
Method Article
Visualización de la dinámica de acoplamiento de actina y microtúbulos in vitro mediante microscopía de fluorescencia de reflexión interna total (TIRF)
En este artículo
Resumen
Este protocolo es una guía para visualizar actina dinámica y microtúbulos utilizando un ensayo de microscopía de fluorescencia interna total (TIRF) in vitro .
Resumen
Tradicionalmente, los citoesqueletos de actina y microtúbulos se han estudiado como entidades separadas, restringidas a regiones o procesos celulares específicos, y reguladas por diferentes conjuntos de proteínas de unión únicas para cada polímero. Muchos estudios ahora demuestran que la dinámica de ambos polímeros citoesqueléticos está entrelazada y que esta diafonía es necesaria para la mayoría de los comportamientos celulares. Ya se han identificado varias proteínas involucradas en las interacciones actina-microtúbulos (es decir, Tau, MACF, GAS, forminas y más) y están bien caracterizadas con respecto a la actina o los microtúbulos solos. Sin embargo, relativamente pocos estudios mostraron ensayos de coordinación actina-microtúbulo con versiones dinámicas de ambos polímeros. Esto puede ocluir los mecanismos de enlace emergentes entre la actina y los microtúbulos. Aquí, una técnica de reconstitución in vitro basada en microscopía de fluorescencia interna total (TIRF) permite la visualización de la dinámica de la actina y los microtúbulos a partir de la única reacción bioquímica. Esta técnica preserva la dinámica de polimerización del filamento de actina o de los microtúbulos individualmente o en presencia del otro polímero. La proteína Tau disponible comercialmente se utiliza para demostrar cómo cambian los comportamientos de actina-microtúbulos en presencia de una proteína de reticulación citoesquelética clásica. Este método puede proporcionar información funcional y mecanicista confiable sobre cómo las proteínas reguladoras individuales coordinan la dinámica actina-microtúbulo a una resolución de filamentos individuales o complejos de orden superior.
Introducción
Históricamente, la actina y los microtúbulos se han visto como entidades separadas, cada una con su propio conjunto de proteínas reguladoras, comportamientos dinámicos y ubicaciones celulares distintas. Abundante evidencia ahora demuestra que los polímeros de actina y microtúbulos participan en mecanismos funcionales de diafonía que son esenciales para ejecutar numerosos procesos celulares, incluida la migración, el posicionamiento del huso mitótico, el transporte intracelular y la morfología celular 1,2,3,4. Los diversos comportamientos coordinados que subyacen a estos ejemplos dependen de un intrincado equilibrio de factores de acoplamiento, señales y propiedades físicas. Sin embargo, los detalles moleculares que sustentan estos mecanismos aún se desconocen en gran medida porque la mayoría de los estudios se centran en un solo polímero citoesquelético en un momento 1,2,5.
La actina y los microtúbulos no interactúan directamente 6,7,8. La dinámica coordinada de la actina y los microtúbulos observada en las células está mediada por factores adicionales. Se han identificado muchas proteínas que se cree que regulan la diafonía actina-microtúbulo y sus actividades están bien caracterizadas con respecto a cualquiera de los polímeros citoesqueléticossolos 1,2. La creciente evidencia sugiere que este enfoque de polímero único ha ocultado las funciones duales de algunas de las proteínas / complejos que permiten eventos de acoplamiento actina-microtúbulo 7,8,9,10,11,12,13. Los experimentos en los que ambos polímeros están presentes son raros y a menudo definen mecanismos con un solo polímero dinámico y una versión estática estabilizada de los otros 6,8,9,10,11,14,15,16,17,18 . Por lo tanto, se necesitan métodos para investigar las propiedades emergentes de las proteínas de coordinación actina-microtúbulo que solo pueden entenderse completamente en sistemas experimentales que emplean ambos polímeros dinámicos.
La combinación de enfoques de etiquetado directo de proteínas, etiquetas de afinidad codificadas genéticamente y microscopía de fluorescencia de reflexión interna total (TIRF) se ha aplicado con gran éxito en sistemas de reconstitución biomimética 19,20,21,22,23. Muchos esquemas de abajo hacia arriba no contienen todos los factores que regulan las proteínas en las células. Sin embargo, la tecnología de "bioquímica en un vidrio de cubierta" ha refinado muchos mecanismos de la dinámica de la actina y los microtúbulos a altas escalas espaciales y temporales, incluidos los componentes necesarios para el ensamblaje o desmontaje de polímeros y el movimiento de proteínas motoras 5,12,23,24,25,26,27 . Aquí se describe un enfoque de filamento único de componente mínimo para investigar el acoplamiento actina-microtúbulo in vitro. Este protocolo se puede utilizar con proteínas purificadas disponibles comercialmente o altamente puras, proteínas marcadas fluorescentemente, cámaras de perfusión y extenderse a esquemas más complicados que contienen extractos celulares o sistemas sintéticos. Aquí, la proteína Tau disponible comercialmente se utiliza para demostrar cómo cambia la dinámica citoesquelética en presencia de una proteína de acoplamiento actina-microtúbulo, pero puede ser sustituida por otros supuestos factores de coordinación actina-microtúbulo. La principal ventaja de este sistema sobre otros enfoques es la capacidad de monitorear simultáneamente la dinámica de múltiples polímeros citoesqueléticos en una reacción. Este protocolo también proporciona a los usuarios ejemplos y herramientas simples para cuantificar los cambios en los polímeros citoesqueléticos. Por lo tanto, los usuarios del protocolo producirán datos confiables, cuantitativos y de resolución de un solo filamento para describir los mecanismos que subyacen a cómo las diversas proteínas reguladoras coordinan la dinámica actina-microtúbulo.
Protocolo
1. Lavado de las fundas
NOTA: Lavar (24 mm x 60 mm, #1.5) los cubrehojas según Smith et al., 201328.
- Coloque las fundas en un contenedor de plástico deslizante.
- Sumergir los cubrehojas secuencialmente en las siguientes soluciones y sonicar durante 30-60 min, enjuagando con ddH2O 10 veces entre cada solución: ddH2O con una gota de jabón para platos; 0,1 M KOH. Almacene los cubrehojas en etanol al 100% durante un máximo de 6 meses.
NOTA: No toque superficies de vidrio con los dedos sin amor. Use fórceps en su lugar.
2. Recubrimiento limpio (24 mm x 60 mm, #1.5) de fundas con mPEG- y biotina-PEG-silano
NOTA: Este protocolo utiliza específicamente un sistema de biotina-estreptavidina para posicionar la actina y los microtúbulos dentro del plano de imágenes TIRF. Se pueden utilizar otros recubrimientos y sistemas (por ejemplo, anticuerpos, poli-L-lisina, miosina NEM, etc.).
- Descongelar las alícuotas de peg-silano y biotina-PEG-silano en polvos.
- Disolver polvos de PEG en etanol al 80% (pH 2.0) para generar soluciones de recubrimiento de 10 mg/mL de mPEG-silano y 2-4 mg/mL de biotina-PEG-silano, justo antes del uso.
NOTA: Los polvos de PEG a menudo aparecen disueltos, pero pueden no estar a nivel microscópico. La resuspensión adecuada toma ~ 1-2 minutos con pipeteo constante. Se anima a los usuarios a pipetear 10 veces más después de la aparición de la disolución del polvo.
PRECAUCIÓN: Use guantes para proteger la piel del HCl concentrado cuando produzca etanol al 80% (pH 2.0). - Retire el estuche limpio (24 mm x 60 mm, #1.5) del almacenamiento de etanol con fórceps. Secar con gas nitrógeno y almacenar en una placa de Petri limpia.
- Cubra los cubrehojas con 100 μL de solución de recubrimiento: una mezcla de 2 mg/ml de mPEG-silano (MW 2.000) y 0,04 mg/ml de biotina-PEG-silano (MW 3.400) en etanol al 80% (pH 2,0).
NOTA: Para un recubrimiento disperso (recomendado) use 2 mg/mL mPEG-silano y 0.04 mg/mL de biotina-PEG-silano. Para el recubrimiento denso use 2 mg/mL mPEG-silano, 4 mg/mL biotina-PEG-silano. - Incubar los recubiertos a 70 °C durante al menos 18 h o hasta su uso.
NOTA: Las fundas recubiertas se degradan si se almacenan a 70 °C durante más de 2 semanas.
3. Montaje de cámaras de flujo de imágenes
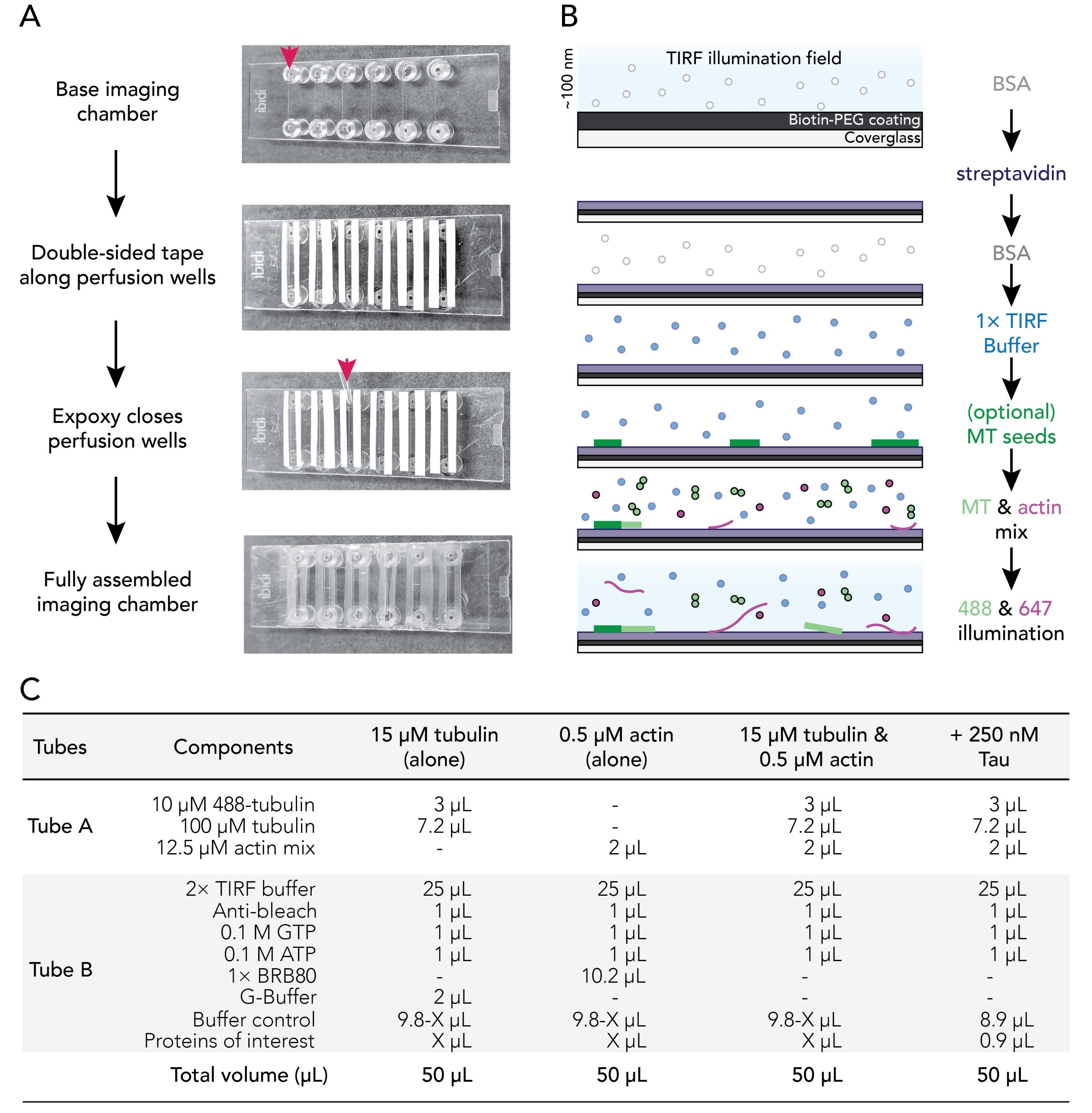
- Cortar 12 tiras de cinta de doble cara de doble respaldo a una longitud de 24 mm. Retire un lado del soporte de la cinta y fije trozos de cinta adyacentes a las seis ranuras presentes en una cámara de imágenes limpia.
NOTA: La cinta debe ser plana para un montaje adecuado, de lo contrario las cámaras de imágenes tendrán fugas. Retire con cuidado el respaldo de la cinta para evitar golpes. Se recomienda deslizar las cámaras con cinta adhesiva sobre una superficie limpia para alisar los contactos de la cámara de cinta. - Retire la segunda pieza de soporte de cinta para exponer el lado pegajoso de la cinta a lo largo de cada ranura de la cámara. Coloque la cinta de la cámara de lado hacia arriba sobre una superficie limpia.
- Mezcle resina epoxi y soluciones endurecedoras 1:1 (o según las instrucciones del fabricante) en un bote de pesaje pequeño.
- Use una punta P1000 para colocar una gota de epoxi mixto entre las tiras de cinta al final de cada ranura de la cámara de imágenes (flecha roja; Figura 1A). Coloque la cinta de la cámara/el lado epoxi hacia arriba sobre una superficie limpia.
- Retire una funda recubierta de la incubadora a 70 °C. Enjuague las superficies recubiertas y no recubiertas de los cubrehojas con ddH2O seis veces, seque con gas nitrógeno filtrado y luego fije a la cámara de imágenes con el recubrimiento de la cubierta hacia el lado hacia la cinta.
- Utilice una punta de pipeta P200 o P1000 para aplicar presión sobre la interfaz cinta-vidrio para garantizar un buen sellado entre la cinta y la cubierta.
NOTA: Con un sello adecuado, la cinta de doble cara se vuelve translúcida. Las cámaras de imágenes que carecen de suficientes contactos de cámara de cinta tendrán fugas. - Incube las cámaras ensambladas a temperatura ambiente durante al menos 5-10 minutos para permitir que el epoxi selle completamente los pozos de la cámara antes de su uso. Las cámaras de perfusión expiran dentro de las 12-18 h posteriores al montaje.
NOTA: Dependiendo de la colocación de la cinta y el grosor de la cinta de doble cara utilizada, la cámara ensamblada tendrá un volumen final de 20-50 μL.
4. Acondicionamiento de cámaras de perfusión
- Utilice una bomba de perfusión (velocidad establecida en 500 μL/min) para intercambiar secuencialmente soluciones de acondicionamiento en la cámara de perfusión de la siguiente manera:
- Flujo de 50 μL de BSA al 1% para cebar la cámara de imagen. Retire el exceso de amortiguación del depósito de conexión de bloqueo Luer.
- Flujo 50 μL de 0,005 mg/ml de estreptavidina. Incubar durante 1-2 min a temperatura ambiente. Retire el exceso de amortiguación del depósito.
- Flujo de 50 μL de BSA al 1% para bloquear la unión inespecífica. Incubar durante 10-30 s. Retire el exceso de amortiguación del depósito.
- Flujo 50 μL de tampón TIRF caliente (37 °C) 1x (1x BRB80, 50 mM KCl, 10 mM TDT, 40 mM de glucosa, 0,25% (v/v) metilcelulosa (4.000 cp)).
NOTA: No retire el exceso de amortiguación del depósito. Esto evita que la cámara se seque, lo que puede introducir burbujas de aire en el sistema. - Opcional: Flujo de 50 μL de semillas de microtúbulos estabilizados al29 y 50% biotiniladas diluidas en 1x tampón TIRF.
NOTA: La dilución adecuada debe determinarse empíricamente y contener la variabilidad de lote a lote. Se recomiendan protocolos a partir del 27,29 como puntos de partida. Una dilución que produce 10-30 semillas por campo de visión funciona bien con esta configuración.
5. Preparación del microscopio
NOTA: Las reacciones bioquímicas que contienen filamentos dinámicos de actina y microtúbulos se visualizan/realizan utilizando un microscopio de fluorescencia de reflexión interna total (TIRF) invertido equipado con láseres de estado sólido de 120-150 mW, un objetivo TIRF de inmersión en aceite de 63x corregido a la temperatura y una cámara EMCCD. Las proteínas de este ejemplo se visualizan en las siguientes longitudes de onda: 488 nm (microtúbulos) y 647 nm (actina).
- Configure el dispositivo de calentador de escenario / objetivo para mantener 35-37 ° C al menos 30 minutos antes de obtener imágenes de la primera reacción bioquímica.
- Establezca los parámetros de adquisición de imágenes de la siguiente manera:
- Establezca el intervalo de adquisición en cada 5 s durante 15-20 min.
- Ajuste las exposiciones láser 488 y 647 a 50-100 ms a una potencia del 5%-10%. Establezca el ángulo TIRF apropiado para el microscopio.
NOTA: Independientemente de la configuración del microscopio, la forma más sencilla de establecer la potencia del láser, la exposición y el ángulo TIRF es realizar ajustes en las imágenes de cualquiera de los polímeros solos (consulte 5.2.2.1 y 5.2.2.2, a continuación). Se recomienda encarecidamente a los usuarios que utilicen la configuración de potencia y exposición láser más baja que aún permita la detección.- Ajuste la reacción de polimerización (Figura 1C) para iniciar el ensamblaje del filamento de actina y adquirir imágenes a 647 nm. Haga los ajustes apropiados.
- Ajustar la reacción de polimerización en un segundo pozo de perfusión acondicionado para iniciar el ensamblaje de microtúbulos (Figura 1C) y visualizar a 488 nm. Haga los ajustes apropiados.
6. Preparación de mezclas de reacción de proteínas
- Prepare la solución madre de tubulina marcada fluorescentemente.
- Determine la concentración de tubulina casera sin etiquetar mediante espectrofotometría en Abs280, de la siguiente manera:
- Espectrofotómetro en blanco con 1xBRB80 sin GTP.
- Calcular la concentración de tubulina utilizando el coeficiente de extinción determinado de 115.000 M-1 cm-1 y la siguiente fórmula:

- Resuspend 488-tubulina marcada con lisina liofilizada de fabricación comercial a 10 μM (1 mg/mL; etiqueta al 100%) con 20 μL de 1x BRB80 sin GTP.
- Descongelar una alícuota de 7,2 μL de tubulina29 reciclada sin etiquetar de 100 μM sobre hielo.
NOTA: La tubulina reciclada es fundamental para el ensamblaje exitoso de microtúbulos in vitro porque elimina los dímeros incompetentes para la polimerización formados en reservas de proteínas congeladas29,30. - Combine 3 μL de 10 μM de 488-tubulina con la alícuota de 7,2 μL de tubulina sin etiquetar 100 μM, no más de 15 min antes de su uso.
- Determine la concentración de tubulina casera sin etiquetar mediante espectrofotometría en Abs280, de la siguiente manera:
- Prepare la solución madre de actina marcada fluorescentemente.
- Para las proteínas caseras, determine la concentración y el porcentaje de la etiqueta de actina a través de la espectrofotometría Abs290 y Abs650, de la siguiente manera:
- Espectrofotómetro en blanco con G-buffer.
- Calcular la concentración de actina no etiquetada utilizando el coeficiente de extinción determinado de 25.974 M-1 cm-1 y la siguiente fórmula:

- Calcule la concentración de lisina etiquetada alexa-647-actina utilizando el coeficiente de extinción de actina no etiquetada, el factor de corrección de flúor de 0.03 y la siguiente fórmula:
[Alexa-647 actina], μM = (Abs290 -
- Calcule la etiqueta porcentual de Alexa-647-actina utilizando el ε determinado para Alexa-647 de 239,000 M-1 cm-1, de la siguiente manera:
% etiqueta de Alexa-647-actina = (Abs290 - (
- Descongelar una alícuota de 2 μL de biotina-actina 3 μM 100% marcada (etiquetada sobre residuos de lisina). Diluir 10 veces añadiendo 18 μL de tampón G.
- Combine 3 μL de actina biotinilada diluida, volúmenes apropiados de actina sin etiquetar y etiquetada (arriba) de modo que la mezcla final sea de 12,5 μM de actina total con etiqueta fluorescente de 10% -30%.
NOTA: Más del 30% por ciento de monómeros de actina fluorescente (final) pueden comprometer la resolución de la imagen a medida que los filamentos se vuelven difíciles de discernir del fondo.
- Para las proteínas caseras, determine la concentración y el porcentaje de la etiqueta de actina a través de la espectrofotometría Abs290 y Abs650, de la siguiente manera:
- Preparar mezclas de reacción (Figura 1C).
- Preparar la mezcla de citoesqueletos (tubo A) combinando 2 μL de la mezcla de actina de 12,5 μM (6.2.3) con la mezcla de material de tubulina (6.1.4), no más de 15 minutos antes de la toma de imágenes. Conservar en hielo hasta su uso.
- Prepare la mezcla de reacción de proteínas (tubo B) combinando todos los demás componentes experimentales y proteínas, incluyendo: 2x tampón TIRF, anti-blanqueador, nucleótidos, tampones y proteínas accesorias. Un ejemplo se muestra en la Figura 1C.
NOTA: La dilución final da como resultado un tampón TIRF 1x que contiene ATP, GTP y fuerza iónica dentro del rango fisiológico estimado.
- Incubar el tubo A y el tubo B por separado a 37 °C durante 30-60 s. Para iniciar la reacción, mezcle y agregue el contenido del tubo B al tubo A (abajo).
7. Dinámica de la actina y los microtúbulos de la imagen
- Acondicionar bien la perfusión (Figura 1B; paso 4, arriba).
- Inicie el ensamblaje de actina y microtúbulos simultáneamente agregando el contenido del tubo B (mezcla de reacción) al tubo A (mezcla de citoesqueletos) (Figura 1C).
- Flujo de 50 μL de reacción que contiene 1x tampón TIRF suplementado con 15 μM de tubulina libre, 1 mM de GTP y monómeros de actina de 0,5 μM y volúmenes apropiados de controles tampón.
- Grabe películas de lapso de tiempo utilizando el software de microscopio para adquirir cada 5 s durante 15-20 minutos.
NOTA: El inicio de la dinámica de la actina y los microtúbulos ocurre dentro de los 2-5 min (Figura 2). Los retrasos más largos indican problemas con el control de la temperatura o problemas relacionados con la concentración de proteínas en la mezcla de reacción. - Acondicionar un nuevo pozo de perfusión (paso 4) y reemplazar el volumen tampón con proteínas reguladoras de interés (es decir, Tau) y controles tampón (Figura 1C). Adquirir como se describe en el paso 7 (arriba) para evaluar las proteínas reguladoras para las funciones emergentes de actina-microtúbulos.
8. Procesar y analizar imágenes utilizando el software FIJI31
- Abra las películas TIRF guardadas y véalas como un compuesto.
- Analice la dinámica de los microtúbulos (Figura 3A), de la siguiente manera:
- Genere una proyección Z máxima basada en el tiempo desde el menú pilas de imágenes.
- Sincronizar la ventana Z-projection con la película TIRF original desde el menú Analizar> Herramientas> Sincronizar Windows .
- Dibuje una línea usando la herramienta de línea recta a lo largo de un microtúbulo de interés en la imagen proyectada en el tiempo.
- Abra el administrador de región de interés (ROI) desde el menú analizar (Analizar> Herramientas> Administrador de ROI).
- Guarde las ubicaciones individuales de los microtúbulos presionando "t". Repetir para todos los microtúbulos de interés.
- Traza kymographs de líneas seleccionadas usando "/" o ejecuta la macro multi-kymo que genera un video y kymograph para cada microtúbulo en el gestor de ROI31.
- Agregue barras de escala de longitud (μm) y tiempo (min) a los kymographs desde el menú Analizar> Herramientas> barras de escala .
- Mida las velocidades de crecimiento de los microtúbulos a partir de las pendientes del kymograph (Figura 3A, 1-2; pendiente de las líneas negras).
- Contar eventos dinámicos de microtúbulos (catástrofe o rebrote) a partir del kymograph generado o utilizando macros de análisis disponibles 5,8,18,25. Las líneas punteadas rojas en la Figura 3A, 1-2 representan eventos de catástrofe/desmontaje rápido.
- Analice la dinámica de la actina (Figura 3B), de la siguiente manera:
- Mida la nucleación de actina, de la siguiente manera:
- Contar el número de filamentos de actina presentes en el campo de visión 100 s después del inicio de la reacción y expresarlos por el área (filamentos por μm2).
- Registre y guarde los datos en el administrador de ROI, como en el paso 8.2.3.1, anterior.
- Mida las tasas de elongación del filamento de actina (Figura 3B), de la siguiente manera:
- Dibuje una línea a lo largo de un filamento de actina de interés utilizando la herramienta de línea segmentada.
- Agregue una línea al administrador de ROI como en el paso 8.2.3.1, anterior.
- Repita el procedimiento siguiendo la línea (agregando cada medición al administrador de ROI) durante al menos cuatro fotogramas de película.
NOTA: Se recomienda medir de siete a ocho fotogramas consecutivos, sin embargo, algunas condiciones ralentizan la polimerización de actina por debajo del límite detectable que puede resolverse mediante configuraciones de objetivos / microscopios. En tal caso, las mediciones se pueden realizar a intervalos regulares sobre fotogramas no consecutivos (por ejemplo, cada cinco fotogramas). - Trazar valores de longitud medidos durante el tiempo transcurrido. La pendiente de la línea generada es la tasa de elongación de la actina en micras/s.
- Transmitir las tasas finales calculadas como subunidades s-1 μM-1 utilizando un factor de corrección de 370 subunidades para tener en cuenta el número de monómeros de actina en una micra de filamento32.
- Mida la nucleación de actina, de la siguiente manera:
- Realizar análisis correlativos para regiones de asociación paralelo actina-microtúbulos (Figura 3C), de la siguiente manera:
- Dibuje una línea a lo largo de un microtúbulo de interés en un punto de tiempo específico (es decir, 300 s después del inicio de la reacción), utilizando la herramienta de línea recta.
- Agregue una línea al administrador de ROI como en el paso 8.2.3.1, anterior.
- Traza la intensidad de la fluorescencia a lo largo de la línea en cada canal.
- Seleccione cada canal con el control deslizante de imagen y trace las intensidades a lo largo de la línea usando "comando k".
- Guarde o exporte valores haciendo clic en el botón "lista" en la ventana de salida.
- Exprese los eventos de acoplamiento actina-microtúbulo como una relación (acción superpuesta con microtúbulos) a partir de eventos individuales o como un recuento de eventos en un campo de visión dado en un punto de tiempo consistente (Figura 3C).
- Alternativa: Utilice software para determinar el porcentaje de superposición de ambos canales 5,12.
- Dibuje una línea a lo largo de un microtúbulo de interés en un punto de tiempo específico (es decir, 300 s después del inicio de la reacción), utilizando la herramienta de línea recta.
Resultados
Con las condiciones descritas anteriormente (Figura 1), los polímeros de actina y microtúbulos deben ser visibles (y dinámicos) dentro de los 2 minutos posteriores a la adquisición de la imagen (Figura 2). Al igual que con cualquier protocolo basado en la bioquímica, la optimización puede ser necesaria para diferentes proteínas reguladoras o lotes de proteínas. Por estas razones, el ángulo TIRF y las exposiciones de la imagen se establecen primero con reacciones que contienen cada polímero individual. Esto confirma que las proteínas almacenadas son funcionales y que hay suficiente proteína marcada para la detección. Si bien no siempre es necesario (y no se realiza aquí), el posprocesamiento de películas (es decir, resta de fondo, promedio o transformaciones de Fourier) se puede utilizar para mejorar el contraste de la imagen (particularmente de los microtúbulos)5,25,33. La visualización directa de filamentos y microtúbulos de actina individuales ofrecida por este ensayo apoya la determinación cuantitativa de varias medidas dinámicas para el componente del citoesqueleto solo o en conjunto, incluidos los parámetros de polimerización (es decir, la tasa de nucleación o elongación), los parámetros de desmontaje (es decir, las tasas de contracción o eventos de catástrofe) y la coalignización / superposición de polímeros (Figura 3 ). Además, estas medidas se pueden utilizar como punto de partida para descifrar la unión o influencia de ligandos reguladores como Tau (Figura 3). Muchas mediciones de filamentos o microtúbulos de actina individuales se pueden hacer a partir de una película TIRF. Sin embargo, debido a las variaciones en el recubrimiento de la cubierta, el pipeteo y otros factores, las mediciones confiables también deben incluir múltiples reacciones / películas de réplica técnica.
Muchas facetas de la dinámica de los microtúbulos se pueden determinar a partir de kymographs de ejemplo, incluida la tasa de elongación de los microtúbulos, así como la frecuencia de los eventos de catástrofe y rescate (Figura 3A). El uso de kymographs para medir la dinámica de la actina en este sistema no es tan sencillo porque los filamentos de actina son más enrevesados que los microtúbulos. Como consecuencia, los parámetros de la dinámica del filamento de actina se miden a mano, lo que consume mucho tiempo y mano de obra. Los recuentos de nucleación se miden como el número de filamentos de actina presentes en un punto de tiempo consistente para todas las condiciones. Estos recuentos varían ampliamente entre los campos de imágenes TIRF, pero se pueden usar con muchas réplicas o para complementar las observaciones de otros ensayos de polimerización. Los recuentos de nucleación también se pueden usar para los microtúbulos si las condiciones de prueba carecen de semillas de microtúbulos estabilizadas. Las tasas de elongación del filamento de actina se miden como la longitud del filamento a lo largo del tiempo de al menos cuatro fotogramas de película. Los valores de velocidad se transmiten por actina micromolar con un factor de corrección de 370 subunidades para tener en cuenta el número de monómeros de actina en una micra de filamento (Figura 3B)32. Las mediciones para definir los comportamientos coordinados entre la actina y los microtúbulos están menos bien definidas. Sin embargo, se han aplicado análisis correlativos para medir la coincidencia de ambos polímeros incluyendo escaneos de línea (Figura 3C) o software de superposición 5,11,34.
Disponibilidad de datos:
Todos los conjuntos de datos asociados con este trabajo se han depositado en Zenodo y están disponibles con una solicitud razonable en: 10.5281 / zenodo.6368327.

Figura 1. Esquemas experimentales: montaje de cámara de flujo a adquisición de imágenes. (A) Montaje de la cámara de imágenes. De arriba a abajo: las cámaras de imágenes IBIDI se pegan con cinta adhesiva a lo largo de los pozos de perfusión (denotados por flecha); se retira la segunda capa (blanca) de soporte de cinta (dejada en la imagen que se muestra para orientar mejor a los usuarios) y se aplica epoxi en el borde de la cámara de perfusión (flecha). Nota: Para orientar más fácilmente a los usuarios dónde colocar el epoxi, se dejó el respaldo blanco en esta imagen. La cubierta limpia y recubierta se une a la cámara de imágenes con el lado del recubrimiento mirando hacia el interior del pozo de perfusión. (B) Diagrama de flujo que ilustra los pasos para acondicionar las cámaras de imágenes para los enlaces biotina-estreptavidina. (C) Ejemplos de reacciones utilizadas para adquirir películas TIRF de microtúbulos dinámicos y filamentos de actina. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2. Secuencias de imágenes de filamentos y microtúbulos de actina en crecimiento en ausencia o presencia de Tau. Montaje de imágenes de lapso de tiempo de ensayos TIRF que contienen 0,5 μM de actina (10% de Alexa-647-actina y 0,09% de biotina-actina marcada) y 15 μM de tubulina libre (4% de HiLyte-488 marcado) en ausencia (A) o presencia (B) de 250 nM Tau. Se muestra el tiempo transcurrido desde el inicio de la reacción (mezcla del tubo A y el tubo B). Barras de escala, 25 μm. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3. Ejemplo de mediciones de microtúbulos y dinámica de filamentos de actina. (A) La proyección de tiempo promedio del canal de tubulina visualiza eficientemente las longitudes totales de microtúbulos para los escaneos de línea utilizados para generar gráficos de kymograph. Las líneas punteadas negras corresponden a los dos kymographs de ejemplo de microtúbulos dinámicos que se muestran a la derecha. El crecimiento (líneas negras sólidas) y las fases de desmontaje (líneas rosadas dotadas; dos denotadas con flechas rosas) de los microtúbulos se muestran en cada kymograph. Barra de escala de tiempo, 3 min. Barra de escala de longitud, 10 μm. Reaction contiene 0,5 μM de actina (10% de etiqueta 647) y 15 μM de tubulina libre (4% de etiqueta de 488-HiLyte). Solo se muestra el canal de tubulina. (B) Dos montajes de imágenes de lapso de tiempo de ejemplo que representan filamentos de una sola actina polimerizando activamente. Las tasas de elongación se calculan como la pendiente de las gráficas de la longitud de los filamentos de actina a lo largo del tiempo por actina micromolar. Por lo tanto, se debe aplicar un factor de corrección de dos a las reacciones de actina de 0,5 μM para la comparación de las tasas típicamente determinadas a la concentración de actina de 1 μM. Ejemplos de cinco filamentos se muestran a la derecha. Barras de escala, 10 μm. Reaction contiene 0,5 μM de actina (10% de etiqueta 647) y 15 μM de tubulina libre (4% de etiqueta de 488-HiLyte). Solo se muestra el canal de actina. (C) Imágenes TIRF de microtúbulos dinámicos (MT) (verde) y filamentos de actina (púrpura) polimerizando en ausencia (izquierda) o presencia de 250 nM Tau (centro). Las líneas de puntos azules y las flechas marcan dónde se dibujó una línea para los gráficos de escaneo de línea correspondientes a cada condición (debajo de cada imagen). La superposición entre los microtúbulos y las regiones de actina (que se muestra como negro) se puede puntuar en un punto de tiempo establecido por área (derecha). Barras de escala, 25 μm. Las reacciones contienen 0,5 μM de actina (10% de etiqueta 647) y 15 μM de tubulina libre (4% de etiqueta de 488-HiLyte) con o sin 250 nM tau. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Discusión
El uso de la microscopía de fluorescencia de reflexión interna total (TIRF) para visualizar proteínas purificadas ha sido un enfoque fructífero y convincente para diseccionar mecanismos únicos de regulación citoesquelética 5,23,24,25,26,27,35. En comparación con los ensayos bioquímicos tradicionales, las reacciones TIRF requieren volúmenes muy pequeños (50-100 μL), y las mediciones cuantitativas de la dinámica citoesquelética se pueden obtener de un ensayo individual. La mayoría de los estudios de dinámica citoesquelética se centran en un solo sistema polimérico (es decir, filamentos de actina o microtúbulos), por lo que las mediciones detalladas de la diafonía o los comportamientos emergentes entre los filamentos de actina y los microtúbulos que se observan típicamente en las células han permanecido esquivas y difíciles de recapitular en el tubo de ensayo. Para resolver este problema, este protocolo describe un sistema de microscopía TIRF de filamento único que permite la visualización directa de polímeros dinámicos de actina y microtúbulos en la misma reacción bioquímica. Por lo tanto, este método va más allá de los ensayos tradicionales que recapitulan el comportamiento dinámico de los filamentos de actina o microtúbulos solos. Esta técnica también se realizó con Tau como ejemplo de cómo varias propiedades dinámicas cambian en presencia de un factor de acoplamiento del citoesqueleto. Este protocolo se puede utilizar con proteínas adicionales conocidas o sospechadas para coordinar la dinámica de la actina o los microtúbulos, incluyendo (pero no limitado a) MACF, GAS, forminas y más. Finalmente, los análisis de ejemplo proporcionados se pueden utilizar como guía para cuantificar los datos adquiridos con este protocolo.
"Ver para creer" es una razón convincente para realizar ensayos basados en microscopía. Sin embargo, se requiere precaución en la ejecución e interpretación de los experimentos de microscopía TIRF. Uno de los principales desafíos de los ensayos de ensamblaje citoesquelético es que muchas condiciones de imagen de uso común no son compatibles con cada polímero. Los microtúbulos y la actina suelen tener diferentes requisitos de tampón, temperatura, sal, nucleótidos y concentración para la polimerización. La actina, la tubulina, las proteínas reguladoras de interés y los tampones utilizados en este protocolo son sensibles a los ciclos de congelación-descongelación. Por lo tanto, es necesario un manejo cuidadoso de proteínas y tampones para ejecutar con éxito este protocolo. Para aliviar muchas de estas preocupaciones, se recomienda encarecidamente el uso de tubulina recién reciclada (congelada durante <6 semanas) y la limpieza previa de actinas congeladas / resuspendidas a través de la ultracentrifugación. Estas consideraciones también se aplican a la miríada de proteínas reguladoras que se evaluarán con este procedimiento, que pueden ser sensibles a los ciclos de congelación-descongelación o a la concentración de sales tampón 5,11,36.
Desafortunadamente, no existe un búfer único para todos sin compensaciones experimentales. Para asignar más volumen para proteínas de menor concentración, se puede incluir ATP y GTP en la solución tampón 2x TIRF (Figura 1C). Sin embargo, debido a que estos nucleótidos son extremadamente sensibles a los ciclos de congelación-descongelación, no se recomienda. Los compuestos de eliminación de oxígeno utilizados aquí (es decir, catalasa y glucosa-oxidasa) son necesarios para visualizar proteínas durante largos períodos de tiempo (minutos a horas), pero se sabe que restringen la polimerización de microtúbulos a altas concentraciones5. En relación con estas consideraciones tampón, una limitación de este protocolo es que algunas proteínas reguladoras canónicas asociadas a microtúbulos pueden requerir más o menos sal para recapitular las funciones que se encuentran en las células o ensayos utilizando microtúbulos solos (sin actina). Cambiar la naturaleza o la concentración de sal para abordar estas preocupaciones probablemente influirá en las tasas de polimerización de filamentos de actina y / o los parámetros de la dinámica de los microtúbulos. Se requieren mediciones de múltiples parámetros descriptivos (mínimamente, nucleación, tasas de elongación y estabilidad) (Figura 3) para confirmar el éxito del protocolo o para documentar explícitamente los efectos de tampones específicos o proteínas reguladoras. Por ejemplo, demasiada polimerización de filamentos de actina puede oscurecer los eventos de acoplamiento actina-microtúbulo en cuestión de segundos. En consecuencia, el ajuste fino de las condiciones experimentales mediante la reducción de la concentración general de actina o la inclusión de proteínas adicionales para suprimir la nucleación de actina (es decir, la profilina) extenderá el período general en el que se pueden ver claramente las actividades coordinadas de actina-microtúbulos. Los controles que abordan estos requisitos previos y las réplicas técnicas (más allá de múltiples campos de visión) son fundamentales para que los usuarios generen resultados confiables y reproducibles.
Los estudios basados en células brindan una oportunidad limitada para observar las relaciones directas proteína-proteína o la acción de complejos reguladores. En contraste, algunos de los mecanismos obtenidos de los ensayos in vitro no siempre reflejan los comportamientos exactos de las proteínas observadas en las células. Este clásico dilema bioquímico puede ser abordado en futuras aplicaciones de esta técnica con modificaciones específicas. Por ejemplo, la adición de proteínas de acoplamiento funcionales marcadas fluorescentemente expande este método de estudios de filamento único a estudios de molécula única. Los ensayos se pueden modificar aún más para usar extractos celulares que pueden agregar los factores clave desconocidos "faltantes" necesarios para recapitular fenómenos similares a las células. Por ejemplo, los ensayos basados en TIRF que emplean extractos de levadura o Xenopus han reconstituido anillos de actomiosina contráctil37, husos mitóticos26,38, componentes de actina o ensamblaje de microtúbulos39,40, e incluso dinámica en el centrosoma y los cinetocoros 36,41,42,43 . Además, tales sistemas pueden allanar el camino hacia sistemas celulares artificiales que tienen lípidos o factores de señalización presentes 44,45,46.
Divulgaciones
No hay conflictos de intereses que revelar.
Agradecimientos
Agradezco a Marc Ridilla (Repair Biotechnologies) y Brian Haarer (SUNY Upstate) por sus útiles comentarios sobre este protocolo. Este trabajo fue apoyado por los Institutos Nacionales de Salud (GM133485).
Materiales
| Name | Company | Catalog Number | Comments |
| 1% BSA (w/v) | Fisher Scientific | BP1600-100 | For this purpose (blocking TIRF chambers), BSA is resuspended in ddH20 and filtered through a 0.22 µm filter. |
| 1× BRB80 | Homemade | 80 mM PIPES, 1 mM MgCl2, 1 mM EGTA, pH 6.8 with KOH | |
| 10 mg/mL (1000 U) glucose oxidase | Sigma Aldrich Inc, St. Louis, MO | G2133-50KU | Combined with catalase, aliquot and store at -80 oC until use |
| 100 µM tubulin | Cytoskeleton Inc, Denver, CO | T240 | Homemade tubulins should be recycled before use to remove polymerization-incompetent tubulin (Hyman et al. (1992)29; Li and Moore (2020)30). Commercially available tubulins are often too dilute to recycle, but function well if resuspended according to manufacturer’s instructions and pre-cleared via ultracentrifugation (278,000 × g) for 60 min, before use. |
| 100 mM ATP | Gold Biotechnology Inc, Olivette, MO | A-081 | Resuspended in ddH20 (pH 7.5) and filter sterilized. |
| 100 mM GTP | Fisher Scientific | AC226250010 | Resuspended in 1× BRB80 (pH 6.8) and filter sterilized. |
| 120-150 mW solid-state lasers | Leica Microsystems | 11889151; 11889148 | |
| 2 mg/mL catalase | Sigma Aldrich Inc, St. Louis, MO | C40-100 | Combined with glucose oxidase, aliquot and store at -80 oC until use |
| 2× TIRF buffer | Homemade | 2× BRB80, 100 mM KCl, 20 mM DTT, 80 mM glucose, 0.5% (v/v) methylcellulose (4,000 cp); Note: 1 µL of 0.1M GTP and 1 µL of 0.1M ATP added separately to TIRF reactions to avoid repeated freeze-thaw cycles. | |
| 24 × 60 mm, #1.5 coverglass | Fisher Scientific, Waltham, MA | 22-266882 | Coverglass must be extensively washed before use (Smith et al. (2014)22) |
| 37 oC heatblock | |||
| 37 oC water bath | |||
| 5 mg/mL Streptavidin (600x stock) | Avantor, Philadelphia, PA | RLS000-01 | Resuspended in Tris-HCl (pH 8.8); dilute the aliquot to 1× in HEK buffer on day of use |
| 5 min Epoxy resin and hardener | Loctite, Rocky Hill, CT | 1365736 | Combined resin and hardener may take up to 30 min to cure. |
| 50% biotinylated-GpCpp microtubule seeds | Cytoskeleton Inc; Homemade | T333P | (optional) GppCpp or Taxol stabilized microtubule seeds can more efficiently mediate microtubule polymerization. Taxol and GppCpp stabilize microtubules in different ways that can affect the microtubule lattice structure and ability of certain regulatory proteins to bind to the stabilized portion of the microtubule. A method to make diverse kinds of microtubule seeds is outlined in Hyman et al. (1992). |
| 70 oC incubator | |||
| Actin mix stock | Homemade; this protocol | A 12.5 µM actin mix comprised of labeled (fluorescent and biotinylated) and unlabeled actin for up to six reactions. 2 µL of stock is used in the final TIRF reaction. The final concentration of actin used in each reaction is 0.5 µM (10% Alexa-647; 0.09% biotin-labeled). | |
| Appropriate buffer controls | Homemade | Combination of buffers from all proteins being assessed | |
| Biotin-PEG-silane (MW 3,400) | Laysan Bio Inc | biotin-PEG-SIL-3400 | Dispensed into 2-5 mg aliquots, backfilled with nitrogen, parafilmed closed, and stored at -20 oC with desiccant until use |
| Biotinylated actin | Cytoskeleton Inc; Homemade | AB07 | Biotin-actin is made by labeling on lysine residues and thus assumed to be at least 100% labeled, but varies with different lots/preparations. Optimal biotinylated actin concentrations must be empirically determined for particular uses/experimental designs. Higher concentrations permit more efficient tracking, but may impede polymerization or interactions with regulatory proteins. Here a small percentage (0.09% or 900 pM) biotinylated actin is present in the final TIRF reaction. |
| Dishsoap | Dawn, Procter and Gamble, Cincinnati, OH | For unknown reasons, the blue version cleans coverslips more efficiently than other available colors. | |
| Dry ice | |||
| FIJI Software | www.https://imagej.net/software/fiji/downloads | Schneider et al. (2012)31. | |
| Fluorescently labeled actin | Cytoskeleton Inc; Homemade | AR05 | Homemade fluorescently labeled actin is stored in G-buffer supplemented with 50% glycerol at -20 oC (Spudich et al. (1971)47; Liu et al. (2022)48). Fluorescently labeled actin is dialyzed against G-buffer and precleared via ultracentrifugation for 60 min at 278,000 × g before use. |
| Fluorescently labeled tubulin | Cytoskeleton Inc | TL488M, TLA590M, TL670M | Resuspended in 20 µL 1× BRB80 (10 µM final concentration) and pre-cleared via ultracentrifugation (278,000 × g) for 60 min, before use. |
| G-buffer | Homemade | 3 mM Tris-HCl (pH 8.0), 0.2 mM CaCl2, 0.5 mM DTT, 0.2 mM ATP | |
| HEK Buffer | Homemade | 20 mM HEPES (pH 7.5), 1 mM EDTA (pH 8.0), 50 mM KCl | |
| Ice | |||
| Ice bucket | |||
| Imaging chambers | IBIDI, Fitchburg, WI | 80666 | Order chambers with no bottom to utilize different coverslip coatings |
| iXon Life 897 EMCCD camera | Andor, Belfast, Northern Ireland | 8114137 | |
| LASX Premium microscope software | Leica Microsystems | 11640611 | |
| Methylcellulose (4,000 cp) | Sigma Aldrich Inc | M0512 | |
| Microscope base equipped with TIRF module | Leica Microsystems, Wetzlar, Germany | 11889146 | |
| mPEG-silane (MW 2,000) | Laysan Bio Inc, Arab, AL | mPEG-SIL-2000 | Dispensed into 10-15 mg aliquots, backfilled with nitrogen, parafilmed closed, and stored at -20 oC with desiccant until use |
| Objective heater and heated stage insert | OKO labs, Pozzioli, Italy | 8113569 | Set temperature controls to 35-37 oC. Use manufacturer suggestions for accurate calibration. |
| Perfusion pump | Harvard Apparatus, Holliston, MA | 704504 | A syringe and tubing can be substituted. |
| Petri Dish, 100 x 15 mm | Genesee Scientific, San Diego, CA | 32-107 | |
| Plastic slide mailer container | Fisher Scientific | HS15986 | |
| SA-S-1L-SecureSeal 0.12 mm thick | Grace Biolabs, Bend, OR | 620001 | Double-sided tape of precise manufactured dimensions is strongly recommended. |
| Small styrofoam container | Abcam, Cambridge, UK | Reused from shipping | |
| Small weigh boat | Fisher Scientific | 02-202-100 | |
| Spectrophotometer | |||
| Tau | Cytoskeleton Inc | TA01 | Three isoforms of Tau are present in the commercially available preparation of Tau. The concentration in this protocol was determined from the highest molecular weight band (14.3 µM, when resuspended per manufacturer’s recommendations with 50 µL of ddH20). |
| Temperature corrected 63× Plan Apo 1.47 N.A. oil immersion TIRF objective | Leica Microsystems | 11506319 | |
| Tubulin stock | Homemade; this protocol | A tubulin stock consisting of 7.2 µL recycled 100 µM unlabeled tubulin and 3 µL of 10 µM resuspended commercially available fluorescently labeled tubulin. One tubulin stock is used per reaction and thawed/stored on ice. The final concentration of free tubulin in each reaction is 15 µM (4% labeled). More than 15 µM tubulin will result in hyperstabilized (not dynamic) microtubules, whereas concentrations below 7.5 µM free tubulin do not polymerize well. Careful determination of protein concentration and handling is required. | |
| Unlabeled actin (dark) | Cytoskeleton Inc; Homemade | AKL99 | Actin nucleates are almost always present in commercially available (lyophilized) or frozen actins and contribute to variability in quantitative measurements (Spudich et al. (1971)47; Liu et al. (2022)48). Rabbit muscle actin is stored in G-buffer at -80 oC and precleared via ultracentrifugation for 60 min at 278,000 × g before use. Several actin stock solutions are made throughout the day (making no more than enough for six reactions at a time is strongly recommended). |
Referencias
- Pimm, M. L., Henty-Ridilla, J. L. New twists in actin-microtubule interactions. Molecular Biology of the Cell. 32 (3), 211-217 (2021).
- Dogterom, M., Koenderink, G. H. Actin-microtubule crosstalk in cell biology. Nature Reviews. Molecular Cell Biology. 20 (1), 38-54 (2019).
- Etienne-Manneville, S. Actin and microtubules in cell motility: which one is in control. Traffic. 5 (7), 470-477 (2004).
- Rodriguez, O. C., et al. Conserved microtubule-actin interactions in cell movement and morphogenesis. Nature Cell Biology. 5 (7), 599-609 (2003).
- Prezel, E., et al. TIRF assays for real-time observation of microtubules and actin coassembly: Deciphering tau effects on microtubule/actin interplay. Methods in Cell Biology. 141, 199-214 (2017).
- Griffith, L. M., Pollard, T. D. The interaction of actin filaments with microtubules and microtubule-associated proteins. The Journal of Biological Chemistry. 257 (15), 9143-9151 (1982).
- Henty-Ridilla, J. L., Rankova, A., Eskin, J. A., Kenny, K., Goode, B. L. Accelerated actin filament polymerization from microtubule plus ends. Science. 352 (6288), 1004 (2016).
- Elie, A., et al. Tau co-organizes dynamic microtubule and actin networks. Scientific Reports. 5 (1), 1-10 (2015).
- Preciado López, M., et al. Actin-microtubule coordination at growing microtubule ends. Nature Communications. 5 (1), 1-9 (2014).
- Oberhofer, A., et al. Molecular underpinnings of cytoskeletal cross-talk. Proceedings of the National Academy of Sciences of the United States of America. 117 (8), 3944-3952 (2020).
- Nakos, K., et al. Septins mediate a microtubule-actin crosstalk that enables actin growth on microtubules. bioRxiv. , (2022).
- Kučera, O., Gaillard, J., Guérin, C., Théry, M., Blanchoin, L. Actin-microtubule dynamic composite forms responsive active matter with memory. bioRxiv. , (2022).
- Kundu, T., Dutta, P., Nagar, D., Maiti, S., Ghose, A. Coupling of dynamic microtubules to F-actin by Fmn2 regulates chemotaxis of neuronal growth cones. Journal of Cell Science. 134 (13), 252916 (2021).
- Roth-Johnson, E. A., Vizcarra, C. L., Bois, J. S., Quinlan, M. E. Interaction between microtubules and the Drosophila formin Cappuccino and its effect on actin assembly. The Journal of Biological Chemistry. 289 (7), 4395-4404 (2014).
- Gaillard, J., et al. Differential interactions of the formins INF2, mDia1, and mDia2 with microtubules. Molecular Biology of the Cell. 22 (23), 4575-4587 (2011).
- Bartolini, F., et al. The formin mDia2 stabilizes microtubules independently of its actin nucleation activity. The Journal of Cell Biology. 181 (3), 523-536 (2008).
- Sider, J. R., et al. Direct observation of microtubule-F-actin interaction in cell free lysates. Journal of Cell Science. 112 (12), 1947-1956 (1999).
- Alkemade, C., et al. Cross-linkers at growing microtubule ends generate forces that drive actin transport. Proceedings of the National Academy of Sciences of the United States of America. 119 (11), 2112799119 (2022).
- Axelrod, D. Total internal reflection fluorescence microscopy in cell biology. Methods in Enzymology. 361, 1-33 (2003).
- Amann, K. J., Pollard, T. D. Direct real-time observation of actin filament branching mediated by Arp2/3 complex using total internal reflection fluorescence microscopy. Proceedings of the National Academy of Sciences of the United States of America. 98 (26), 15009-15013 (2001).
- Al-Bassam, J. Reconstituting dynamic microtubule polymerization regulation by TOG domain proteins. Methods in Enzymology. 540, 131-148 (2014).
- Smith, B. A., Gelles, J., Goode, B. L. Single-molecule studies of actin assembly and disassembly factors. Methods in Enzymology. 540, 95-117 (2014).
- Wioland, H., Jégou, A., Romet-Lemonne, G. Celebrating 20 years of live single-actin-filament studies with five golden rules. Proceedings of the National Academy of Sciences of the United States of America. 119 (3), 2109506119 (2022).
- Ganzinger, K. A., Schwille, P. More from less - bottom-up reconstitution of cell biology. Journal of Cell Science. 132 (4), (2019).
- Mahamdeh, M., Howard, J. Implementation of interference reflection microscopy for label-free, high-speed imaging of microtubules. Journal of Visualized Experiments. (150), e59520 (2019).
- King, M. R., Petry, S. Phase separation of TPX2 enhances and spatially coordinates microtubule nucleation. Nature Communications. 11 (1), 270 (2020).
- Ramirez-Rios, S., et al. A TIRF microscopy assay to decode how tau regulates EB's tracking at microtubule ends. Methods in Cell Biology. 141, 179-197 (2017).
- Smith, B. A., et al. Three-color single molecule imaging shows WASP detachment from Arp2/3 complex triggers actin filament branch formation. eLife. 2, 01008 (2013).
- Hyman, A. A., Salser, S., Drechsel, D. N., Unwin, N., Mitchison, T. J. Role of GTP hydrolysis in microtubule dynamics: information from a slowly hydrolyzable analogue GMPCPP. Molecular Biology of the Cell. 3 (10), 1155-1167 (1992).
- Li, G., Moore, J. K. Microtubule dynamics at low temperature: evidence that tubulin recycling limits assembly. Molecular Biology of the Cell. 31 (11), 1154-1166 (2020).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Pollard, T. D., Blanchoin, L., Mullins, R. D. Molecular mechanisms controlling actin filament dynamics in nonmuscle cells. Annual Review of Biophysics and Biomolecular Structure. 29, 545-576 (2000).
- Kapoor, V., Hirst, W. G., Hentschel, C., Preibisch, S., Reber, S. MTrack: Automated detection, tracking, and analysis of dynamic microtubules. Scientific Reports. 9 (1), 3794 (2019).
- Willige, D., et al. Cytolinker Gas2L1 regulates axon morphology through microtubule-modulated actin stabilization. EMBO reports. 20 (11), (2019).
- Hirst, W. G., Kiefer, C., Abdosamadi, M. K., Schäffer, E., Reber, S. In vitro reconstitution and imaging of microtubule dynamics by fluorescence and label-free microscopy. STAR Protocols. 1 (3), 100177 (2020).
- Farina, F., et al. The centrosome is an actin-organizing centre. Nature Cell Biology. 18 (1), 65-75 (2016).
- Mishra, M., et al. In vitro contraction of cytokinetic ring depends on myosin II but not on actin dynamics. Nature Cell Biology. 15 (7), 853-859 (2013).
- Groen, A. C., Ngyuen, P. A., Field, C. M., Ishihara, K., Mitchison, T. J. Glycogen-supplemented mitotic cytosol for analyzing Xenopus egg microtubule organization. Methods in Enzymology. 540, 417-433 (2014).
- Pollard, L. W., Garabedian, M. V., Alioto, S. L., Shekhar, S., Goode, B. L. Genetically inspired in vitro reconstitution of Saccharomyces cerevisiae actin cables from seven purified proteins. Molecular Biology of the Cell. 31 (5), 335-347 (2020).
- Bergman, Z. J., Wong, J., Drubin, D. G., Barnes, G. Microtubule dynamics regulation reconstituted in budding yeast lysates. Journal of Cell Science. 132 (4), 219386 (2018).
- Inoue, D., et al. Actin filaments regulate microtubule growth at the centrosome. The EMBO Journal. 38 (11), (2019).
- Colin, A., Singaravelu, P., Théry, M., Blanchoin, L., Gueroui, Z. Actin-network architecture regulates microtubule dynamics. Current Biology. 28 (16), 2647-2656 (2018).
- Torvi, J. R., et al. Reconstitution of kinetochore and microtubule dynamics reveals a role for a kinesin-8 in establishing end-on attachments. bioRxiv. , (2022).
- Nguyen, P. A., et al. Spatial organization of cytokinesis signaling reconstituted in a cell-free system. Science. 346 (6206), 244-247 (2014).
- Abu Shah, E., Keren, K. Symmetry breaking in reconstituted actin cortices. eLife. 3, 01433 (2014).
- Vendel, K. J. A., Alkemade, C., Andrea, N., Koenderink, G. H., Dogterom, M. In vitro reconstitution of dynamic co-organization of microtubules and actin filaments in emulsion droplets. Cytoskeleton Dynamics. 2101, 53-75 (2020).
- Spudich, J. A., Watt, S. The regulation of rabbit skeletal muscle contraction. Biochemical studies of the interaction of the tropomyosin-troponin complex with actin and the proteolytic fragments of myosin. The Journal of Biological Chemistry. 246 (15), 4866-4871 (1971).
- Liu, X., Pimm, M. L., Haarer, B., Brawner, A. T., Henty-Ridilla, J. L. Biochemical characterization of actin assembly mechanisms with ALS-associated profilin variants. European Journal of Cell Biology. 101 (2), 151212 (2022).
- Pimm, M. L., Liu, X., Tuli, F., Lojko, A., Henty-Ridilla, J. L. Visualizing functional human profilin in cells and in vitro applications. bioRxiv. , (2021).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados