
Method Article
Visualiser le nœud et la notochorde plaque en Gastrulating des embryons de souris à l’aide de la microscopie électronique à balayage et toute monture Immunofluorescence
Dans cet article
Résumé
Le nœud et la plaque de la notochorde sont transitoires organisateurs dans le développement des embryons de souris qui peuvent être visualisées à l’aide de plusieurs techniques de signalisation. Ici, nous décrivons en détail comment effectuer deux des techniques pour étudier leur structure et la morphogenèse : 1) microscopie électronique (MEB) ; et 2) support entier immunofluorescence (WMIF).
Résumé
L’embryon de souris après l’implantation subit des changements de forme majeure après le début de la gastrulation et morphogenèse. Une caractéristique de la morphogenèse est la formation des organisateurs transitoires, le nœud et la notochorde plaque, des cellules qui ont traversé la ligne primitive. La formation adéquate de ces centres de signalisation est indispensable pour l’élaboration de ce plan de corps et les visualiser, les techniques sont d’un grand intérêt pour les biologistes du développement de la souris. Le nœud et la plaque de la notochorde se trouvent sur la face ventrale du gastrulating autour du jour embryonnaire (E) 7.5 du développement des embryons de souris. Le nœud est une structure en forme de coupe, dont les cellules possèdent un seul mince CIL chaque. La localisation sous-cellulaire appropriée et la rotation des cils dans la fosse de nœud détermine l’asymétrie gauche-droite. Les cellules de la notochorde plaque possèdent également des cils unique bien que plus courtes que celles des cellules nœud. La plaque de la notochorde forme la notochorde qui agit comme un important organisateur de signalisation pour somitogenèse et structuration neurale. Parce que les cellules du nœud et notochorde plaque sont transitoirement présents sur la surface et possèdent des cils, ils peuvent être visualisées à l’aide de la microscopie électronique à balayage (SEM). Parmi les autres techniques permettant de visualiser ces structures au niveau cellulaire est support entier immunofluorescence (WMIF) en utilisant les anticorps contre les protéines qui sont fortement exprimés dans le nœud et la plaque de la notochorde. Dans ce rapport, nous décrivons nos protocoles optimisés pour exécuter SEM et WMIF du nœud et plaque notochorde dans des embryons de souris en développement pour aider à l’évaluation de la forme des tissus et une organisation cellulaire de type sauvage et embryons mutants de la gastrulation.
Introduction
Gastrulation et les mouvements morphogénétiques qui l’accompagne sont indispensables pour former l' embryon de souris1. Les changements dans l’organisation au cours de la morphogenèse et forme cellulaire dictent des données de position pour régler le sort de la cellule et permettent également les voies de signalisation qui s’ensuivit précisément leurs fonctions afin de diversifier le nouvellement formé des couches de germe1. La formation d’organiser les structures transitoires et de signalisation des centres comme le nœud et la notochorde est essentielle pour l’exécution du programme de perfectionnement2. Biologistes du développement ont utilisé une variété de techniques pour étudier la morphogenèse de ces structures, plus notables dont est l’utilisation des cellulaires reporters et direct ex vivo imagerie pour suivre la dynamique des comportements cellulaires et subcellulaires2 ,3,4. Dans ce rapport, nous nous concentrons sur décrivant les détails de nos protocoles optimisés pour deux de ces techniques : scanning electron microscopy (SEM) et immunofluorescence support entier (WMIF), qui ont été et sont toujours joué un rôle important dans l’étude de la morphogénèse du nœud et la plaque notochorde, le précurseur de la notochorde.
Le nœud embryonnaires de souris est une coupe en forme de larme de cellules qui se trouve sur la surface ventrale de l’embryon de souris autour du début de pli tête tardivement au cours de la gastrulation et morphogenèse (jour embryonnaire, E7.5-E8)2,5, 6,7. La plaque de la notochorde, morphologiquement antérieurement découle le noeud3. Chaque cellule dans le nœud et la plaque de la notochorde se caractérise par un seul CIL qui fait saillie vers l’extérieur, qui est plus long dans les cellules de noeud, mais dont la longueur varie selon le stade de développement2. La rotation des cils dans la fosse de nœud s’est avérée être important pour la signalisation qui détermine asymétrie gauche-droite4. La plaque de la notochorde est le précurseur de la notochorde, le centre de signalisation qui est important pour la structuration des somites adjacents et le sus-jacente de tube neural3.
À cause des attributs de localisation (surface), forme (coupe) et possédant des structures cellulaires externes distincts (cils), SEM a été utilisé traditionnellement pour visualiser le nœud et la plaque de la notochorde et étudier leur structure et la formation2, 7. SEM est également utilisé pour étudier les changements dans la structure du nœud lui-même ou les cils sur ses cellules dans les mutations qui affectent la gastrulation, morphogenèse, ainsi que cils formation8,9,10. SEM est une technique qui utilise un faisceau focalisé d’électrons pour interroger l’ultrastructure topologique de la surface extérieure des matériaux tels que les spécimens biologiques11. L’échantillon est généralement fixe, séché et puis par pulvérisation cathodique-plaqués de métaux pour l’observation sous un microscope électronique à balayage comme on décrit à l’étape 1.
WMIF est une technique de coloration pour visualiser des produits de gène, comme les protéines, en trois dimensions (3D). WMIF des tissus, organes ou organismes entiers même fournit des informations spatiales sur la distribution du signal et la forme de la structure résultante en 3D. La technique repose sur la fixation de l’échantillon, puis il coloration avec fluorescents conjugués. Des embryons de souris ~ E7.5 sont petites et transparentes et donc idéal pour les protocoles WMIF visualiser le nœud et la plaque de la notochorde. Par exemple, le facteur de transcription Barchyury (T) est exprimé dans les noyaux du nœud et plaque de la notochorde et dans une moindre mesure dans la ligne primitive, autour de E7.5-E8 du développement embryonnaire et bon travail anticorps contre T de WMIF sont dans le commerce disponibles et faire la procédure de marquage possible. Les cellules du nœud et notochorde plaque sont aussi caractérisés par des surfaces apicales rétrécies, qui font face à l’extérieur et donc peuvent être colorés avec la phalloïdine conjugué fluorescence repère F-actine à la constriction apicale. En utilisant ces réactifs comme exemples, la combinaison de T et F-actine, coloration de WMIF fournit une représentation du nœud et plaque notochorde en 3D dans des embryons de souris gastrulating comme nous le montrer dans l’étape 2, 8. Cependant, marqueurs des cils, comme ARL13B ou acétylée tubuline, ainsi qu’autres marqueurs du nœud et plaque notochorde, tels que FOXA2, permet également d’effectuer WMIF embryons en développement les souris3,4.
Nous avons montré que striatin-interacting protein 1 (STRIP1) est essentielle pour la gastrulation normale et morphogenèse dans l' embryon de souris8. STRIP1 est un composant essentiel de l’interaction striatin phosphatases et complexes de kinases (STRIPAK), qui nous et autres avons impliquée dans l’actine cytosquelette organisation8,12. Un défaut majeur chez les embryons mutants Strip1 est dans la formation du mésoderme axial (nœud et notochorde plaque) et l’extension de l’axe antéro-postérieur de corps. Nous avons utilisé des SEM et WMIF pour analyser le nœud et la notochorde plaque de type sauvage (WT) et d’embryons de mutant Strip1 que nous montrons dans les Résultats de représentant et les chiffres correspondants.
Protocole
Toutes les expériences impliquant des expérimentations animales ont été approuvées par les autorités responsables du Nord-Westphalie Rhein (LANUV-NRW).
1. microscopie électronique du nœud embryonnaires de souris
- Sacrifier la souris femelle enceinte à ~ E7.5 (stade de segments de 2-4) par dislocation cervicale. Une explication détaillée avec schémas des étapes 1.1 - 1.7 est disponible à l’embryon de souris laboratoire manuels13.
- Ouvrir l’abdomen à travers la peau et les mésentères et enlever l’utérus à l’aide de ciseaux et pinces fines.
- Rincer l’utérus brièvement dans l’eau distillée et placez-le dans un petit plat de Petri propre (6 cm) contenant 1 x solution saline tamponnée au phosphate (PBS).
- Une binoculaire et en utilisant des pinces fines, enlever les muscles utérins pour libérer les deciduae individuels ou les sites d’implantation.
- Tenez chaque caduque avec une paire de pinces et utilisez l’autre paire de faire une incision longitudinale de pleine épaisseur entre la partie rouge (futur placenta) et la partie blanche (où l’embryon se trouve). Faire des perforations superficielles verticalement le long de la partie blanche de la caduque contigus par l’incision. Séparez la caduque horizontalement en deux moitiés et évidez soigneusement l’embryon dans la partie blanche de la caduque.
- Transfert de l’embryon d’une nouvelle boîte de Pétri (35 mm) avec frais PBS stérile filtré. Répétez pour tous les deciduae/embryons.
- Enlever la membrane de Reichert, une membrane relativement opaque enveloppant l’embryon, de chaque embryon en les taquinant loin comme une chaussette à partir du cône d’ectoplacental (site d’implantation rougeâtre). Pour le génotypage, prendre un petit morceau (~ 0,1 mm2) du sac vitellin à ce stade.
- Sous une hotte chimique et portant une protection adéquate (gants), transférer les embryons à EM grade fixateur composé de glutaraldéhyde à 2,5 % dans du PBS stérile filtrée dans un tube de microcentrifuge (1,5 mL) à la température ambiante. Difficulté les embryons pendant la nuit à 4 ° C.
- Retirez délicatement le fixateur de glutaraldéhyde de tube sans toucher les embryons et jeter dans une poubelle appropriée. Lavez les embryons trois fois dans du PBS stérile filtré, de 15 min chacun, à la température ambiante.
- Déshydrater les embryons dans une série d’éthanol de 5 min chacun : 50 %, 70 %, 85 % et trois fois à 100 % ou éthanol absolu. Conserver les embryons à −20 ° C dans de l’éthanol ou passez directement à l’étape suivante.
- Transfert des embryons dans l’éthanol à paniers pour séchage (CPD) dans une machine sèche point critique des points critiques. Remplir la chambre à l’éthanol pour couvrir les paniers complètement.
- Échanger de l’éthanol en rinçant soigneusement avec du liquide CO2 pour dix fois à 10 ° C. Vidange liquide de CO2 après la dernière étape jusqu'à ce que la chambre est à moitié pleine. Chauffer jusqu'à 40 ° C jusqu'à ce que la pression atteint 80 bars (point critique) et le liquide de CO2 passe au gaz. Attendez pendant 10 min, puis souffler lentement le gaz pendant environ 45 min.
- Comme une alternative plus facile de CPD pour le séchage, ajouter hexaméthyldisilazane (HMDS) selon un ratio de 1:1 pour les embryons dans l’éthanol pendant 30 min. Puis transférez les embryons au HMDS pur pendant 30 min. Retirer les embryons de liquide à l’aide d’une pipette et laissez-les sécher pendant 30 min.
Remarque : Les deux méthodes de séchage a travaillé aussi bien dans nos mains. - Utilisez une brosse fine pour monter les embryons séchés avec la face ventrale (nœud) vers le haut sur un talon de SEM avec ruban adhésif double face.
- Insérez les stubs avec les embryons dans une machine de revêtement par pulvérisation cathodique pour revêtement de particules d’or, ce qui est préférable pour charger les cils long et minces. Appliquer une couche de 120-150 Å ; le temps dépend du courant, qui varie avec chaque échantillon.
- Placer les talons enrobés avec des embryons dans un microscope SEM, vide et observer le nœud embryonnaire et les cellules de la notochorde plaque cils à grossissements allant de 1000 X à 15, 000 X.
2. toute monture Immunofluorescence des souris noeud et notochorde plaque
- À l’aide de PBS glacée avec 0,05 % Tween 20 (PBSTw), suivez les étapes ci-dessus de 1,1 à 1,7 pour retirer les embryons à E7.75 et placez-les dans PBSTw en un 35 mm boîte de Pétri sur glace.
- Sous une hotte chimique et portant une protection adéquate (gants), transférer les embryons dans une solution de fixation de paraformaldéhyde à 4 % dans du PBS dans un tube de microcentrifuge. Difficulté les embryons pendant la nuit à 4 ° C.
- Retirez délicatement le fixateur de paraformaldéhyde de tube sans toucher les embryons et jeter dans une poubelle appropriée. Lavez les embryons de trois fois dans du PBS contenant 0,2 % Triton X-100 (Ref) de 5 min chacun, à la température ambiante. Exécuter toutes les laver et les prochaines étapes de l’incubation sur un agitateur oscillant.
- Supprimez le dernier lavage et blocage solution contenant Ref avec 10 % de sérum d’inactivés par la chaleur (à partir de l’espèce hôte de l’anticorps secondaire). Bloc de 2 h à une nuit (ou plus) sur un nutator à 4 ° C.
- Retirer le blocage et ajouter ~ 1 mL de l’anticorps primaire dilué en bloquant la solution, par exemple un anticorps anti-T à la dilution de 1/500. Incuber pendant la nuit (ou plus) sur un nutator à 4 ° C.
- Supprimer l’anticorps primaire et l’enregistrer pour une utilisation ultérieure en ajoutant l’azoture de sodium à une concentration finale de 0,02 % (1 µL de stock de 20 % à 1 mL de solution d’anticorps). L’anticorps peut être réutilisé ~ 10 fois. Rincez les embryons deux fois avec REF et lavez-les ensuite trois fois de 30 min chacun, sur un nutator à 4 ° C.
- Remplacer le lavage avec un anticorps secondaire conjugué fluorescence, contre les espèces hôtes anticorps primaire, dilués à ~ 1/1000 une nuit (ou plus) sur un nutator à 4 ° C.
- Retirez l’anticorps secondaire et rincez deux fois avec la Ref, puis laver trois fois pendant 30 min avec Ref.
- Remplacer le dernier lavage avec Ref contenant 1/500 conjugué fluorescence phalloïdine, à colorer F-actine et 1/1000 DAPI, pour souiller des noyaux, pendant 1 h à température ambiante.
- Rincer deux fois dans la Ref et laver une fois avec Ref pendant 30 min à température ambiante.
- Remplacez Ref par PBS et laisser les embryons sur glace. Préparer les diapositives propre-chargé positivement (60 x 24 mm) et lamelles couvre-objet (24 x 24 mm) et supports de montage de glycérol-base aqueuse, par exemple, 90 % de glycérol dans 1xPBS et un réactif antifade, pour monter les embryons.
- Mettre deux morceaux de ruban adhésif transparent à une distance de ~ 15 mm les uns des autres sur la partie claire de la diapositive. Cela va créer suffisamment d’espace 3D (dans la dimension Z) qui permettrait l’aplatissement des embryons, mais pas complètement empiète sur eux.
- Sous un microscope à dissection, déplacer délicatement les embryons à l’aide d’une pipette de P200 coupée (espace suffisant pour permettre à l’embryon d’être transférés et non endommagé) à la diapositive.
- À l’aide de pinces fines, faire deux incisions complets sur les faces latérales du sac vitellin se dérouler l’embryon. Placer l’embryon avec la face ventrale (nœud et plaque notochorde) vers le haut (dorsale du tube neural vers le bas de la diapositive).
- Ajouter 50 µL de montage des médias sur l’embryon. Placez les 4 et 5 embryons par diapositive. Ajouter un peu de montage des médias sur le côté de la lamelle qui tout d’abord appuyer sur la lame (côté haut ou en bas), puis placez-la à cheval sur les deux morceaux de ruban et laissez-la s’enfoncer lentement sur les embryons à l’aide de la pince fine ou une aiguille tordue fine tout en évitant de créer des bulles d’air.
- Nettoyer les supports de montage excès à l’aide d’un chiffon absorbant. Veillez à ne pas déplacer la lamelle couvre-objet dans le processus.
- En utilisant une quantité généreuse de vernis à ongles, sceller les côtés de la lamelle sans le déplacer.
- Observer sous un microscope confocal à balayage.
Résultats
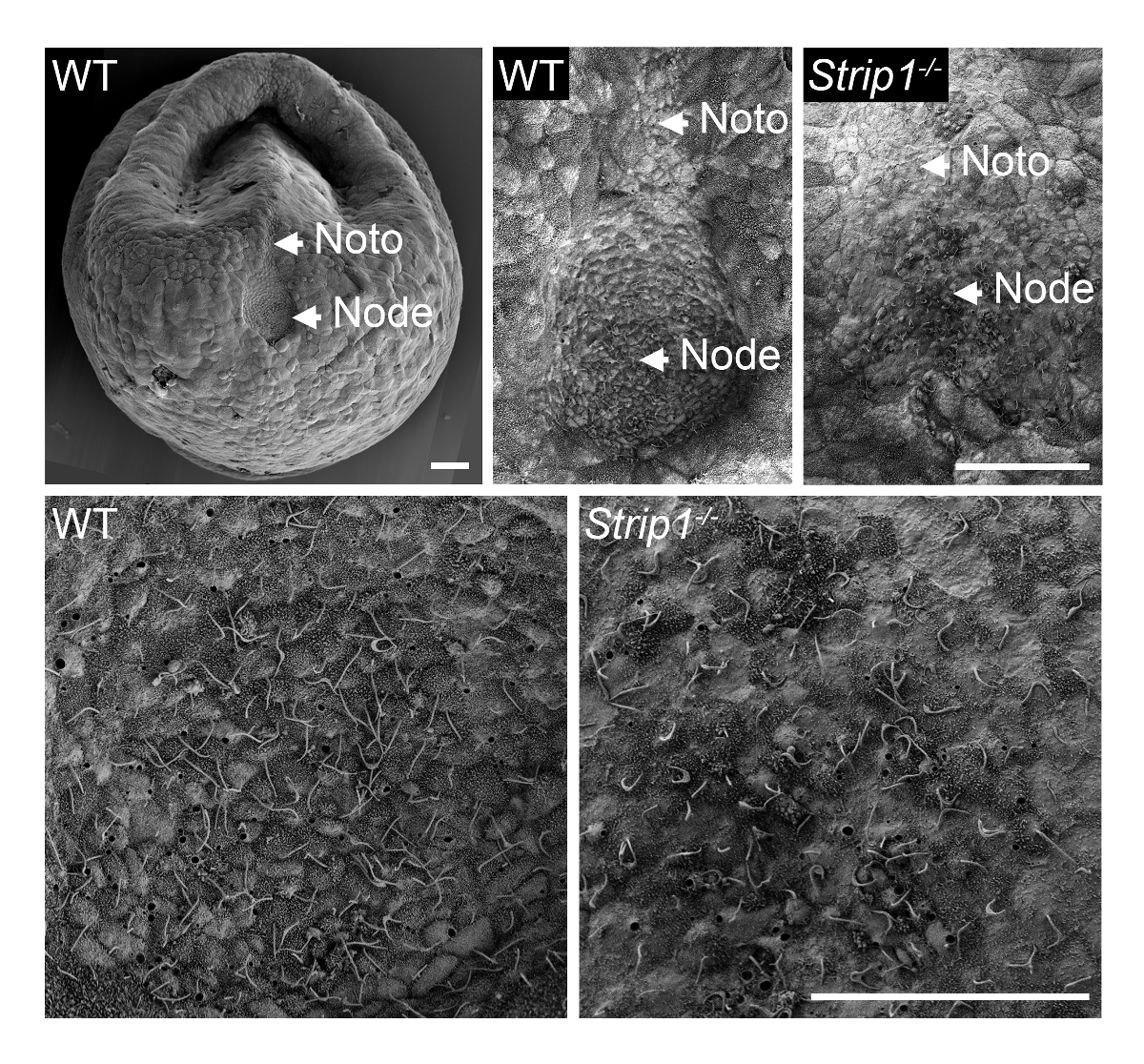
Afin d’examiner la formation du nœud dans des embryons mutants WT et Strip1 au ~ E7.5, nous avons utilisé SEM tel que décrit à l’étape 1 et illustré à la Figure 18. Les détails ultrastructurales de la topologie de l’extérieur à l’aide de SEM étaient très instructifs et il était immédiatement évident qu’à la différence du nœud puits-formées dans des embryons WT, les embryons mutants un nœud aplati et irrégulière. Un grossissement supérieur des embryons ne présentaient les cils caractéristiques des cellules de noeud qui eux identifié sans ambiguïté. La densité apparente inférieure des cils chez le mutant pourrait être attribuable à la perte de structure de stands de nœud et courbure ou un nombre inférieur de cellules de noeud. La notochorde plaque qui apparaît émane du nœud était également irrégulier chez les embryons mutants. Ils étaient identifiables avec leurs cils plus courts. Par conséquent, il importait SEM révéler les défauts de la morphogenèse de nœud Strip1 mutants8. Nous avons également utilisé des SEM dans des études antérieures à démontrer l’absence de cils dans le nœud embryonnaire des mutants qui ne disposaient pas de centrioles, qui fournissent le modèle pour cils9.
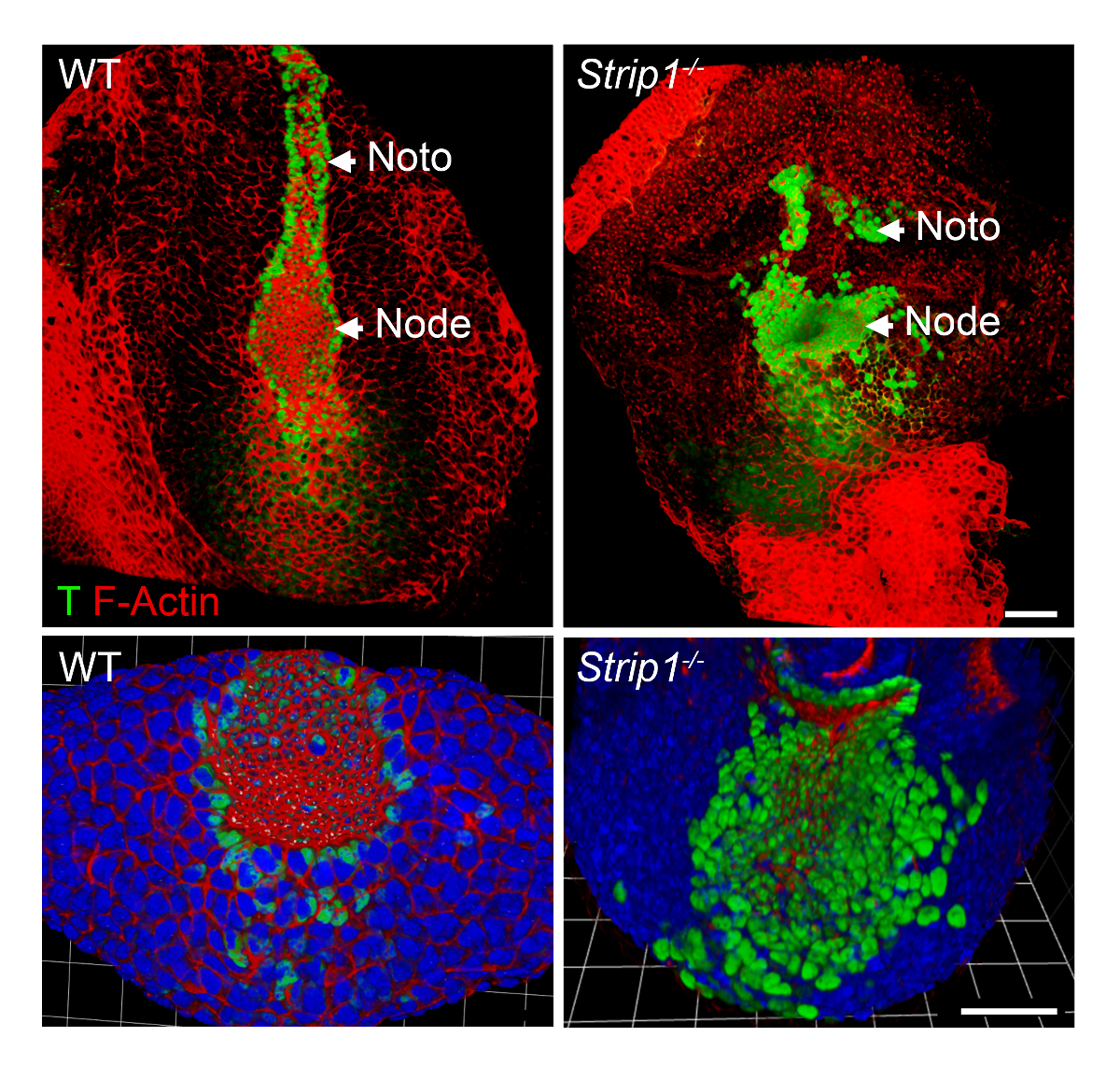
Afin d’étudier les défauts mésoderme axial de formation chez les embryons mutants Strip1 au niveau cellulaire, nous avons utilisé WMIF tel que décrit à l’étape 2 et illustré à la Figure 2. En utilisant cette technique, le nœud et la plaque de la notochorde, ont été facilement identifiés par actine F et T la coloration. Nœud WT et les cellules de la notochorde plaque ont étranglée apicales domaines où actine F s’est enrichie, et la coloration des noyaux T était évidente. La plaque de la notochorde étendu rostralement dans le WT mais était courte et irrégulière chez le mutant. Les données montrent que les organisation F-actine est anormale dans les différentes couches de germe des embryons mutants dont le mésoderme axial8. Ainsi, WMIF a contribué à étudier les vices du nœud et la formation de la notochorde plaque en Strip1 embryons mutants.

Figure 1 . Microscopie électronique à balayage révèle les défauts dans la morphogenèse des nœuds dans des embryons de souris mutantes Strip1 . (En haut) Analyses de SEM de WT et Strip1 mutants nœuds embryonnaires ventrales et notochorde plaques (Noto)8. Un exemple d’une image de faible grossissement d’un embryon de WT est montré sur la gauche. (En bas) Grossissements du Centre des nœuds montré sur le dessus, révélant le monocilia longue saillie sur les cellules de noeud. Antérieure est en place dans tous les panneaux. Barreaux de l’échelle : 30 µm. s’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}

Figure 2 . Support entier immunofluorescence montre le nœud anormale et notochorde au niveau cellulaire chez les embryons mutants Strip1 . (En haut) Rendu 3D ventral (logiciel Volocity) de WMIF sur des embryons mutants WT et Strip1 en utilisant une combinaison de phalloïdine conjugué à fluorescence (F-actine, rouge) et la souillure des anticorps (vert) de T. (En bas) Plus d’exemples de la coloration ci-dessus en se concentrant sur le nœud avec zoom supérieure et y compris DAPI. Antérieure est en place dans tous les panneaux. Barreaux de l’échelle : 30 µm. s’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
{kind=link}
Discussion
Dans ce travail, nous démontrons comment faire SEM et WMIF pour visualiser le nœud embryonnaires de souris et la plaque de la notochorde. La petite taille des embryons de souris gastrulating ~ E7.5 et la présence de ces structures sur la surface les rendent idéales pour étudier à l’aide des techniques décrites2,7,8. La disponibilité de bons anticorps, comme les marqueurs T et les cils, donne des informations 3D excellentes à l’aide de WMIF sur la structure, l’organisation et la formation de ces organisateurs embryonnaires essentiel8.
Parce que le développement embryonnaire chez la souris progresse à un rythme très rapid et le nœud et la plaque de la notochorde ne sont transitoirement présents sur la surface de l’embryon, le moment est essentiel pour le succès de ces expériences2,3. Par exemple, 2-4 somite embryons sont bonnes pour l’analyse de SEM d’une fosse de nœud mature avec longs cils. Chez les embryons de beaucoup antérieures ou ultérieures (par exemple, de 12 h avant ou après), le nœud peut-être pas présent à la surface. WMIF est un peu plus flexible à cet égard, mais les structures elles-mêmes sont également transitoires pendant le développement et le calendrier dépend dans ce cas les intérêts des chercheurs.
La pureté des réactifs est également essentielle pour le succès de ces techniques, en particulier dans la détection de l’ultrastructure par SEM. Tiny les impuretés qui se collent aux embryons généralement entraîner des artefacts énormes.
Nous avons testé deux méthodes différentes de fixation de l’embryon pour SEM l’un à l’aide de fixateur de Karnovsky moitié (glutaraldéhyde à 2,5 %, 2 % paraformaldéhyde et 0,1 M cacodylate buffer) et une plus simple glutaraldéhyde à 2,5 % dans du PBS 1 x. Nous préférons utiliser le glutaraldéhyde et fixateur de PBS comme décrit à l’étape 1, mais nous et autres avons également utiliser le fixateur de Karnovsky la moitié avec succès pour SEM.
Nous avons également comparé deux méthodes de séchage les embryons pour SEM et trouvé aucune différence dans la qualité de l’échantillon en utilisant un point critique sèche ou HMDS tel que décrit à l’étape 1 et signalée ailleurs14.
Pour l’étape 2, nous avons testé l’incorporation des embryons après les étapes de lavage final dans l’agarose de fusion faible 1 % monté sur un plat de fond en verre de 35 mm et puis la garniture avec ~ 10 µL de milieu de montage. Cette méthode d’incorporation fonctionne et préserve la structure 3D d’origine de l’embryon et des structures connexes ; Toutefois, un microscope multiphoton est nécessaire à l’image de l’échantillon car un microscope confocal ordinaire ne peut pas atteindre aussi profond dans les embryons intacts (~ 1 mm).
Nous croyons que ces deux méthodes donne des informations complémentaires sur la structure du nœud et la plaque de la notochorde au cours du développement normal et chez les mutants qui présentent des défauts dans la formation de ces structures.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
H.B. est pris en charge par financement de démarrage de la faculté de médecine et de la SFB829 de l’Université de Cologne. C.X. est pris en charge par DFG accorder BA 5810/1-1. Nous tenons à remercier les installations d’imagerie de la CECAD research center et le Memorial Sloan Kettering Cancer Center (New York, USA). Nous remercions Joaquín Grego-Bessa (espagnol National Center for Cardiovascular Research, Madrid, Espagne) pour sa perspicacité sur les embryons de montage pour WMIF.
matériels
| Name | Company | Catalog Number | Comments |
| 1,1,1,3,3,3 Hexamethyldisilazane (HMDS) | Carl Roth | 3840 | |
| Anti-T antibody | R&D Systems | AF2058 | |
| Critical Point Dryer | Blazers Union | CPD 020 | |
| DAPI | AppliChem | A4099,0005 | |
| Glutardialdehyde solution 25% | Merck | 1042390250 | |
| Triton X-100 | Sigma Aldrich | X100-100ML | |
| Tween 20 | AppliChem | A4974,0500 | |
| SEM coating unit PS3 | Agar Aids for Electron Microscopy | PS3 | |
| SEM microscope Quantum FEG 250 | ThermoFisher Scientific (FEI) | Quantum FEG 250 |
Références
- Rivera-Pérez, J. A., Hadjantonakis, A. K. The dynamics of morphogenesis in the early mouse embryo. Cold Spring Harbor Perspectives in Biology. , (2015).
- Lee, J. D., Anderson, K. V. Morphogenesis of the node and notochord: The cellular basis for the establishment and maintenance of left-right asymmetry in the mouse. Developmental Dynamics. 237 (12), 3464-3476 (2008).
- Balmer, S., Nowotschin, S., Hadjantonakis, A. K. Notochord morphogenesis in mice: Current understanding and open questions. Developmental Dynamics. 245 (5), 547-557 (2016).
- Yoshiba, S., et al. Cilia at the node of mouse embryos sense fluid flow for left-right determination via Pkd2. Science. , (2012).
- Jurand, A. Some aspects of the development of the notochord in mouse embryos. Journal of Embryology and Experimental Morpholog. 32 (1), 1-33 (1974).
- Poelmann, R. E. The head-process and the formation of the definitive endoderm in the mouse embryo. Anatomy and Embryology (Berl). , 41-49 (1981).
- Sulik, K., et al. Morphogenesis of the murine node and notochordal plate. Developmental Dynamics. 201 (3), 260-278 (1994).
- Bazzi, H., Soroka, E., Alcorn, H. L., Anderson, K. V. STRIP1, a core component of STRIPAK complexes, is essential for normal mesoderm migration in the mouse embryo. Proceedings of the National Academy of Sciences of the United States of America. 114 (51), 10928-10936 (2017).
- Bazzi, H., Anderson, K. V. Acentriolar mitosis activates a p53-dependent apoptosis pathway in the mouse embryo. Proceedings of the National Academy of Sciences of the United States of America. 111 (15), 1491-1500 (2014).
- Huangfu, D., Liu, A., Rakeman, A. S., Murcia, N. S., Niswander, L., Anderson, K. V. Hedgehog signalling in the mouse requires intraflagellar transport proteins. Nature. 426 (6962), 83-87 (2003).
- McMullan, D. Scanning electron microscopy 1928-1965. Scanning. 17 (3), 175-185 (2006).
- Bai, S. W., et al. Identification and characterization of a set of conserved and new regulators of cytoskeletal organization, cell morphology and migration. BMC Biology. 9, (2011).
- Behringer, R., Gertsenstein, M., Vintersen Nagy, K., Nagy, A. Manipulating the Mouse Embryo: A Laboratory Manual, Fourth Edition. Cold Harb Lab Press. , (2014).
- Braet, F., De Zanger, R., Wisse, E. Drying cells for SEM, AFM and TEM by hexamethyldisilazane: a study on hepatic endothelial cells. Journal of Microscopy. 186, Pt 1 84-87 (1997).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.