
Method Article
Nettoyage sélectif des nématodes sauvages de Caenorhabditis pour enrichir les bactéries du microbiome intestinal
Dans cet article
Résumé
Les nématodes sauvages de Caenorhabditis sont associés à de nombreux microbes, souvent dans la lumière intestinale ou infectant l’intestin. Ce protocole détaille une méthode pour enrichir les microbes inculturables colonisant l’intestin, en tirant parti de la résistance de la cuticule de dauer.
Résumé
Caenorhabditis elegans (C. elegans) s’est avéré être un excellent modèle pour étudier les interactions hôte-microbe et le microbiome, en particulier dans le contexte des intestins. Récemment, l’échantillonnage écologique de nématodes sauvages de Caenorhabditis a découvert un large éventail de microbes associés, y compris des bactéries, des virus, des champignons et des microsporidies. Beaucoup de ces microbes ont des phénotypes de colonisation ou d’infection intéressants qui justifient une étude plus approfondie, mais ils sont souvent inculturables. Ce protocole présente une méthode pour enrichir les microbes intestinaux souhaités chez C. elegans et les nématodes apparentés et réduire la présence des nombreux microbes contaminants adhérant à la cuticule. Ce protocole consiste à forcer les animaux au stade de développement et à utiliser une série de lavages antibiotiques et détergents pour éliminer la contamination externe. Comme les animaux dauer ont des changements physiologiques qui protègent les nématodes des conditions environnementales difficiles, tous les microbes intestinaux seront protégés de ces conditions. Mais, pour que l’enrichissement fonctionne, le microbe d’intérêt doit être maintenu lorsque les animaux se développent en dauers. Lorsque les animaux quittent le stade de dauer, ils se propagent individuellement en lignes individuelles. Les populations F1 sont ensuite sélectionnées pour les microbes ou les phénotypes d’infection souhaités et contre la contamination visible. Ces méthodes permettront aux chercheurs d’enrichir les microbes inculturables dans la lumière intestinale, qui font partie du microbiome naturel de C. elegans et des agents pathogènes intestinaux intracellulaires. Ces microbes peuvent ensuite être étudiés pour la colonisation ou les phénotypes d’infection et leurs effets sur la condition physique de l’hôte.
Introduction
L’organisme modèle génétique C. elegans est un excellent système in vivo pour étudier les interactions hôte-microbe1,2. Ils ont une physiologie relativement simple par rapport aux autres animaux, mais une grande partie de leur biologie cellulaire est fondamentalement similaire à celle des mammifères, ce qui en fait un bon modèle pour la recherche biologique1,3,4. De plus, ils sont microscopiques, faciles à entretenir et restent transparents tout au long de leur courte durée de vie. Ces propriétés permettent des études rapides sur les mécanismes régissant les interactions hôte-microbe et la visualisation de l’infection in vivo et de la colonisation des hôtes génétiquement souples5,6. Enfin, C. elegans réagit rapidement aux infections bactériennes, fongiques et virales, ce qui en fait un excellent modèle pour étudier les interactions hôte-microbe et le microbiome intestinal7,8,9.
L’augmentation de l’échantillonnage de C. elegans sauvages et d’autres nématodes a permis des recherches sur l’écologie des nématodes libres et la variation génétique naturelle10,11. Parallèlement, l’échantillonnage a également augmenté la découverte d’agents pathogènes biologiques naturels et de microbes qui interagissent avec C. elegans12,13,14,15, conduisant à la mise en place de nombreux systèmes modèles hôte-microbe qui étudient les interactions avec les virus, les bactéries, les microsporidies, les oomycètes ou les champignons16,17,18,19,20 . Typiquement, C. elegans sauvage se trouve dans les tiges et les fruits en décomposition, souvent dans les climats plus tempérés, et la plupart du temps ils sont autoreproductifs21. Lorsque ces échantillons sont amenés en laboratoire, les nématodes sauvages sont isolés dans des populations clonales, porteuses d’un ensemble de microbiotes associés. Lors de la découverte de nouveaux microbes d’intérêt chez les nématodes de Caenorhabditis, les animaux sont souvent directement dépistés pour l’infection ou la colonisation par microscopie à l’aide de phénotypes facilement visualisés. Par exemple, l’infection virale peut être visualisée comme une désintégration des structures intestinales, et les stades microsporidiens peuvent être vus à l’intérieur des cellules hôtes comme des spores ou des méronts14,22. Lorsqu’un microbe d’intérêt est découvert pour une étude future, il doit être séparé des autres microbes contaminants trouvés dans les nématodes sauvages afin qu’il puisse être étudié isolément. Dans de nombreux cas, le microbe d’intérêt ne peut pas être cultivé in vitro, ce qui rend essentiel l’enrichissement du microbe dans le nématode hôte.
Par exemple, ce protocole décrit un isolat sauvage de C. tropicalis contenant une bactérie qui colonise dans la lumière intestinale des nématodes, adhérant aux cellules épithéliales intestinales de manière directionnelle. Phénotypiquement, la bactérie se développe perpendiculairement le long des côtés internes de la lumière intestinale, lui donnant un aspect hérissé, visualisé sur un microscope Normarski standard à tous les stades de l’animal, y compris le stade dauer. La plaque du milieu de croissance des nématodes (NGM) sur laquelle cette souche sauvage de C. tropicalis a été cultivée contenait une contamination visible par d’autres microbes. Ce protocole a été développé pour réduire la croissance microbienne contaminante supplémentaire sur les plaques pour l’étude de cette bactérie adhérente inconnue. Les nématodes ont été forcés au stade dauer pour protéger les bactéries dans la lumière, puis nettoyés à l’aide d’une série de lavages. Par la suite, l’espèce bactérienne inconnue a été identifiée par dissection des intestins et amplification par PCR de l’ADN ribosomique 16S pour le séquençage.
Dans l’ensemble, ce protocole peut potentiellement enrichir tout microbe d’intérêt associé à un nématode capturé à l’état sauvage. Par la suite, les chercheurs identifieront le microbe cible, visualiseront in vivo les phénotypes d’infection ou de colonisation par microscopie et étudieront les effets sur la condition physique de l’hôte ou d’autres aspects des interactions hôte-microbe. L’isolement et l’étude de nouvelles espèces microbiennes qui interagissent avec les nématodes de Caenorhabditis peuvent révéler les mécanismes génétiques de l’immunité de l’hôte et de nouveaux paradigmes d’interactions hôte-microbe pertinents pour la pathogenèse microbienne et les études sur le microbiome.
Protocole
1. Induire la formation de dauer pour les nématodes sauvages sur les plaques NGM
- Cultivez la souche sauvage de Caenorhabditis sur des plaques NGM standard avec OP50-1 E. coli comme source de nourriture après avoir obtenu les vers avec un microbe d’intérêt inculturable. Incuber les nématodes à 20 °C.
REMARQUE: La température standard pour cultiver les nématodes Caenorhabditis est comprise entre 15 et 25 ° C, mais doit être déterminée empiriquement pour éviter la perte du microbe d’intérêt. - Affamer la plaque des animaux à 20 ° C pendant 4-5 jours jusqu’à ce que tout l’OP50-1 soit consommé et que la majorité ait atteint le stade dauer.
REMARQUE: Les Dauers sont des larves à longue durée de vie qui se développent en raison de l’absence de nutrition et ont des cuticules protectrices.
2. Laver les nématodes
- Ajouter 5 mL de milieux de sels minimaux M9 (42 mM Na2HPO4, 22 mM KH2PO4, 8,6 mM NaCl, 19 mM NH4Cl) à une plaque de 6 cm de vers affamés.
- À l’aide d’une pipette et d’une ampoule en verre stérile, pipettez le M9 et les vers de la plaque et transférez-les dans un tube de centrifugeuse stérile de 15 mL.
- À l’aide d’une centrifugeuse clinique, faire tourner les vers dans le tube à 1000 x g pendant 30 s à température ambiante.
- À l’aide d’une pipette stérile de 15 mL, retirer et jeter le surnageant M9 au-dessus de la pastille de vers. Ne pas déranger les vers vivants au fond du tube en laissant environ 50 μL de M9 au-dessus des vers.
- Ajouter 10 mL de M9 + 0,05 % de Triton X-100 dans le même tube de centrifugeuse et serrer adéquatement le bouchon du tube.
- Incuber le tube sur un nutateur pendant 20 min à température ambiante pour éliminer les microbes externes. Retirez le tube du nutateur et faites tourner les vers à 1000 x g pendant 30 s.
- À l’aide d’une pipette stérile, retirez et jetez le surnageant M9 au-dessus de la pastille de vers. Ne pas déranger les vers vivants au fond du tube en laissant environ 50 μL de M9 au-dessus des vers.
- Suivez et répétez les étapes 2.5 à 2.8 trois fois de plus.
3. Désinfecter les nématodes
- Préparer une solution antibiotique et SDS dans 10 mL de tampon M9 en ajoutant 0,25 % de FDS (250 μL de FDS à 10 %), 50 μg/mL de carbenicilline, 25 μg/mL de kanamycine, 12,5 μg/mL de tétracycline, 100 μg/mL de gentamycine, 50 μg/mL de streptomycine, 37,5 μg/mL de chloramphénicol et 200 μg/mL de céfotaxime (voir tableau des matériaux).
- Incuber le tube contenant des nématodes dans la solution antibiotique et SDS sur un nutateur à température ambiante pendant la nuit.
REMARQUE: Tous les vers et embryons non dauer mourront, mais de nombreux vers dauer survivront à ce processus de nettoyage.
4. Retrait de l’antibiotique et de la solution SDS
- Faites tourner les vers dans le tube à 1000 x g pendant 1 min à température ambiante. À l’aide d’une pipette stérile, retirez le surnageant du tube de centrifugeuse sans déranger les vers au fond du tube.
- Ajouter 10 mL de M9 + 0,05 % de Triton X-100 et serrer adéquatement le capuchon du tube. Incuber le tube sur un nutateur pendant 20 min à température ambiante.
- Retirez le tube du nutator et faites tourner les vers à 1000 x g pendant 1 min. Suivez et répétez les étapes 4.2 à 4.3 trois fois.
- Après le dernier lavage, laissez la pastille de ver intacte au fond dans 100 μL de la solution et jetez le reste.
5. Propager une souche de nématode propre
- À l’aide d’une pipette en verre stérile, transférer 100 μL du surnageant et de la pastille au centre d’une plaque NGM de 10 cm ensemencée avec OP50-1. Laissez sécher la plaque sans perturber le liquide au centre.
- Laissez les dauers ramper hors du centre et à travers la pelouse OP50-1 pendant 5-10 min.
- Prenez soigneusement un seul dauer et plaquez-le sur une plaque ensemencée NGM de 6 cm. Au total, cueillez au moins 10 dauers et plaquez-les dans des plaques NGM individuelles de 6 cm ensemencées d’OP50-1 (10 plaques au total).
- Incuber les plaques pendant 4 à 5 jours à 20 ° C jusqu’à ce que les dauers aient grandi et que la génération suivante (F1) ait atteint le stade adulte. Vérifiez visuellement la contamination sur toutes les plaques, considérée comme une croissance microbienne non OP50-1.
- Pour chaque plaque propre, vérifier la propagation du microbe d’intérêt via Normarski ou la microscopie fluorescente, telle que l’hybridation fluorescente in situ (FISH)12, selon le phénotype d’intérêt.
6. Dissection intestinale et identification par PCR des espèces microbiennes
- Élevez les animaux à 20 °C pendant 3-4 jours pour leur permettre de mourir de faim et de réduire la quantité de bactéries OP50-1. Ajouter 5 mL de milieu M9 à la plaque des vers affamés.
- À l’aide d’une pipette et d’une ampoule en verre stérile, pipetez les vers M9 + de la plaque et transférez-les dans un tube de centrifugeuse stérile de 15 mL.
- À l’aide d’une centrifugeuse clinique, faire tourner les vers dans le tube à 1000 x g pendant 30 s à température ambiante.
- À l’aide d’une pipette stérile de 15 mL, retirer le surnageant du tube de centrifugeuse sans perturber les vers vivants au fond du tube.
- Ajouter 10 mL de M9 + 0,05 % de Triton X-100 et incuber le tube sur un nutateur pendant 20 min pour éliminer les microbes externes. Répétez quatre fois pour laver les vers.
- Après le dernier lavage, à l’aide d’une pointe de pipette stérile, retirer le surnageant sans perturber la pastille au fond dans 100 μL de la solution.
- À l’aide d’une pointe de pipette stérile, transférez M9 et les vers sur une plaque NGM non ensemencée et laissez la plaque sécher pendant que les vers rampent pendant 20 minutes pour aider à éliminer OP50-1 de la cuticule et de l’intestin.
- Ajouter 250 μL de M9 à la plaque séchée et utiliser une pipette en verre pour transférer les vers M9 + vers une nouvelle plaque NGM non ensemencée. Encore une fois, laissez sécher cette assiette et laissez les vers ramper pendant 20 minutes.
- Ajouter 250 μL de M9 à la plaque et transférer 100 μL des vers M9 + sur un verre de montre propre.
- À l’aide de deux aiguilles stériles de seringue de 26 G, décapitez les nématodes en tenant le ver vers le bas avec une aiguille et en utilisant l’autre aiguille pour couper le ver. Après la décapitation, l’intestin (granuleux) et la gonade (transparente) du corps sortiront naturellement du corps.
- Coupez un morceau de l’intestin exposé en maintenant l’intestin avec une aiguille et en le coupant avec l’autre.
REMARQUE: Si possible, vérifiez que le microbe d’intérêt est toujours présent dans les intestins éventrés à l’aide de la microscopie Normarski, fish ou de l’immunohistochimie23. - Transférer un seul intestin disséqué dans un tube PCR de 0,5 mL contenant 10 μL d’eau stérile. Répétez l’opération pour un total d’au moins 5 tubes PCR contenant des intestins d’animaux différents.
- Congelez les tubes PCR à -80 °C pendant au moins 5 min. Retirez les tubes PCR du congélateur et décongelez les échantillons.
- Pipettez le liquide de haut en bas plusieurs fois pour séparer les bactéries de l’intestin.
- Effectuer la PCR en utilisant des paires d’amorces universelles contre la séquence d’ADN de la petite sous-unité ribosomique de bactéries, de levures ou de microsporidies, selon le type de microbe suspecté.
REMARQUE : Par exemple, un exemple de protocole utilisant des amorces bactériennes universelles 16S est présenté dans le tableau 1. - Purifiez l’amplicon à l’aide d’un kit de nettoyage et de concentration d’ADN à base de filtre (voir Tableau des matériaux) et effectuez le séquençage de Sanger.
Résultats
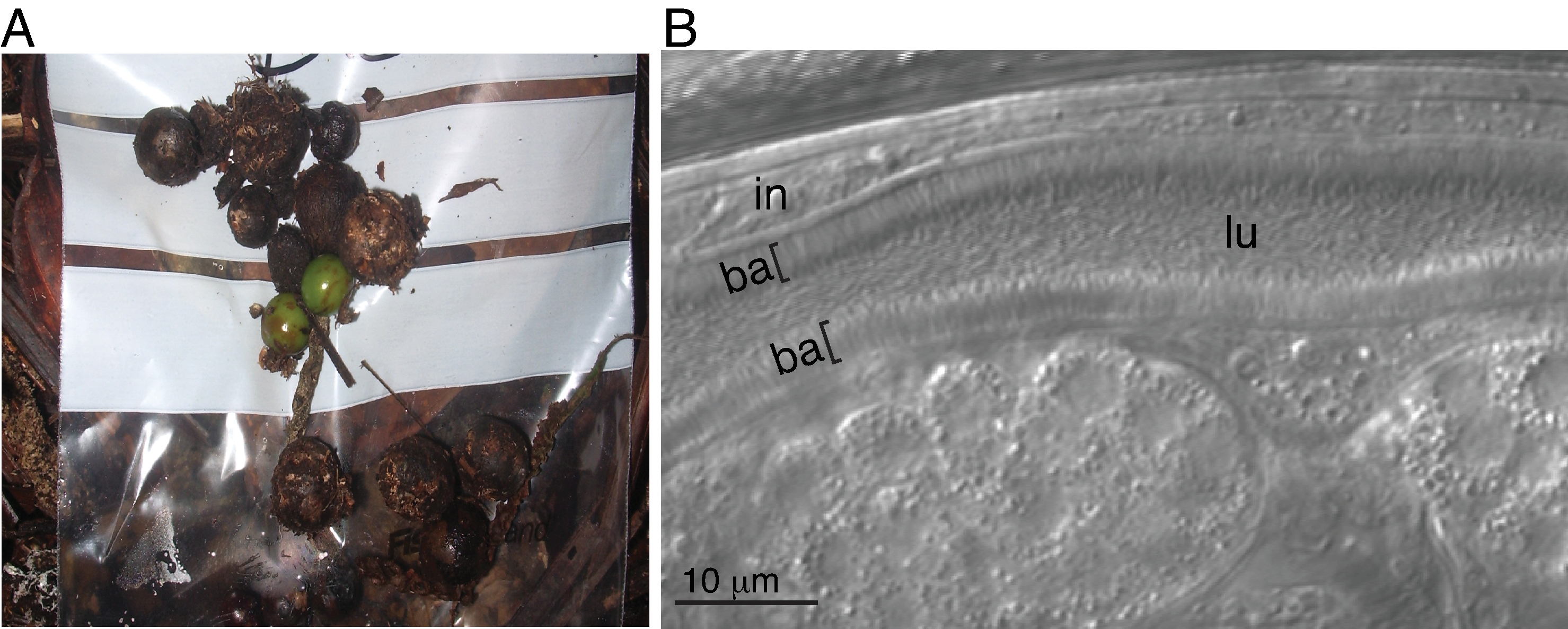
Une souche sauvage de C. tropicalis (JU1848) a été isolée à partir de fruits de palmiers pourris dans la forêt de Nouragues en Français En Guyane (Figure 1A)24. Cette souche s’est avérée avoir des microbes minces qui colonisent la lumière de l’intestin de manière directionnelle (Figure 1B). Ce microbe a été facilement transféré à la souche N2 de C. elegans par co-culture avec la souche JU1848, où il a colonisé la lumière de l’intestin de la même manière.
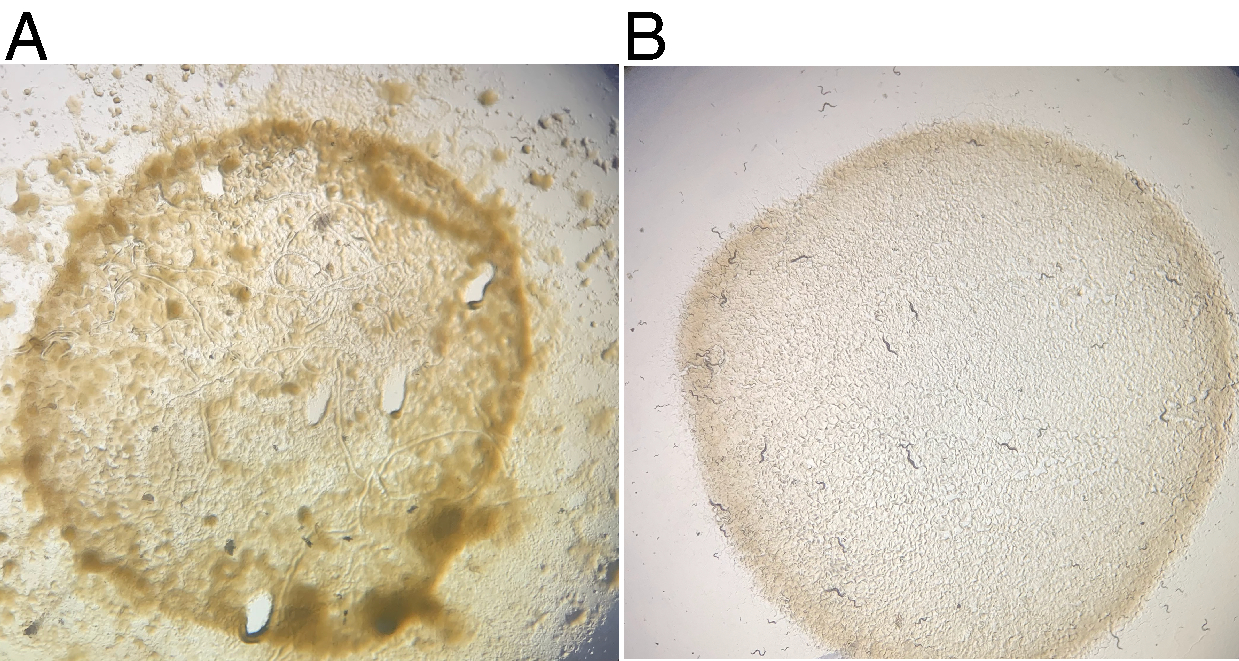
La propagation de JU1848 sur des plaques NGM standard ensemencées avec E. coli OP50-1 sur plusieurs générations a continuellement entraîné une contamination visible, considérée comme diverses colonies mucoïdes sombres sur et hors de la pelouse OP50-1 (Figure 2A). Une assiette de JU1848 sauvage a été affamée pour forcer les animaux à entrer dans la dauer et nettoyée comme décrit. Les animaux dauer individuels qui ont survécu au nettoyage ont été plaqués sur des plaques individuelles de NGM de 6 cm ensemencées avec OP50-1 et laissées pousser pendant 4 jours à 20 ° C. Plusieurs plaques de descendance F1 ont été observées sans contamination microbienne visible (figure 2B). Il a été vérifié que la descendance F1 contenait toujours des bactéries adhérentes dans la lumière de l’intestin (voir ci-dessous).
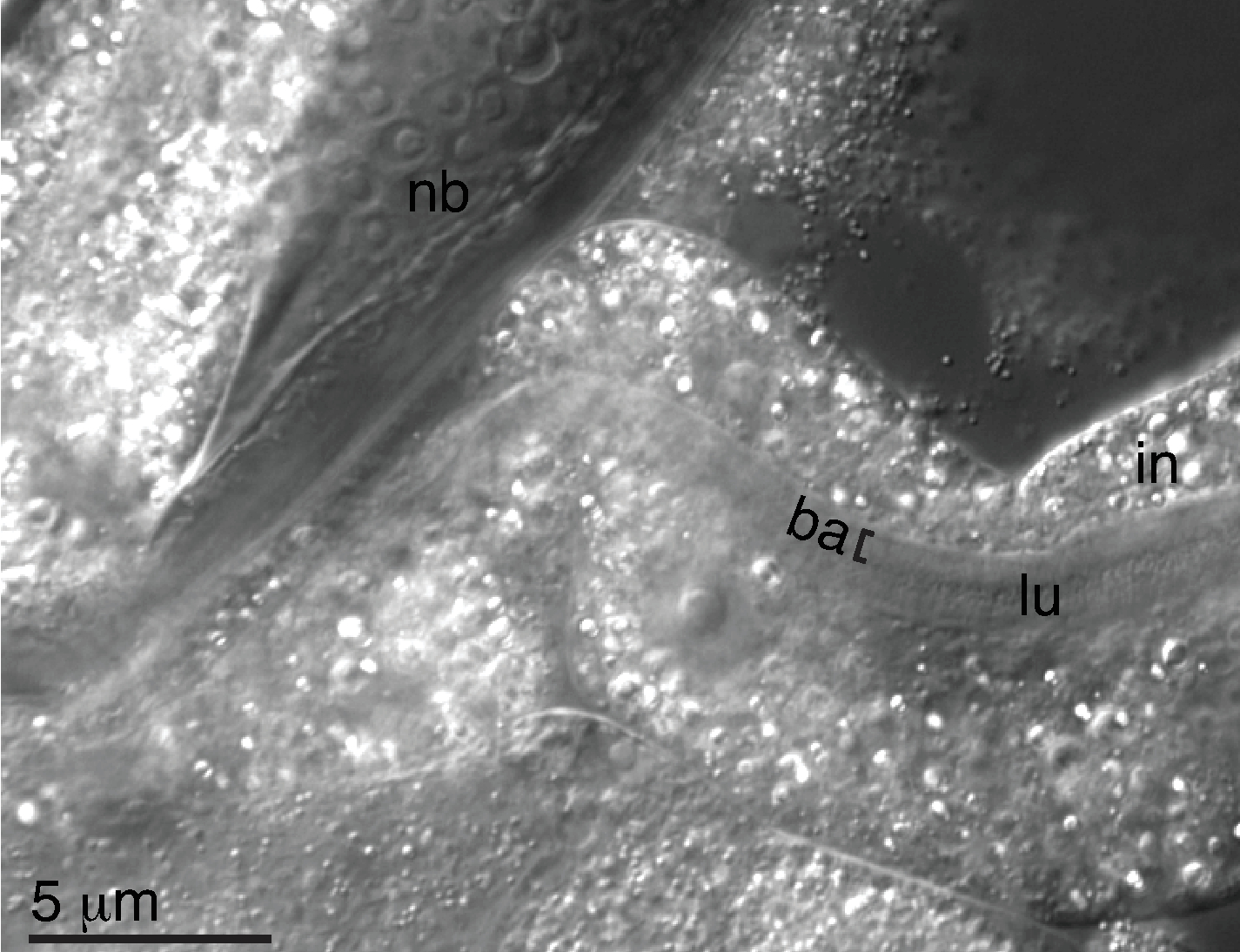
Les animaux JU1848 propres ont été lavés et décapités pour isoler les morceaux intestinaux comme décrit dans le protocole (étapes 6.1-6.12). L’adhésion des bactéries dans la lumière de l’intestin disséqué a été vérifiée par microscopie Nomarski (Figure 3). Le microbe dans la lumière de JU1848 était soupçonné d’être une bactérie, de sorte que les intestins disséqués ont été utilisés comme modèle pour la PCR en utilisant des amorces bactériennes universelles 16S, 27F et 1492R. Sur un total de six intestins disséqués individuels, les produits de PCR ont été séquencés via Sanger, et des chromatographes propres ont montré que les six séquences étaient identiques. Sur la base de ces séquences, cette bactérie a été identifiée comme une nouvelle espèce dans la classe des Alphaprotéobactéries, mais n’a pas pu être classée dans un ordre ou un genre connu (Dossier supplémentaire 1).

Figure 1 : Bactéries adhérentes colonisant la lumière d’un C. tropicalis sauvage. (A) Image d’échantillon de terrain de fruits de palmiers pourris d’Euterpe sp. (Famille : Arecaceae) dans la forêt de Nouragues de Français Guyane. (B) Image de Nomarski de la souche JU1848 vue avec des milliers de bactéries longues et minces qui forment un aspect de brosse dans la lumière (lu) de l’intestin hôte (in). Les couches bactériennes (ba) recouvrant l’intestin sont indiquées par des parenthèses ([). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : La croissance microbienne contaminante est perdue après le nettoyage des nématodes. (A) La souche sauvage JU1848 propage une croissance microbienne notable sur des plaques NGM standard de 6 cm ensemencées de bactéries E. coli OP50-1. (B) Après le nettoyage, une plaque de descendance F1 d’un seul dauer ne montre aucune contamination microbienne visible après 4 jours d’incubation à 20 °C. Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}

Figure 3 : Les bactéries adhérentes sont observées dans la lumière de l’intestin disséqué. Image de Nomarski d’un animal JU1848 propre qui a été décapité de sorte que les intestins se répandent. Les bactéries colonisatrices (ba) sont indiquées par un crochet ([) et sont vues dans la lumière (lu) d’un morceau de l’intestin (in) qui se trouve à l’extérieur du corps du nématode (nb). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
| Réactif | Concentration | Quantité |
| Amorce 27F (5'-AGAGTTTGATCMTGGCTCAG-3') | 20 mM | 2,5 μL |
| Amorce 1492R (5'-GGTTACCTTGTTACGACTT-3') | 20 mM | 2,5 μL |
| Intestin disséqué dans l’eau | N/A | 3 μL |
| 10x tampon PCR | 10x | 5 μL |
| dNTP | 10 mM | 1 μL |
| Taq Polymérase | 5 U/μL | 0,5 μL |
| Eau | N/A | 35,5 μL |
Tableau 1 : Exemple de protocole de PCR utilisant des amorces bactériennes universelles et un intestin disséqué.
Dossier supplémentaire 1: Alignement MUSCULAIRE de séquences bactériennes d’ADNr 16S dérivées de la PCR de six intestins JU1848 disséqués. Veuillez cliquer ici pour télécharger ce fichier.
Discussion
Ce protocole décrit l’isolement et l’identification des microbes des nématodes Caenorhabditis isolés à l’état sauvage à l’aide d’une série de procédures de nettoyage. De nombreux microbes sont associés à des nématodes isolés à l’état sauvage, et certains d’entre eux ont des phénotypes passionnants qui peuvent être utilisés pour de futures études sur les interactions hôte-microbe et l’immunité innée. De nombreux microbiomes cultivables et bactéries pathogènes ont été isolés à partir de nématodes sauvages de Caenorhabditis à l’aide de techniques standard de croissance bactérienne in vitro25,26. Cependant, tous les microbes ne peuvent pas être cultivés in vitro et il devient nécessaire de les enrichir chez les nématodes sauvages. Certains microbes ont un stade de spore résistant, comme les microsporidies, et des concentrations élevées de FDS peuvent être utilisées pour tuer la plupart des bactéries et des champignons, permettant un enrichissement spécifique des spores12. Ce protocole présente une méthode pour enrichir les microbes intestinaux inculturables qui ne sont pas résistants aux FDS et aux traitements antibiotiques.
La technique présentée ici tire parti de la résistance environnementale observée chez les animaux dauer en raison de changements physiologiques tels que le renforcement de la cuticule, la suppression du pompage pharyngé et la couverture de la bouche avec un bouchon buccal27. Une étape critique de ce protocole est l’incubation de nuit avec divers antibiotiques et 0,25% de FDS. Cette étape est utilisée pour tuer tous les microbes externes tout en laissant les microbes internes intacts. Alors qu’il a été démontré que C. elegans dauers survit à des concentrations de SDS aussi élevées à 10% pendant 30 min27, ce protocole utilise une incubation modérée mais prolongée non seulement pour tuer les microbes, mais aussi pour exposer davantage les bactéries aux antibiotiques. De plus, une concentration modérée de FDS peut aider à assurer la survie des dauers d’autres espèces de Caenorhabditis , car l’exposition de C. tropicalis à 1 % de FDS pendant la nuit a entraîné la mort de tous les animaux dauer. Si tous les dauers meurent, la concentration de FDS et/ou la durée d’exposition aux FDS doivent être réduites. Inversement, si les plaques de génération F1 présentent encore une contamination visible après le nettoyage, la concentration de FDS et le temps d’incubation doivent être augmentés.
Une autre étape critique est l’isolement des animaux dauer individuels après le nettoyage. Cette étape est cruciale car tous les animaux ne sont pas propres après un SDS et un traitement antibiotique. Par conséquent, les animaux sont placés au centre d’une plaque NGM de 10 cm avec OP50-1 et autorisés à ramper radialement vers l’extérieur. Souvent, il est préférable de choisir plus d’animaux distaux, car le crawl prolongé à travers OP50-1 semble aider à éliminer tous les microbes survivants potentiels attachés à la cuticule. Cependant, cela conduit à une limitation du protocole, car il sera plus difficile de l’enrichir pour un microbe d’intérêt s’il n’est pas présent dans la population à une fréquence élevée. Ici, les alphaprotéobactéries adhérentes étaient présentes dans 90% à 95% de la population; par conséquent, la plupart des plaques propres avaient la bactérie du microbiome. Cependant, si un microbe d’intérêt est présent à une fréquence beaucoup plus faible dans la population, il peut être nécessaire de filtrer beaucoup plus de plaques F1 .
Ce protocole pourrait probablement être utilisé pour isoler un certain nombre de microbes non cultivables d’intérêt trouvés dans les nématodes sauvages. Cependant, le microbe doit être dans un tissu protégé par la cuticule dauer, capable de survivre chez les animaux dauer, et avoir un phénotype observable chez l’hôte. En tant que telle, cette technique peut être utilisée pour enrichir d’autres bactéries du microbiome dans la lumière intestinale en plus des espèces d’Alphaprotéobactéries décrites ici, y compris les bactéries qui n’adhèrent pas. En outre, le protocole a été utilisé pour enrichir une bactérie intracellulaire facultative, Bordetella atropi, qui infecte le nématode Oscheius tipulae28. Après enrichissement, on a constaté que B. atropi formait des colonies sur des plaques de NGM, ce qui montre qu’un microbe d’intérêt peut être découvert cultivable in vitro une fois que les contaminants à croissance plus rapide sont éliminés. Cette technique fonctionnerait probablement pour les microsproidiens et les virus, y compris le virus Orsay, compte tenu de cette capacité à enrichir une bactérie intracellulaire. Cependant, ces microbes doivent être capables de survivre à la transition vers et hors de dauer.
Il est important de se rappeler que même si ce protocole peut être appliqué dans un laboratoire de niveau de biosécurité 1, une technique stérile doit être maintenue tout au long pour prévenir toute contamination microbienne supplémentaire. Le protocole peut être modifié en fonction des besoins du chercheur, y compris les types / concentrations d’antibiotiques, le pourcentage de FDS et / ou l’ajout d’antifongiques tels que la nystatine. Souvent, le nombre de microbes contaminants trouvés dans un nématode isolé à l’état sauvage peut varier considérablement. Ici, la perte apparente de la croissance d’E. coli non-OP50-1 sur les plaques NGM a été utilisée comme lecture pour une souche de nématode propre. Mais, il peut y avoir des populations non cultivables de microbes contaminants présents, il est donc essentiel de mener une méthode métagénomique telle que le séquençage amplicon de l’ARNr 16S pour voir l’étendue de la contamination26. Une fois la souche de ver nettoyée, elle peut être congelée et stockée pour de futures études. Dans l’ensemble, ce protocole permet aux chercheurs d’enrichir les microbes inculturables chez les nématodes sauvages, ce qui leur permet d’étudier les effets sur la condition physique de l’hôte, de caractériser les phénotypes de colonisation ou d’infection et de tirer parti des outils génétiques pour comprendre les mécanismes sous-jacents aux interactions hôte-microbe.
Déclarations de divulgation
Les auteurs ne déclarent aucun conflit d’intérêts.
Remerciements
Merci au Dr Christian Braendle et au Centre National de la Recherche Scientifique (CNRS) Nouragues Field Station.
matériels
| Name | Company | Catalog Number | Comments |
| Agarose | Fisher Scientific | BP1356 | |
| 10% SDS | Invitrogen | AM9822 | |
| BD PrecisionGlide Needle - 26 G | Fisher Scientific | 305115 | |
| Carbenicillin | Millipore-Sigma | C1389-1G | |
| Cefotaxime | Millipore-Sigma | C7039-500mg | |
| Chloramphenicol | Millipore-Sigma | C0378-25G | |
| DNA Clean and Concentrator Kit | Zymo Research | 11-303C | |
| DreamTaq Polymerase | Fisher Scientific | EP0711 | |
| Gentamycin | Millipore-Sigma | G1264-250mg | |
| Kanamycin | Millipore-Sigma | K1876-1G | |
| KH2PO4 | Fisher Scientific | P-286 | |
| NaCl | Fisher Scientific | S-671 | |
| NH4Cl | Fisher Scientific | A-661 | |
| Streptomycin | Millipore-Sigma | S6501-50G | |
| Tetracyclin | Millipore-Sigma | T7660-5G | |
| Triton X-100 | Fisher Scientific | BP-151 | |
| Watch glasses | VWR | 470144-850 |
Références
- Balla, K. M., Troemel, E. R. Caenorhabditis elegans as a model for intracellular pathogen infection. Cellular Microbiology. 15, 1313-1322 (2013).
- Pukkila-Worley, R., Ausubel, F. M. Immune defense mechanisms in the Caenorhabditis elegans intestinal epithelium. Current Opinion in Immunology. 24, 3-9 (2012).
- Bossinger, O., Fukushige, T., Claeys, M., Borgonie, G., McGhee, J. D. The apical disposition of the Caenorhabditis elegans intestinal terminal web is maintained by LET-413. Developmental Biology. 268, 448-456 (2004).
- Dimov, I., Maduro, M. F. The C. elegans intestine: organogenesis, digestion, and physiology. Cell and Tissue Research. 377, 383-396 (2019).
- Zhang, F., et al. Caenorhabditis elegans as a model for microbiome research. Frontiers in Microbiology. 8, 485 (2017).
- Szumowski, S. C., Botts, M. R., Popovich, J. J., Smelkinson, M. G., Troemel, E. R. The small GTPase RAB-11 directs polarized exocytosis of the intracellular pathogen N. parisii for fecal-oral transmission from C. elegans. Proceedings of the National Academy of Sciences of the United States of America. 111, 8215-8220 (2014).
- Bakowski, M. A., et al. Ubiquitin-mediated response to microsporidia and virus infection in C. elegans. PLoS Pathogens. 10, 1004200 (2014).
- Sowa, J. N., et al. The Caenorhabditis elegans RIG-I Homolog DRH-1 Mediates the Intracellular Pathogen Response upon Viral Infection. Journal of Virology. 94, 01173 (2020).
- Zugasti, O., et al. Activation of a G protein-coupled receptor by its endogenous ligand triggers the innate immune response of Caenorhabditis elegans. Nature Immunology. 15, 833-838 (2014).
- Félix, M. -. A., Duveau, F. Population dynamics and habitat sharing of natural populations of Caenorhabditis elegans and C. briggsae. BMC Biology. 10, 59 (2012).
- Lee, D., et al. Balancing selection maintains hyper-divergent haplotypes in Caenorhabditis elegans. Nature Ecology and Evolution. 5, 794-807 (2021).
- Luallen, R. J., et al. Discovery of a natural microsporidian pathogen with a broad tissue tropism in Caenorhabditis elegans. PLOS Pathogens. 12, 1005724 (2016).
- Osman, G. A., et al. Natural infection of C. elegans by an oomycete reveals a new pathogen-specific immune response. Current Biology. 28, 640-648 (2018).
- Felix, M. A., et al. Natural and experimental infection of Caenorhabditis nematodes by novel viruses related to nodaviruses. PLoS Biology. 9, 1000586 (2011).
- Zhang, G., et al. A large collection of novel nematode-infecting microsporidia and their diverse interactions with Caenorhabditis elegans and other related nematodes. PLoS Pathogens. 12, 1006093 (2016).
- Félix, M. -. A., Wang, D. Natural viruses of Caenorhabditis nematodes. Annual Review Genetics. 53, 313-326 (2019).
- Grover, M., Barkoulas, M. C. elegans as a new tractable host to study infections by animal pathogenic oomycetes. PLoS Pathogens. 17, 1009316 (2021).
- Bakowski, M. A., Luallen, R. J., Troemel, E. R. Microsporidia Infections in Caenorhabditis Elegans and Other Nematodes. Microsporidia: Pathogens of Opportunity: First Edition. , (2014).
- Taffoni, C., Pujol, N. Mechanisms of innate immunity in C. elegans epidermis. Tissue Barriers. 3, 1078432 (2015).
- Dirksen, P., et al. CeMbio - The Caenorhabditis elegans microbiome resource. G3: Genes, Genomes, Genetics. 10, 3025-3039 (2020).
- Frézal, L., Félix, M. -. A. C. elegans outside the Petri dish. eLife. 4, 05849 (2015).
- Troemel, E. R., Félix, M. -. A., Whiteman, N. K., Barrière, A., Ausubel, F. M. Microsporidia are natural intracellular parasites of the nematode Caenorhabditis elegans. PLoS Biology. 6, 2736-2752 (2008).
- Luallen, R. J., Bakowski, M. A., Troemel, E. R. Characterization of microsporidia-induced developmental arrest and a transmembrane leucine-rich repeat protein in Caenorhabditis elegans. PLoS One. 10, 0124065 (2015).
- Félix, M. -. A., et al. Species richness, distribution and genetic diversity of Caenorhabditis nematodes in a remote tropical rainforest. BMC Evolutionary Biology. 13, 10 (2013).
- Samuel, B. S., Rowedder, H., Braendle, C., Félix, M. -. A., Ruvkun, G. Caenorhabditis elegans responses to bacteria from its natural habitats. Proceedings of the National Academy of Sciences of the United States of America. 113, 3941-3949 (2016).
- Berg, M., et al. Assembly of the Caenorhabditis elegans gut microbiota from diverse soil microbial environments. ISME Journal. 10, 1998-2009 (2016).
- Androwski, R. J., Flatt, K. M., Schroeder, N. E. Phenotypic plasticity and remodeling in the stress-induced C. elegans dauer. Wiley Interdisciplinary Reviews: Developmental Biology. 6, 278 (2017).
- Tran, T. D., Ali, M. A., Lee, D., Félix, M. -. A., Luallen, R. J. Bacterial filamentation is an in vivo mechanism for cell-to-cell spreading. bioRxiv. , (2021).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationExplorer plus d’articles
This article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.