Method Article
הקלטות ארוכות טווח בו זמנית בשני שלבים עצביים עיבוד במתנהגים דבורי דבש
In This Article
Summary
הקלטות ארוכות טווח תאיים בו זמנית משני neuropiles המוח שונה או שני קטעים אנטומיים שונים הוקמו בדבורי דבש. הקלטות אלה מאפשרות החקירה של היבטים זמניים של עיבוד עצבי על פני אזורים שונים במוח בנוירון הבודד, כמו גם ברמת ההרכב בבעלי חיים מתנהגים.
Abstract
בשני יונקים וחרקים מידע עצבי מעובד במרכזים במוח מסדר גבוה יותר ונמוך יותר שונים. מרכזים אלה הם מצמידים באמצעות מתכנסים וקשרים אנטומיים מסתעף כולל הזנה קדימה וחיווט משוב. יתר על כן, מידע מאותו המוצא נשלח באופן חלקי דרך שבילים מקבילים לשונה ולפעמים לאותם אזורים במוח. כדי להבין את היתרונות אבולוציוניים, כמו גם את היתרונות חישוביים של אסטרטגיות החיווט הללו ובמיוחד התלות הזמנית שלהם אחד על השני, זה הכרחי כדי לקבל גישה בו זמנית לנוירונים בודדים של שטחים או neuropiles שונים באותה ההכנה ברזולוציה גבוהה זמנית. כאן אנו להתרכז בדבורי דבש על ידי הוכחת גישה לטווח ארוך תאית ייחודית להקליט פעילות יחידה רבת בשתי neuropiles הבא 1, באונה המחושים (AL), השלב הראשון חוש הריח העיבוד וגוף הפטרייה (MB), invo מרכז אינטגרציה מסדר גבוה יותרlved בלמידה וביצירת זיכרון, או שני קטעים עצביים במקביל 2 חיבור AL עם מגה בייט. האחרון נבחר כדוגמה ויהיה מתואר במלואו. בסרטון התמיכה בבנייה וההחדרה קבועה של אלקטרודות חוט רבת ערוצים גמישים באו לידי ביטוי. הגברה ההפרש Pairwise של ערוצי אלקטרודה חוט מיקרו באופן דרסטי מפחיתה את הרעש ומוודאת כי המקור של האות קשור קשר הדוק למצב של הקצה האלקטרודה. הגמישות מכאנית של אלקטרודות החוט משמשים מאפשרת הקלטות ארוכות טווח פולשנית יציבה לאורך שעות רבות עד ימים, וזה יתרון ברור בהשוואה לטכניקות הקלטה קונבנציונליות נוספות ותאיות בגוף חי.
Introduction
דבורי דבש, כמו גם רוב חרקים אחרים להסתמך בכבדות על חוש ריח. בין השאר הם משתמשים ברמזי חוש הריח להתמצאות, הזדווגות, תקשורת עם בני מין, וחיפוש אחר מזון. מערכת חוש הריח פירט גם תורמת לרפרטואר עשיר של התנהגויות למידה הקשורות לגירויי ריח פרחוניים. ניתן ללמוד התנהגויות אלה בקלות בתנאי מעבדה מבוקרת (לסקירה ראתה 3 - 5). "המוח של המיני" שלהם (cp. 6) עם המספרים הקטנים יחסית של נוירונים עושה דבורת דבש אורגניזם מודל מתאים גם ללימוד קידוד חוש הריח והלמידה במהלך הניטור של פעילות עצבית.
מערכת חוש הריח בחרקים, כמו גם ביונקים מציגה ארגון מקביל במידה רבה (לסקירה ראה 7,8). בדבורי דבש כ -80,000 קולט נוירונים 9 ממוקמים בsensillae לאורך האנטנות 10,11 לתרגם את גירוי הריח הסביבתי לneurאות onal. האקסונים מתאי עצב קולטני ריח מעצבבים את אונת המחושים (AL), שבה יש ארגון גלומרולרי דומה לנורת חוש הריח החוליות. AL מהווה כ 164 glomeruli מחובר אחד עם השני על ידי כ -4,000 interneurons המקומי (LN) (לסקירה ראה 12). במיוחד בדבורת הדבש זה הוכח לאחרונה כי LNs לספק קישוריות לרוחב אחידה ושאוכלוסיות שונות בעלי תכונות הרחה יסודות וconfigural קידוד 13,14. AL הוצג להיות מחולקים לאונת הגחון וחמים הגבי והוליד המדיאלי והדרכים לרוחב מחושים אונה (m-ו-L-ALT; לשעבר כינה מ-ו-L-APT להמדיאלי וprotocerebral אונת מחושים לרוחב מערכת 15-17). הנה מינוח בדרכי חדש הוצג על ידי מאמץ האחרון למינוח אחיד של מוח החרק ישמש 18. שני alts (l-ו-M-ALT) לשלב גם 410 proje uniglomerular (l-ALT) או 510 (m-ALT)נוירונים ction (PN), בהתאמה 15,16,19. לאחרונה PNS של שני קטעים הוכחו ריחות קוד ב2 מקבילים (לסקירה ראה 17,20), ושני קטעי synaptically ליצור קשרים מסועפים עם תאי קניון (KC), גוף הפטרייה (MB) נוירונים עיקריים. כל MB מכיל כ 172,000 KCS 21-23. מ"ב ידועים להיות מעורב בשילוב גירוי, למידה, ויצירת זיכרון. דנדריטים axo של KCS להקים peduncle (הגזע של הפטרייה), שבו יש שני אזורי פלט עיקריים: vertica או אלפא אונה ו22,24 אופקיים או בטא אונה. הפלט של MB מתכנס רק על 400 נוירונים חיצוניים (EN) 24. ENS אחראי על עיבוד מידע הרחה בעיקר מעצבב את היבט הגחון של האונה האנכית 22. לאחרונה, זה כבר הראה כי ENS נרשם באזור זה לקודד את עמותת הפרס ריח 25.
זמני כמוpects בתוך מערכת חוש הריח של חרקים, כמו גם בעלי חוליות הפכו היבט חשוב ומשמעותי כעיקרון קידוד פוטנציאל 26-29. כדי להיות מסוגל להקליט בו זמנית מספר רב של נוירונים מאתרים שונים ברזולוציה גבוהה זמנית, הקמנו טכניקות הקלטת יחידה כפולות רב באמצעות אלקטרודות תיל ערוץ מותאמות אישית רב הציגו לאזורי יעד שונים במערכת ההרחה של הדבורה. גישה זו מאפשרת לנו לנתח ולהשוות בין עיבוד זמני במערכת ההרחה דבורת הדבש ברמה של תאי עצב ואוכלוסיות של תאי עצב או בין מסלולי הרחה מקבילים, המסלול הכפול ההרחה 2 או בין neuropils שלאחר מכן שונה 1 אחד. לאחרונה עם גישה ניסויית דומה במערכת ההרחה ארבה 30 שימוש בתצורה של אלקטרודות שונה היו מסוגלים לנתח את מנגנון קידוד spatiotemporal להכרה ריח רקע בלתי משתנה ביום 31. האותנו, את ההקלטות כפולים הוקמו מאפשרות איסוף מידע מרחבי על פרופילים של פעילות עצביות בו זמנית.
בהשוואה לדגימה מרחבית הרחבה יותר המתקבלת מהדמית סידן בשיטה זו מאפשרת הקלטה משתי נקודות בלבד. עם זאת, היתרון בהשוואה לשיטות הדמיה סידן הוא הדיוק הגבוה הזמני של הקלטות פוטנציאל פעולה, אשר אינו יכול להיות מסופקות על ידי או הדמיה CCD קונבנציונלית או רכישת הדמיה 2 פוטונים. אלקטרודות תאי שתוארו כאן באופן קבוע מושתלות וקבוע ביחס למוח וכמוסת ראש הימנעות נדידת האלקטרודה. זהו יתרון ברור בהשוואה לשימוש באלקטרודות תאית חדה. יתרון נוסף בהשוואה להקלטות תאית והדמיה סידן הוא זמן תצפית עצבית המורחב החל משעות רבות עד ימים. זה תנאי חשוב לחקור וקושרת עצבית של היווצרות למידה וזיכרון. יתרונות נוספים של רבהקלטות יחידה מתוארות נוסף בסעיף הדיון.
בסקירה מתודולוגית זה הליך הייצור של אלקטרודות תיל עיצוב המותאם אישית תוכלו לראות, שהותאם מ32,33 ומתאים להקלטות יחידה רבת לטווח ארוך במוח דבורת הדבש. בנוסף, דוגמא כיצד סוגים אלה של האלקטרודות מושתלים באופן קבוע בשני אתרי הקלטה שונים בתוך מערכת ההרחה דבורת הדבש כדי להקליט בו זמנית l-וה-M-ALT על פני תקופות זמן כדי לאפשר לפרוטוקולי גירוי רבים ארוכות מוצגת 2. לצורך אימות עמדות הקלטת דוגמא ופרוטוקול להדמיה מכתים והודעת הקלטה של אתרי ההקלטה מסופקת.
Protocol
1. אלקטרודה בנייה (איור 1)
- ייצור של מתאם אלקטרודה שמתאים את האלקטרודה ממשק הלוח של 1,2,25 מערכות מגבר רב ערוצים מסחריים.
- השתמש בצלחת פלסטיק קטנה דבוקה לבסיס מחבר 18 פין.
- חבר את הבסיס עם 3 קטעים קצרים של תיל מבודד ל3 זיזי הלחמה נפרדים מוברגים לצלחת הפלסטיק (איור 1 A1-A3).
- הכנס חריץ לתוך צלחת הפלסטיק שבו נימי זכוכית יכולות בקלות לעבור ולהיות מוחזקות במקום על ידי בורג (איור B1 1).
- להאריך את נימי זכוכית על 5 מ"מ באמצעות סיכת minutien.
- צרף את חוטי האלקטרודה מיקרו לאורך סיכת minutien ונימי הזכוכית כדי להבטיח ייצוב ותמיכה.
- ייצור חוטי מייקר רב ערוצית (שאומץ מRyuichy אוקאדה 32,33)
- תוחלת 3 חוטי מיקרו (חוטי נחושת פוליאוריטן מצופים, 15קוטר מיקרומטר), באופן שהם ממוקמים אחד ליד שני (איור B2 1).
- שימוש במחט הלחמת 12 V להפיץ שכבה דקה של שעוות שיניים התכה נמוכה (C ° 50) באופן חלקי לאורך החוטים כדי להדביקם (קצה האלקטרודה) (איור 1 B3). השאר כמה סנטימטרים unglued (סוף אלקטרודה) כסעיף זה ישמש מאוחר יותר לחבר את חוטי מיקרו עם מתאם האלקטרודה.
- חבר את כבל מיקרו הרב ערוצים למתאם אלקטרודה
- הסר את נימי זכוכית מהמחזיק ולצרף אותו עם סיכת minutien לקצה האלקטרודה. להביא אותו לעמדה מקבילה למייקרו אלקטרודה (B3 Figure1).
- מדביקים את הקצה האלקטרודה לפין minutien באמצעות שעוות שיניים התכה נמוכה ולחתוך את האלקטרודה מיקרו בקצה, בולט 2-3 סנטימטר מפיני minutien ובסוף האלקטרודה (חיצים קטנים איור B3 1).
- Slip sl הנימיםightly בחזרה לתוך מתאם האלקטרודה. השתמש בבורג כדי לתקן את זה (איור B4 1).
- הלחם את הקצוות החופשיים של שלושה חוטים לזיזי הלחמה באמצעות אקדח הלחמה עם טמפרטורה של כ 360 ° C כדי להבטיח היתוך של הבידוד (איור B4 1). לאחר הלחמה, להבטיח כי יש קשר נאות חשמלי (~ 300 kOhm).
- הר אחד של אלקטרודות (אב) לאלקטרודה ממשק הלוח של headstage ולתקן את האלקטרודה האחר רב ערוצים (עבדים) במתאם נפרד. חבר את הערוצים של העבד להאלקטרודה השנייה (איור 1 C1). בנוסף, לרתך את ההתייחסות, כמו גם אלקטרודות שרירים לבסיס האלקטרודה השנייה (איור 1 C2).
2. Bee הכנה (איור 2)
בניסויים שתוארו אלה, דבורת הדבש (Apis mellifera), אשר הוא בעלי חיים חסרי חוליותולכן אינו דורש אישורים אתיים ספציפיים לשימוש, הוא בשימוש.
- לתפוס מלקטי מזון דבורת דבש (mellifera א) בכניסה הכוורת בבוקר, כפי שמוצג על ידי אחרים 34,35.
- צ'יל הדבורים על קרח כתוש עד חוסר תנועה (5-10 דקות) ולתקן אחד בבעל פרספקס סטנדרטי או צינור מתכת בצורה שהראש חשוף (איור 2). לצמצום תנועות הראש להשתמש בשעוות שיניים התכה נמוכה (~ 50 מעלות צלזיוס) ולתקן את הראש בזהירות לבעל סביב הבסיס של העיניים מורכבות והצוואר.
- השתמש בשעוות התכה נמוכה לקבע scapi של האנטנות על הקפסולה הראש (איור 2 ב ') בלי לגעת שבוטון. השוטון של האנטנה צריך להיות הצביע קדימה. ודא שהדבורה יכולה להזיז חוטם שלה באופן חופשי.
- לגלח את ראש הקפסולה כדי להבטיח נוף וגישה לחלק העליון של הראש באין מפריע.
- להאכיל את הדבורים בתמיסת סוכרוז 30% עד saturatiעל מנת להבטיח הרטבה מספקת של רקמת המוח ויכולת קיום טוב של בעל החיים (איור 2 C).
- לעשות חתכים זהירים אנכי לאורך הגבולות של העיניים מורכבות ואופקיים מעל בסיסי מחושים כמו גם מתחת לocelli ולהסיר את החתיכה הרופפת של ציפורן (איור 2 ד ').
- להגדיר בזהירות בצד בלוטות hypopharyngeal ולהסיר את קנה הנשימה כדי להבטיח נוף וגישה למוח לפני אלקטרודה הכנסה (2E דמויות, 2F) ברורים.
3. אלקטרודה קלטי
במקרה הדוגמא באיור 2, אלקטרודה אחת ממוקמת מכוון l-ALT, והשני מכוון על M-ALT 2. באמצעות ציוני דרך מסוימות, אזורי יעד אחרים אפשריים, כמו גם, לדוגמא אזורי הפלט אל על וMB 1.
- מקם את האלקטרודות באמצעות micromanipulators באזור של עניין ( איור 2G ו 3 א). כדי למקד את המקום מ '-ALT האלקטרודה בין AL ומדיאלית מהאונה האנכית של מגה בייט. ודא להיות מעל נקודת PNS ALT mediolateral הסתעפות אתר ההכנסה. בולט אלקטרודה למוח עם עומק של כ -180 (2E איור) מיקרומטר. למקום הקלטת l-ALT PN האלקטרודה מתחת protocerebrum לרוחב (LH) באמצע קו דמיוני בין הצד לרוחב של אונה אנכית ואמצע AL. הכנס את האלקטרודה עם עומק של כ -300 מיקרומטר (איור 2E)
- הכנס את ההתייחסות (חוט כסף, על 25 מיקרומטר קוטר) לעין המורכבת ipsilateral דרך חתך קטן בציפורן. הכנס חוט כסף נוסף לאזור הקרנת שרירים מתחת לocelli לרוחב. הערה: אם יש צורך בהתנהגות של דבורי הלמידה יכולה להיות במעקב עם דיוק גבוה זמני על ידי הקלטת השרירים M17, שהוא מעורב in תגובת הארכה חוטם (PER) של הדבורה 36 כפי שמתוארת ב25.
- לעגן באופן יציב אלקטרודות בתוך המוח ואת הקפסולה הראש, מכסה את החלל כולו מעל המוח עם שני סיליקון הרכיב (איור 2H), אשר ימנע את המוח מהתייבשות. הערה: ההקלטות יכולות להימשך שעות עד ימים, והוא יכול למשל להיות מוקלטים הדבורים במהלך הליך קלאסי אוויר (איור 2H) או מגורה עם פנל גדול של ריחות שונים.
4. רכישת נתונים ועיבוד מקדים
- השתמש בתוכנת רכישה ראויה שעונה על הדרישות הבאות: קצב דגימה של kHz דקות 25; אנלוגי 1,25 או בידול צלב 2 דיגיטלי בין ערוצי האלקטרודה; להקה לעבור סינון מ300 הרץ עד 8,000 Hz לחלץ אירועי ספייק.
- השתמש בתוכנת ספייק מיון זמינה כדי לחלץ את פעילות יחידה אחת, למשל templאכל טכניקות התאמה כפי שנכלל בתוכנת Spike2 (איור 3).
- לניתוח נוסף להשתמש בבולים של יחידות חילוץ בזמן כדי לחשב יחידה אחת בממוצע תגובות ריח (איור 4) או לחשב וקטורי אוכלוסייה לקרן רכיב ניתוח (PCA) (איור 5) באמצעות תוכנה זמינה באופן מסחרי. כדי לנתח יחידה אחת וחביון תגובת אוכלוסייה נוסף אנא להשוות פרסומים אחרונים 1,2,25.
5. ויזואליזציה של עמדת אלקטרודה יחסית (איור 4)
- טובלים את הטיפים אלקטרודות לתוך תמיסה של 5% או Alexa hydrazide 568 או 5% Alexa hydrazide 488 אשר מומסים ב0.5 כלוריד פתרון M אשלגן לפני ניסויי ההקלטה.
- הסר את האלקטרודות וסיליקון הכיסוי בקפידה לאחר הניסויים, יש לשטוף את המוח עם פתרון Ringer הדבורה, להסיר את הבלוטות ואת קנה הנשימה ולהכניס גבישים זעירים של tetramethylrhodamin dextran או להכניס פתרון 5% פתר ב1.0 אצטט M אשלגן לתוך AL לתייג alts anterogradely. בצע את השלבים הבאים בחושך.
- לאפשר לצבע שיש לנקוט ומועבר על ידי נוירונים ההקרנה לאורך שטחי אקסון (30-45 דקות) (איור 4 א), לפני שטיפת המוח עם פתרון Ringer דבורים שלוש פעמים לעוד דקות 30-45.
- צ'יל הדבורה על קרח עד חוסר תנועה ולהסיר בזהירות את המוח מכמוסת הראש. לקבע את המוח על ידי שטיפתו בתמיסת 0.1 M PBS המכיל פורמלין 4% ולשמור אותו לילה בשעה 4 ° C.
- לחכות שעה לפחות 12 לפני שטיפת המוח פי שניים ב0.1 M PBS (10 דקות כל אחד).
- שטוף את המוח 3x עבור 20 דקות ב0.2% טריטון X-100 מדוללים ב0.1 M PBS לפני התייבשותו בסדרת אלכוהול עולה (30%, 50%, 70%, 90%, 95%, 3x 100% אתנול, 20 דקות כל שלב).
- שבץ המוח מיובש בMethylsalicylate בשקופית מיקרוסקופ ויםeal אותו עם כיסוי שקופיות.
- השתמש במיקרוסקופ לייזר confocal סריקה ולסרוק את המוח, כמו חלקים אופטיים כל מיקרומטר 2-5 באמצעות תרכובת הרמונית מטרת התכנית Apochromat (10X טבילה 0.4 NA). להלהיב את הרקמות באמצעות אורך גל 568 ננומטר לdextran tetramethylrhodamin ואורך גל של 488 ננומטר לתפקיד האלקטרודה.
- לשחזר את המבנים המוכתמים המוח ונתיבי האלקטרודה מערימות התמונה ב-3D עם תוכנת שחזור (למשל. עמירה או פיג'י) (איור 4).
תוצאות
"הפרוטוקול הנוכחי מאפשר הקלטות בו זמנית בשני שלבי עיבוד שונים בתוך דבורים בודדים ובנוסף מאפשר לבחון את המנגנון בסיסי של למידה וזיכרון באמצעות דוגמא., PER האוויר בתוך דבורי דבש מאופקים". זהו תנאי הכרחי לניתוח היבטים זמניים של עיבוד עצבי. השיטה היא להתאמה בקלות לגישות מדעיות שונות לפענח את הרשת העצבית של מערכת חוש הריח של הדבורה. לדוגמא שיטה זו משמשת (i) כדי לנתח עיבוד זמני של PNS בתוך מסלול ההרחה הכפול של דבורת הדבש, l-ו-M-ALT PNS (איור 5). באיור 5A דוגמא אחת PN l-ALT נרשם בו זמנית עם PN של m-ALT ניתן כעשר ממוצע משפט וממחישה את כוחם התגובה והשהיה בגין חמישה ריכוזי ריח שונים כמו עלילת חום בקודי צבע. בממוצע של שבעה דבורים עם 11 l-ו13 mALT PNS (5B דמויות, 5C) ממחיש כי גם כוח התגובה, כמו גם ההשהיה התגובה, במידה מסוימת, משקף את ריכוז odorant. כך PNS בדוגמא זו להגדיל את כוח התגובה שלהם בזמן עם ריכוז odorant הגדלת חביון התגובה שלהם ירד (5B דמויות, 5C). תוצאה זו היא מוגבלת למדי ותקפה רק לodorant ניתח, אבל הוא עדיין עולה בקנה אחד עם מודלים חישוביים האחרונים של 37 AL. אם קידוד ריכוז הריח בAL של הדבורה בבסיס חישובים לא ליניארי או בבסיס מאפייני קידוד אחרים עדיין צריך להיות מנותחים בעתיד. יתר על כן השיטה יכולה לשמש (ii) כדי להשוות היבטים זמניים בפעילות האוכלוסייה בשני שלבים שלאחר מכן עיבוד, AL-וMB-הפלט (איור 6). ניתוח עיקרי רכיב (PCA) ממחיש כי חישוב ריח הוא ממושך ולשרוד את מצגת הריח כולו ברמת PN ואילו בENזה רק הריח לסירוגין סט היו מיוצג בפעילות האוכלוסייה (איור 6). וכך אוכלוסיית EN הגיעה הפעילות המקסימלי שלהם כבר בנקודת זמן שבו פעילות PN עדיין מתפתחת (סרט cp. 1).

.. איור 1 ייצור אלקטרודות מיקרו חוט שלושת ערוצי A1) הסיומות של שלושה חוטים מולחמות לפיני IC בפוזיציות 11, 13 ו16 A2) ארבעה זיזי הלחמה דפוקים על צלחת בסיס פלסטיק פרספקס.; סיכת minutien מוכנסת לתוך הקצה של נימי זכוכית אשר מחוברת אז לצלחת הפלסטיק. A3) צלחת הפלסטיק מודבקת על גבי פיני IC ואת הקצה החופשי של כל חוט הוא מולחם לאחד מההלחמה העליונה זיזים. B1) כדי לצייד את בעל אלקטרודה עם חוטי נחושת הדקים, הנימים יש לקחת החוצה. B2 פעם נוסף) הבסיס של קרס ואז קבועים במנהג עשה יישור מכשיר. שלושה חוטי מייקר נחושת מיושרים לאורך החריץ וקבוע עם דבק בכל קצה B3) חוטי המייקר המקבילים מודבקים יחד עם שעוות שיניים והנימים היא להחזיר במקום (חיצים ארוכים).; אז חוטי מיקרו דבוקים מצורפים לנימים בשעוות שיניים וקצותיו מנותקים (חיצים קצרים). B4) שלושה הקצוות החופשיים של חוטי הנחושת מולחמים לשלושה זיזי ההלחמה העליונים ובכך להביא אותם למגע חשמלי עם ניתן לחבר שתי אלקטרודות פיני IC. C1) התאספו באופן מלא לכל אחד אחר לשימוש עם headstage אחת. C2) לצורך כך את הסיכות של אלקטרודה אחת (משמאל, עבדים) מחוברות באמצעות חוטים מבודדים לאלקטרודה אחרת (מימין, תורןER), אשר מחובר לבמה את הראש; האלקטרודה headstage מחוברת יכולה גם לאסוף קלט משריר (M17) ואלקטרודה השוואתית (נ"צ).

איור 2. הכנה והחדרת אלקטרודה קבועה למוח הדבורה.) דבורה מוכנסת לתוך בעל פרספקס לאחר קיבוע על קרח. אנטנות עם השוטון (פלורידה) וScapus (SC) מצוינים. ב ') את הראש ואנטנות קבועות באמצעות שעוות שיניים. C) הקפסולה הראש מגולחת והדבורים מוזנים במי סוכר. ד') את הראש הקפסולה נפתחה. E) לאחר הסרת בלוטות וקנה נשימה מהחלק העליון של המוח, יכולים בקלות להיות מכובדת neuropiles שונה וציוני הדרך העיקרית. המסלולים של alts מסומנים יחד עםסימן שבו האלקטרודות מוכנסות. (MB: גוף פטריות, אל על: אונת מחושים, OL: אונה אופטית, α: אונת אלפא או באונה אנכית, ALT: מערכת אונת מחושים, E1: צד החדרת אלקטרודה להקלטות מ-ALT, E2: צד החדרת אלקטרודה לl-ALT הקלטות). F) הפניה (REF) ואלקטרודות שריר (M17) מוכנסות לתוך הקפסולה הראש דרך חורים קטנים בציפורן או עין המורכבת. G) אלקטרודות החוט מוכנסות לתוך המוח באתרים המתאימים. H) לאחר תיקון אלקטרודות במקום באמצעות שני סיליקון רכיב, הדבורה עדיין מראה PER ויכולה להיות מותנית (למשל., שימוש במי סוכר).

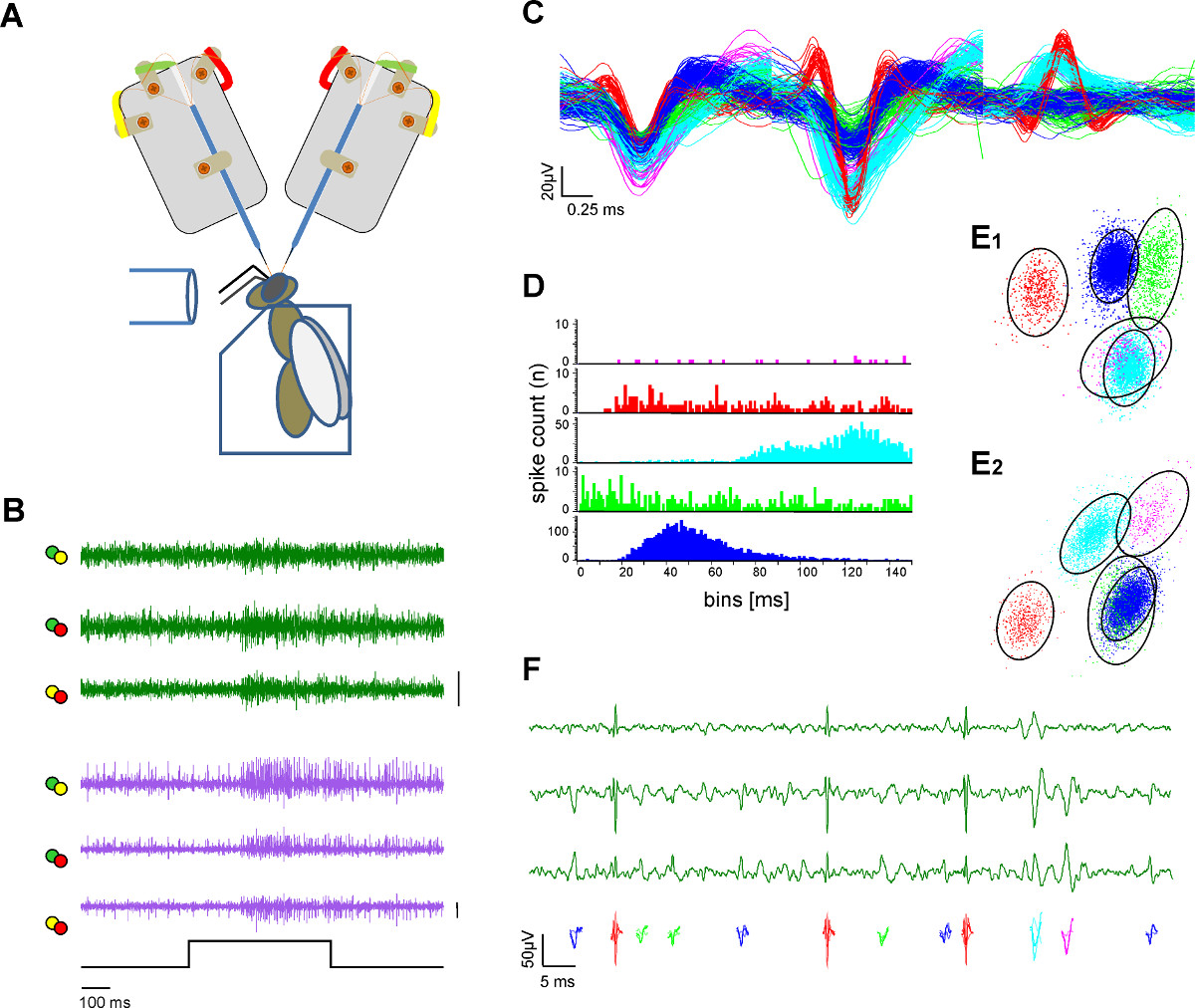
הקלטת איור 3. תאי בשני שטחים עצביים והפקה אחת יחידה (מיון ספייק). ) ציור סכמטי של הגדרת הניסוי. הדבורה היא מקובעת בבעל פרספקס. גירוי ריח מסופק באמצעות שפופרת זכוכית. שני אנקס אלקטרודה מקליט ממוח הדבורה החשוף. ב ') הקלטות בו זמנית מl-ו-M-ALT PNS (ירוק ועקבות סגולות) מראה תגובות מעוררות בשני קטעים לגירוי 500 ריח אלפיות של דבש בפתרון מים בריכוז של 1:100 ב33 ° C. כל שורה זממה מייצגת את הערוצים מובחנים כתווית בצבעים מזיזי הלחמת אלקטרודה בבר א:. 50 μV ג) לאחר הליכי מיון ספייק פוטנציאל פעולה בודד מסודרים וקודי צבע. היסטוגרמה מרווח כיסוי של היחידות ממוינות ממחיש את ההפרדה של צורות הגל. ד) ספייק מציינת את איכות הפרדת היחידות ממוינות כהוכחה למיון ספייק הולם. שים לב שאין ספייק בתוך התקופה עקשן של היחידה. ה) שתי תצוגות (E1, E2) מאשכולות 3D של יחידת המיון עם ניתוח מרכיבים ראשי המציין את המרחק של היחידות ממוינות אחד לשני. עיגולים מציינים את מרחק מלנוביס פי 2.5 אשר דומה SD בחלל ומציין בידול משמעותי של האשכולות בF מרחב המרכיב העיקרי יחידות) צבע מקודד מתארות את פוטנציאל הפעולה גלוי בהגדלה של שלושה ערוצים מהקלטת מערכת אחת. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}

איור 4. הדמיה לאחר הקלטה ו3D-שחזור של עמדת ההקלטה. א) השקפת הקרנה של Ortho-פרוסות לאורך Z-הצירים עם יישור עוצמה מקסימלית של Anteroולמלא את המשבצות מדרדר של נוירונים הקרנת uniglomerular מקרינים מAL לMB ו-LH. נותב תאיים Microruby (dextran tetramethylrhodamin) היה מוכנס לתוך AL לאחר ניסויי ההקלטה. glomeruli המוכתם בתוך תצוגת הקרנת AL מכתים הוכחה נכונה PN. ב) לפרוסות Ortho לאורך Z-הצירים עם יישור עוצמה מקסימלית מהכתמה של שתי אלקטרודות עם אלקסה hydrazide 488 מציין את מיקום האלקטרודה למ-ALT (E1, החץ ) ו-L-ALT (E2, חץ). נותב Alexa Hydrazide 488 נודד לרקמות המקיפות את האלקטרודה ומכתים את אתר החדרת אלקטרודה. שים לב, הצביעה הבולטת בAL היא artefact שטחי. C) 3D שחזורים של התאים המוכתמים היעד (PNS) ואתר הכנסת האלקטרודה מA, B (בצד ימין) יחד עם סקירה סכמטית של מערכת חוש הריח דבורת הדבש (משמאל צד) עם הציון של מסלולי l-ו-M-ALT. שים לב, רק uniglomerשטחי התואר בלבד PN מוצגים. : עצב מחושים, אל על: אונת מחושים, LH: קרן לרוחב, MB: גוף פטרייה, E1, E2: אתרי החדרה אלקטרודה, מ-ALT: מערכת אונת מחושים המדיאלי, L-ALT: מערכת אונת מחושים לרוחב, ג: הזנב, r: מקורי, מ ': המדיאלי, l:. לרוחב אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}

איור 5. דוגמא לקידוד ריכוז ריח במסלול הריח הכפול.) חלקות חום להמחיש את התגובה של l-ALT בודד (ירוקה) ו-M-ALT אחד PN (סגול) רכש בו זמנית מדבורת דבש בודדת בתגובה לodorant hexanal בהגדלת ריכוזי ריח (מ 1: 10 -6 ל1:100). כל שורה היא ממוצע של עשרה סטים משפטulation. שיעור הירי מוצג כשינוי עוצמה יחסי המהווה את קצב הירי ביחס לפעילות הספונטנית מופחת. ב ') חביון תגובת אוכלוסיית 11 l-ו13 מ-ALT PNS מ7 דבורים רשמו. בכל אחד דבורת PNS משני חלקות נרשמו בו זמנית. חביון תגובת אוכלוסיית תערוכות השהיה יורדת עם הגדלת ריכוז ריח בPNS משני קטעים. תחילת תגובת ריח במחושים ב99 אלפיות שניים נרשמה באמצעות electroantennograms ונגרעה משיהוי תגובת PN. שים לב כי בריכוזים הנמוכים ביותר תגובות חלשות מדי למדידות השהיה, ולכן לא נכללו בתגובת אוכלוסייה. C) ירי שיעור מאותה PNS כמו בב 'עם ריכוז ריח הגדלת כוח התגובה הולך וגדל. L-ALT מציג חוזק תגובה חזק יותר. בB, ניתנים C הממוצע ו-SD.

איור 6. השוואת פעילות אוכלוסייה בשני שלבי עיבוד שלאחר מכן לאורך מסלול חוש הריח של דבורת הדבש. נתונים נרשמו ב20 בעלי חיים אשר היו מגורה עם 1-hexanol ו2-octanone.) כל שורה מייצגת צבע השווא מקודד קצב ירי ממוצע של היטל אחד נוירון (PN) מחושב על פני 10 חזרות ריח של 1-hexanol. מצגת ריח מתחילה בזמן 0 ונמשכה שלוש שניות. ב ') מציג את אותו הדבר כמו ב) אבל לנוירונים חיצוניים גוף פטרייה (EN). המטריצה שמוצגת ב) ניתן לראות כוקטור אוכלוסיית PN במהלך גירוי ריח עם 1-hexanol. חישבנו את אותו סוג של וקטור אוכלוסייה במהלך גירוי ריח עם 2-octanone והשתמשנו בשני הווקטורים בניתוח מרכיבים עיקריים (PCA) שמירה הזמני ממד. C) שלושת המרכיבים העיקריים הראשונים (PC1, 2 ו -3) היוזמם נגד אחד את השני כדי להמחיש את הפרדת הריח בפעילות הרכב PN במוצא אונת המחושים. הזמן לפני הופעת ריח מסומן בשחור. פעילות במהלך שלוש שניות של גירוי עם 1-hexanol מוצגת בכחול. הפעילות במהלך גירוי עם 2-octanol מוצגת באדום. יתר על כן, אנו מראים 1.5 שניות (הודעה) של הפעילות לאחר הריח של סט של 1-hexanol (כחול אור) ו -2 octanonen (ורוד). שים לב כי ברמת הרכב PN, שני הריחות מעוררי מסלולים שונים מאוד להתיישב ב" נקודה קבועה ", שמאריכה ימים אחרי כל תקופת גירוי ריח. רק אחרי הריח לקזז את המסלולים לחזור לפעילות הבסיסית ללא גירוי ריח. ד ') אותו הניתוח נעשה ברמת הרכב EN המייצגת את הפעילות במוצא גוף פטרייה. בהשוואה לפעילות PN ריחות לעורר מסלול פחות מובחן. יתר על כן "נקודה קבועה" אינה נצפות. בתחילה המסלולים מושרה ריח Intermingle עם פעילות בסיסית למרות שהריח הוא עדיין קיים. רק הריח לקזז עורר מסלול נוסף.
. סרט 1 זמן נפתר הערכה של מסלול ריח מושרה לאחר ניתוח מרכיבים ראשי של וקטור PN-אוכלוסייה (משמאל) ווקטור EN אוכלוסייה. (מימין;. Cp איור 6) החלקים העליונים כוללים שלושה מרכיבים העיקריים הראשונים (PC1, 2 ו -3) זממו אחד נגד השני. הלוחות הנמוכים להמחיש את ההערכה של PC1, 2 ו -3 לאורך זמן. גירוי ריח מסומן בפס האפור. כל הלוחות היו מסונכרנים. שים לב שפעילות אוכלוסיית EN מתחילה מעט לפני פעילות אוכלוסיית PN, תופעה אשר נראתה כי contraintuitive אבל יכולה להיות מוסברת על ידי הקישוריות ומאפיינים של השכבות מעורבות, אשר דנו קודם לכן 1.
Discussion
מאמר זה מדגים את הייצור ושימוש בחוט אלקטרודות מיקרו רב ערוצים מותאמים אישית שתוכננה. אלקטרודות שתוארו מתאימות להקלטה שניהם יחידה אחת ופעילות אוכלוסייה אשר היא שימושי במיוחד עבור מדידות חביון ומאפייני תגובה זמניות אחרים של נוירונים שונים וneuropils שונה בתוך דגימה אחת (לפרטים ראו 1,2,25). בנוסף יש לנו לראות כיצד ליישם באופן קבוע את האלקטרודות חוט מיקרו כדי לאפשר הקלטות יציבים לטווח ארוך במתנהגים דבורי דבש להימשך שעות עד ימים.
הקלטות רבת יחידה תאית הפכו לכלי נוח כדי להשיג רזולוציה גבוהה זמנית בשילוב עם מידע מרחבי. במקרה שלנו, אלה הם שני קטעים מקבילים עצביים 2 או שתיים neuropils שונה 1. ניתן להקליט נוירונים מרובים ונותחו ברמת תא העצב הבודד במקביל וברזולוציה גבוהה זמנית. שיא Multi-יחידהings יושמו לראשונה ביונקי 38 ומאוחר יותר גם בחרקים 39-41. התקדמות משמעותית הושגה עם הפיתוח והשיפור של טכניקות הקלטת רב ערוצית תאיים 42,43. זה, למשל, כולל את הפיתוח של אלקטרודות חדשות 44 או אלגוריתמי מיון וקיבוץ באשכול של ספייק רומן 45. שיטות כלליות של טכניקות הקלטה רבת יחידה תאית מתוארות היטב 46-48. אלקטרודות העצמית שנבנה שמוצגים בסרטון הזה בנוסף ניתן להתאים על ידי הוספה יותר microwires לאלקטרודה או יכולים להיות מעוות חוטי מיקרו כדי לזכות במרחקים קבועים למדידה בין הטיפים. שני ההליכים היו, עם זאת, להביא לירידה בגמישות והגדלת עובי של האלקטרודה.
בהשוואה לבדיקות סיליקון המשמשות בדרך כלל להקלטות תאי בחרקים הרבה יותר גדולים כמו עש נץ, ארבה והמקק 40,49 - 51 חוט אלקטרודות מיקרו תיארה הם קטנים יותר, גמישות ויכולה להתמודד בקלות עם תנועות מוח פוטנציאליות, ולכן ניתן להשתמש בו באופן מהימן בחרקים חברתיים קטנים כמו דבורים ונמלים המציגים רפרטואר התנהגותי רחב הרבה יותר. רוב בדיקות סיליקון שוק חד כמו מבני חיתוך האקסונים ורקמה עצבית לאורך ערוץ הכנסתם, ואילו חוטי מיקרו תיארו הם עגולים, גמישים וקטנים יותר ולכן הם פחות מזיקים לרקמה הסובבת וזה יתרון ברור אם המטרה היא ללמוד עוד פלסטיות טווח בבעלי חיים בשלמותה ומתנהגים. יתרון נוסף של אלקטרודות חוט מיקרו הוא הייצור שלהם בעלות הנמוכה והטיפול קל. במקום לנקות בדיקה סיליקון יקרה בזהירות את חוטי האלקטרודה הם טריים חתוכים לקראת כניסה המוח, ולכן בעיות חוסר הגודש. כמו כן ניתן להשתמש באלקטרודה חוט מיקרו יותר מפעם אחת באותו ההכנה הוכנסה גם בneuropiles שונה 1 oשטחי r 2 כפי שאנו מראים כאן. גישה זו היא חיובית במיוחד כדי לנתח ולהשוות היבטים זמניים כמו שיהוי תגובה ואינטראקציות ברמות עיבוד עצביים שונות.
אנו מודעים לעובדה שאות נרשמה תאית אינה משקפת את פעילות תא בודדת כשלעצמו. זה תמיד תרכובת של פעילות מתח סביב הקצה האלקטרודה. כדי לאתר את מקור אות ההבדל של שני ערוצי חוט מייקר שכנים בתוך אלקטרודה אחת תמיד מחושב. כך המקור לאותות ספייק משמשים כדי לחלץ את פעילות יחידה אחת תמיד היה קרוב מאוד לאחד או ערוץ האלקטרודה האחר וכתוצאה מכך גל ספייק להבחין בקלות. אותות ממקום מרוחק יותר, כמו פעילות שרירים או פעילות של neuropils השכן, יגיעו שני אלקטרודות באותו הזמן מעורר צורות ואמפליטודות דומות וייפסלו על ידי הליך זה. באמצעות טכניקת התאמת תבנית של Spike2,אנחנו בטוחים מאוד כדי להשיג פעילות יחידה אחת, שזה לא אותו הדבר, אבל קרובים מאוד לפעילות תא עצב יחידה. עם זאת, סוגיית מיון ספייק יכולה להימנע על ידי שימוש בטכניקות הקלטה תאית.
הקלטות תא בודדות עם שני אלקטרודות חדות או טפטפות תיקון תאפשר ידע מעמיק על מאפיינים פיסיולוגיים של נוירון בודד. עם זאת, בשל גודלו הקטן של נוירונים חרקים וneurites (לדוגמא., מיקרומטר פחות מ 1 לדבורת הדבש PNS 52) הקלטות קצר טווח הן לניהול. יתר על כן, הקלטות סלולריות התוך עשויות להיות פולשנית ועלולה לפגוע בתא שבו הוא אולי סיבה נוספת למגבלות זמן. בvivo הקלטות תאית בחרקים נדירים לשרוד שעה אחת. חלון זמן שהיה מספיק לעבודתו החלוצית של מרטין האמר 53 שנרשמו תאיים מתא עצב אחד מזוהה, תא עצב maxilar # הגחון מזווג 1 (VUMmx1). הוא יכול lדיו את פעילותה באופן ישיר למסלול הגמול. יוליאנה Mauelshagen 54 רשום intracellularly הפעילות של נוירון החיצוני מזוהה פטריות גוף, pedunculus נוירון החיצוני # 1 (PE1) בהתניה קלאסית. אותו תא העצב היה במוקד של מנזל ומנז 55 כאשר הם מצאו LTP לאחר גירוי חשמלי של תאי קניון. עם זאת, אוקאדה ועמיתי 56 יכולים להשתמש בדפוס intracellularly המאופיין היטב spiking (קוצים כפולים ומשולשים) לזיהוי PE1 במהלך הקלטות תאיים. אחרי הכל שילוב של שני השיטות, הקלטות תאיים מתא עצב מזוהה והקלטות ארוכות טווח תאיים עשויות להיות כלי רב עוצמה לחקירות עתידיות.
עם זאת, באמצעות אלקטרודות חדות להקליט תאים מרובים (יחידות) בו זמנית ברמות עיבוד שונות על פני שעות רבות עד ימים כדי לנתח את מערכות היחסים שלהם זמני תגובה ו / או אפילו שינויי פלסטיק iזה כמעט בלתי אפשרי.
עם ההדמיה סידן הראשונה מתקרבת ב57,58 דבורת הדבש באמצעות צבעי סידן רגיש לניתוח דפוסים מרחביים של תגובות ריח היה נגיש 59-62. עם זאת, במקרים רבים הצבעים הרגישים סידן צריכה להיות הציגו לרקמת המוח באמצעות מניפולציות פולשנית ששוב להגביל את תוחלת החיים של הדבורה ותכונות פנימיות של התאים נותחו. בעיה זו היא להתגבר באורגניזמים מודל אחרים כמו פריזבוב באמצעות חיישני סידן הציגו גנטי 63,64. עם זאת, באופן כללי, חיישני סידן עשויים להציג את מגבלות אחרות כפי שהם יכולים לשמש כמאגרי סידן שצפויה להשפיע על המאפיינים זמניים של תגובות ריח. הקלטות תאיים בו זמנית בשילוב עם הדמיה סידן או גישות חישוביות יכולות להוכיח הפתרון הזמני הנכון של תהליכי הדמיה 65,66. עם זאת, ההחלטה הזמנית של תהליך ההדמיה עצמה היא Rather מוגבל. רכישת מערכות אופטיות בדרך כלל להשתמש CCD-הדמיה עם רזולוציה זמנית של 5-20 הרץ 67, אם כי 2-Photon-Imaging יוכל לרכוש רצפים מהירים יותר 68. עם זאת, קצב דגימת הגדלת תמיד הולך יחד עם הפסד ברזולוציה מרחבית. יתר על כן צבעי סידן רגיש לשימוש בדבורת הדבש לעבור הלבנת, אשר גם מפחיתה את זמן רכישה 69.
בהשוואה לטכניקות הקלטה פיסיולוגיות אחרות בחרקי אלקטרודות מיקרו החוט שלנו גמישה רב הערוצים להבטיח גישה ארוך זמן ליחידה אחת ופעילות עצבית באוכלוסייה מתנהג דבורים.
הראינו כיצד להשתמש בשני אלקטרודות אלה בשלבי עיבוד שונים באותה החיה, המאפשרת הניתוח של היבטי קידוד זמניים בין אתרי הקלטה השונים. תלוי בבעית המחקר ומודל חרקים השיטה הבסיסית של בניין האלקטרודה הפגין כאן הוא להרחיב בקלותמסוגל ו / או יכול להיות מותאם. לדוגמא ניתן לשער להשתמש יותר משלושה חוטים אחד לייצר אלקטרודות רב ערוצית. בנוסף, מספר אתרי הקלטה ניתן להאריך והתבוננות היבטים זמניים של יותר משני שטחים או neuropils אינה ריאלי. התקווה שלנו היא כי שיטה זו תשמש השראה למדענים רבים ותתרום באופן חיובי להבנה של עיבוד עצבי מתוחכם במוח קטן.
Disclosures
החוקרים מצהירים כי אין להם אינטרסים כלכליים מתחרים.
Acknowledgements
The authors would like to thank Isabelle Reus for establishment of tracing the electrode insertion side, Tobias Rosenbaum for LabView programming, Anneke Meyer for data analyzes and helpful discussions. We thank Randolf Menzel for discussion and practical help during early stage of electrode development. Furthermore we thank Brian Smith for postdoctoral association to MS-B. This work was supported by the Deutsche Forschungsgemeinschaft (DFG, SPP 1392, Ro1177/5-2) to WR.
Materials
| Name | Company | Catalog Number | Comments |
| Paraffin oil | Fluka | 76235 | |
| Odors | Sigma Aldrich | ||
| PBS | pH 7.2 | ||
| 4% Formaldehyde | ThermoScientific | 28908 | Methanol free |
| Triton X | BioChemica | A1388 | |
| Methylsalicylate | Roth | 4529.1 | |
| Tetramethylrhodamin dextran, 10,000 MW (Microruby) | Invitrogen | D7162 | keep dark |
| Alexa 488 hydrazide | Invitrogen | A-10436 | keep dark |
| Alexa 568 hydrazide | Invitrogen | A-10437 | keep dark |
| Bee Ringer Solution | see 2 | ||
| Polyurethane-coated copper wire | Elektrisola | 15µm diameter & P155 insulation | |
| Dental Wax | Densply Detrey | 64103015S1 | moderate melting point |
| Dental Wax | Flexaponal | 124-202-00 | low-melting Wax |
| KWIK SIl | WPI | 03L | |
| 18 Pin Socket | Conrad Electronic | 189634-62 | |
| Hot melting glue | Conrad Electronic | 827673 | |
| soldering needle | Conrad Electronics | 830283 | 12 V |
| Soldering terminal lug | Conrad Electronic | 531901 | |
| Glaselectrodes | WPI | 1B100F-3 | |
| Minutien Pins | Fine Science Tools | 26002-20 | V2A 0.2 x 12 mm |
| switchable headstage | Tucer Davis Technologies | SH16 | |
| Headstage connection module | NPI | INT-03M | |
| Amplifier Module | NPI | PDA-2F | |
| Data Acquisition boards | National Instruments | NI-6123, Ni-6143 | |
| Acquisition Software | National Instruments | Lab View 8.2 | custom design |
| Spike-Sorting | CED | Spike 2 v7.11 | |
| Matlab | Mathworks | R2008B | |
| Micromanipulator | Leitz | manual | |
| AG-wires | WPI | AGT05100 | |
| Confocal laser scanning microscope | Leica | TCS SP2 AOBS | |
| AMIRA | Mercury Computer Systems | 2/5/2000 |
References
- Strube-Bloss, M. F., Herrera-Valdez, M. a., Smith, B. H. Ensemble response in mushroom body output neurons of the honey bee outpaces spatiotemporal odor processing two synapses earlier in the antennal lobe. PLoS ONE. 7 (11), (2012).
- Brill, M. F., Rosenbaum, T., Reus, I., Kleineidam, C. J., Nawrot, M. P., Rössler, W. Parallel processing via a dual olfactory pathway in the honeybee. The Journal of neuroscience the official journal of the Society for Neuroscience. 33 (6), 2443-2456 (2013).
- Menzel, R. The honeybee as a model for understanding the basis of cognition. Nature Reviews Neuroscience. 13 (11), 758-768 (2012).
- Sandoz, J. Behavioral and neurophysiological study of olfactory perception and learning in honeybees. Frontiers in Systems Neuroscience. 5, (2011).
- Giurfa, M. Cognition with few neurons: higher-order learning in insects. Trends in Neurosciences. 36 (5), 1-10 (2013).
- Menzel, R., Giurfa, M. Cognitive architecture of a mini-brain: the honeybee. Trends in cognitive sciences. 5 (2), 62-71 (2001).
- Hildebrand, J. G., Shepherd, G. M. Mechanisms of olfactory discrimination: converging evidence for common principles across phyla. Annual Review of Neuroscience. 20, 595-631 (1997).
- Wilson, R. I., Mainen, Z. F. Early events in olfactory processing. Annu. Rev. Neurosci. 29, 163-201 (2006).
- Streinzer, M., Kelber, C., Pfabigan, S., Kleineidam, C. J., Spaethe, J. Sexual dimorphism in the olfactory system of a solitary and a eusocial bee species. The Journal of comparative neurology. 521 (12), (2013).
- Nishino, H., Nishikawa, M., Mizunami, M., Yokohari, F. Functional and topographic segregation of glomeruli revealed by local staining of antennal sensory neurons in the honeybee Apis mellifera. The Journal of comparative neurology. 515 (2), 161-180 (2009).
- Schneider, D. Elektrophysiologische Untersuchungen von Chemo- und Mechanorezeptoren der Antenne des Seidenspinners Bombyx mori L. Zeitschrift für Vergleichende Physiologie. 40 (1), 8-41 (1957).
- Menzel, R., Rybak, J. Antennal lobe of the honeybee. Handbook of brain microcircuits. , 427-432 (2011).
- Girardin, C. C., Kreissl, S., Galizia, C. G. Inhibitory connections in the honeybee antennal lobe are spatially patchy. Journal of neurophysiology. 109 (2), 332-343 (2013).
- Meyer, A., Galizia, C. G. Elemental and configural olfactory coding by antennal lobe neurons of the honeybee (Apis mellifera). Journal of comparative physiology. A, Neuroethology, sensory, neural, and behavioral. 198 (2), 159-171 (2012).
- Abel, R., Rybak, J., Menzel, R. Structure and response patterns of olfactory interneurons in the honeybee, Apis mellifera. The Journal of comparative neurology. 437 (3), 363-383 (2001).
- Kirschner, S., Kleineidam, C. J., Zube, C., Rybak, J., Grünewald, B., Rössler, W. Dual olfactory pathway in the honeybee, Apis mellifera. The Journal of comparative neurology. 499 (6), 933-952 (2006).
- Galizia, C. G., Rössler, W. Parallel olfactory systems in insects: anatomy and function. Annual review of entomology. 55 (August), 399-420 .
- Ito, K., Shinomiya, K., et al. A coordinated nomenclature system for the insect brain. Neuron. 81 (4), 755-765 (2014).
- Rybak, J. The digital honey bee brain atlas. Honeybee Neurobiology and Behavior. , 125-140 (2012).
- Rössler, W., Brill, M. F. Parallel processing in the honeybee olfactory pathway: structure, function, and evolution. Journal of comparative physiology. A, Neuroethology, sensory, neural, and behavioral. 199 (11), (2013).
- Mobbs, P. The brain of the honeybee Apis mellifera. I. The connections and spatial organization of the mushroom bodies. Philosophical transactions of the Royal Society of London. Series B, Biological. 298 (1091), 309-354 (1982).
- Strausfeld, N. J. Organization of the honey bee mushroom body: representation of the calyx within the vertical and gamma lobes. The Journal of comparative neurology. 450 (1), 4-33 (2002).
- Witthöft, W. Absolute Anzahl und Verteilung der Zellen im Hirn der Honigbiene. Zeitschrift für Morphologie der Tiere. 61 (1), 160-184 (1967).
- Rybak, J., Menzel, R. Anatomy of the mushroom bodies in the honey bee brain: the neuronal connections of the alpha-lobe. The Journal of Comparative Neurology. 465, 444-465 (1993).
- Strube-Bloss, M. F., Nawrot, M. P., Menzel, R. Mushroom body output neurons encode odor-reward associations. The Journal of neuroscience : the official journal of the Society for Neuroscience. (8), 3129-3140 (2011).
- Haddad, R., Lanjuin, A., Madisen, L., Zeng, H., Murthy, V. N., Uchida, N. Olfactory cortical neurons read out a relative time code in the olfactory bulb. Nature neuroscience. (May), 1-11 (2013).
- Martin, J. P., Beyerlein, A., et al. The neurobiology of insect olfaction: Sensory processing in a comparative context. Progress in neurobiology. 95 (3), 427-447 (2011).
- Nawrot, M. P. Dynamics of sensory processing in the dual olfactory pathway of the honeybee. Apidologie. 43 (3), 269-291 (2012).
- Farkhooi, F., Froese, A., Muller, E., Menzel, R., Nawrot, M. P. Cellular adaptation facilitates sparse and reliable coding in sensory pathways. PLoS computational biology. 9 (10), e1003251 (2013).
- Saha, D., Leong, K., Katta, N., Raman, B. Multi-unit recording methods to characterize neural activity in the locust (Schistocerca americana) olfactory circuits. Journal of visualized experiments JoVE. (71), (2013).
- Saha, D., Leong, K., Li, C., Peterson, S., Siegel, G., Raman, B. A spatiotemporal coding mechanism for background-invariant odor recognition. Nature neuroscience. 16 (12), 1-13 (2013).
- Mizunami, M., Okada, R., Li, Y., Strausfeld, N. J. Mushroom Bodies of the Cockroach Activity and Identities of Neurons. Journal of Comparative Neurology. 519 (July), 501-519 (1998).
- Okada, R., Ikeda, J., Mizunami, M. Sensory responses and movement-related activities in extrinsic neurons of the cockroach mushroom bodies. Journal of Comparative Physiology A: Sensory, Neural, and Behavioral Physiology. 185 (2), 115-129 (1999).
- Haehnel, M., Froese, A., Menzel, R. In vivo Ca2+ imaging of mushroom body neurons during olfactory learning in the honey bee. Journal of visualized experiments JoVE. (30), (2009).
- Felsenberg, J., Gehring, K. B., Antemann, V., Eisenhardt, D. Behavioural pharmacology in classical conditioning of the proboscis extension response in honeybees (Apis mellifera). Journal of visualized experiments JoVE. (47), (2011).
- Rehder, V. Quantification of the honeybee’s proboscis reflex by electromyographic recordings. Journal of Insect Physiology. 33 (7), 501-507 (1987).
- Serrano, E., Nowotny, T., Levi, R., Smith, B. H., Huerta, R. Gain control network conditions in early sensory coding. PLoS computational biology. 9 (7), e1003133 (2013).
- Hubel, D. H., Wiesel, T. N. Receptive fields and functional architecture of monkey striate cortex. The Journal of physiology. 195 (1), 215-243 (1968).
- Christensen, T. A., Pawlowski, V. M., Lei, H., Hildebrand, J. G. Multi-unit recordings reveal context-dependent modulation of synchrony in odor-specific neural ensembles. Nature neuroscience. 3 (9), 927-931 (2000).
- Byers, K. J. R. P., Sanders, E., Riffell, J. A. Identification of olfactory volatiles using gas chromatography-multi-unit recordings (GCMR) in the insect antennal lobe. Journal of visualized experiments JoVE. (72), e4381 (2013).
- Perez-Orive, J., Mazor, O., Turner, G. C., Cassenaer, S., Wilson, R. I., Laurent, G. Oscillations and sparsening of odor representations in the mushroom body. Science. 297 (5580), 359-365 (2002).
- Stevenson, I. H., Kording, K. P. How advances in neural recording affect data analysis. Nature. 14 (2), 139-142 (2011).
- Buzsáki, G. Large-scale recording of neuronal ensembles. Nature neuroscience. 7 (5), 446-451 (2004).
- Viventi, J., Kim, D. -. H., et al. Flexible, foldable, actively multiplexed, high-density electrode array for mapping brain activity in vivo. Nature neuroscience. 14 (12), 1599-1605 (2011).
- Pouzat, C., Mazor, O., Laurent, G. Using noise signature to optimize spike-sorting and to assess neuronal classification quality. Journal of neuroscience methods. 122 (1), 43-57 (2002).
- Lewicki, M. S. A review of methods for spike sorting: the detection and classification of neural action potentials. Network. 9 (4), R53-R78 (1998).
- Quian Quiroga, R., Panzeri, S. Extracting information from neuronal populations: information theory and decoding approaches). Nature reviews. Neuroscience. 10 (3), 173-185 (2009).
- Einevoll, G. T., Franke, F., Hagen, E., Pouzat, C., Harris, K. D. Towards reliable spike-train recordings from thousands of neurons with multielectrodes. Current opinion in neurobiology. 22 (1), 11-17 (2012).
- Riffell, J. a., Lei, H., Abrell, L., Hildebrand, J. G. Neural basis of a pollinator’s buffet: olfactory specialization and learning in Manduca sexta. Science. 164 (6), 877-892 (2013).
- Bender, J. a., Pollack, A. J., Ritzmann, R. E. Neural activity in the central complex of the insect brain is linked to locomotor changes. Current biology : CB. 20 (10), 921-926 (2010).
- Perez-Orive, J., Bazhenov, M., Laurent, G. Intrinsic and circuit properties favor coincidence detection for decoding oscillatory input. The Journal of neuroscience the official journal of the Society for Neuroscience. 24 (26), 6037-6047 (2004).
- Rybak, J. . Die strukturelle Organisation der Pilzkörper und synaptische Konnektivität protocerebraler Interneuronen im Gehrin der Honigbiene, Apis mellifera.: eine licht- und elektronenmikroskopische Studie. , (1994).
- Hammer, M. An identified neuron mediates the unconditioned stimulus in associative olfactory learning in honeybees. Nature. 366, 59-63 (1993).
- Mauelshagen, J. Neural correlates of olfactory learning paradigms in an identified neuron in the honeybee brain. Journal of neurophysiology. 69, 609-625 (1993).
- Menzel, R., Manz, G. Neural plasticity of mushroom body-extrinsic neurons in the honeybee brain. The Journal of experimental biology. 208 (22), 4317-4332 (2005).
- Okada, R., Rybak, J., Manz, G., Menzel, R. Learning-related plasticity in PE1 and other mushroom body-extrinsic neurons in the honeybee brain). The Journal of neuroscience: the official journal of the Society for Neuroscience. 27 (43), 11736-11747 (2007).
- Joerges, J., Küttner, A., Galizia, C. G., Menzel, R. Representation of odours and odour mixtures visualized in the honeybee brain. Nature. 387, 285-288 (1997).
- Galizia, C. G., Joerges, J., Küttner, A., Faber, T., Menzel, R. A semi-in-vivo preparation for optical recording of the insect brain. Journal of neuroscience. 76 (1), 61-69 (1997).
- Galizia, C. G., Sachse, S., Rappert, A., Menzel, R. The glomerular code for odor representation is species specific in the honeybee Apis mellifera. Nature. 2 (5), 473-478 (1999).
- Sandoz, J. -. C. Odour-evoked responses to queen pheromone components and to plant odours using optical imaging in the antennal lobe of the honey bee drone Apis mellifera L. The Journal of experimental biology. 209 (18), 3587-3598 (2006).
- Fernandez, P. C., Locatelli, F. F., Person-Rennell, N., Deleo, G., Smith, B. H. Associative conditioning tunes transient dynamics of early olfactory processing. The Journal of neuroscience the official journal of the Society for Neuroscience. 29 (33), 10191-10202 (2009).
- Locatelli, F. F., Fernandez, P. C., et al. Nonassociative plasticity alters competitive interactions among mixture components in early olfactory processing. European Journal of Neuroscience. 37 (1), (2013).
- Silbering, A. F., Bell, R., Galizia, C. G., Benton, R. Calcium imaging of odor-evoked responses in the Drosophila antennal lobe. Journal of visualized experiments JoVE. (61), 1-7 (2012).
- Strutz, A., Völler, T., Riemensperger, T., Fiala, A., Sachse, S. Calcium imaging of neural activity in the olfactory system of Drosophila. Genetically Encoded Functional Indicators. 72, 43-70 (2012).
- Galizia, C. G., Kimmerle, B. Physiological and morphological characterization of honeybee olfactory neurons combining electrophysiology, calcium imaging and confocal microscopy. Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology. 190 (1), 21-38 (2004).
- Helmchen, F., Waters, J. Ca2+ imaging in the mammalian brain in vivo. European journal of pharmacology. 447 (2-3), 119-129 (2002).
- Stierle, J. S., Galizia, C. G., Szyszka, P. Millisecond stimulus onset-asynchrony enhances information about components in an odor mixture. Journal of Neuroscience. 33 (14), 6060-6069 (2013).
- Haase, A., Rigosi, E., et al. In-vivo two-photon imaging of the honey bee antennal lobe. Biomedical optics express. 2 (1), 131-138 (2010).
- Becker, P. L., Fay, F. S. Photobleaching of fura-2 and its effect on determination of calcium concentrations. The American journal of physiology. 253 (4 pt 1), C613-C618 (1987).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved