuja fuera del Oct o no se pegará.
- Una vez seca, rehidrata la muestra colocando PBS en la sección del tejido durante 5 min.
NOTA: El volumen puesto en la sección depende del tamaño de la sección. Por lo general, usamos de 100 a 300 ol. No deje que las secciones se sequen una vez que se rehidraten. - Enjuague con PBS al menos dos veces para asegurarse de que la sección esté bien adherida a la diapositiva.
-
Agregue la solución de bloqueo a la sección para disminuir la unión no específica de los anticuerpos.
- Preparar la solución de bloqueo de la siguiente manera: PBS con 0,1% Triton X100, 2% de suero de pantorrilla fetal (FCS), bloqueador de receptores Fc de 2,5 g/mL (cd16/32 antiratón). A continuación, agregue entre un 2 y un 5% de suero normal de las especies de cada anticuerpo secundario del panel de tinción.
NOTA: Si los anticuerpos se conjugan/biotinylan directamente, añada un 5% de suero normal de las especies de cada anticuerpo primario. Si un anticuerpo primario y uno de los anticuerpos secundarios son de la misma especie (por ejemplo, anticuerpo primario criado en conejo sin efecto y conejo secundario antirrárico), no utilice las especies séricas normales, ya que aumentará la señal de fondo. - Retire suavemente el PBS de la sección por aspiración y agregue 100 sl de la solución de bloqueo por sección de muestra. Incubar en una cámara húmeda cubierta durante un mínimo de 1 h a RT.
- Preparar la solución de bloqueo de la siguiente manera: PBS con 0,1% Triton X100, 2% de suero de pantorrilla fetal (FCS), bloqueador de receptores Fc de 2,5 g/mL (cd16/32 antiratón). A continuación, agregue entre un 2 y un 5% de suero normal de las especies de cada anticuerpo secundario del panel de tinción.
-
Mancha con anticuerpos primarios.
- Diluir los anticuerpos primarios a la concentración óptima en la solución de bloqueo. La concentración general de anticuerpos de punto de partida es de 5 g/ml, pero debe optimizarse para cada anticuerpo y tejido.
NOTA: Si los anticuerpos se conjugan directamente, centrifugar la mezcla de anticuerpos a 17.135 x g (13.500 rpm) durante 15 minutos a 4 oC antes de usarla. Los fluoróforos pueden precipitarse. Este paso pellet los precipitados y así evitar la deposición inespecífica de los anticuerpos precipitados en la diapositiva. - Sustituya la solución de bloqueo por la mezcla de anticuerpos primarios para cada muestra.
- Incubar durante 4 h a RT o durante la noche (OVN) a 4oC en una cámara húmeda cubierta.
- Diluir los anticuerpos primarios a la concentración óptima en la solución de bloqueo. La concentración general de anticuerpos de punto de partida es de 5 g/ml, pero debe optimizarse para cada anticuerpo y tejido.
-
Realice el lavado.
- Prepare Wash Buffer agregando 2% FCS a PBS.
- Lavar 4 veces con tampón de lavado: uno rápido (sin incubación), uno para 10 min y dos por 5 min. A continuación, realice un lavado final con PBS durante 5 min.
-
Mancha con anticuerpos secundarios.
- Diluir los anticuerpos secundarios de interés a la concentración óptima en la solución de bloqueo. Centrifugar la mezcla, como se describe para los anticuerpos primarios.
- Retire la solución de lavado final. Añadir la mezcla secundaria de anticuerpos en la parte superior de la sección e incubar durante 1-4 h a RT en una cámara húmeda cubierta.
- Lavar 4 veces con tampón de lavado: uno rápido (sin incubación), uno para 10 min y dos por 5 min. A continuación, realice un lavado final con PBS durante 5 min.
- Retire la solución de lavado final. Deje que el PBS se evapore pero no seque en exceso la sección. Coloque una gota del medio de montaje encima de la muestra y coloque cuidadosamente el vidrio de la cubierta encima de ella. El medio de montaje debe recuperar toda la sección. Dejar que la polimerasa OVN en RT protegido de la luz.
NOTA: Dibuje un círculo alrededor de la sección en el reverso de la diapositiva antes de aplicar el medio de montaje. Una vez que se aplica el medio de montaje, el tejido puede llegar a ser difícil de ver. - Almacene las diapositivas en la oscuridad a 4 oC hasta que estén listas para la imagen.
8. Imágenes y análisis
- Realice imágenes de la tinción con un microscopio confocal.
NOTA: En este protocolo, se utilizó un microscopio de escaneo láser espectral invertido (ver la Tabla de Materiales),junto con los objetivos 10x/NA 0.40 o 60x/NA 1.4 (para el análisis de la localización subcelular de citoquinas). En la Tabla de Materialesse muestran longitudes de onda de excitación y emisión para cada fluoróforo y proteína fluorescente. - Realizar análisis y cuantificación según sea necesario utilizando un software de procesamiento de imágenes (por ejemplo, Imaris o Fiji).
Resultados
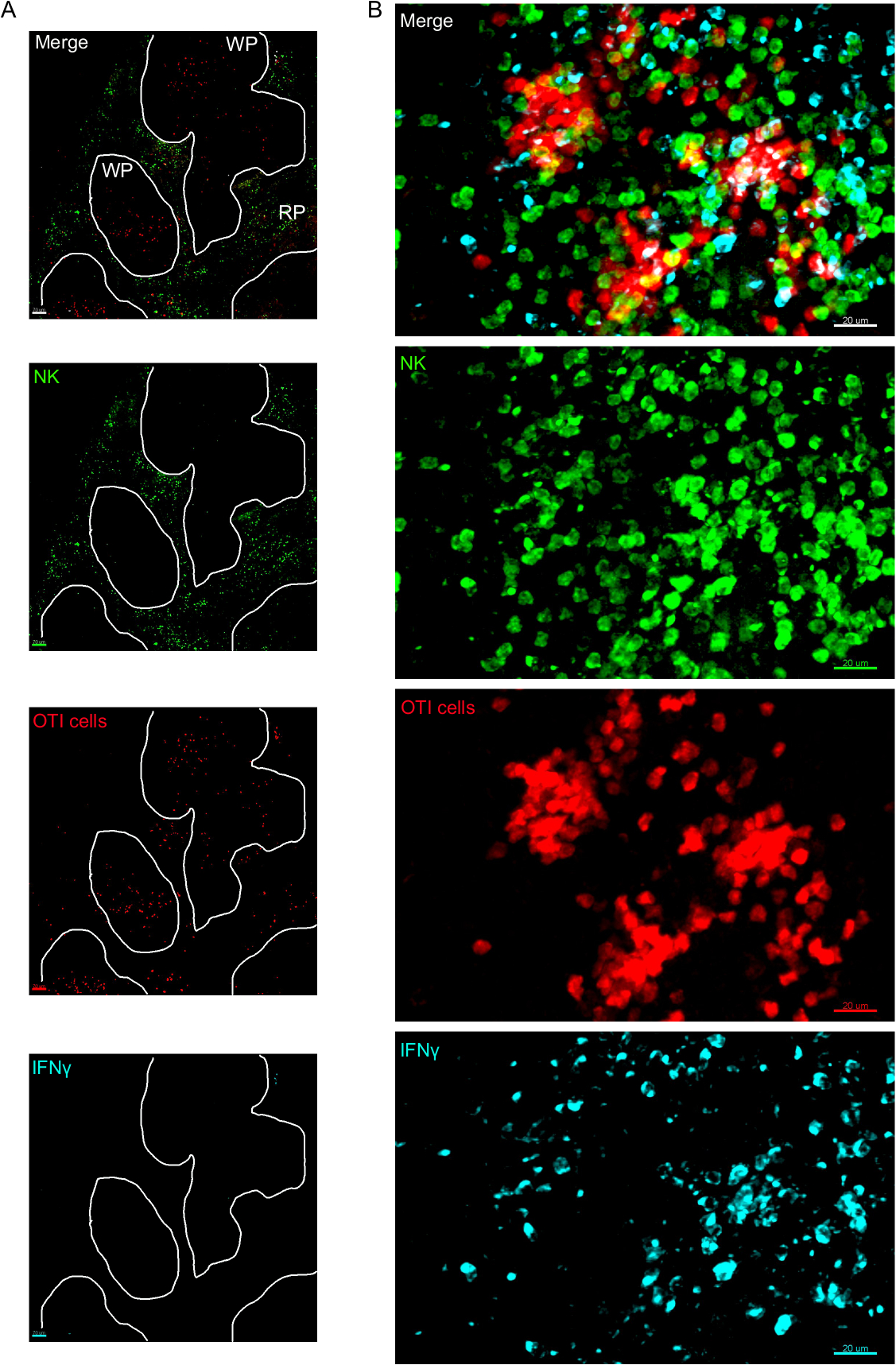
El IFN producido dentro de las primeras 24 h después de la infección por Listeria monocytogenes es fundamental para controlar la propagación de este patógeno. Usando este protocolo, podemos visualizar no sólo qué células están produciendo IFN, sino también si se encuentran en un microambiente específico. Para ayudarnos a delinear la arquitectura del bazo, etiquetamos las células que se sabe que tienen una ubicación particular dentro del bazo. El marcador F4/80 etiqueta todos los macrófagos y resalta la pulpa roja. El marcador B220 etiqueta las células B y resalta los folículos celulares B que rodean la zona de la célula T. El marcador CD169 etiqueta los macrófagos de zona marginal, que rodean la pulpa blanca (Figura1). La mayoría de las células OTI, ya sean que expresan IFN o no, están presentes en la pulpa blanca y, como tal, todas las imágenes son las de la pulpa blanca, a menos que se indique.
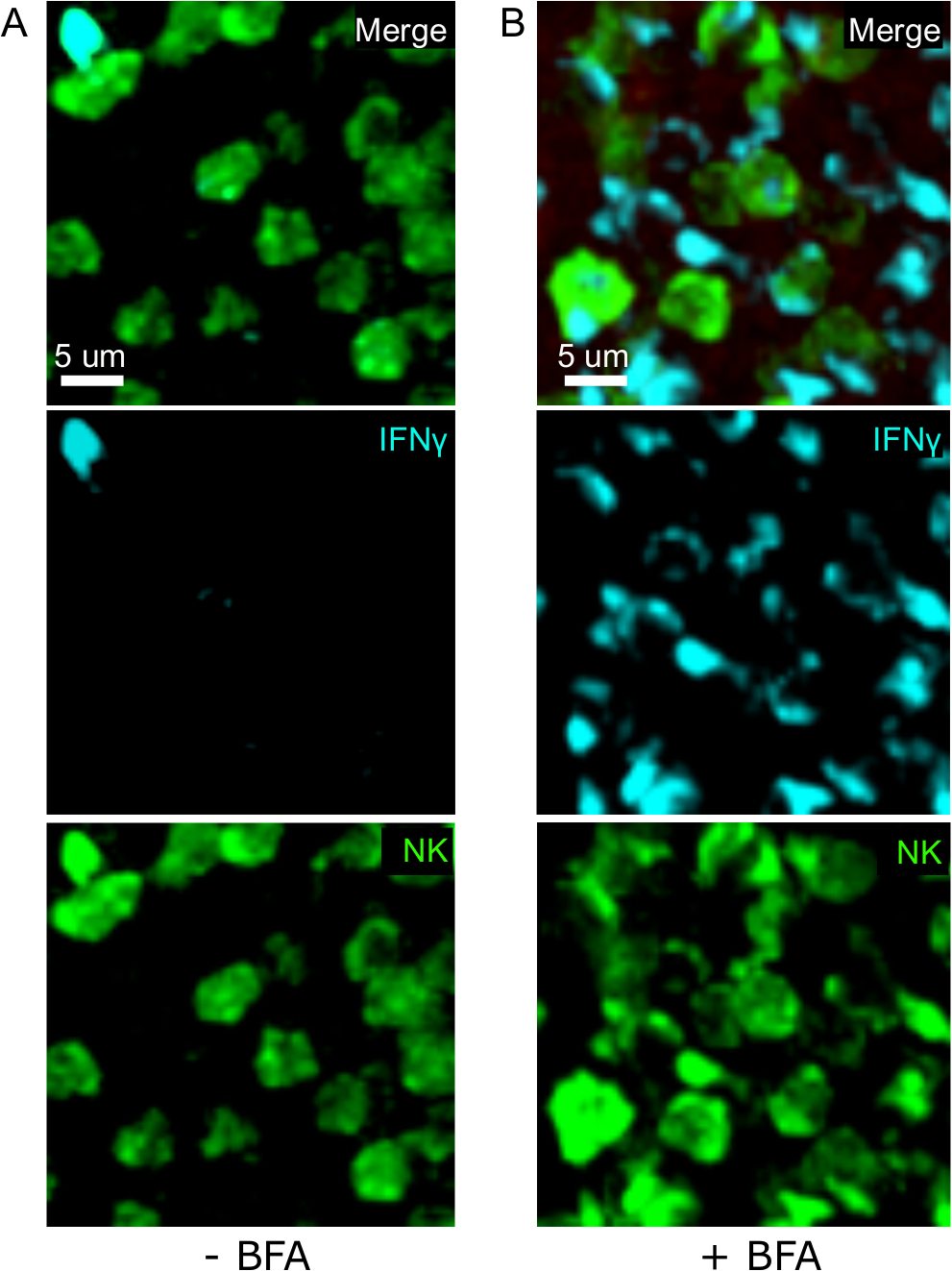
Un paso crítico en este protocolo es el uso de BFA para inhibir la secreción de citoquinas. De hecho, la detección de IFN por células NK se viomuy afectada cuando los ratones no fueron tratados con BFA (Figura 2). Usando nuestro protocolo, podríamos encontrar que al menos dos tipos de células producen IFN-24 h después de la infección —células NK y células CD8+ T específicas del antígeno (Figura3), de manera similar a lo que se ha encontrado anteriormente por la citometría de flujo3.
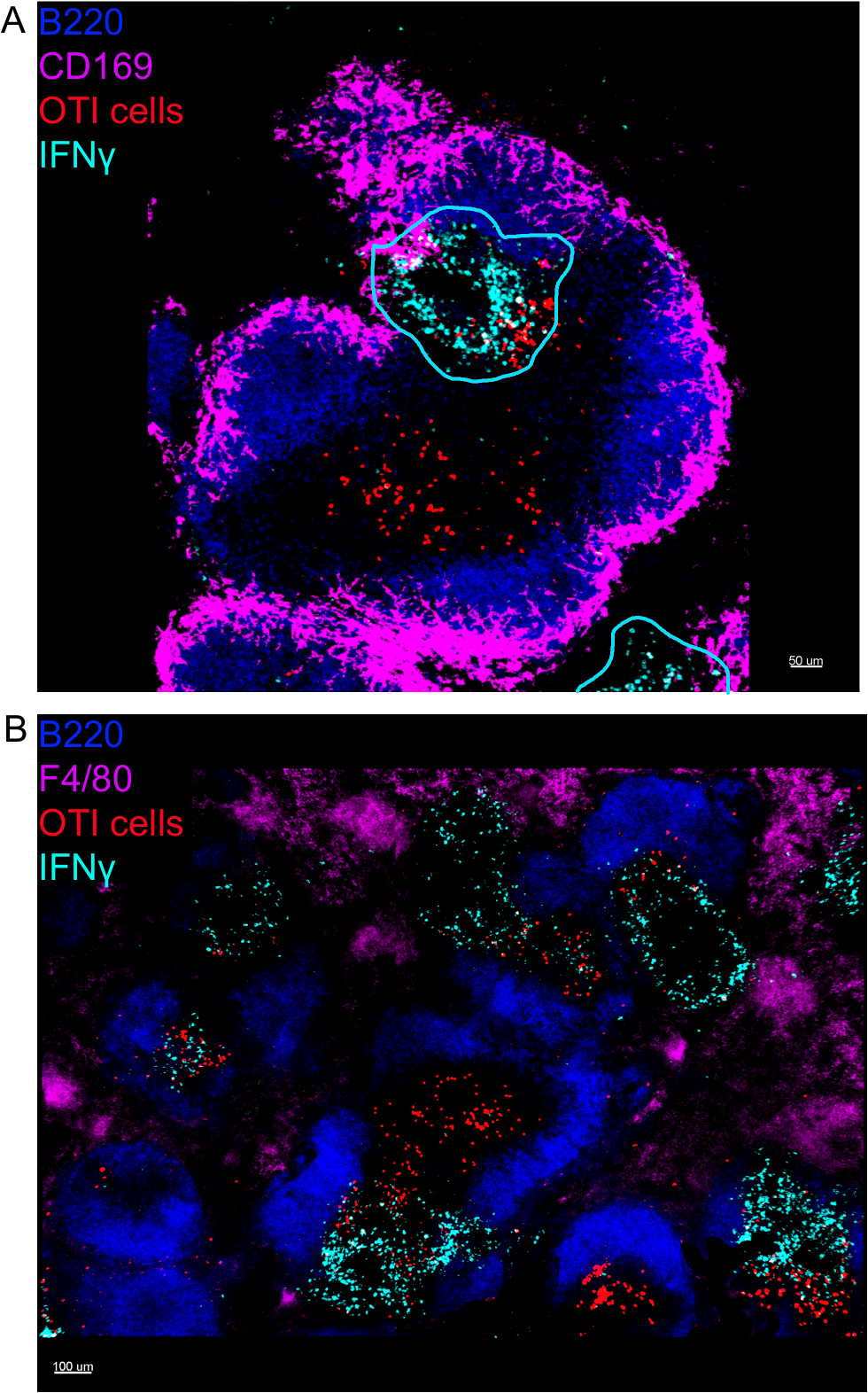
La imagen in situ de las células productoras de IFN reveló que la producción de IFN no se extiende por todo el bazo, sino que se concentra en áreas discretas (Figura4). De hecho, descubrimos que las células T se activaban a lo largo del bazo (resaltado por la agrupación de células T), y esto no necesariamente se correlacionaba con la producción de IFN. Una explicación probable es que la producción de IFN está restringida a la ubicación de las células infectadas15,16y la activación de células T, representadas por la agrupación en clústeres, pueden ser compatibles tanto con las infectadas (IFN positivo) como por las no infectadas (IFN-positivo) negativos) células que presentan antígenos. Se requerirán otras manchas para identificar la ubicación exacta y obtener una indicación del mecanismo que restringe la producción de IFN a esta área y su relación con la transferencia de antígenos. Curiosamente, encontramos que las células T activadas, agrupadas y específicas del antígeno se encuentran a lo largo de la pulpa blanca del bazo, pero producen IFN sólo en regiones donde las células NK están coexistiendo con ellos (Figura5). Como tal, la presencia de células NK delinea un microambiente específico en la pulpa blanca, en el que las células T agrupadas producen IFN en lugar de células T agrupadas en la otra parte de la pulpa blanca. Esto sugiere que la activación de la célula T no es suficiente para dictar la producción de IFN en este momento.
Otra característica interesante destacada por nuestro protocolo es la diferente localización subcelular de IFN en NK versus CD8+ T células5. Tal y como se muestra en de la Figura6, mientras que la localización IFN en las células NK se difunde en el citosol, las células CD8+ T a menudo reclutan el IFN hacia otra célula T.

Figura 1: Marcadores que resaltan la arquitectura del bazo. Los ratones se infectaron con 2 x 104 CFU LM-OVA y eutanasia 24 h después de la infección. El bazo fue explantado y procesado como se describe en el protocolo. (A) Se tiñó las secciones para las células NK (anti-NCR1 seguidade de IgG-FITC anticabras; verde), células OTI-RFP (roja) y macrófagos (anti-F4/80-APC; magenta). RP - Pulpa Roja; WP - Pulpa Blanca. Barra de escala a 200 m. (B) Se teñieron las secciones para las células B (azul anti-B220-Pacífico; Células Azules), células OTI-GFP (señal GFP mostrada en rojo) y macrófagos de zona marginal (anti-CD169-Alexa647; magenta). RP - Pulpa Roja; FBF - Folículo celular B; TZ - Zona de células T. Barra de escala a 50 m. Se trata de una imagen representativa de 3 experimentos independientes (N .4). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2: El tratamiento con BFA permite la detección in situ de ifN intracelular. La producción de no está restringida a áreas específicas del bazo Los ratones fueron infectados con 2 x 104 CFU LM-OVA y tratados con BFA (A) o no tratados (B) después de 18 h. Los ratones fueron eutanetaizados 24 horas después de la infección. El bazo fue explantado y procesado como se describe en el protocolo. Las secciones se teñían para las células NK (anti-NCR1 seguido de igG-FITC anticabras; verde), células OTI-RFP (roja) e IFN (anti-IFN-BV421; cian). Barra de escala a 5 m. Se trata de una imagen representativa de las áreas ricas en células NK de 3 experimentos independientes (N.o 3). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3: Células productoras de IFN en el bazo. Los ratones se infectaron con 2 x 104 CFU LM-OVA cuando se indica y trataron con BFA después de 18 h. Los ratones fueron eutanetamos 24 h después de la infección. El bazo fue explantado y procesado como se describe en el protocolo. Las secciones se teñían para las células NK (anti-NCR1 seguido de igG-FITC anticabras; verde), células OTI-RFP (roja) e IFN (anti-IFN-BV421; cian). (A) Imagen representativa de un bazo de un ratón ingenuo no infectado para demostrar la ausencia de tinción inespecífica de IFN. Las líneas blancas delinean la pulpa blanca. WP - Pulpa blanca; RP - Pulpa roja. (B) Imagen representativa de la pulpa blanca del bazo de un ratón infectado por LM-OVA, mostrando la invasión de células NK a la pulpa blanca y la producción de IFN por células NK, células OTI y células no etiquetadas. Las imágenes son representativas de 4 experimentos independientes (N .4). Barras de escala a 70 m (A); y 20 m (B). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 4: La producción de IFN está restringida a áreas específicas en el bazo después de la infección por LM-OVA. Los ratones fueron infectados con 2 x 104 CFU LM-OVA y tratados con BFA después de 18 h. Los ratones fueron eutanasiados 24 h después de la infección. El bazo fue explantado y procesado como se describe en el protocolo. Todas las secciones se teñieron para las células B (B220-Pacific Blue Ab, Blue) y IFN (anti-IFN-biotina seguida de streptavidina-PE; cian). Células OTI-GFP (señal GFP mostrada en rojo). Las líneas cian corresponden a áreas de alta producción de IFN. Se trata de imágenes representativas de 4 experimentos independientes (N.o 4). (A) Se han manchado las secciones para los macrófagos de zona marginal (anti-CD169-Alexa 647, magenta). Barra de escala a 50 m. (B) Se tiñeron las secciones para todos los macrófagos (F4/80). Barra de escala a 100 m. Por favor, haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 5: La producción de IFN por células OTI activadas se produce en un microambiente específico. Los ratones fueron infectados con 2 x 104 CFU LM-OVA y tratados con BFA después de 18 h. Los ratones fueron eutanasiados 24 h después de la infección. Los bazos fueron explantados y procesados como se describe en el protocolo. Las secciones se teñían para las células NK (anti-NCR1 seguido de igG-FITC anticabras; verde), células OTI-RFP (roja) e IFN (anti-IFN-BV421; cian). Las líneas verdes y rojas resaltan las zonas de celda NK y OTI, respectivamente. La flecha blanca indica ejemplos de clústeres de celdas T que no producen IFN. Ejemplos de flechas verdes de clústeres de células T que producen IFN. Barra de escala a 100 m. Se trata de una imagen representativa de cuatro experimentos independientes (N .4). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 6: Localización subcelular de IFN en células NK y células T. Los ratones fueron infectados con 2 x 104 CFU LM-OVA y tratados con BFA después de 18 h. Los ratones fueron eutanasiados 24 h después de la infección. El bazo fue explantado y procesado como se describe en el protocolo. Todas las secciones fueron manchadas para IFN (anti-IFN-BV421; cyan). Las líneas blancas delinean los bordes de las celdas y las flechas blancas muestran la direccionalidad de la secreción. Se trata de una imagen representativa de dos experimentos independientes (N.o 5). (A)- Las celdas OTI-RFP se muestran en rojo. Barra de escala de5 m. (B ) Se tiñeron las secciones para las células NK (anti-NCR1 seguida de IgG-FITC anticabra; verde. Barra de escala de 2 m. Por favor, haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Discusión
En este manuscrito, presentamos un método para visualizar la producción de IFN en el bazo después de la infección de L. monocytogenes en ratones. Este protocolo es simple y se puede adaptar a otros tejidos y desencadenantes de citoquinas, pero los siguientes aspectos tienen que ser considerados. Las células a menudo secretan rápidamente las citoquinas que producen, y las citoquinas son rápidamente recogidas por las células vecinas. Como tal, es difícil detectar citoquinas in situ. Un método común para volver a iniciar rápidamente la producción de citoquinas es reestimular las células ex vivo seguidas de la detección de citoquinas en los medios mediante un ensayo inmunoabsorbente ligado a enzimas. En este contexto, se pierde cualquier información sobre la localización espacial de las células productoras de citoquinas. Además, la producción de citoquinas después de la reestimulación no refleja necesariamente si las citoquinas se producen y secretan in vivo, sino que indica la capacidad de una población celular determinada para producir citoquinas. Por lo tanto, ambos métodos proporcionarán información diferente y uno debe considerar qué información es más valiosa para su experimento.
Con el fin de detectar citoquinas intracelulares, nuestro método utiliza un inhibidor de transporte de proteínas intracelular para atrapar citoquinas dentro de las células y aumentar la detección de señales. Sin embargo, es importante tener en cuenta que estos inhibidores afectan el transporte normal de proteínas desde el retículo endotelial (RE) al aparato Golgi y a la vesícula secretor que deteriora su liberación, lo que podría causar toxicidad. Como consecuencia, BFA, u otro inhibidor, debe utilizarse durante un corto período de tiempo, por lo general no más de unas pocas horas. Por lo tanto, es importante encontrar el equilibrio adecuado entre la dosis del inhibidor y el tiempo de tratamiento con el fin de optimizar el nivel de citoquinas atrapadas dentro de la célula sin causar efectos citotóxicos graves. Estas variables pueden diferir entre las citoquinas y la vía de administración para el BFA. En nuestro modelo de infección, el BFA se administró por vía intraperitoneal con el fin de proporcionar una rápida dispersión sistémica, pero también se puede administrar por vía intravenosa.
Los inhibidores de transporte de proteínas intracelulares más utilizados son el BFA, utilizado aquí, y la monensina (MN). Estos inhibidores se utilizan a menudo indistintamente para acumular y estudiar la producción de citoquinas, pero tienen ligeras diferencias en sus mecanismos de acción. MN inhibe el transporte de proteínas dentro del aparato Golgi, por lo tanto, la acumulación de proteínas en el Golgi17 mientras que el BFA previene el reclutamiento de proteínas coatómeros-I, inhibiendo el movimiento retrógrado de las proteínas al retículo endoplasmático (ER) y promoviendo así la acumulación de citoquinas en los ERes18. Como tal, elegir el mejor inhibidor de transporte de proteínas intracelular dependerá de diferentes factores, como la citoquina que se detectará. Por ejemplo, se ha demostrado en la tinción intracelular inducida por lipopolisacáridos de monocitos que el BFA es más eficiente para medir las citoquinas IL-1, IL-6 y TNF que MN19.
Este protocolo implica la visualización de la citoquina in situ por microscopía confocal y, por lo tanto, sólo hay un número limitado de marcadores que se pueden utilizar para estudiar las células productoras de citoquinas y su microambiente. También es necesario tener en cuenta que los inhibidores del transporte de proteínas como bfA o MN perturban la expresión normal de varias proteínas y, por lo tanto, su uso al estudiar la expresión simultánea de ciertos marcadores de superficie celular de activación debe abordarse Cuidadosamente. Por ejemplo, BFA pero no MN bloquea la expresión de CD69 en linfocitos murinos20. A pesar de esta limitación, la imagen confocal permite la localización subcelular de las citoquinas, así como la dirección de la secreción de citoquinas dentro de la célula. Los datos generados utilizando este protocolo sugieren que las células NK tienden a secretar IFN-y en un patrón difuso, mientras que las células CD8+ T parecen dirigir la secreción de IFN hacia otras células CD8+ T que están en interacción directa con ellas5.
Para concluir, este protocolo es adecuado para visualizar una variedad de citoquinas in situ e identificar las células productoras y su microambiente después de muchos desencadenantes como la infección o la autoinmunidad. La información obtenida es fundamental para comprender la importancia de la orquestación espacial in vivo de los diferentes tipos de células y la citoquina que producen, necesaria para una respuesta inmune eficiente.
Divulgaciones
Los autores no tienen nada que revelar.
Agradecimientos
Agradecemos al personal del Centro de Imágenes del Instituto Kennedy por su asistencia técnica con imágenes. Este trabajo fue apoyado por subvenciones del Kennedy Trust (a A.G.), y del Consejo de Investigación de Biotecnología y Ciencias Biológicas (BB/R015651/1 a A.G.).
Materiales
| Name | Company | Catalog Number | Comments |
| Brefeldin A | Cambridge bioscience | CAY11861 | |
| Paraformaldehyde | Agar scientific | R1018 | |
| L-Lysin dihydrochloride | Sigma lifescience | L5751 | |
| Sodium meta-periodate | Thermo Scientific | 20504 | |
| D(+)-saccharose | VWR Chemicals | 27480.294 | |
| Precision wipes paper Kimtech science | Kimberly-Clark Professional | 75512 | |
| O.C.T. compound, mounting medium for cryotomy | VWR Chemicals | 361603E | |
| Fc block, purified anti-mouse CD16/32, clone 93 | Biolegend | 101302 | Antibody clone and Concentration used: 2.5 mg/ml |
| Microscope slides - Superfrost Plus | VWR Chemicals | 631-0108 | |
| anti-CD169 - AF647 | Biolegend | 142407 | Antibody clone and Concentration used: clone 3D6.112 1.6 mg/ml Excitation wavelength: 650 Emission wavelength: 65 |
| anti-F4/80 - APC | Biolegend | 123115 | Antibody clone and Concentration used: clone BM8 2.5 mg/ml Excitation wavelength: 650 Emission wavelength: 660 |
| anti-B220 - PB | Biolegend | 103230 | Antibody clone and Concentration used: clone RA3-6B2 1.6 mg/mL Excitation wavelength: 410 Emission wavelength: 455 |
| anti-IFNg - biotin | Biolegend | 505804 | Antibody clone and Concentration used: clone XMG1.2 5 mg/mL |
| anti-IFNg - BV421 | Biolegend | 505829 | Antibody clone and Concentration used: clone XMG1.2 5 mg/mL Excitation wavelength: 405 Emission wavelength: 436 |
| anti-Nkp46/NCRI | R&D Systems | AF2225 | Antibody clone and Concentration used: goat 2.5 mg/mL |
| anti-goat IgG-FITC | Novusbio | NPp 1-74814 | Antibody clone and Concentration used: 1 mg/mL Excitation wavelength: 490 Emission wavelength: 525 |
| Streptavidin - PE | Biolegend | 405203 | Antibody clone and Concentration used: 2.5 mg/mL Excitation wavelength: 565 Emission wavelength: 578 |
| Streptavidin - FITC | Biolegend | 405201 | Antibody clone and Concentration used: 2.5 mg/mlL Excitation wavelength: 490 Emission wavelength: 525 |
| Fluoromount G | SouthernBiotech | 0100-01 | |
| Cover glasses 22 mm x 40 mm | Menzel-Glazer | 12352128 | |
| Liquid blocker super PAP PEN mini | Axxora | CAC-DAI-PAP-S-M | |
| Imaris - Microscopy Image Analysis Software | Bitplane | ||
| Confocal microscope - Olympus FV1200 Laser scanning microscope | Olympus | ||
| Cryostat - CM 1900 UV | Leica | ||
| Base mould disposable | Fisher Scientific UK Ltd | 11670990 | |
| PBS 1x | Life Technologies Ltd | 20012068 | |
| BHI Broth | VWR Brand | 303415ZA | |
| GFP | Excitation wavelength: 484 Emission wavelength: 507 | ||
| RFP | Excitation wavelength: 558 Emission wavelength: 583 | ||
| Insulin syringe, with needle, 29 G | VWR International | BDAM324824 | |
| C57BL/6 wild type mice | Charles River |
Referencias
- Iwasaki, A., Medzhitov, R. Control of adaptive immunity by the innate immune system. Nature Immunology. 16 (4), 343-353 (2015).
- Harty, J. T., Bevan, M. J. Specific immunity to Listeria monocytogenes in the absence of IFN gamma. Immunity. 3 (1), 109-117 (1995).
- Kubota, K., Kadoya, Y. Innate IFN-gamma-producing cells in the spleen of mice early after Listeria monocytogenes infection: importance of microenvironment of the cells involved in the production of innate IFN-gamma. Frontiers in Immunology. 2 (26), (2011).
- Dunn, P. L., North, R. J. Early gamma interferon production by natural killer cells is important in defense against murine listeriosis. Infection and Immunity. 59 (9), 2892-2900 (1991).
- Krummel, M. F., et al. Paracrine costimulation of IFN-gamma signaling by integrins modulates CD8+ T cell differentiation. Proceedings of the National Academy of Sciences of the United States of America. 115 (45), 11585-11590 (2018).
- Curtsinger, J. M., Agarwal, P., Lins, D. C., Mescher, M. F. Autocrine IFN-gamma promotes naive CD8+ T cell differentiation and synergizes with IFN-alpha to stimulate strong function. Journal of Immunology. 189 (2), 659-668 (2012).
- Hosking, M. P., Flynn, C. T., Whitton, J. L. Antigen-specific naive CD8++ T cells produce a single pulse of IFN-gamma in vivo within hours of infection, but without antiviral effect. Journal of Immunology. 193 (4), 1873-1885 (2014).
- Croxford, A. L., Buch, T. Cytokine reporter mice in immunological research: perspectives and lessons learned. Immunology. 132 (1), 1-8 (2011).
- Gerard, A., et al. Secondary T cell-T cell synaptic interactions drive the differentiation of protective CD8++ T cells. Nature Immunology. 14 (4), 356-363 (2013).
- Engelhardt, J. J., et al. Marginating dendritic cells of the tumor microenvironment cross-present tumor antigens and stably engage tumor-specific T cells. Cancer Cell. 21 (3), 402-417 (2012).
- Matheu, M. P., Cahalan, M. D. Isolation of CD4+ T cells from mouse lymph nodes using Miltenyi MACS purification. Journal of Visualized Experiments. (9), 409 (2007).
- Matheu, M. P., Parker, I., Cahalan, M. D. Dissection and 2-photon imaging of peripheral lymph nodes in mice. Journal of Visualized Experiments. (7), 265 (2007).
- Pope, C., et al. Organ-specific regulation of the CD8+ T cell response to Listeria monocytogenes infection. Journal of Immunology. 166 (5), 3402-3409 (2001).
- Jones, G. S., D'Orazio, S. E. Listeria monocytogenes: cultivation and laboratory maintenance. Current Protocols in Microbiology. 31, 1-7 (2013).
- Kang, S. J., Liang, H. E., Reizis, B., Locksley, R. M. Regulation of hierarchical clustering and activation of innate immune cells by dendritic cells. Immunity. 29 (5), 819-833 (2008).
- Chang, S. R., et al. Characterization of early gamma interferon (IFN-gamma) expression during murine listeriosis: identification of NK1.1+ CD11c+ cells as the primary IFN-gamma-expressing cells. Infection and Immunity. 75 (3), 1167-1176 (2007).
- Mollenhauer, H. H., Morre, D. J., Rowe, L. D. Alteration of intracellular traffic by monensin; mechanism, specificity and relationship to toxicity. Biochimica et Biophysica Acta. 1031 (2), 225-246 (1990).
- Helms, J. B., Rothman, J. E. Inhibition by brefeldin A of a Golgi membrane enzyme that catalyses exchange of guanine nucleotide bound to ARF. Nature. 360 (6402), 352-354 (1992).
- Schuerwegh, A. J., Stevens, W. J., Bridts, C. H., De Clerck, L. S. Evaluation of monensin and brefeldin A for flow cytometric determination of interleukin-1 beta, interleukin-6, and tumor necrosis factor-alpha in monocytes. Cytometry. 46 (3), 172-176 (2001).
- Nylander, S., Kalies, I. Brefeldin A, but not monensin, completely blocks CD69 expression on mouse lymphocytes: efficacy of inhibitors of protein secretion in protocols for intracellular cytokine staining by flow cytometry. Journal of Immunology Methods. 224 (1-2), 69-76 (1999).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados