
Method Article
Contrôle de l’adhérence des cellules à l’aide de techniques de modelage d’hydrogel pour des applications en microscopie à force de traction
Dans cet article
Résumé
La lithographie quasi UV et la microscopie à force de traction sont combinées pour mesurer les forces cellulaires sur des hydrogels à micromotifs. La libération ciblée de cellules individuelles induite par la lumière permet un grand nombre de mesures sur le même échantillon.
Résumé
La microscopie à force de traction (TFM) est la principale méthode utilisée en mécanobiologie pour mesurer les forces cellulaires. Il est couramment utilisé pour les cellules adhérant à des substrats plats et mous qui se déforment sous la traction cellulaire (2D-TFM). TFM repose sur l’utilisation de matériaux élastiques linéaires, tels que le polydiméthylsiloxane (PDMS) ou le polyacrylamide (PA). Pour 2D-TFM sur PA, la difficulté d’atteindre un débit élevé résulte principalement de la grande variabilité des formes de cellules et des tractions, ce qui nécessite une standardisation. Nous présentons un protocole pour fabriquer rapidement et efficacement des hydrogels DE PA à micromotifs pour les études 2D-TFM. Les micromotifs sont d’abord créés par photolithographie sans masque en utilisant une lumière proche des UV où les protéines de la matrice extracellulaire ne se lient qu’aux régions micromotifs, tandis que le reste de la surface reste non adhésif pour les cellules. Le micromodèlement des protéines de la matrice extracellulaire est dû à la présence de groupes aldéhydes actifs, ce qui entraîne des régions adhésives de différentes formes pour accueillir des cellules individuelles ou des groupes de cellules. Pour les mesures TFM, nous utilisons des hydrogels PA d’élasticité différente en faisant varier les quantités d’acrylamide et de bis-acrylamide et en suivant le déplacement des billes fluorescentes incorporées pour reconstruire les champs de traction cellulaire avec la cytométrie de traction à transformée de Fourier régularisée (FTTC).
Pour obtenir un enregistrement précis des forces cellulaires, nous décrivons l’utilisation d’une dose contrôlée de lumière à motifs pour libérer des tractions cellulaires dans des régions définies pour des cellules individuelles ou des groupes de cellules. Nous appelons cette méthode la microscopie à force de traction par éclairage UV local (LUVI-TFM). Avec le traitement enzymatique, toutes les cellules sont détachées de l’échantillon simultanément, alors qu’avec LUVI-TFM, les forces de traction des cellules dans différentes régions de l’échantillon peuvent être enregistrées en séquence. Nous démontrons l’applicabilité de ce protocole (i) pour étudier les forces de traction cellulaire en fonction de l’adhérence contrôlée au substrat, et (ii) pour obtenir un plus grand nombre d’observations expérimentales à partir du même échantillon.
Introduction
Lorsqu’elles interagissent avec leur environnement extracellulaire, les cellules adhérentes exercent des forces qui sont principalement médiées par des adhérences focales à base d’intégrine reliant la matrice extracellulaire (ECM) au cytosquelette d’actine. Les adhérences focales sont des assemblages multiprotéiques centrés sur la liaison des intégrines aux protéines ECM comme la fibronectine et le collagène. Le regroupement de l’intégrine et la croissance des adhérences focales sont cruciaux non seulement pour l’établissement d’une connexion mécaniquement stable, mais aussi pour le recrutement d’autres protéines d’adhésion focale, y compris celles qui activent la voie RhoA pour la régulation de la contractilité cellulaire1. La contractilité rhoa-dépendante du cytosquelette d’actine permet aux cellules de se propager et de migrer sur l’ECM sous-jacent, ainsi que de détecter sa rigidité2. La distribution des forces de traction dépend fortement de la zone d’étalement et de la forme des cellules, qui reposent toutes deux sur les propriétés de la matrice et ont donc un impact sur l’organisation du cytosquelette, formant finalement une boucle de rétroaction fermée entre la mécanique de la matrice et l’organisation du cytosquelette3,4.
Les techniques de micromodèlement de surface permettent un contrôle défini de la forme des cellules en créant des régions de la taille d’un micron présentant des protéines adhésives ECM; selon la taille de ces régions, des cellules individuelles ou des groupes de cellules qui adhèrent au micromodèle5. Les protéines ECM peuvent être modelées sur des substrats de verre par différentes approches telles que l’impression par microcontact, le photo-motif ou le motif laser6. L’utilisation de la lumière UV (λ = 185 ou 375 nm) combinée à des stratégies antisalissures de surface offre la flexibilité nécessaire pour concevoir différentes formes et tailles et l’immobilisation de plusieurs types de protéines avec une grande précision près des bords de surface7,8. Les régions recouvertes de produits chimiques protéiques tels que le polyéthylène glycol (PEG) sont protégées par un photomasque au chrome ou un système de lithographie sans masque à base de miroir numérique (DMD). Les motifs dans les masques permettent l’exposition à la lumière UV de régions qui seront ensuite modelées avec des protéines ECM. La modelation des surfaces d’hydrogel avec des protéines ECM nécessite une étape de transfert pour éliminer les protéines de la surface du verre et les relier aux motifs. Alternativement, le motif sur des matériaux mous peut être obtenu en recouvrant d’abord le photomasque d’une protéine répulsive, puis en brûlant les régions non masquées à l’aide d’un éclairage UV profond. Étant donné que les UV profonds génèrent de l’ozone et rendent la surface réactive pour la liaison aux protéines, les régions non masquées sont recouvertes de protéines ECM et enfin le gel est polymérisé directement sur les régions non masquées9,10.
Les hydrogels à motifs peuvent être utilisés pour effectuer le TFM, une technique qui mesure les forces cellulaires à l’interface cellule-matériau11. En 2D-TFM, on utilise la surface plane d’un film polymère épais, dans lequel des billes de marqueurs ont été incorporées pour suivre les déformations12,13,14,15. Afin d’extraire des vecteurs de déplacement, il est essentiel de combiner deux images, l’une de l’état déformé et l’autre image de référence sans déformations. Les deux images sont ensuite mappées l’une sur l’autre avec traitement d’image. À haute densité de marqueurs, cela se fait généralement avec la vélocimétrie par image de particules (PIV), qui est une méthode bien établie pour reconstruire l’écoulement hydrodynamique. À faible densité de marqueurs et en 3D-TFM, cela se fait généralement avec la vélocimétrie de suivi des particules (PTV), qui comprend des caractéristiques spécifiques de l’ensemble de données expérimentales. Un exemple d’alternative moins coûteuse en calcul est le flux optique, tel que l’algorithme Kanade-Lucas-Tomasi (KLT)16. Dans le cas des substrats d’hydrogel, les billes fluorescentes sont généralement incorporées à haute densité pendant la polymérisation du matériau, et les images sont enregistrées avant et après la libération des cellules lors du détachement enzymatique. Le détachement enzymatique des cellules adhérentes, par exemple par trypsinisation, conduit à la libération simultanée de tractions cellulaires de toutes les cellules sur les hydrogels, ce qui rend difficile l’obtention d’une analyse détaillée à partir d’un grand nombre de cellules.
Nous présentons ici un protocole pour préparer des hydrogels à micromotifs pour contrôler la forme et la localisation des cellules et une méthode pour mesurer efficacement les forces de traction en séquence en utilisant les UV pour détacher les cellules individuelles du substrat. Pour les mesures de force de traction, nous présentons une technique pour produire des hydrogels PA à double couche, où les billes fluorescentes sont incorporées uniquement dans la couche supérieure, ce qui augmente leur densité et réduit leur étalement vertical. La combinaison de la libération par les UV des forces de traction cellulaires avec le micromotif permet d’obtenir un contrôle spatial sur le détachement cellulaire (par exemple, des cellules individuelles sans affecter l’adhésion d’autres cellules dans la région d’intérêt), à condition qu’il y ait une distance suffisante entre les cellules à motifs. La traction cellulaire est ensuite reconstruite à l’aide de la méthode la plus efficace et la plus fiable pour la 2D-TFM, à savoir la cytométrie de traction à transformée de Fourier (FTTC) avec régularisation17,18.
Protocole
1. Préparation des couvercles méthacrylés
REMARQUE: Les couvercles en verre sont méthacrylés pour lier de manière covalente les hydrogels PA et donc empêcher leur détachement pendant l’incubation avec les cellules. Les couvercles en verre de microscope sont utilisés pour obtenir une qualité d’image élevée.
- Pour préparer une solution de 300 mL, prenez un bécher en verre propre de 400 mL et placez-le à l’intérieur d’une hotte aspirante.
- Ajouter 10 mL d’eau distillée double (ddH2O) dans le bécher. Ajouter 18,75 mL d’acide acétique (≥99,8 % pro analyse, p.a.), 18,75 mL de méthacrylate de 3-(Triméthoxysilyl)propyle et 252,5 mL d’éthanol (≥99,8 % par an) à l’aide d’une pipette sérologique. Versez la solution dans un plat cristallisant.
- Nettoyez les couvercles en verre rond de 24 mm avec des lingettes de précision. Placez les couvercles dans un rack en polytétrafluoréthylène sur mesure.
- Immerger le rack dans la solution et incuber pendant 15 min à température ambiante (RT).
- Prenez la grille et rincez tous les couvercles avec de l’éthanol (99,8% par an). Sécher les couvercles sous le flux d’air.
REMARQUE: Les couvercles méthacrylés peuvent être conservés jusqu’à 1 mois à RT.
2. Micromodélisation des protéines de la matrice extracellulaire sur des couvercles en verre
REMARQUE: Les couvercles en verre sont d’abord recouverts d’une couche de molécules, comprenant des protéines et un répulsif cellulaire. Cette couche est ensuite enlevée à l’aide d’une technique de photomodélisme, pour permettre le dépôt ultérieur de protéines de matrice extracellulaire dans des régions micromotifs.
- Prenez un couvercle en verre rond de 15 mm et placez-le sur une boîte de Pétri. Utilisez un stylo diamanté pour marquer la face supérieure du couvercle. Ensuite, passez au nettoyage par traitement au plasma à l’oxygène à 0,4 mbar et 200 W pendant 2 min.
- Pipette 100 μL de solution de poly-L-lysine à 0,01% sur la surface de chaque couvercle. Incuber pendant 30 min à TA. Laver les couvercles avec 10 mM d’acide 4-(2-hydroxyéthyl)-1-pipérazineéthanesulfonique (HEPES) pH 8,5. Enlevez tout excès de liquide, mais gardez la surface humide.
- Pipette 100 μL de 50 mg/mL d’acide poly(éthylène glycol) méthyl éther succinimidyl valérique (mPEG-SVA) dans 10 mM HEPES pH 8,5 sur la surface de chaque couvercle. Incuber pendant 1 h à RT. Rincer les couvercles avec 10 mM HEPES pH 8,5 et sécher sous le flux d’air.
- Ajouter 2 μL de photoinitiateur sensible aux UV (p. ex. gel PLPP) suivi de 40 μL d’éthanol (≥99,8 % par an) sur la surface. Pour obtenir une distribution homogène du gel et de l’éthanol à la surface, inclinez doucement la boîte de Petri d’avant en arrière. Attendez 5 min pour que la solution polymérise.
- Placez un seul couvercle à l’intérieur d’une boîte de Petri de 35 mm avec un trou de 20 mm au fond. Cela permet au faisceau laser venant de dessous d’atteindre directement la surface du verre.
- Allumez le microscope et un module de modélisation de la lumière (travailler avec cet appareil n’est autorisé qu’après une introduction de sécurité par les agents de sécurité laser). Calibrez le laser UV-A sur l’objectif d’air 20x. Mettez l’échantillon avec le photoinitiateur sur la scène. Ajustez la mise au point sur la surface du verre et activez le contrôle de mise au point. Chargez et verrouillez le motif pré-dessiné.
- Préparez le motif à l’aide d’un logiciel graphique comme Inkscape. Assurez-vous que le fichier de motif ne dépasse pas les dimensions DMD (1824 x 1140 px, ce qui correspond à 552 x 325 μm sur objectif 20 x (0,28 μm/px)).
REMARQUE: Il est recommandé d’utiliser la taille DMD (1824 x 1140 pixels) comme taille du fichier de motif, car cela garantira l’absence d’erreur lors de la modélisation. Par exemple, ne produisez pas de fichier de motif avec une dimension de 1830 x 1130 pixels. - Aux fins du transfert de motif au polyacrylamide, assurez-vous que le motif n’est effectué que jusqu’à 60% à 70% du rayon du centre du verre au bord.
- Pour modéliser une seule cellule, utilisez un diamètre de 50 à 100 μm avec une distance de 100 μm entre les motifs et un diamètre de 150 à 300 μm pour un groupe de cellules. La taille de motif appropriée dépend fortement de la taille typique de la cellule, et elle doit être suffisamment grande pour permettre aux cellules d’adhérer.
- Préparez le motif à l’aide d’un logiciel graphique comme Inkscape. Assurez-vous que le fichier de motif ne dépasse pas les dimensions DMD (1824 x 1140 px, ce qui correspond à 552 x 325 μm sur objectif 20 x (0,28 μm/px)).
- Commencez le modelage par une dose UV de 30 mJ/mm2. La durée de la modélisation dépend du nombre de motifs. Faire un seul motif prend environ 1 s.
- Après avoir terminé l’étape de modelage, rincez la surface avec une solution saline tamponnée au phosphate (PBS) trois fois. Incuber l’échantillon avec 100 μL de 25 μg/mL de fibronectine dissoute dans 1x PBS et 25 μg/mL de conjugué de fibrinogène Alexa488 dans pbS pendant 1 h à RT. Pour les autres protéines ECM, trouvez les concentrations optimales couvrant complètement les régions à motifs.
- Rincez la surface avec 1x PBS trois fois. Assurez-vous que le verre à motifs est immédiatement utilisé pour le transfert verre-PA. Conservez le verre à motifs dans PBS at RT pendant la préparation de l’hydrogel.
3. Fabrication d’hydrogels de polyacrylamide à motifs
REMARQUE: Des hydrogels de polyacrylamide sont préparés, y compris l’acrylamide N-hydroxyéthyle oxydé (HEA) pour présenter des groupes aldéhydes réactifs pour la liaison covalente des protéines de la matrice à la surface. De plus, une approche à double couche pour intégrer des billes fluorescentes uniquement sur la couche supérieure de l’hydrogel est utilisée pour améliorer l’enregistrement du déplacement des billes lors d’expériences de microscopie à force de traction.
- Mettez les couvercles en verre méthacrylé dans une boîte de Pétri.
- Pour préparer une solution fraîche oxydée de HEA, ajouter 9,55 mL d’eau double distillée dans un tube de centrifugeuse de 15 mL. Ensuite, ajoutez 0,5 mL de HEA et 42 mg de (méta)periodate de sodium pour obtenir 10 mL de la solution. Remuer continuellement la solution dans l’obscurité pendant 4 h.
- Mélanger l’acrylamide, le bis-acrylamide et l’eau double distillée selon le tableau 1, pour obtenir 10 mL d’hydrogel en solution mère (faites-le sous la hotte, les monomères sont neurotoxiques). Dégazez et fermez le couvercle à un joint étanche à l’air.
REMARQUE: La solution mère peut être conservée dans des aliquotes jusqu’à 1 an à 4 ° C. Caractérisez toujours le module de Young de l’hydrogel avant utilisation. - Préparez un hydrogel PA à double couche (faites-le sous la hotte, les monomères sont neurotoxiques).
- Commencez par la couche inférieure en mélangeant doucement 99,3 μL de solution mère, 0,5 μL de persulfate d’ammonium (APS) à 1 % et 0,2 μL de tétraméthyléthylènediamine (TEMED) dans un tube de 1,5 mL. Prélever 10 μL de la solution et la pipeter goutte à goutte sur le centre du couvercle méthacrylé.
- Placez soigneusement un couvercle rond de 15 mm sur la gouttelette et attendez 45 minutes pour la polymérisation. Détachez délicatement le couvercle supérieur avec un scalpel.
- Pour la couche supérieure, mélanger délicatement 93,3 μL de solution mère avec 1 μL de solution HEA fraîche oxydée, 5 μL de billes fluorescentes (correspondant à trois billes par micron carré pour les billes de 200 nm), 0,5 μL de APS à 1 % et 0,2 μL de TEMED dans un tube de 1,5 mL. Prélever 5 μL de la solution et la pipeter goutte à goutte sur le centre de la couche inférieure déjà polymérisée.
- Placez doucement le couvercle à micromotifs sur la gouttelette. Attendez 45 min à TA pour que la gouttelette se polymérise. Assurez-vous que le côté à motifs touche la gouttelette. Détachez délicatement le couvercle avec un scalpel.
- Collez les couvercles de 24 mm au fond de plaques de 6 puits percées sur mesure à l’aide de colle silicone à deux composants. Après 5 min, ajoutez PBS aux puits.
- Caractériser le module de Young des échantillons d’hydrogel de polyacrylamide par microscopie à force atomique (AFM).
- Montez une pointe sphérique en porte-à-faux sur le support AFM.
- Calibrez chaque porte-à-faux en acquérant une courbe force-distance (point de consigne de 3 à 4 V) sur un substrat dur (p. ex. verre ou mica), extrapolez la sensibilité en porte-à-faux, rétractez-vous de la surface et effectuez un réglage thermique pour obtenir leur ressort constant.
- Placez les porte-à-faux étalonnés sur l’échantillon d’hydrogel et acquérez des courbes de force dans le tampon PBS, en utilisant les paramètres suivants : point de consigne de force de 20 à 30 nN, vitesse d’approche et de rétraction de 5 ou 10 μm/s, taille de rampe de 5 μm.
REMARQUE: Un microscope optique inversé, équipé d’un objectif d’air 40x et d’un filtre à protéine fluorescente verte (GFP) a été utilisé pour visualiser et cibler les zones à micromodenture ECM dans les échantillons d’hydrogel PA. - Tracez les courbes de force par rapport à distance acquises avec le logiciel d’analyse AFM.
- Trouvez le point de contact et convertissez les courbes de force par rapport à la distance en courbes de force par rapport aux courbes d’indentation.
- Ajuster la partie indentation de la courbe avec un modèle hertz (ou un modèle mécanique approprié en fonction de la géométrie de la pointe), en utilisant un rapport de Poisson compris entre 0,2 et 0,5.
4. Libération locale des forces de traction cellulaire par éclairage laser UV-A (LUVI-TFM)
REMARQUE: Le laser UV-A est appliqué pour libérer des forces de traction dans des cellules localisées dans des régions définies des hydrogels. Le laser UV-A (c’est-à-dire λ = laser à semi-conducteurs de 375 nm, <15 mW) est un laser de classe 3B. Le faisceau laser non blindé est dangereux pour les yeux et souvent pour la peau. La lumière et le rayonnement réfléchis et diffusés peuvent être dangereux. De plus, le rayonnement UV peut causer le cancer de la peau. Le travail avec des appareils n’est autorisé qu’après l’introduction de la sécurité par les agents de sécurité laser.
- Aspirez le PBS des puits.
- Ensemencez 3 x 106 cellules par plaque de 6 puits en milieu de croissance. Pour les fibroblastes, utilisez le milieu Eagle modifié (DMEM) de Dulbecco contenant de la L-glutamine et complété par 10% de sérum fœtal bovin (FBS) et 1% de pénicilline / streptomycine. Incuber les cellules pendant la nuit à 37 °C/5 % de CO2. Lavez les échantillons pour enlever les cellules flottantes avant de commencer la microscopie.
- Allumez la chambre d’incubation du microscope et l’alimentation en gaz pour obtenir une température de 37 °C et une atmosphère de CO2 de 5 %.
- Allumez le microscope et le module de modelage laser. Si nécessaire, recalibrez le laser UV-A sur l’objectif d’air 20x à l’aide du logiciel Leonardo. Placez la plaque de 6 puits sur mesure avec les échantillons sur la scène. Réglez la mise au point sur la surface du PA.
- Configurez le motif d’éclairage et marquez les cellules d’intérêt. Recentrez-vous sur la surface de l’hydrogel. Acquérir l’image des cellules dans le canal de champ lumineux. Passez au canal Cy5 (filtre rouge lointain, excitation de 650 nm, émission de 670 nm) et prenez une image des billes à l’état déformé.
- Passez au canal laser. Allumez le laser pendant 3 min. Dans notre configuration particulière, cela correspondait à une dose légère de 6 000 mJ/mm2.
- Passez au canal Cy5 et obtenez une image des perles à l’état non déformé.
REMARQUE: Pour l’expérience utilisant des fibroblastes embryonnaires de souris, attendez 15 minutes après l’exposition pour enregistrer l’image de référence / non déformée (voir la section Résultats représentatifs ). Cela peut être différent pour d’autres types de cellules. Répétez les étapes 4.5 à 4.7 pour une ou plusieurs autres cellules d’intérêt. - Pour mesurer le stress oxydatif élevé après l’éclairage UV-A, utilisez le réactif disponible dans le commerce (CellRox). CellRox est non fluorescent à l’état réduit et, lorsqu’il est oxydé par des espèces réactives de l’oxygène, fluoresces avec un maximum d’émission d’environ 665 nm.
- Selon le protocole fourni, ajouter un réactif de 5 μM dans le support d’imagerie et incuber pendant 30 min à 37 °C/5 % de CO2. Ensuite, lavez les cellules 3x avec 1x PBS chaud et remplacez le support d’imagerie. Enregistrez le signal Cy5 par imagerie de fluorescence avant et après l’éclairage UV-A en utilisant une exposition à la lumière similaire.
5. Traitement d’images, vélocimétrie d’images de particules et calcul des forces de traction
REMARQUE: Les forces de traction des cellules sont enregistrées en analysant le déplacement des billes fluorescentes et calculées à l’aide d’outils d’analyse d’images basés sur la vélocimétrie d’images de particules.
- Ouvrir des images de perles fluorescentes, avant (c’est-à-dire déformées) et après le laser (c’est-à-dire non déformées) à l’aide de Fidji. Fusionner deux images en une seule pile (Image | Piles | Images à empiler).
- Corriger la dérive latérale entre les deux images à l’aide du plugin StackReg (P. Thévenaz, Ecole polytechnique fédérale de Lausanne). Changer les images en 8 bits (Image | Type | 8 bits) et redimensionner à 1024 x 1024 pix (Image | Échelle). Enregistrez en tant que séquence d’images (format TIFF) avec l’image de référence toujours au début.
- Acquérir le champ vectoriel de déplacement à l’aide d’une technique de corrélation croisée19 à partir du champ PIV telle qu’implémentée dans le projet OpenPIV.
- Assurez-vous que l’image détendue (non déformée) et l’image déformée sont couvertes par des fenêtres de recherche de taille wR et wD en pixels. Pour chaque fenêtre de recherche, définissez une fonction de corrélation croisée à l’aide de la formule suivante :

Ici, R(i,j) et D(i,j) décrivent les champs d’intensité des deux images tronqués à la fenêtre de recherche sélectionnée et ensuite rembourrés en miroir. et
et  décrire leur valeur moyenne. Les tailles de fenêtre sont choisies pour être wR = 32 et wD = 32 et un chevauchement de 70% est utilisé entre les fenêtres de recherche voisines.
décrire leur valeur moyenne. Les tailles de fenêtre sont choisies pour être wR = 32 et wD = 32 et un chevauchement de 70% est utilisé entre les fenêtres de recherche voisines. - Pour chaque fenêtre de recherche, déterminez un vecteur
 de déplacement qui optimise la fonction de corrélation croisée et affectez-la à la position centrale de la fenêtre de recherche. La précision des sous-pixels est obtenue à l’aide d’un ajustement gaussien autour de la région des maxima comme décrit20.
de déplacement qui optimise la fonction de corrélation croisée et affectez-la à la position centrale de la fenêtre de recherche. La précision des sous-pixels est obtenue à l’aide d’un ajustement gaussien autour de la région des maxima comme décrit20. - Recherchez et supprimez les vecteurs de déplacement ambigus. Les vecteurs de déplacement correspondant aux fenêtres de recherche où le rapport entre deux maxima locaux les plus élevés de la fonction de corrélation croisée en dessous d’un seuil de 1,5 sont considérés comme ambigus21.
- Écartez les vecteurs de déplacement qui échouent au test médian normalisé pour un seuil résiduel de 1,522.
- (Facultatif) Corrigez la dérive latérale restante en soustrayant un vecteur
 fixe de tous les déplacements. Ceci est fait parce que le déplacement dans une zone sélectionnée loin de la cellule disparaît.
fixe de tous les déplacements. Ceci est fait parce que le déplacement dans une zone sélectionnée loin de la cellule disparaît. - Interpolez le champ vectoriel de déplacement résultant sur une grille régulière avec un espacement de 4 px des images d’entrée à l’aide d’une interpolation polynomiale cubique. Les points de données manquants sont remplis à l’aide d’une extrapolation de spline bivariée lisse. Un vecteur de déplacement en pixels est lié à une déformation locale par le rapport de pixels de l’image (0,33 μm/px pour une lentille d’air 20x).
- (Facultatif) Tamponner l’image pour réduire les artefacts de sonnerie dans l’analyse suivante.
- Multipliez le champ de déformation par une fonction de fenêtre Tukey pour éliminer l’effet de bord dû à la correction de dérive, avec un paramètre de 0,2-0,3. Dans cette expérience, la zone à micromotifs qui se trouve au centre du champ de vision est intéressante.
- Assurez-vous que l’image détendue (non déformée) et l’image déformée sont couvertes par des fenêtres de recherche de taille wR et wD en pixels. Pour chaque fenêtre de recherche, définissez une fonction de corrélation croisée à l’aide de la formule suivante :
- Reconstruisez la traction à l’aide de la cytométrie de traction à transformée de Fourier (FTTC)18 régularisée. Comme régularisation, nous utilisons le choix raisonnable le plus simple, à savoir, la régularisation Tikhonov (L2) de 0ème ordre23,24,25. Un paramètre de régularisation optimal λ est choisi de telle sorte qu’il minimise la fonction de validation croisée généralisée (GCV) définie par26 :

Ici, τλ est un vecteur empilé contenant les composantes x et y du champ de traction reconstruit pour la valeur donnée de λ pour tous les modes d’échantillonnage de Fourier et G est l’opérateur linéaire mappant les champs de traction à leur champ de déplacement correspondant, tel que défini pour FTTC. La somme s’exécute sur des composants vectoriels. ui sont les composantes x et y du champ de déplacement pour tous les modes d’échantillonnage de Fourier. La GCV est largement utilisée dans le domaine des problèmes inverses mal posés et constitue une bonne alternative au critère de la courbe en L souvent utilisé dans la TFM. Un filtre Lanzcos est utilisé pour réduire les artefacts introduits en raison de l’espace de fréquence limité et du suréchantillonnage du champ de déplacement. Le calcul de traction dépend du module de Young du PA (par exemple, 11,3 kPa) et du rapport de Poisson (par exemple, pour le PA compris entre 0,2 et 0,5 selon la concentration du réticulant).
Résultats
Les hydrogels de PA ont été polymérisés sur des couvercles en verre méthacrylé, qui présentent des groupes réactifs pour la liaison covalente du PA. De cette façon, lors de la mise en place des hydrogels dans une solution aqueuse, leur détachement du substrat a été empêché (Figure 1A-D). Pour obtenir une forte densité de marqueurs fiducial près de la surface de l’hydrogel, nous avons développé la préparation d’un hydrogel PA à double couche. La couche inférieure, sans perles fluorescentes, a été polymérisée sur le couvercle méthacrylé. Par la suite, une autre couche contenant des billes a été polymérisée sur le dessus de la couche inférieure (Figure 1E-H), remplaçant le besoin de sédimentation, qui est souvent utilisé pour obtenir la distribution des billes dans un seul plan focal et augmenter la densité des billes. Pour contrôler la forme des cellules lors des mesures TFM, nous avons produit des hydrogels PA à micromolymérisation par transfert direct de microstructures à motif de fibronectine d’un couvercle en verre vers le PA prépolymérisé (Figure 1F-F'). L’ajout de 1% de réticulant non conventionnel (HEA oxydé) dans la solution de couche supérieure d’hydrogel fournit des groupes aldéhydes qui se lient de manière covalente aux groupes amines de la fibronectine.
Nous avons préparé des hydrogels PA, d’une épaisseur d’environ 60 μm, et des billes fluorescentes confinées dans la couche supérieure près de la surface de l’hydrogel. Ce type d’hydrogel est un substrat approprié pour l’imagerie des tractions cellulaires avec des microscopes inversés et suffisamment épais (l’épaisseur minimale pour effectuer le TFM a été rapportée à 20-30 μm)18 pour éviter tout impact du substrat de verre. La localisation des billes fluorescentes a été imagée par microscopie confocale (Figure 1I). Pour visualiser le transfert de protéines sur l’hydrogel, nous avons utilisé des protéines ECM marquées par fluorescence et les avons imagées par microscopie à épifluorescence (Figure 1J). Nous avons préparé des cercles de fibronectine à micromotifs d’un diamètre de 100 μm (côté gauche) et de 50 μm (côté droit) pour permettre l’adhésion de groupes de cellules ou de cellules individuelles, comme le montre la superposition d’images en champ lumineux de cellules adhérentes, et de perles fiduciales marquées par fluorescence et de micromotifs de fibronectine. Sur un autre échantillon, la réponse d’adhésion de cellules individuelles à la fibronectine à micromotifs a été validée par microscopie à immunofluorescence indirecte pour imager la localisation de protéines d’adhésion focale telles que la paxilline (Figure 1K).
Le revêtement protéique ECM et la rigidité de l’hydrogel sont des paramètres cruciaux pour l’adhésion cellulaire26. Pour caractériser les propriétés mécaniques de nos hydrogels PA, nous avons réalisé des expériences de nanoindentation par AFM27, couplées à un microscope à épifluorescence. Des substrats d’hydrogel de trois rigidités différentes ont été préparés et testés avec notre configuration (Figure 2 et Tableau 1). Les porte-à-faux AFM en forme de sphère ont été approchés cycliquement et rétractés à partir d’hydrogels PA non modélisés et de fibrinogène conjugués à des zones micromotifs Alexa488, tout en enregistrant des courbes force-distance. Par la suite, l’évaluation du point de contact et l’application du modèle Hertz nous ont permis d’estimer le module de Young (E) de chaque échantillon. Le micromodèle ECM n’a pas modifié le module de Young, qui est resté comparable à chacun des hydrogels PA non modélisés.
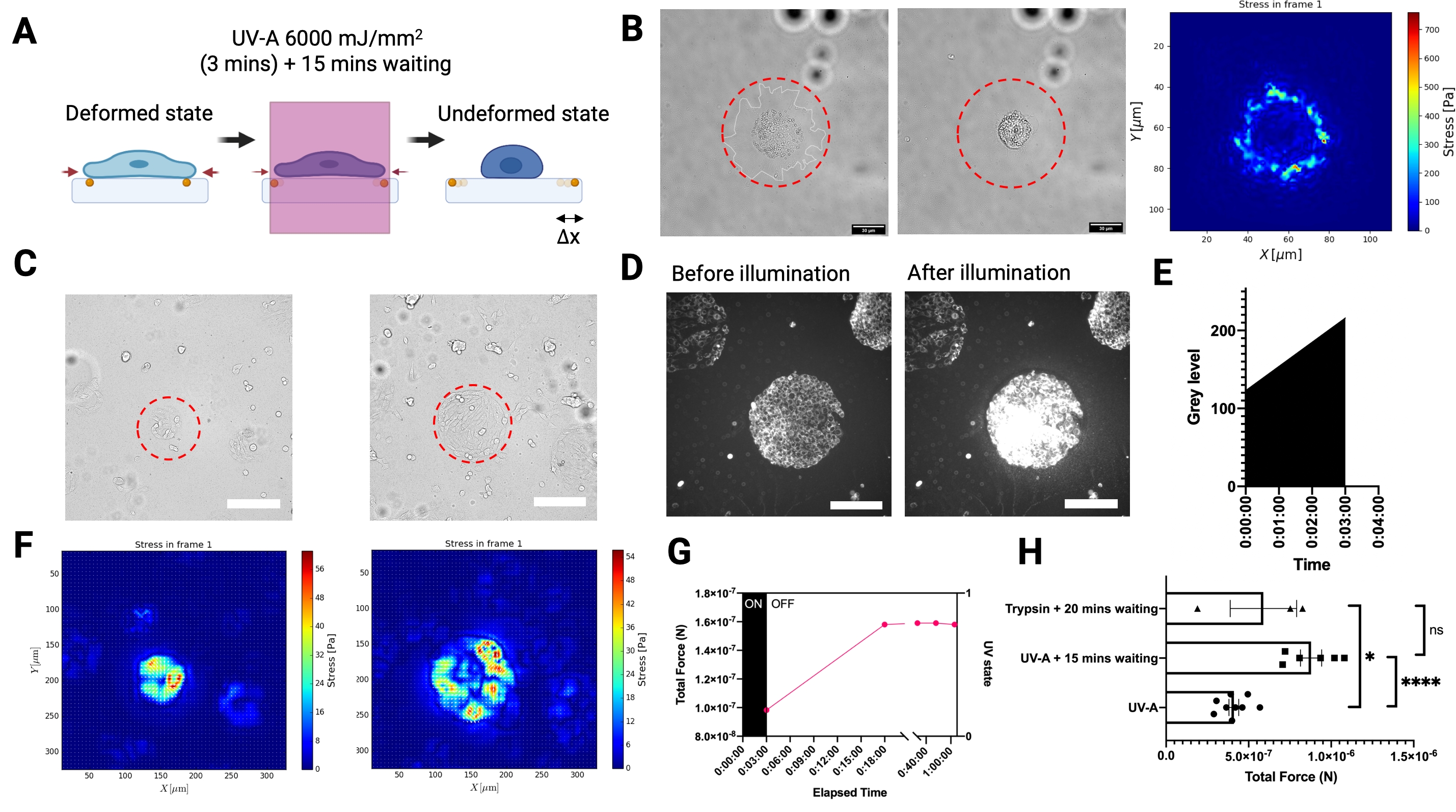
Pour mesurer les forces de traction exercées par les cellules adhérentes sur le substrat, nous avons développé une configuration expérimentale de TFM de référence réalisée sur un microscope à épifluorescence à large champ équipé d’un module laser proche UV (UV-A 6000 mJ/mm2) pour libérer les tractions cellulaires (Figure 3A). Comme l’éclairage du faisceau laser peut être contrôlé spatio-temporellement, non seulement il est possible d’exposer sélectivement une seule cellule ou un groupe de cellules avec une dose laser élevée, mais aussi et surtout, l’étape intermédiaire perturbatrice de l’élimination cellulaire de l’échantillon entier à l’aide d’une enzyme digestive comme la trypsine n’est plus nécessaire. L’éclairage des cellules avec une dose laser aussi élevée a induit un stress oxydatif élevé conduisant à la mort cellulaire (Figure 3B). La mort cellulaire a entraîné la libération de tractions du substrat, comme l’indique le déplacement des billes (illustré à la figure 3A). Nous avons combiné l’éclairage UV-A avec un module de motif lumineux pour éclairer sélectivement des régions de la taille d’un micron des hydrogels PA (Figure 3B-C). De cette façon, il est possible de libérer les tractions d’amas de cellules à micromotifs (Figure 3C,F) ou de cellules individuelles (Figure 3B). Il est important de noter qu’à nos échelles de temps, les propriétés mécaniques des hydrogels à motif ECM n’étaient pas significativement affectées par l’exposition aux UV (Figure 2C). L’augmentation du stress oxydatif est illustrée à la figure 3D,E. Les forces de traction ont été reconstruites à l’aide de FTTC régularisés avec un paramètre de régularisation choisi par la validation croisée généralisée (Figure 3B,F). La libération de forces pour les cellules individuelles s’est produite sur une période de 15 minutes, et le résultat du LUVI-TFM est comparable au TFM à base de trypsine (Figure 3G-H).

Figure 1: Schéma de préparation du substrat de fibronectine à motifs pour TFM. (A) La solution pour la couche inférieure est pipetée sur une surface méthacrylée. (B) Un couvercle propre plus petit est soigneusement placé sur la gouttelette. (C) Le temps de gélification est de 45 min. (D) Le couvercle supérieur est détaché. La couche inférieure est prête. (E) La solution pour la couche supérieure est pipetée sur l’hydrogel. (F) Le couvercle à motifs est soigneusement placé sur la gouttelette. (F') Le micromodèlement des protéines adhésives sur le verre est produit par lithographie sans masque près des UV, puis transféré du verre à l’hydrogel PA. Les groupes amines libres des protéines adhésives, par exemple la fibronectine, se lient de manière covalente aux groupes aldéhydes à la surface de l’AP. (G) Le temps de gélification est de 45 min. (H) Le couvercle supérieur est détaché. L’hydrogel PA à motifs est prêt. (I) Représentation 3D d’une micrographie confocale xyz montrant une forte densité de perles fiduciales près de la surface. (J) Une micrographie de fibronectine marquée par fluorescence (magenta) à la surface de PA avec des perles fluorescentes incorporées (vert), recouvertes d’une image en champ lumineux des cellules. (K) Imagerie par microscopie par immunofluorescence indirecte d’une seule cellule adhérant à un micromotif de fibronectine en forme de cercle (100 μm). Noyau (bleu), paxilline (rouge). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : Caractérisation des propriétés mécaniques de l’échantillon par AFM. (A) Configuration expérimentale de nanoindentation AFM. Un porte-à-faux en forme de sphère est utilisé pour sonder les micromotifs ECM avant ou après le traitement UV, et les régions non modelées des zones PA à double couche (5% Acrylamide, 0,3% Bis-acrylamide). (B) Force représentative par rapport à la courbe d’indentation pour le micromoptère ECM (noir). L’ajustement Hertz (rouge) est utilisé pour calculer le module de Young (E) de l’échantillon. (C) Mesures mécaniques pour les zones PA à double couche non modelées (pas d’ECM), les micromotifs ECM et les micromotifs ECM après exposition aux UV-A. Les barres montrent des ± moyennes S.E.M. (test ANOVA de Brown-Forsythe et Welch). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Graphique 3. L’éclairage local UV-A TFM (LUVI-TFM) permet de mesurer la force de traction locale dans un large champ de vision. (A) Schéma d’éclairage UV-A local TFM. Les cellules sont traitées avec une forte dose de laser UV-A pour obtenir l’image non déformée (référence). (B) L’image en champ lumineux d’une seule cellule avant l’éclairage laser UV-A. Milieu : L’image en champ lumineux d’une seule cellule après l’éclairage laser UV-A. Droite : Force de traction d’un seul MEF adhérant à un hydrogel PA revêtu de fibronectine (E = 5,74 kPa) éclairé par un faisceau de lumière UV de 50 μm de diamètre (ligne pointillée rouge). (C) À gauche : Enregistrement des forces de traction d’un groupe de fibroblastes embryonnaires de souris (MEF) adhérant à la fibronectine à micromotif (cercle, diamètre de 100 μm). L’amas a été éclairé par un faisceau de lumière UV de 200 μm de diamètre (ligne pointillée rouge). Droite : Enregistrement des forces de traction d’un groupe de fibroblastes embryonnaires de souris (MEF) adhérant à la fibronectine à micromotif (cercle, diamètre de 300 μm). L’amas a été éclairé par un faisceau de lumière UV de 300 μm de diamètre (ligne pointillée rouge). Barre d’échelle = 200 μm. (D,E) Une augmentation du stress oxydatif dans les cellules indiquée par l’augmentation de l’intensité du signal Cy5 après une dose élevée de laser UV-A est détectée par microscopie à fluorescence. Le stress oxydatif conduit à la mort cellulaire. Barre d’échelle = 200 μm. (F) Gauche : Carte thermique de contrainte d’un groupe de fibroblastes embryonnaires de souris (MEF) adhérant à la fibronectine à micromotif (cercle, 100 μm de diamètre). Droite : La carte thermique de stress d’un groupe de fibroblastes embryonnaires de souris (MEF) adhérant à la fibronectine à micromotif (cercle, diamètre de 300 μm). (G) Les cellules MEF adhérant à des micromotifs de fibronectine de 100 μm libèrent lentement leurs tractions après une exposition aux UV-A. La libération complète est obtenue après 15 min (l’image de référence a ensuite été prise 15 min après l’exposition). (H) Une comparaison avec le TFM conventionnel à base de trypsine pour les cellules MEF adhérant à la fibronectine à motifs de 300 μm de diamètre. Le résultat de l’éclairage UV-A (6 000 mJ/mm2) suivi d’une attente de 15 minutes n’est pas significativement différent du traitement conventionnel à la trypsine (c’est-à-dire 0,05 % pendant 20 min). Les barres indiquent une moyenne de S.E.M. ( test t de Student à deux queues, ****P < 0,0001, * P < 0,05, ns P ≤0,5). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
| Acrylamide (%) | Bis-acrylamide (%) | Acrylamide à partir d’une solution mère à 40 % (ml) | Bis-acrylamide à partir d’une solution mère à 2 % (ml) | Eau (ml) | Module de Young (kPa) |
| 4 | 0.1 | 1 | 0.5 | 8.5 | 5,74 ± 0,53 |
| 5 | 0.15 | 1.25 | 0.75 | 8 | 9,69 ± 0,68 |
| 5 | 0.3 | 1.25 | 1.5 | 7.25 | 11,33 ± 1,06 |
Tableau 1 : Mélanges de solution mère d’hydrogel et élasticité résultante
Toutes les données sont déposées dans le dépôt de données en libre accès de la Société Max Planck (Edmond) et peuvent être consultées à l’adresse suivante: https://edmond.mpdl.mpg.de/imeji/collection/JTu8PlWqpbymN9Qf
Discussion
Dans ce protocole, nous décrivons la préparation d’hydrogels DE PA à micromotifs contenant des billes fluorescentes, qui sont utilisés comme marqueurs fiducaux pour les études TFM. Notre approche est basée sur trois étapes : 1) préparation d’hydrogels PA à double couche ; 2) le micromodèlement des protéines ECM et leur transfert sur la surface de l’hydrogel; 3) utilisation de lumière proche UV à motifs pour TFM. La configuration expérimentale pour analyser les tractions cellulaires vers le substrat nécessite l’utilisation de matériaux élastiques linéaires avec des valeurs de rigidité connues pour calculer les forces liées au déplacement des billes fluorescentes26. Les hydrogels PA sont faciles à préparer, la rigidité peut être facilement réglée et ils sont couramment utilisés pour la détection de rigidité et TFM18,28. Cependant, pour obtenir des temps de polymérisation reproductibles et une polymérisation homogène de l’ensemble de l’hydrogel, il convient de prêter attention aux conditions de stockage et au temps de conservation des réactifs, par exemple, l’APS doit être conservé dans un dessiccateur pour éviter de perdre son activité; TEMED doit être protégé de la lumière directe. L’utilisation de HEA oxydé permet la liaison covalente de protéines matricielles à la surface de l’hydrogel, ce qui pourrait être avantageux pour obtenir la formation d’une couche protéique complète et stable. La solution HEA oxydée doit être préparée fraîche chaque fois que des hydrogels de PA sont fabriqués. L’hydrogel PA à double couche offre trois avantages principaux: 1) il offre un moyen alternatif d’obtenir de manière reproductible une distribution homogène des billes de fiducial près de la surface de l’hydrogel, sans avoir besoin de rendre le gel extrêmement mince (c’est-à-dire <20 μm). Le contrôle de l’épaisseur de l’hydrogel est crucial pour obtenir des mesures précises avec TFM. Lorsque le substrat élastique est trop mince, de fortes cellules adhérentes telles que les fibroblastes peuvent détecter et répondre mécaniquement au substrat de verre rigide sous-jacent29,30. Les hydrogels épais rendent l’acquisition d’images pour la reconstruction de la force plus difficile. De plus, de nombreux microscopes n’auront pas assez d’espace pour les accueillir, compte tenu de l’épaisseur supplémentaire du substrat de verre utilisé pour fixer l’hydrogel, à moins que des lames de microscopie ultramince ne soient utilisées. 2) Dans l’hydrogel PA à double couche, la distribution homogène des billes fiduciales près de la surface de l’hydrogel PA est obtenue sans utiliser de centrifugeuse, mais plutôt avec une simple incubation de quantités précises de solutions d’hydrogel et de perles fluorescentes. Une densité de billes élevée est d’un avantage significatif lors de l’analyse PIV car elle augmente la résolution des forces de traction et le rapport signal/bruit sans avoir besoin de microscopie confocale. 3) Le confinement des billes dans une fine couche proche de l’interface cellule-matériau permet d’imager les forces de traction avec des microscopes à épifluorescence ainsi que des microscopes confocaux. Lors de la préparation de l’hydrogel, l’utilisateur doit s’assurer qu’il adhère fermement au verre inférieur avant de procéder aux étapes suivantes du protocole. Nous recommandons de suivre le temps d’incubation indiqué pour la polymérisation des couches d’hydrogel, car il pourrait être difficile d’enlever le verre sur le dessus de la surface sans endommager la surface de l’hydrogel.
Les techniques les plus connues pour mesurer les propriétés élastiques sont l’AFM, la nanoindentation, les tests de traction et la rhéométrie. Cependant, la nanoindentation induit des contraintes très élevées sur les matériaux qui peuvent affecter la détermination des propriétés élastiques. Les essais de traction et la rhéométrie, quant à eux, sont des techniques de mesure macroscopiques, tandis que les cellules interagissent à l’échelle microscopique31,32. L’AFM permet des mesures à l’échelle microscopique avec des déformations réduites dans des conditions physiologiques. La fiabilité des mesures AFM peut être affectée négativement si des détails expérimentaux sont manquants (par exemple, la force d’indentation et la vitesse) ou si des données insuffisantes sont enregistrées27. Huth et al. décrivent un algorithme pour extraire les modules de Young des données AFM, qui met l’accent sur le maintien des détails de mesure constants27. Cet algorithme offre une détermination précise et fiable des modules de Young et a été utilisé pour nos expériences. De plus, nous avons mesuré de nombreuses courbes sur des échantillons fabriqués à différents jours et obtenu des résultats très similaires (variation des valeurs moyennes d’environ 1-2 kPa). Cela montre que la rigidité de nos gels peut être prédite de manière fiable.
Dans ce protocole, nous utilisons un module de photomotifs pour créer des régions micromotifs sur verre, qui sont ensuite transférées sur les surfaces d’hydrogel. Le micromodèlement montré dans ce protocole est basé sur la lithographie sans masque sans masque DMD (λ = 375 nm)7. Un DMD se compose d’un grand nombre de micromiroirs sur une puce. Un seul pixel correspond à un seul micro-miroir. Le fichier image de motif pixélisé d’un ordinateur est projeté via DMD et mis au point sur la surface à l’aide d’un objectif. Pour le micromodélisme des protéines, la lumière laser focalisée est utilisée pour cliver les brosses en polymère répulsives à l’aide d’un photoinitiateur. Ensuite, les régions exposées sont remplies de protéines ECM. Cette méthode d’ablation sans masque offre une grande flexibilité dans la conception de nouveaux motifs, car elle ne repose pas sur l’utilisation d’un photomasque. La conception et l’application d’un motif sont très faciles car cela ne prend que quelques minutes en utilisant un logiciel gratuit comme Inkscape. Cependant, le nombre de motifs et d’échantillons produits en peu de temps est un inconvénient majeur, car cette méthode ne peut être utilisée que pour modeler un seul substrat à chaque fois. Le module de photomotif utilise une source laser à semi-conducteurs proche des UV qui émet plusieurs milliwatts. Le faisceau laser non blindé est dangereux pour les yeux et la peau. La lumière et le rayonnement réfléchis et diffusés peuvent également être dangereux. La manipulation doit être accompagnée d’une consigne de sécurité de la part des agents laser. L’étape la plus critique du protocole lors de l’utilisation d’un module de photomodélisme est de s’assurer que pendant le micropatterning et l’ablation, le laser est correctement focalisé sur la surface. Une dose d’éclairage constante (intensité multipliée par le temps) de lumière UV pendant la modélisation dépend de la façon dont le laser est focalisé sur la surface. Une faible intensité à la surface due à une mauvaise mise au point peut entraîner l’échec du transfert de l’ECM à la surface de l’hydrogel, ce qui entraîne l’absence de cellules attachées à l’hydrogel.
Dans les expériences TFM, après l’imagerie initiale des cellules adhérentes et des marqueurs fiducial, les cellules sont libérées de l’hydrogel PA par traitement à la trypsine pour enregistrer leur état détendu. Un inconvénient de cette étape est la manipulation de l’échantillon au stade de la microscopie. Sans chambre de perfusion, l’ouverture du couvercle du plat, l’aspiration du milieu, le rinçage et le pipetage de la solution de trypsine représentent un défi pour les débutants et les utilisateurs expérimentés. En fait, ces procédures sont une source majeure de la dérive dans les axes xyz , entraînant une perte de position et de concentration. Notre protocole d’éclairage UV local fait du TFM une technique plus accessible pour les débutants. Il convient de noter que nous avons utilisé un module de photomotif sans masque basé sur la microscopie disponible dans le commerce, mais en principe, tout système d’éclairage UV-A pourrait être utilisé, éventuellement en combinaison avec un masque pour protéger les régions des substrats, où les forces de traction cellulaire ne devraient pas être enregistrées. L’exposition de cellules avec une dose importante de lumière visible de faible longueur d’onde (par exemple, la lumière violette qui excite l’émission de DAPI) entraînera une augmentation du stress oxydatif, ce qui peut entraîner une phototoxicité et la mort cellulaire. Par conséquent, cette technique peut être appliquée même dans un microscope à épifluorescence sans module laser UV. Cependant, comme l’intensité est beaucoup plus faible, il sera beaucoup plus facile d’accomplir la TFM avec le traitement enzymatique dans ce cas.
Avec LUVI-TFM, il est possible d’utiliser le même échantillon pour plusieurs mesures en raison du détachement local de cellules individuelles ou de petits groupes de cellules. Cependant, il faut faire attention à la sélection des cellules à détacher, pour éviter d’enregistrer les forces des cellules voisines. Ainsi, pour les mesures à cellule unique en l’absence de micromotifs, les régions surpeuplées doivent être évitées; pour les mesures sur des cellules à micromotifs uniques, les motifs doivent être conçus de telle sorte que la distance entre les structures individuelles soit au moins deux fois supérieure au diamètre de la zone à motifs. Nous recommandons également de ne pas utiliser les cellules de motifs adjacents dans l’ordre, mais plutôt de les échantillonner dans des régions éloignées sur le substrat. Notre mesure est bien réalisée sur un microscope à épifluorescence équipé d’une fonction de contrôle de la mise au point et d’une lentille 40 x air NA = 0,9, où la distance de travail de la lentille est suffisamment longue et le mouvement rapide d’un puits à l’autre est très accordable. Le détachement ciblé des cellules pour les applications TFM est efficace pour mesurer la force cellulaire d’une seule cellule ou d’un groupe entier de petites cellules (par exemple, jusqu’à 300 μm de diamètre). À l’aide de notre configuration, nous avons observé l’arrondi et le détachement des cellules pour le traitement UV des cellules individuelles (Figure 3A), alors que le détachement se produisait rarement pour les grappes de petites cellules. Cela pourrait conduire à une sous-estimation des forces de traction cellulaire. Pour les grappes de cellules plus grandes, le détachement enzymatique est recommandé, car les utilisateurs doivent utiliser un objectif d’air 20x pour imager l’ensemble du cluster et une fonction de contrôle de la mise au point est nécessaire. Comme la profondeur de mise au point de l’objectif d’air 20x est beaucoup plus longue, la manipulation ne sera pas aussi critique que l’objectif de grossissement plus élevé. L’utilisateur doit s’assurer que la mise au point est correctement réglée pour l’ablation des cellules, car la dose d’éclairage dépend de la mise au point laser sur la surface. Bien que nous soyons conscients des limites possibles de l’utilisation de LUVI-TFM dans les collectifs cellulaires en raison des interactions mécaniques possibles avec les cellules voisines non traitées, cet aspect pourrait en fait s’avérer utile pour les études sur la mécanique de l’extrusion cellulaire ciblée, par exemple à partir de monocouches épithéliales.
En conclusion, avec notre approche TFM combinée à la libération induite par la lumière de cellules micromotifs, nous fournissons un protocole robuste et à haut débit pour mesurer les forces d’adhésion cellulaire. La polyvalence de cette méthode pourrait être exploitée davantage en utilisant la microscopie et la configuration d’imagerie visant à améliorer la résolution et la sensibilité.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Nous remercions Mme Rebecca Alvarado pour le soutien apporté à la production vidéo du protocole et M. Stephen Casale pour les critiques constructives sur le manuscrit. Nous remercions les collègues du Département de biophysique cellulaire de l’Institut Max Planck de recherche médicale pour leurs discussions utiles. Le soutien financier de la Max-Planck-Gesellschaft à E.A.C.-A. et la Deutsche Forschungsgemeinschaft (DFG SFB1129, Projektnummer 240245660, P15 à E.A.C.-A. et P4 à U.S.S.; DFG EXC 2082/1-390761711 à U.S.S.) est également grandement reconnu. J.B. remercie la Fondation Carl Zeiss pour son soutien financier. E.A.C.-A., C.S. et U.S.S. reconnaissent le financement par le biais de l’école Max Planck Matter to Life soutenu par le ministère fédéral allemand de l’Éducation et de la Recherche (BMBF). C.S. est soutenu par le Conseil Européen de la Recherche (Consolidator Grant PHOTOMECH, n°101001797).
matériels
| Name | Company | Catalog Number | Comments |
| 3-(Trimethoxysilyl)propyl methacrylate | #440159 | Sigma Aldrich | |
| Acetic acid | #33209 | Honeywell | |
| Acrylamide 40 % | #1610140 | Bio-Rad | CAUTION : toxic, work under a fume hood |
| AFM cantilever | #CP-CONT-BSG-A-G | NanoAndMore | 5 μm spherical tip |
| AFM system | #NanoWizard3 | JPK | Coupled to optical microscope equipped with 40x air objective and GFP filter |
| Ammonium persulfate | #A3678 | Sigma | |
| Bis-acrylamide 2 % | #1610142 | Bio-Rad | CAUTION : toxic, work under a fume hood |
| Camera sCMOS | #C11440-42U30 | Hamamatsu | |
| Camera sCMOS | #ANDORZYLA4.2 | Oxford Instrument | |
| CellRox | #C10422 | Thermo Fisher | |
| Custom incubator chamber | EMBLEM | ||
| Dental glue | #1300 100 | Picodent | |
| DMEM | #41965 | Thermo Fisher | |
| Epifluorescence microscope | #Eclipse Ti2E | Nikon | |
| Epifluorescence microscope | #Axiovert200 | Zeiss | |
| Ethanol | #9065.3 | Carl Roth | |
| FBS South America | #S181T | Thermo Fisher | |
| Fibrinogen | #F13191 | Invitrogen | Alexa488 conjugate |
| Fibronectin | #F1141 | Sigma | |
| Fluorescent beads 200 nm | #F8805 | Invitrogen | Carboxylated (365/415) |
| Fluorescent beads 200 nm | #F8848 | Invitrogen | Carboxylated (505/515) |
| Fluorescent beads 200 nm | #F8810 | Invitrogen | Carboxylated (580/605) |
| Fluorescent beads 200 nm | #F8807 | Invitrogen | Carboxylated (660/680) |
| Glass coverslip 15 mm round | #41001115 | Assistent | |
| Glass coverslip 24 mm round | #41001124 | Assistent | |
| Microscope Objective | #MRH08230 | Nikon | 20x air NA 0.45 |
| Mouse Embryonic Fibroblasts | #CRL-2991 | ATCC | |
| mPEG-SVA | #MPEG-SVA-5000 | Laysan Bio | |
| N-Hydroxyethyl acrylamide | #697931 | Aldrich | |
| Plasma cleaner | #100-E | TePla | |
| PLPP gel | #PLPPgel | Alveole | |
| Poly-L-lysine | #P4823 | Sigma Aldrich | |
| Poly(L-lysine)-graft-poly(ethylene glycol) | #PLL(20-g[3.5]-PEG(2) | SuSoS | |
| Sodium(meta) periodate | #S1878 | Sigma Aldrich | |
| TEMED | #17919 | Thermo Scientific | |
| UV Patterning module | #PRIMO | Alveole | |
| UVO cleaner | #342-220 | Jelight |
Références
- Geiger, B., Bershadsky, A. Exploring the neighborhood: adhesion-coupled cell mechanosensors. Cell. 110 (2), 139-142 (2002).
- Beningo, K. A., Hamao, K., Dembo, M., Wang, Y. -l, Hosoya, H. Traction forces of fibroblasts are regulated by the Rho-dependent kinase but not by the myosin light chain kinase. Archives of Biochemistry and Biophysics. 456 (2), 7(2006).
- Rape, A. D., Guo, W. H., Wang, Y. L. The regulation of traction force in relation to cell shape and focal adhesions. Biomaterials. 32 (8), 2043-2051 (2011).
- Kumar, R., Saha, S., Sinha, B. Cell spread area and traction forces determine myosin-II-based cortex thickness regulation. Biochimica Biophysica Acta. Molecular Cell Research. 1866 (12), 118516(2019).
- Chen, C. S., Mrksich, M., Huang, S., Whitesides, G. M., Ingber, D. E. Geometric control of cell life and death. Science. 276 (5317), 1425-1428 (1997).
- Thery, M. Micropatterning as a tool to decipher cell morphogenesis and functions. Journal of Cell Science. 123, 4201-4213 (2010).
- Strale, P. O., et al. Multiprotein printing by light-induced molecular adsorption. Advanced Materials. 28 (10), 2024-2029 (2016).
- Azioune, A., Carpi, N., Tseng, Q., Théry, M., Piel, M. Microtubules: in vivo Methods in Cell Biology. , 133-146 (2010).
- Tseng, Q., et al. A new micropatterning method of soft substrates reveals that different tumorigenic signals can promote or reduce cell contraction levels. Lab on a Chip. 11 (13), 2231-2240 (2011).
- Vignaud, T., Ennomani, H., Thery, M. Polyacrylamide hydrogel micropatterning. Methods Cell Biology. 120, 93-116 (2014).
- Munevar, S., Wang, Y., Dembo, M. Traction force microscopy of migrating normal and H-ras transformed 3T3 fibroblasts. Biophysical Journal. 80 (4), 1744-1757 (2001).
- Style, R. W., et al. Traction force microscopy in physics and biology. Soft Matter. 10 (23), 4047-4055 (2014).
- Polacheck, W. J., Chen, C. S. Measuring cell-generated forces: a guide to the available tools. Nature Methods. 13 (5), 415-423 (2016).
- Schwarz, U. S., Soine, J. R. Traction force microscopy on soft elastic substrates: A guide to recent computational advances. Biochimica Biophysica Acta. 1853, 3095-3104 (2015).
- Roca-Cusachs, P., Conte, V., Trepat, X. Quantifying forces in cell biology. Nature Cell Biology. 19 (7), 742-751 (2017).
- Hanke, J., Probst, D., Zemel, A., Schwarz, U. S., Koster, S. Dynamics of force generation by spreading platelets. Soft Matter. 14 (31), 6571-6581 (2018).
- Butler, J. P., Tolic-Norrelykke, I. M., Fabry, B., Fredberg, J. J. Traction fields, moments, and strain energy that cells exert on their surroundings. American Journal of Physiology. Cell Physiology. 282 (3), 595-605 (2002).
- Sabass, B., Gardel, M. L., Waterman, C. M., Schwarz, U. S. High resolution traction force microscopy based on experimental and computational advances. Biophysical Journal. 94 (1), 207-220 (2008).
- Willert, C. E., Gharib, M. Digital particle image velocimetry. Experiments in Fluids. 10, 181-193 (1991).
- Lourenco, L., Krothapalli, A. On the accuracy of velocity and corticity measurements with PIV. Experiments in Fluids. 18, 421-428 (1995).
- Adrian, R. J., Westerweel, J. Particle Image Velocimetry. , Cambridge University Press. (2011).
- Westerweel, J., Scarano, F. Universal outlier detection for PIV data. Experiments in Fluids. 39 (6), 1096-1100 (2005).
- Dembo, M., Wang, Y. Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophysical Journal. 76 (4), 2307-2316 (1999).
- Schwarz, U. S., et al. Calculation of forces at focal adhesions from elastic substrate data: the effect of localized force and the need for regularization. Biophysical Journal. 83 (3), 1380-1394 (2002).
- Huang, Y., Gompper, G., Sabass, B. A Bayesian traction force microscopy method with automated denoising in a user-friendly software package. Computer Physics Communications. 256, 107313(2020).
- Vedadghavami, A., et al. Manufacturing of hydrogel biomaterials with controlled mechanical properties for tissue engineering applications. Acta Biomaterialia. 62, 42-63 (2017).
- Huth, S., Sindt, S., Selhuber-Unkel, C. Automated analysis of soft hydrogel microindentation: Impact of various indentation parameters on the measurement of Young's modulus. PLoS One. 14 (8), 0220281(2019).
- Oria, R., et al. Force loading explains spatial sensing of ligands by cells. Nature. 552 (7684), 219-224 (2017).
- Buxboim, A., Rajagopal, K., Brown, A. E., Discher, D. E. How deeply cells feel: methods for thin gels. Journal of Physics. Condensed Matter. 22 (19), 194116(2010).
- Sen, S., Engler, A. J., Discher, D. E. Matrix strains induced by cells: Computing how far cells can feel. Cell and Molecular Bioengineering. 2 (1), 39-48 (2009).
- Arnold, M., et al. Induction of cell polarization and migration by a gradient of nanoscale variations in adhesive ligand spacing. Nano Letters. 8 (7), 2063-2069 (2008).
- Galluzzi, M., et al. Atomic force microscopy methodology and AFMech Suite software for nanomechanics on heterogeneous soft materials. Nat Communications. 9 (1), 3584(2018).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.