
Method Article
In Vivo Électrophysiologique mesure du potentiel d’Action musculaire composé de membres antérieurs dans des modèles murins de la dégénérescence des motoneurones
Dans cet article
Résumé
La mesure de la conduction nerveuse est un outil utile pour évaluer les modèles murins de la neurodégénérescence, mais il est fréquemment appliqué seulement pour stimuler le nerf sciatique dans les membres postérieurs. Nous décrivons ici une technique pour mesurer les composés muscle potentiel d’action (CMAP) in vivo dans les muscles des membres antérieurs de souris innervés par le plexus brachial.
Résumé
Évaluer les fonctionnalités de l’axone de nerf fournit des informations détaillées sur l’évolution des troubles neuromusculaires. Des enregistrements électrophysiologiques offrent une approche sensible pour mesurer la conduction nerveuse chez les humains et les modèles de rongeurs. Afin d’élargir les possibilités techniques d’électromyographie chez la souris, la mesure des potentiels d’action composé de muscle (CMAPs) des nerfs du plexus brachial dans la patte avant à l’aide d’électrodes aiguilles est décrite ici. Enregistrements de la CMAP après stimulation du nerf sciatique dans les membres postérieurs ont été décrits précédemment. La méthode nouvellement introduite ici permet l’évaluation de la conductibilité nerveuse à un autre site et ainsi donne un aperçu plus profond de la fonction neuromusculaire. La technique fournit des informations sur le nombre relatif des axones fonctionnelles et le niveau de la myélinisation. Ainsi, cette méthode peut être appliquée pour évaluer les maladies axonales ainsi que conditions démyélinisantes. Cette méthode mini-invasive ne nécessite pas l’extraction du nerf et donc il est approprié pour des mesures répétées pour un suivi longitudinal chez le même animal. Des enregistrements similaires sont effectuées dans des configurations cliniques à souligner la pertinence translationnelle de la méthode.
Introduction
Électrophysiologie est utilisé comme un outil de diagnostic dans les maladies neuromusculaires comme les maladies du motoneurone, plexopathies, neuropathies, troubles de la jonction neuromusculaire et myopathies. Dans la sclérose latérale amyotrophique (SLA), dont principalement les motoneurones sont touchés, les lésions axonales et de paralysie musculaire1 sont consignées dans les amplitudes CMAP réduites sur les études de conduction de nerf (NCS). Dans la maladie de Charcot-Marie-Tooth (CMT), dégénérescence axonale et la démyélinisation peuvent être estimées dans les nerfs périphériques à l’aide de NCS2. Cette technique peut être utilisée pour confirmer le diagnostic, ainsi que d’évaluer la progression de maladie3,4. NCS permettent l’estimation de la pathologie axonale, qui se déduite de l’amplitude du potentiel d’action amplitude5, et l’étendue de la démyélinisation - qui se traduit par la vitesse de conduction réduite, prolongée des latences distales, ou bloc de conduction 6.
Mesure de la CMAP est une méthode rapide et sensible pour évaluer la conduction nerveuse tant chez les humains et les souris. Alors que chez les patients les NCS sont effectuées systématiquement à divers endroits pour enregistrer différents nerfs et les muscles, chez les souris, CMAP effectuées les mesures généralement uniquement pour le nerf sciatique afin d’évaluer les fonctionnalités de nerfs dans les membres postérieurs. Toutefois, dans certaines études animales il serait avantageux d’enregistrement CMAP en avant - et membres postérieurs, par exemple, de suivre la progression de la maladie différentiel entre fore - et membres postérieurs chez les modèles murins ALS.
Ici, nous présentons une méthode pour enregistrer les membres antérieurs des souris à l’aide d’électrodes aiguilles CMAPs. En outre, nous fournissons une approche pour mesurer des CMAPs des membres postérieurs, même avec des électrodes aiguilles. La mesure des CMAPs de membres postérieurs avec des électrodes de l’anneau a été présentée plus tôt7,8. L’enregistrement des CMAPs utilisant des électrodes aiguilles est une méthode de mesure rapide, il ne nécessite pas de rasage de la fourrure et la procédure pour mesurer les postérieurs et antérieurs prend seulement 10 min par animal pour un chercheur expérimenté. En outre, cette approche mini-invasive est faisable pour des mesures répétées permettre le suivi longitudinal des nerfs multiples chez les animaux.
Protocole
Tous les animaux étaient logés dans des conditions normalisées selon les lignes directrices de la KU Leuven - Université de Louvain et les directives connexes européens (Union européenne Directive 2010/63/UE pour l’expérimentation animale). Toutes les expériences animales ont été approuvées par le Comité local d’éthique de la KU Leuven.
1. anesthésie et préparation animaux
- Induire l’anesthésie chez la souris avec inhalation isoflurane/oxygène. Utilisez 4 d’isoflurane pour l’induction de l’anesthésie et 2-3 % pour le maintien à 2,5 L/min débit d’oxygène. Ajustez le pourcentage d’isoflurane pour l’entretien de l’anesthésie selon l’état de la souris, c'est-à-dire, petits et faibles souris requièrent moins anesthésiques. Confirmez par exempleanesthésie adéquate, en appliquant une pression légère sur le postérieur marche pad pour vérifier l’absence d’un réflexe de retrait de douleur.
- Contrôler la température de corps de la souris à l’aide d’une plaque chauffante thermostatique à 37 ° C pour éviter la baisse de la température corporelle pendant l’anesthésie.
- Placez la souris avec l’ogive pour l’entretien de l’anesthésie. Ce que l’animal ait un apport suffisant d’oxygène en vérifiant que l’ogive ne bloque pas les voies respiratoires et que l’animal respire régulièrement.
- Lors de l’enregistrement, vérifier si la souris est suffisamment anesthésiée en observant le rythme de la respiration (environ 1 Hz en anesthésie) et l’absence d’un réflexe de retrait sur la légère pression. Augmenter la concentration de l’isoflurane manuellement si l’anesthésie n’est pas assez profond.
- Après les mesures, laisser la souris pour récupérer sur la plaque chauffante ou dans la chaleur d’une lampe infrarouge jusqu'à ce qu’il a repris connaissance suffisante pour maintenir le décubitus sternal, pendant environ 2-5 min. Ne laissez pas la souris et sans surveillance en compagnie d’autres souris jusqu'à ce qu’il a pleinement récupéré de l’anesthésie.
2. mesure de la CMAP dans les membres antérieurs et postérieurs

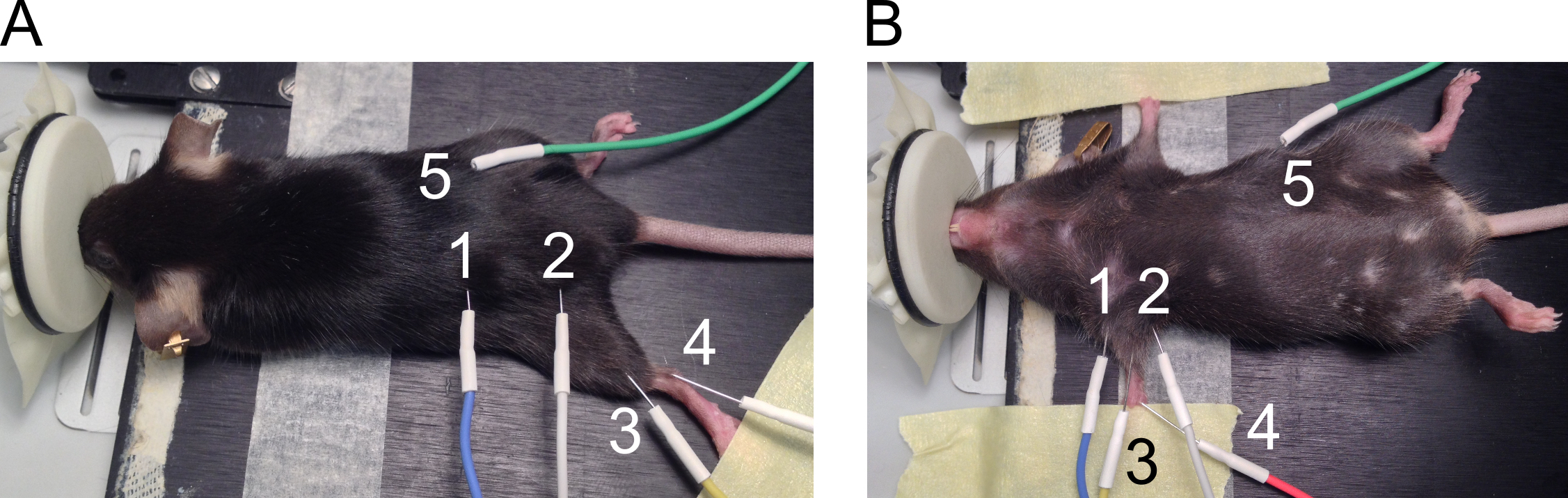
Figure 1. Positionnement des électrodes pour les mesures de la CMAP. La position des électrodes est présentée pour hind-(A) et ses pattes avant (B). Les électrodes sont numérotées comme suit : 1 : anode et 2 : cathode stimulant des électrodes, 3 : électrode de l’enregistrement actif, 4 : électrode de référence et 5 : électrode de mise à la terre. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
- Utilisez les électrodes aiguilles 27 G pour les mesures de CMAP du membre postérieur et des membres antérieurs. Voir la Figure 1 pour les endroits recommandés de positionnement des électrodes.
- Placer les électrodes sur le postérieur comme suit.
- Placez votre souris sur le coussin chauffant en position couchée. Étendre le postérieur au niveau du genou et fixer la patte sur la surface de travail à l’aide de ruban adhésif (Figure 1 a).
- Placer les électrodes de stimulation par voie sous-cutanée des deux côtés de l’échancrure sciatique avec une distance d’environ 2 cm (1 = anode et 2 = cathode) entre les électrodes. Soulever la peau pour insérer l’aiguille perpendiculairement à travers la peau et pousser environ 5 mm de l’aiguille sous la peau sans percer les muscles sous-jacents.
- De même, placer l’électrode d’enregistrement (3) par voie sous-cutanée en alignant le muscle gastrocnémien. Insérer l’électrode de référence (4) par voie sous-cutanée à côté le tendon d’Achille dans un angle de 30 degrés et laisser 2-5 mm de l’aiguille sous la peau. Placer l’électrode de terre (5) par voie sous-cutanée sur le côté de la souris de manière similaire comme les électrodes stimulantes, mais la position de cette électrode n’est pas critique pour la mesure.
- Placer les électrodes sur les membres antérieurs comme suit.
- Positionnez la souris sur le coussin chauffant en position couchée et utiliser du ruban adhésif pour étendre les deux pattes avant sur les côtés du corps (Figure 1 b).
- Placer les électrodes stimulants (1 = anode et 2 = cathode) par voie sous-cutanée des deux côtés de la patte avant pour s’aligner avec les nerfs du plexus brachial. Soulever la peau pour insérer l’aiguille perpendiculairement à travers la peau et pousser environ 5 mm de l’aiguille sous la peau sans percer les muscles sous-jacents.
- Placer l’électrode d’enregistrement (3) par voie sous-cutanée sur le dessus du muscle biceps brachial en soulevant la peau. Placer l’électrode de référence (4) sur les tapis de marche en 3 mm de profondeur à un angle de 30 degrés. Placer l’électrode de terre (5) par voie sous-cutanée sur le côté de la souris.
Remarque : Les électrodes sont à proximité immédiate de l’autre dans cette configuration. Électrodes empêcher de toucher l’autre comme cela déforme l’enregistrement.
3. Acquisition de données
- Commencer la stimulation en appuyant sur le bouton relance récurrente dans l’unité de commande et tournez le bouton de contrôleur d’intensité pour augmenter la stimulation. Stimuler tous les axones par 1 impulsion/s avec durée de stimulation de 0,1 ms. Sélectionnez la bonne fréquence et la durée dans les menus déroulants dans le logiciel.
- Rejoindre des stimuli supramaximal (5-20 mA ; démyélinisantes et le couchage jusqu'à 60 mA), appliquer des stimuli croissantes en tournant le bouton de régulateur d’intensité jusqu'à ce que l’amplitude de la réponse de la CMAP cesse d’augmenter. A partir de là, le stimulus augmenter de 20 %, pour s’assurer que l’amplitude de la CMAP a atteint sa réponse maximale. Fin de la stimulation en poussant le bouton de stimulation récurrente à nouveau.
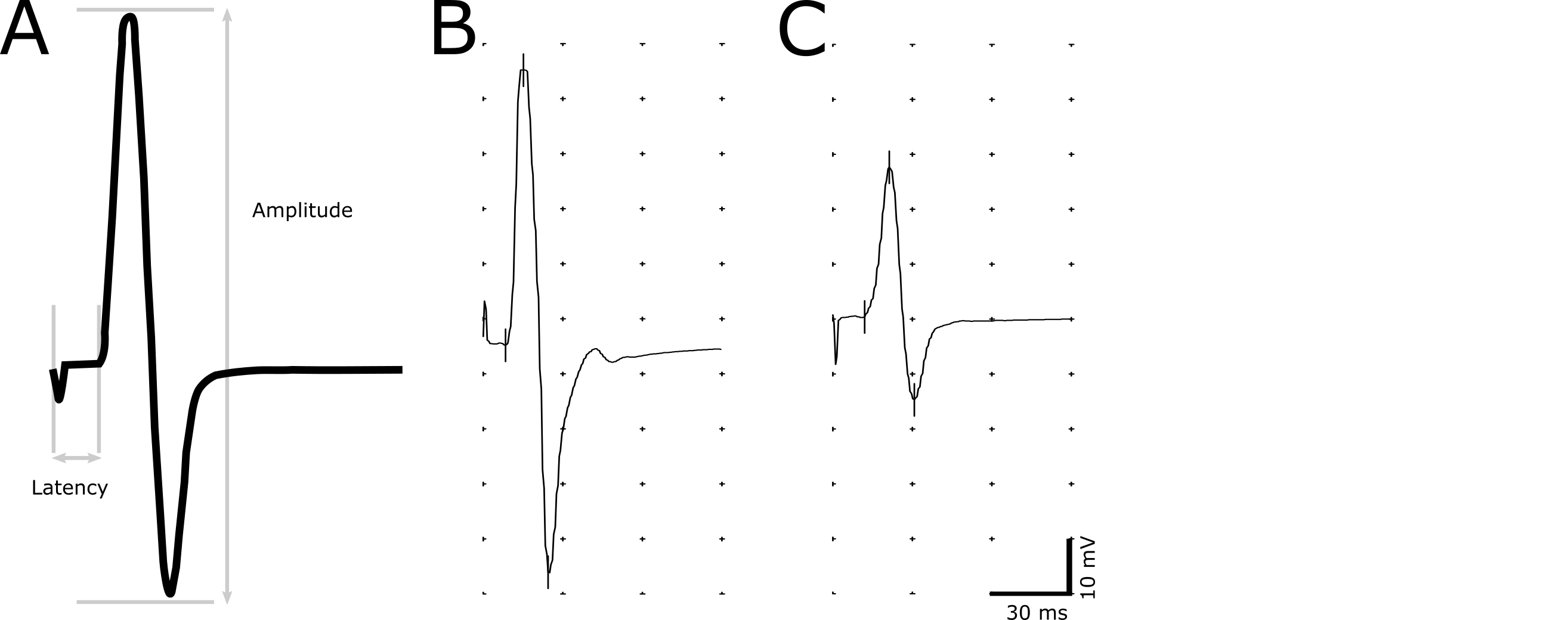
- Utilisez l’outil marqueur pour indiquer les points suivants lors de l’enregistrement : début de la stimulation, l’initiation de la réponse, crête positive et négative maximale (Figure 2).
- Déterminer le temps de latence (en ms) comme un retard de l’ouverture du stimulus à l’initiation de la réponse (Figure 2). Définir l’initiation de la réponse comme le point le plus rapproché où l’amplitude commence à augmenter. La latence permet d’évaluer la démyélinisation dans les axones.
- Mesurer l’amplitude (mV) de la maximale négative à positive maximale (Figure 2). L’ampleur de l’amplitude permet d’établir une corrélation entre le nombre d’axones fonctionnelles.

Figure 2. Une image représentative de la réponse de la CMAP. Une réponse CMAP descriptive indiquant les points utilisés pour le calcul de l’amplitude et la latence (A). Temps de latence est déterminée par le retard de la stimulation à l’apparition de la réponse de la CMAP. Amplitude crête à crête est mesurée entre la maximale négative et la positif de crête de l’onde biphasique. Enregistrements représentatifs d’un animal sain non transgéniques (B) et un animal malade avec des temps de latence prolongée et amplitude réduite (C). S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
- Étant donné que l’emplacement exact des électrodes peut affecter la valeur de résultat de l’enregistrement, remplacer les électrodes et mesurer le nerf même trois fois à l’aide de stimuli supramaximal pour s’assurer que la plus forte réponse est obtenue. Utiliser la moyenne des enregistrements.
Résultats
Des mesures électrophysiologiques des CMAPs utilisant des électrodes aiguilles est une méthode mini-invasive et très sensible pour suivre la fonction neuromusculaire au fil du temps. La technique décrite ici permet l’évaluation de la conduction nerveuse des membres antérieurs chez les souris et ainsi, permet de mieux comprendre la fonctionnalité du nerf.
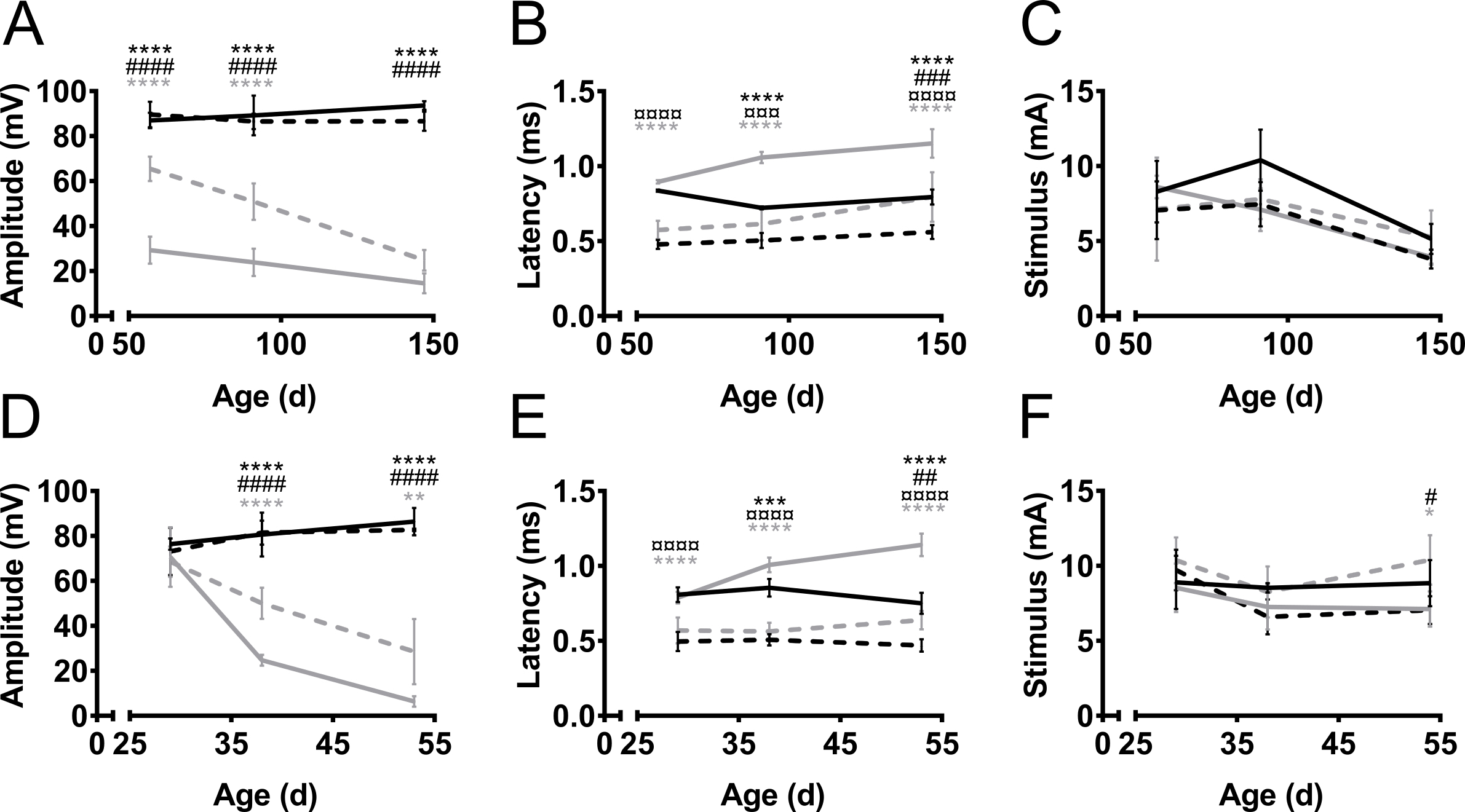
Les amplitudes de la CMAP et les latences ont été mesurés des membres antérieurs et postérieurs au cours de l’évolution de la maladie dans deux modèles de souris de la SLA, SOD1-G93A9 et PrP-hFUS-WT310 (Figure 3) et dans un modèle murin de CMT, C61-PMP2211,12 (Figure 4). ALS souris modèles ont été créés par la surexpression des gènes humains liés à l’ALS, nommément soit muté SOD1 ou type sauvage FUS. Dans les deux modèles, les souris développent ALS ressemblant à une dégénérescence progressive des neurones moteurs, conduisant à la paralysie. Chez les témoins de la même portée non transgéniques, l’amplitude de la CMAP des postérieurs et antérieurs n’a pas changé au fil du temps (Figure 3 a). En revanche, l’amplitude de la CMAP du nerf sciatique depuis le membre postérieur a été diminué de façon spectaculaire chez les souris SOD1-G93A, avant même le début des symptômes vers l’âge de 60 jours (alors que les premiers symptômes moteurs sont habituellement observés à l’âge de trois mois)13 . L’amplitude était 90 mV à cet âge en non-transgéniques de la même portée (non-tg), tandis que chez les souris SOD1-G93A, il était seulement de 30 mV. Il y avait seulement minime nouvelle diminution de l’amplitude que la maladie progressait au stade symptomatique tardif à l’âge de 150 jours. La diminution de l’amplitude de la CMAP, et par conséquent la dégénérescence des axones, a été retardé dans le nerf de plexus brachial de membres antérieurs en comparaison avec le nerf sciatique provenant des membres postérieurs. Dans les membres antérieurs, la progression de la maladie est également plus notable comme le CMAP amplitude a diminué de 70 mV à 30 mV lorsqu’elle est mesurée avant et après la manifestation des déficits moteurs chez ces souris.
Dans le modèle murin de PrP-hFUS-WT3 de la SLA, l’apparition de déficits moteurs commence environ à l’âge de 28 jours10, qui coïncide avec le début du déclin de l’amplitude de la CMAP. Il s’agit d’un modèle de maladie plus accéléré que les souris atteint terminale à l’âge de 65 jours environ. La diminution de l’amplitude de la CMAP s’effectue plus rapidement dans le nerf sciatique des membres postérieurs en comparaison avec le nerf de plexus brachial dans le membre antérieur, ce qui indique une plus tôt de la dégénérescence axonale dans les membres postérieurs (Figure 3D). Cette observation supporte l’observation clinique dans deux de ces modèles de souris que les membres postérieurs sont paralysées notamment plus tôt que les membres antérieurs qui restent fonctionnels jusqu'à les derniers stades de la maladie.
En général, la latence du stimulus à l’initiation du potentiel d’action est plus courte dans les membres antérieurs par rapport aux membres postérieurs (Figure 3 b, E). Il s’agit simplement en raison de la distance la plus courte entre la stimulation et les électrodes d’enregistrement. La latence fournit une indication du niveau de la myélinisation des axones. Notre observation est que les latences de la CMAP sont prolongent au cours de la progression de la maladie dans des modèles murins de la SLA, bien que la SLA n’est pas une maladie démyélinisante. Cela est probablement dû à la perte de plus gros, plus vite mener des axones moteurs.
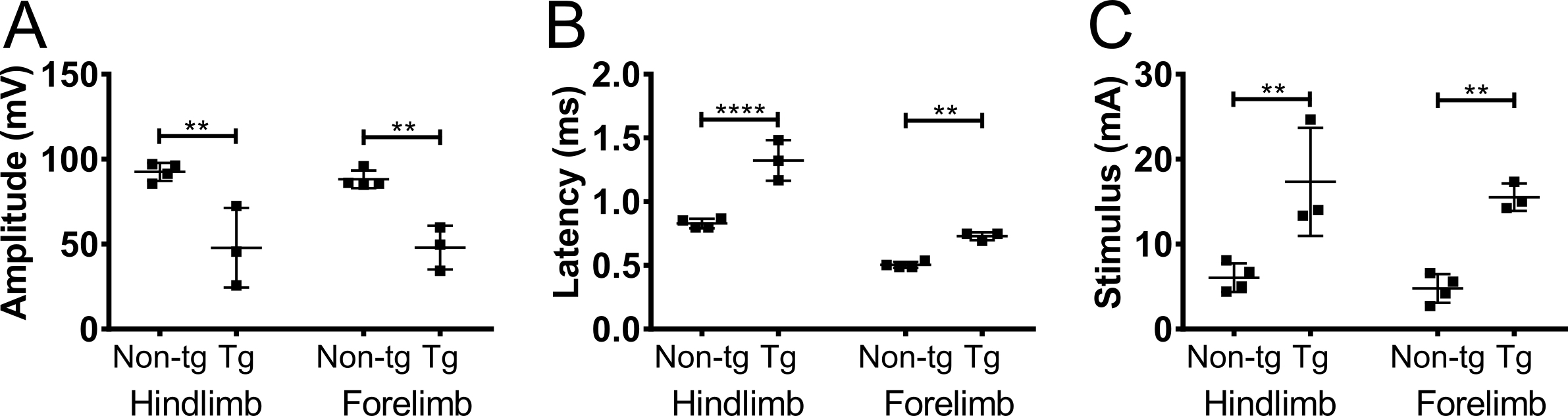
Les C61-PMP22 souris surexprimant 3-4 exemplaires de la PMP22 humain et les souris hétérozygotes récapitulent un phénotype de la maladie très doux CMT1A avec démyélinisation douce et CMAPs réduits, mais avec aucun phénotype visible11,12. 1,5 à 2 années de souris d’âge C61-PMP22, les amplitudes de la CMAP sont réduits et les latences prolongées dans les membres postérieurs et antérieurs (Figure 4). Enregistrements représentant affichage amplitude diminuée et une réponse retardée par rapport à un enregistrement d’un sujet sain sont présentés dans la Figure 2B, respectivement. Les latences de la CMAP dans les membres antérieurs ne sont pas affectés autant que dans les membres postérieurs. Cela est compatible avec les patients CMT1A, car le plus souvent, les patients ont sévèrement réduits ou indétectables CMAPs dans les membres inférieurs en raison de la nature physiopathologique de CMT, comme un trouble longueur14. En outre, le degré de gravité de la maladie est corrélé avec amplitude CMAP, plutôt que de latence ou la conduction de la vitesse, que les amplitudes sont en corrélation avec le degré d’intégrité axonale14,15. Néanmoins, les résultats indiquent que cette méthode est assez sensible pour la détection démyélinisantes processus tels que ceux observés dans CMT1A.
Variation de l’amplitude et la latence était plus faible dans les groupes non transgéniques (coefficient de variation 2-15 % et de 1 à 13 %, respectivement). Dans tous les cas transgéniques, il n’y avait plus de variations dans les mesures (coefficient de variation d’amplitude 8-51 % et latence 1-21 %), qui, très probablement, est causée par des différences dans la progression de la maladie chez les animaux. Dans tous les cas, la variation a été similaire dans les membres antérieurs et postérieurs. La variation dans l’utilisation des aiguilles et des électrodes de surface a été signalée à être similaires16.
L’intensité du stimulus nécessaire ne varie pas beaucoup entre non transgénique et modèles ALS (Figure 3, F). De même, le stimulus nécessaire pour atteindre supramaximal relance dans ces cas était semblable pour fore - et les membres postérieurs et varie de 5 à 12 mA. Dans Sam, l’obligation pour des intensités de stimulation accrue a été reconnu17 et le même phénotype a été observé chez les souris C61-PMP22 (Figure 4). Le phénomène a été expliqué par une impédance électrique accrue du endoneural hypertrophique changements17.
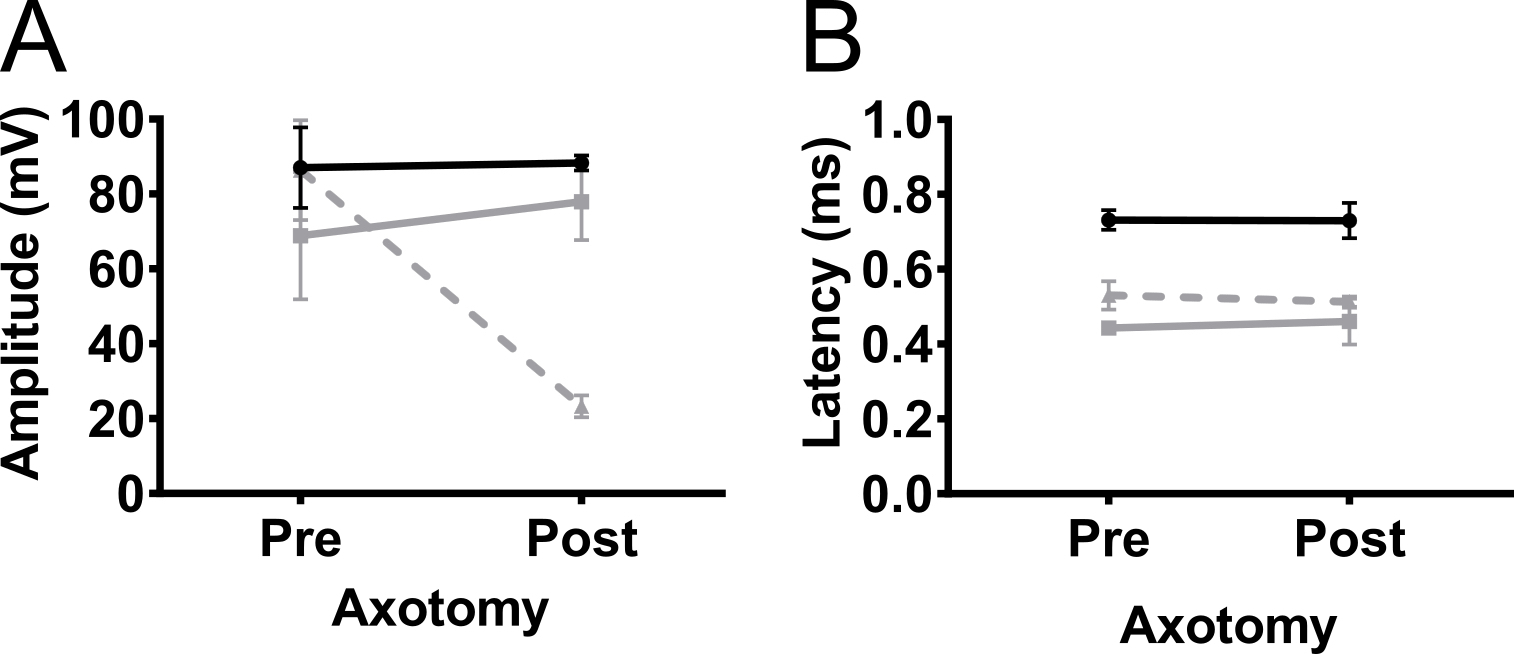
Pour confirmer que l’amplitude de la CMAP enregistrée dans les membres antérieurs était due à une stimulation nerveuse et pas la stimulation musculaire, nous avons effectué une axotomie partielle unilatérale sur le nerf du plexus brachial dans 5 mois non transgéniques C57BL/6Jax souris (mâles et femelles) ( Figure 5). Axotomie réduit l’amplitude de la CMAP de 90 mV à 20 mV, ce qui indique que la plupart des axones ont été déconnectée dans l’opération. Il n’y avait aucun changement dans l’amplitude dans le membre controlatéral antérieur ou dans les membres postérieurs. Ces résultats indiquent fortement que la réponse détectée dans le biceps brachial est due à la stimulation nerveuse et qu’elle ne résulte pas d’une stimulation musculaire.

Figure 3. Amplitude de la CMAP, la latence et stimulus nécessaire au cours de l’évolution de la maladie dans le hind - et ses pattes avant dans la SLA modèles murins. SOD1-G93AA–(C) et des souris transgéniques (tg) PrP-hFUS-WT3 (D–F) et non transgéniques (non-tg) même portée ont été mesurée à l’apparition des symptômes moteurs, au stade symptomatique et dans le processus de phase tardive-symptomatique de la maladie, âge 57, 91 et 147 jours (d) ou à 29, 38 et 53 jours chez les souris SOD1-G93A et de PrP-hFUS-WT3 respectivement. Noir : Non transgéniques postérieur, noir pointillée : membres antérieurs non transgéniques, gris : transgénique postérieur, pointillé gris : membres antérieurs transgéniques. Les résultats sont présentés en moyenne ± SD. Amplitudes (A, D) étaient stables au fil du temps chez les animaux non transgéniques dans les membres antérieurs et postérieurs. Chez les animaux transgéniques, les amplitudes a diminué au cours du processus de la maladie. Temps de latence (B, E) ont été moins touchés par la maladie et des différences importantes ont été observées entre les postérieurs et antérieurs, quel que soit le génotype. La variation dans le stimulus nécessaire (C, F) est minimale dans tous les groupes. Pour SOD1-G93A N = 4 dans tous les groupes sauf tg 147D, N = 3. Pour la PrP-hFUS-WT3 souris dans les groupes d’âge 29, 38 et 53, N est pour non-tg, 4, 5 et 4 et tg 7, 5 et 3, respectivement. Symboles représentent la différence entre les groupes comme suit : * : membre postérieur non-tg vs GÉ du membre postérieur, #: membre antérieur non-tg vs forelimb tg, ¤ : membre postérieur non-tg vs non-tg des membres antérieurs, gris * : membre postérieur tg vs forelimb tg. ANOVA bidirectionnelle avec Tukey du test de comparaisons multiples, * : p < 0,05, ** : p < 0,01, *** : p < 0,001, *** : p < 0,0001. #: p < 0,05, ## : p < 0,01, ### : p < 0,001, ### : p < 0,0001. ¤ : p < 0,05, ¤¤ : p < 0,01, ¤¤¤ : p < 0,001, ¤¤¤ : p < 0,0001. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 4. CMAP amplitude, la latence et stimulus nécessaire dans le hind - et des membres antérieurs chez les souris CMT1A. C61-PMP22 des souris transgéniques (tg) et non transgéniques (non-tg) même portée ont été mesurée à 1,5 à 2 ans. Amplitude (A) a diminué en hind - et les membres antérieurs chez les souris transgéniques. Latence (B) a été prolongé dans toutes les branches chez les souris CMT et même subtil changement dans ses pattes avant a été détecté avec cette mesure. Exigence pour l’intensité de stimulation (C) a été augmentée chez les souris C61-PMP22, qui ressemble à du phénotype détecté chez les patients CMT1A. Les résultats sont présentés comme moyenne ± écart-type, pour non-tg N = 4 et GÉ N = 3. ANOVA bidirectionnelle avec Sidak du test de comparaisons multiples, ** : p < 0,01, *** : p < 0,0001. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 5. Potentiels d’action des membres antérieurs sont causées par une stimulation nerveuse. Pour exclure la possibilité que la réponse observée de la CMAP a été provoquée par la stimulation musculaire, axotomie (partielle) a été réalisée sur le nerf du plexus brachial. CMAP amplitude (A) et la latence (B) ont été enregistrés avant (pre) et 4 jours après l’axotomie du plexus brachial (poster) chez les souris non-transgéniques adultes. Axotomie diminue l’amplitude CMAP, qui indique que la réponse était due à une stimulation nerveuse. Noir : membre postérieur, gris : forelimb controlatérale, gris anéanti : membre antérieur ipsilatéral. Résultats sont présentés en moyenne ± écart-type, N = 2. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
Discussion
Méthodes d’enregistrement sensibles sont essentielles pour évaluer la progression de la maladie et surtout l’efficacité d’une thérapie dans des modèles animaux de troubles neuronaux. Déterminer les CMAPs est une technique électrophysiologique minimalement invasive, qui est régulièrement utilisée dans les cliniques et dans des configurations expérimentales afin d’évaluer la conduction nerveuse dans les troubles neuromusculaires et neuropathique3,18. Nous décrivons ici une nouvelle application pour CMAP enregistrement chez la souris, afin de mesurer la conduction nerveuse du nerf de plexus brachial des pattes antérieures. La méthode présentée permet une évaluation longitudinale plus polyvalente et plus détaillée de la fonction neuronale dans des souris modèles de la neurodégénérescence.
Les électrodes d’aiguille sont légèrement plus invasives que les électrodes de l’anneau et surtout dans les études longitudinales, il faut minimiser les lésions tissulaires. Un inconvénient possible de la méthode est dommage résultant de percer un nerf ou un muscle. Toutefois, après avoir soigneusement sous-cutanée placement des électrodes, les blessures et les perturbations des muscles et des nerfs peuvent être évités. Contrairement à la méthode à l’aide d’électrodes de l’anneau, la méthode présentée ici ne nécessite pas de rasage de la fourrure de grandes parties du corps. En conséquence, il n’y a aucun inconfort ou effet sur la thermorégulation de l’animal.
Le positionnement des électrodes est essentiel pour l’enregistrement correct et systématique des amplitudes de la CMAP et latences. Il est conseillé pour repositionner les électrodes et effectuer deux ou trois mesures sur chaque site pour confirmer que la stimulation maximale et les réponses sont atteints. Enregistrements corrects devraient produire biphasique courbes comme illustré à la Figure 2. Afin de normaliser la méthode, des souris non-transgéniques sans lésion nerveuse sont les meilleurs modèles pour établir l’électrode correctement et systématiquement de positionnement pour une stimulation optimale. Électrodes aiguilles réutilisables sont adaptés à une utilisation répétée si elles sont régulièrement stérilisés, par exemple au glutaraldéhyde pendant 20 minutes entre les animaux et inspecté pour la netteté.
Chez la souris adulte, les amplitudes de la CMAP a enregistré avec la méthode présentée sont typiquement 80-100 mV après stimulation du nerf sciatique et du plexus brachial. C’est notamment plus grand que les réponses mesurées avec des électrodes de l’anneau, car il y a une impédance plus élevée causée par la peau pour les électrodes de l’anneau qui donne des résultats de 20-40 mV8,19,20. Dans des modèles murins de ALS, les amplitudes de la CMAP après stimulation du nerf sciatique ou du plexus brachial dans les membres paralysés diminuent à 10-30 mV. L’ampleur de l’amplitude de la CMAP est plus faible chez les jeunes animaux puisque l’amplitude de la CMAP augmente au cours du développement,21.
La méthode que nous décrivons ici est particulièrement utile dans des modèles murins de la SLA, dans lequel une dénervation et les déficits moteurs, lieu plus tôt dans les membres postérieurs que dans les pattes avant13. En plus de la dénervation, la méthode pourrait détecter réinnervation qui est déterminée comme empêché ou retardé de baisse de l’amplitude de la CMAP. La diminution spectaculaire de l’amplitude de la CMAP dans les muscles des membres postérieurs, déjà à l’âge de l’apparition des symptômes entrave le suivi de la progression de la maladie plus loin ; comme les amplitudes CMAP atteignent des valeurs très faibles au stade précoce de la maladie, ils ne diminuent pas davantage pendant le processus de la maladie. En revanche, la perte axonale progresse à un rythme plus lent dans le nerf de plexus brachial des membres antérieurs et présente une option plus sensible pour mesurer la progression de la maladie sur une plus longue durée de la maladie. En outre, les membres antérieurs moins dégénérées pourraient fournir un site plus puissant pour l’évaluation des approches thérapeutiques qui visent à renforcer la fonction axonale.
Il est clair que la technique présentée offre des possibilités nouvelles pour la caractérisation des souris modèles de maladies neuromusculaires. CMAP recordings avec électrodes aiguilles du nerf sciatique et le plexus brachial est une méthode rapide et reproductible pour évaluer la perte axonale et démyélinisation en hind-ainsi que dans ses pattes avant. La sensibilité de la méthode permet la détection des déficits axonales même avant que les déficits moteurs notables peuvent être enregistrées et permet donc la quantification rapide de ces défauts. En outre, la possibilité de tests répétés réduit le nombre d’animaux nécessaires et fournit un aperçu détaillé de la progression des maladies neuromusculaires et neuropathiques à différents endroits dans un animal.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Cette recherche a été soutenue par la KU Leuven (« Ouverture à l’avenir » et C1), le Fonds pour la recherche scientifique Flandre (FWO-Vlaanderen), la Fondation Thierry Latran l’Association Belge contre les Maladies neuro-Musculaires (AMBM), la dystrophie musculaire Association (MDA), l’Association ALS et l’ALS Liga (Belgique). PVD détient une recherche senior de FWO-Vlaanderen. RP a été pris en charge par des subventions de l’Irlande Central assainissement clinique (CRC) et est actuellement pris en charge l’Université nationale d’Irlande (NUI) et le FWO.
matériels
| Name | Company | Catalog Number | Comments |
| Resuable subdermal needle electrode, Pl/Ir | Technomed | TE/S61-434 | The Needle is 13 mm (0.51") in length, 0.4 mm (27G) in diameter |
| Natus electrodiagnostic system | Natus Neurology | UltraPro S100 | EMG device |
| Synergy | Natus Neurology | version 20.1.0.100 | EMG software for UltraPro S100 |
| Physitem Controller | Rothacher-Medical GmbH | TCAT-2LV | Heating pad |
| combi-vet Base Anesthesia System Digital Flowmeter with TEC 3 Vaporize | Rothacher & Partner | CV 30-301-D | Isoflurane Vaporizer and flowmeter |
| Iso-Vet 1000 mg/g | Piramal Healthcare UK Limited | AP/DRUGS/220/96 | Isoflurane |
| SOD1-G93A mice | The Jackson Laboratory | #002726 | ALS tg and non-tg control littermates, only females |
| PrP-hFUS-WT3 mice | The Jackson Laboratory | #017916 | ALS tg and non-tg control littermates, all groups balanced for males and females |
| C57BL/6Jax mice | The Jackson Laboratory | #000664 | Non-tg mice for axotomy, male and female |
| C61-PMP22 mice | Mouse line was generously donated by Prof. M. Sereda (The Max Planck Institute of Experimental Medicine, Göttingen, Germany). | CMT tg and non-tg control littermates, all groups balanced for males and females |
Références
- Brown, R. H., Al-Chalabi, A. Amyotrophic Lateral Sclerosis. N Engl J Med. 377 (2), 162-172 (2017).
- Prior, R., Van Helleputte, L., Benoy, V., Van Den Bosch, L. Defective axonal transport: A common pathological mechanism in inherited and acquired peripheral neuropathies. Neurobiol Dis. , 300-320 (2017).
- de Carvalho, M., et al. Electrodiagnostic criteria for diagnosis of ALS. Clin Neurophysiol. 119 (3), 497-503 (2008).
- Krajewski, K. M., et al. Neurological dysfunction and axonal degeneration in Charcot-Marie-Tooth disease type 1A. Brain. 123 (Pt 7), 1516-1527 (2000).
- Raynor, E. M., Ross, M. H., Shefner, J. M., Preston, D. C. Differentiation between axonal and demyelinating neuropathies: identical segments recorded from proximal and distal muscles. Muscle Nerve. 18 (4), 402-408 (1995).
- Zielasek, J., Martini, R., Toyka, K. V. Functional abnormalities in P0-deficient mice resemble human hereditary neuropathies linked to P0 gene mutations. Muscle Nerve. 19 (8), 946-952 (1996).
- Arnold, W. D., et al. Electrophysiological Motor Unit Number Estimation (MUNE) Measuring Compound Muscle Action Potential (CMAP) in Mouse Hindlimb Muscles. J Vis Exp. (103), (2015).
- Schulz, A., Walther, C., Morrison, H., Bauer, R. In vivo electrophysiological measurements on mouse sciatic nerves. J Vis Exp. (86), (2014).
- Gurney, M. E., et al. Motor neuron degeneration in mice that express a human Cu,Zn superoxide dismutase mutation. Science. 264 (5166), 1772-1775 (1994).
- Mitchell, J. C., et al. Overexpression of human wild-type FUS causes progressive motor neuron degeneration in an age- and dose-dependent fashion. Acta Neuropathol. 125 (2), 273-288 (2013).
- Robertson, A. M., et al. Comparison of a new pmp22 transgenic mouse line with other mouse models and human patients with CMT1A. J Anat. 200 (4), 377-390 (2002).
- Huxley, C., et al. Correlation between varying levels of PMP22 expression and the degree of demyelination and reduction in nerve conduction velocity in transgenic mice. Hum Mol Genet. 7 (3), 449-458 (1998).
- Turner, B. J., Talbot, K. Transgenics, toxicity and therapeutics in rodent models of mutant SOD1-mediated familial ALS. Prog Neurobiol. 85 (1), 94-134 (2008).
- Manganelli, F., et al. Nerve conduction velocity in CMT1A: what else can we tell. Eur J Neurol. 23 (10), 1566-1571 (2016).
- Cornett, K. M., et al. Phenotypic Variability of Childhood Charcot-Marie-Tooth Disease. JAMA Neurol. 73 (6), 645-651 (2016).
- Jacobson, W. C., Gabel, R. H., Brand, R. A. Surface vs. fine-wire electrode ensemble-averaged signals during gait. J Electromyogr Kinesiol. 5 (1), 37-44 (1995).
- Parker, V., Warman Chardon, J., Mills, J., Goldsmith, C., Bourque, P. R. Supramaximal Stimulus Intensity as a Diagnostic Tool in Chronic Demyelinating Neuropathy. Neurosci J. 2016, 6796270 (2016).
- Benoy, V., et al. Development of Improved HDAC6 Inhibitors as Pharmacological Therapy for Axonal Charcot-Marie-Tooth Disease. Neurotherapeutics. 14 (2), 417-428 (2017).
- Xia, R. H., Yosef, N., Ubogu, E. E. Dorsal caudal tail and sciatic motor nerve conduction studies in adult mice: technical aspects and normative data. Muscle Nerve. 41 (6), 850-856 (2010).
- Srivastava, A. K., et al. Mutant HSPB1 overexpression in neurons is sufficient to cause age-related motor neuronopathy in mice. Neurobiol Dis. 47 (2), 163-173 (2012).
- Arnold, W. D., et al. Electrophysiological Biomarkers in Spinal Muscular Atrophy: Preclinical Proof of Concept. Ann Clin Transl Neurol. 1 (1), 34-44 (2014).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.