Method Article
Les enregistrements simultanés à long terme à deux étapes de traitement neuronal à se conduire abeilles
Dans cet article
Résumé
Enregistrements à long terme extracellulaires simultanée de deux neuropiles différentes du cerveau ou deux voies anatomiques différents ont été établis dans les abeilles. Ces enregistrements permettent l'enquête des aspects temporels de traitement neuronal dans différentes zones du cerveau à l'unique neurone ainsi que sur le plan d'ensemble dans un animal se comporter.
Résumé
Dans les deux mammifères et les insectes informations neuronale est traitée dans les différents centres d'ordre supérieur et inférieur cerveau. Ces centres sont couplés par convergentes et divergentes connexions anatomiques y compris l'alimentation et le câblage avant de rétroaction. En outre, les informations de la même origine est partiellement envoyé par des voies parallèles pour différentes et parfois dans les mêmes zones du cerveau. Pour comprendre les avantages évolutifs ainsi que les avantages de calcul de ces stratégies de câblage et surtout leurs dépendances temporelles sur l'autre, il est nécessaire d'avoir un accès simultané à neurones individuels de différents secteurs ou neuropiles dans la même préparation à haute résolution temporelle. Ici, nous nous concentrons sur les abeilles en démontrant un accès à long terme extracellulaire unique à enregistrer l'activité de l'unité multiple à deux neuropiles ultérieures 1, le lobe antennaire (AL), la première étape de traitement olfactif et le corps de champignon (MB), un ordre plus élevé centre d'intégration involved dans l'apprentissage et la formation de la mémoire, ou deux voies neuronales parallèles 2 reliant l'AL avec le MB. Ce dernier a été choisi à titre d'exemple et sera décrite dans son intégralité. Dans la vidéo soutenant la construction et de l'insertion permanente d'électrodes de fil de canal multi-flexibles est démontrée. Amplification différentielle par paire des micro-canaux d'électrode de fil permet de réduire considérablement le bruit et vérifie que la source du signal est étroitement liée à la position de la pointe de l'électrode. La flexibilité mécanique des fils-électrodes utilisées permet des enregistrements à long terme envahissantes stables pendant de nombreuses heures à jour, ce qui est un net avantage par rapport aux techniques classiques d'enregistrement in vivo supplémentaires et intracellulaires.
Introduction
Les abeilles ainsi que la plupart des autres insectes comptent beaucoup sur l'olfaction. Parmi d'autres, ils utilisent des indices olfactifs pour l'orientation, l'accouplement, la communication avec leurs congénères, et la recherche de nourriture. Leur système olfactif bien élaboré contribue à un riche répertoire de comportements d'apprentissage liés à des stimuli d'odeurs florales. Ces comportements peuvent être facilement étudiés dans des conditions contrôlées en laboratoire (pour revue voir 3 - 5). Leurs "mini cerveau" (cf. 6) avec leur nombre relativement restreint de neurones fait l'abeille un organisme modèle bien adapté à l'étude de codage olfactif et l'apprentissage lors de la surveillance de l'activité neuronale.
Le système olfactif des insectes ainsi que chez les mammifères montre organisme analogue dans une large mesure (pour revue, voir 7,8). Dans abeilles environ 80.000 neurones récepteurs situés dans 9 sensilles sur les antennes 10,11 traduisent la relance de l'odeur de l'environnement dans un neursignal de onal. Les axones des neurones récepteurs olfactifs innervent le lobe antennaire (AL), qui a une organisation comparable à glomérulaire du bulbe olfactif des vertébrés. L'AL comprend environ 164 glomérules interconnectés les uns aux autres par environ 4.000 interneurones locaux (LN) (pour revue, voir 12). Surtout chez l'abeille, il a été récemment montré que LN offrent une connectivité latérale inégale et que les différentes sous-populations possèdent des propriétés olfactives de codage élémentaires et configural 13,14. L'AL a été montré pour être subdivisé en un lobe ventral et un hémi dorsale donnant lieu à l'interne et les voies de lobe antennaire latérale (m-et l-ALT; anciennement appelé m-et l-APT pour médial et latéral antennaire lobe protocerebral tracter 15 - 17). Voici une nouvelle terminologie des voies introduite par un effort récent pour une nomenclature unifiée du cerveau de l'insecte sera utilisé 18. Les offres anormalement basses (l et m-ALT) combinent soit 410 (l-ALT) ou 510 (m-ALT) proje uniglomerularneurones ction (PN), respectivement 15,16,19. Unités de raccordement des deux voies ont été récemment démontré que les odeurs de code en parallèle 2 (pour revue, voir 17,20), et deux voies divergentes former des synapses connexions avec des cellules de Kenyon (KC), le corps de champignon (MB) neurones principaux. Chaque Mo contient environ 172 000 KC 21 - 23. Les MB sont connus pour être impliqués dans l'intégration de relance, l'apprentissage et la formation de la mémoire. Les dendrites axo de KCs forment le pédoncule (la tige du champignon), qui a deux principales régions de production: la vertica ou alpha-lobe et l'22,24 horizontale ou bêta-lobe. La sortie de la MB converge à seulement environ 400 neurones extrinsèques (FR) 24. ENS responsable de traitement de l'information olfactive surtout innervent la face ventrale du lobe vertical 22. Récemment, il a été montré que les EN enregistrées à cette zone codent pour l'association odeur de récompense 25.
Temporelle commeaspects du système olfactif des insectes ainsi que les vertébrés sont devenus un aspect important et significatif en tant que principe potentiel de codage 26-29. Pour être en mesure d'enregistrer simultanément plusieurs neurones de différents sites à haute résolution temporelle, nous avons établi doubles techniques d'enregistrement de l'unité multi-électrodes à l'aide de fil multicanal personnalisés introduites à différentes régions cibles dans le système olfactif de l'abeille. Cette approche nous permet d'analyser et de comparer le traitement temporel dans le système olfactif des abeilles au niveau des neurones et des populations de neurones, soit entre voies olfactives parallèles, la double voie olfactive 2 ou entre les différents neuropils ultérieures 1 simple. Récemment, avec une approche expérimentale similaire dans le système olfactif acridienne 30 en utilisant une configuration différente des électrodes étaient en mesure d'analyser le mécanisme de codage spatio-temporel pour la reconnaissance des odeurs fond invariant 31. Thnous, les deux enregistrements établis permettent de recueillir de l'information spatiale sur les profils d'activité neuronales simultanées.
Par rapport à l'échantillonnage spatial plus large obtenu à partir de l'imagerie calcique cette méthode permet l'enregistrement de deux points seulement. Cependant, l'avantage par rapport aux techniques d'imagerie de calcium est la précision de la mesure temporelle des enregistrements potentiels qui ne peuvent pas être fournis soit par l'imagerie CCD classique ou acquisition d'imagerie à 2 photons. Les électrodes décrites ici extracellulaires sont implantés de manière permanente et fixe par rapport au cerveau et de la capsule de la tête en évitant la dérive de l'électrode. Il s'agit d'un net avantage par rapport à l'utilisation d'électrodes intracellulaires pointus. Un autre avantage par rapport aux enregistrements intracellulaires et l'imagerie du calcium est le temps d'observation de neurones étendue allant de plusieurs heures à jour. Il s'agit d'une condition préalable importante pour étudier les corrélats neuronaux de l'apprentissage et de formation de la mémoire. Les autres avantages de plusieursenregistrements unitaires sont en outre décrits dans la section discussion.
Dans cet aperçu méthodologique de la procédure de fabrication de fils-électrodes de conception personnalisée sera affiché, adapté de 32,33 et adapté pour les enregistrements à logements multiples à long terme dans le cerveau des abeilles. En outre, par exemple, la façon dont ces types d'électrodes sont implantées de façon permanente dans deux sites d'enregistrement différents dans le système olfactif d'abeilles pour enregistrer simultanément le l-et le m-ALT sur de longues périodes de temps pour permettre à de nombreux protocoles de stimulation est disponible en deux. Pour la vérification des positions d'enregistrement, par exemple, et le protocole pour la coloration et l'enregistrement de poste visualisation des sites d'enregistrement est fourni.
Protocole
1. Électrode du bâtiment (Figure 1)
- Production d'un adaptateur d'électrode qui correspond à l'interface électrode conseil d'commerciale à plusieurs 1,2,25 des systèmes d'amplification du canal.
- Utilisez une petite plaque de plexiglas collé à une base de connecteur 18 broches.
- Raccorder la base avec 3 petits morceaux de fil isolé à 3 pattes à souder séparées vissées sur la plaque de plexiglas (figure 1 A1-A3).
- Insérez une rainure dans la plaque de plexiglas dans lequel un capillaire de verre peut facilement se déplacer et être maintenu en place par une vis (Figure 1 B1).
- Elargir le capillaire en verre d'environ 5 mm à l'aide d'une broche de minuties.
- Attacher les fils d'électrode le long de la micro-tige de minuties et le capillaire de verre à garantir la stabilisation et le soutien.
- Production micro fil multi-canal (adoptée de Ryuichy Okada 32,33)
- Span 3 micro fils (fil de cuivre revêtu de polyuréthane, 15diamètre um) de manière qu'ils soient placés à côté de l'autre (figure 1 B2).
- Utiliser une aiguille 12 V à souder pour étaler une mince pellicule de cire dentaire à faible température de fusion (50 ° C) en partie le long des fils de les coller ensemble (pointe d'électrode) (Figure 1 B3). Laissez quelques centimètres unglued (fin de l'électrode) que cette section sera utilisé plus tard pour connecter les fils micro avec l'adaptateur d'électrode.
- Connectez le canal micro fil multiples à l'adaptateur d'électrode
- Retirez le capillaire en verre de la porte et l'attacher à la tige de minuties de la pointe d'électrode. L'amener dans une position parallèle à la micro-électrode (figure 1 B3).
- Collez le bout de l'électrode à la broche de minuties en utilisant bas point de fusion de la cire dentaire et couper le micro électrode à l'extrémité, dépassant de 2-3 cm de l'axe de minuties et à la fin de l'électrode (petites flèches Figure 1 B3).
- Glissez le sl capillaireightly retour dans l'adaptateur de l'électrode. Utilisez la vis pour le fixer (Figure 1 B4).
- Souder les extrémités libres des fils aux trois cosses à souder à l'aide d'un pistolet de soudage avec une température d'environ 360 ° C pour assurer une fusion de l'isolant (figure 1 B4). Après la soudure, s'assurer qu'il ya un contact électrique adéquat (~ 300 kOhm).
- Montez l'une des électrodes (maître) à l'interface électrode bord de la headstage et fixer l'autre électrode multi-canal (esclave) sur un adaptateur séparé. Branchez les chaînes de l'esclave à l'électrode principale (Figure 1 C1). En outre, la référence souder ainsi que des électrodes de muscles à la base de l'électrode principale (figure 1 C2).
2. Bee Préparation (Figure 2)
Dans ces expériences décrites, l'abeille (Apis mellifera), qui est un animal invertébréet ne nécessite donc pas de permis éthiques spécifiques pour l'utilisation, est utilisé.
- Catch butineuses abeilles (A. mellifera) à l'entrée de la ruche dans la matinée, comme indiqué par les autres 34,35.
- Refroidir les abeilles sur de la glace pilée jusqu'à immobilisation (5 à 10 min) et fixer un dans un support en plexiglas standard ou tube métallique d'une manière que la tête est exposée (Figure 2). Pour minimiser les mouvements de la tête de cire bas point de fusion dentaire (~ 50 ° C) et fixer la tête attentivement le support autour de la base des yeux composés et le cou.
- Utilisez cire à bas point de fusion pour fixer la SCAPI des antennes sur la tête capsule (figure 2 B) sans toucher le flagelle. Le flagelle de l'antenne doit être fait avant. Assurez-vous que l'abeille peut se déplacer librement sa trompe.
- Raser la capsule de la tête pour assurer une vue paisible et l'accès au sommet de la tête.
- Nourrir les abeilles avec une solution de saccharose à 30% jusqu'à saturatià assurer une humidification suffisante de tissu cérébral et une bonne viabilité de l'animal (figure 2 C).
- Faire des incisions minutieuses verticalement le long des frontières des yeux composés et horizontalement au-dessus des bases des antennes ainsi que sous les ocelles et retirer la pièce détachée de la cuticule (figure 2 D).
- Mettez soigneusement de côté hypopharyngiennes et enlever la trachée afin d'assurer une vision claire et l'accès au cerveau avant électrode insertion (figures 2E, 2F).
3. Electrode insertion
Dans le cas de l'exemple illustré à la figure 2, une électrode est positionnée pour but la l-ALT, l'autre en vue de la m-ALT 2. Utilisation de sites particuliers, d'autres régions cibles sont aussi possibles, par exemple les régions de sortie AL et MB 1.
- Placer les électrodes en utilisant micromanipulateurs à la région d'intérêt ( Figure 2G et 3A). Pour cibler l'endroit m-ALT l'électrode entre l'AL et en dedans du lobe vertical de la MB. S'assurer que le site d'insertion étant au-dessus du point de les unités de raccordement d'ALT médio-latérale de branchement. Faire saillie de l'électrode dans le cerveau avec une profondeur d'environ 180 um (figure 2E). Pour l-ALT PN enregistrement lieu l'électrode en dessous de la protocérébron latérale (la LH) dans le milieu d'une ligne imaginaire entre le côté latéral du lobe vertical et au milieu de la AL. Insérer l'électrode à une profondeur d'environ 300 um (figure 2E)
- Insérez la référence (fil d'argent, environ 25 um de diamètre) dans l'œil composé ipsilatéral par une petite incision dans la cuticule. Insérer un autre fil d'argent dans la région de la saillie au-dessous du muscle ocelles latérale. REMARQUE: Si nécessaire, le comportement d'apprentissage de l'abeille peut être contrôlée avec une grande précision temporelle en enregistrant le muscle M17, qui est impliqué in la réponse d'extension du proboscis (PER) de l'abeille 36 comme décrit dans 25.
- Pour ancrer solidement les électrodes dans le cerveau et la capsule de la tête, couvrir tout l'espace au-dessus du cerveau avec deux silicium composant (figure 2H), ce qui empêche le cerveau de se dessécher. REMARQUE: les enregistrements peuvent durer des heures jusqu'à jours, et les abeilles peuvent par exemple être enregistrés au cours d'une procédure de conditionnement classique (figure 2H) ou stimulées avec un large panel de différentes odeurs.
4. Acquisition et prétraitement des données
- Utilisez le logiciel d'acquisition appropriée qui répond aux exigences suivantes: taux d'échantillonnage de 25 kHz min; 1,25 analogique ou numérique de la différenciation croix entre deux canaux d'électrode; Filtre passe-bande de 300 Hz à 8000 Hz pour extraire les événements de pointes.
- Utilisez disponibles logiciel de pointe de tri pour extraire l'activité d'une seule unité, par exemple templmangé techniques qui correspond aussi inclus dans le logiciel Spike2 (Figure 3).
- Pour une analyse plus utiliser l'horodatage des unités extraites de calculer une seule unité moyenne des réponses des odeurs (figure 4) ou pour calculer des vecteurs de la population pour l'analyse en composantes principales (ACP) (Figure 5) en utilisant un logiciel disponible dans le commerce. Pour analyser plus en une seule unité et de la population la latence de réponse s'il vous plaît comparer publications récentes 1,2,25.
5. Visualisation des électrodes Position relative (Figure 4)
- Plonger les électrodes de conseil dans une solution de 5% soit 568 ou Alexa hydrazide 5% hydrazide Alexa 488, qui est dissoute dans une solution de chlorure de potassium 0,5 M avant les expériences d'enregistrement.
- Retirez les électrodes et le silicium de couverture soigneusement après les expériences, rincer le cerveau avec une solution de Ringer abeille, retirez glandes et de la trachée et insérer de minuscules cristaux de tetramethylrhodamin dextran ou insérez une solution à 5% résolu à 1,0 M d'acétate de potassium dans le AL pour étiqueter les offres anormalement basses antérograde. Effectuez les étapes suivantes dans l'obscurité.
- Laisser le colorant à prendre et transporté par les neurones de projection le long de leurs voies axonales (30-45 min) (figure 4A), avant de laver le cerveau avec une solution de Ringer abeille trois fois pour un autre 30-45 min.
- Refroidissez l'abeille sur la glace jusqu'à l'immobilisation et retirer délicatement le cerveau de la capsule de la tête. Fixer le cerveau d'un rinçage dans une solution 0,1 M de PBS contenant 4% de formaldehyde et le maintenir pendant une nuit à 4 ° C.
- Attendre au moins 12 heures avant de laver le cerveau deux fois en PBS 0,1 M (10 min chacun).
- Laver le 3x cérébral pendant 20 minutes dans 0,2% de Triton X-100 dilué dans 0,1 M de PBS avant la déshydratation dans une série d'alcool ascendant (30%, 50%, 70%, 90%, 95%, 3 x 100% d'éthanol, 20 min chaque étape).
- Incluez le cerveau déshydraté en salicylate de méthyle sur une lame et de microscopeeal avec une lamelle.
- Utiliser un microscope confocal à balayage laser microscope et scanner le cerveau comme des sections optiques chaque 2-5 um en utilisant un composé harmonique objectif plan apochromatiques (10X 0,4 NA d'immersion). Exciter le tissu à l'aide de 568 nm de longueur d'onde pour tetramethylrhodamin dextrane et une longueur d'onde de 488 nm pour la position de l'électrode.
- Reconstruire les structures cérébrales colorées et les chemins d'électrodes des piles d'images en 3D avec le logiciel de reconstruction (par exemple. AMIRA ou Fidji) (figure 4).
Résultats
"Le présent protocole permet des enregistrements simultanés à deux étapes différentes de traitement dans les abeilles individuelles et permet en outre de tester les mécanismes sous-jacents de l'apprentissage et de la mémoire par exemple., PAR conditionné dans les abeilles sobres." Il s'agit d'une condition préalable à l'analyse des aspects temporels de traitement neuronal. La méthode est facilement adaptable pour différentes approches scientifiques de démêler le réseau neuronal de système olfactif de l'abeille. Par exemple, cette méthode est utilisée: (i) analyser le traitement temporel des unités de raccordement à l'intérieur de la voie olfactive double de l'abeille, la l-m-ALT et les unités de raccordement (figure 5). Sur la figure 5A un exemple d'un PN l-ALT simultanément enregistrée avec une unité de raccordement de la m-ALT est donnée à dix moyen d'essai, et illustre leur résistance à la réaction et temps de latence par rapport à cinq concentrations d'odeurs différentes comme un code de couleur parcelle de chaleur. Dans une moyenne de sept abeilles avec 11 l et 13 mALT unités de raccordement (figures 5B, 5C) qui illustre à la fois la force de la réponse ainsi que la latence de la réponse, dans une certaine mesure, reflète la concentration de substance odorante. Ainsi les unités de raccordement dans cet exemple augmentent leur force de la réponse tout avec une concentration croissante odorant leur latence de réponse a diminué (figures 5B, 5C). Ce résultat est plutôt limitée et valable uniquement pour l'odorant analysé, mais il est encore conforme aux récentes des modèles de calcul de l'AL 37. Si un codage de concentration d'odeur en AL de l'abeille à la base de calculs non linéaires ou à la base d'autres propriétés de codage doit encore être analysé à l'avenir. En outre, le procédé peut être utilisé (ii) de comparer les aspects temporels de l'activité de la population à deux étapes de traitement ultérieures, le AL-MB-et sortie (figure 6). Analyse en composantes principales (ACP) illustre cette odeur calcul est prolongée et survivre à l'ensemble de la présentation de l'odeur au niveau PN alors que dans l'ENs que l'odeur et hors jeu étaient représentés dans l'activité de la population (figure 6). Ainsi la population FR atteint leur activité maximale déjà à un point dans le temps lorsque l'activité PN est encore en développement (cf. le film 1).

.. Figure 1 La fabrication des trois voies électrodes micro-filaires A1) Les terminaisons de trois fils sont soudés à un connecteur à broches IC en positions 11, 13 et 16 A2) Quatre cosses à souder sont vissées sur une plaque de base en matière plastique plexiglas.; une broche minuties est inséré dans l'extrémité d'un capillaire de verre qui est ensuite fixé à la plaque en plastique. A3) La plaque en matière plastique est collé sur le dessus d'un connecteur à broches IC et l'extrémité libre de chaque fil est soudée à une de la soudure supérieure pattes. B1) Pour équiper le porte-électrode avec les fins fils de cuivre, le capillaire doit être pris une fois de plus. B2) La base du support est alors fixé dans un rendu dispositif d'alignement. Trois cuivre micro fils sont alignés le long de la rainure et fixés avec du ruban adhésif à chaque extrémité B3) Les micro fils parallèles sont collées ensemble avec de la cire dentaire et le capillaire est mis en place (longues flèches).; les fils micro collés sont ensuite attachés à capillaire avec de la cire dentaire et ses extrémités sont coupées (petites flèches). B4) Les trois bouts de fils de cuivre sont soudés à trois pattes supérieures à souder les amenant ainsi en contact électrique avec la Connecteur à broche IC. C1) Deux électrodes entièrement assemblés peuvent être reliés les uns aux autres pour une utilisation avec une seule headstage. C2) A cet effet, les repères de l'une électrode (gauche, esclave) sont reliés par l'intermédiaire de fils isolés à l'autre électrode (à droite, mâter), qui est relié à l'étage de tête; la headstage connecté électrode peut également recueillir les commentaires d'un muscle (M17) et l'électrode de référence (Ref).

Figure 2. Préparation et insertion d'électrode permanente dans le cerveau d'abeille. A) Une abeille est inséré dans un support en plexiglas après immobilisation sur de la glace. Antennes avec flagelle (FL) et scapus (SC) sont indiqués. B) La tête et les antennes sont fixées avec de la cire dentaire. C) La capsule de la tête est rasée et l'abeille est alimenté avec de l'eau sucrée. D) La capsule de la tête est ouvert. E) Après avoir enlevé les glandes et la trachée de la partie supérieure du cerveau, les différents neuropiles et les principaux points de repère peuvent être facilement distingués. Les trajectoires des offres anormalement basses sont indiqués avecune marque où les électrodes sont insérées. (MB: corps de champignon, AL: lobe antennaire, OL: lobe optique, α: Alpha lobe ou lobe vertical, ALT: antennaire voies de lobe, E1: côté d'insertion d'électrode pour les enregistrements m-ALT, E2: côté d'insertion d'électrode pour l-ALT des enregistrements). F) de référence (Ref) et des électrodes de muscle (M17) sont insérés dans la capsule de la tête à travers de petits trous dans la cuticule ou l'œil composé. G) Les fils-électrodes sont insérées dans le cerveau au niveau des sites appropriés. H) Après fixant les électrodes en place à l'aide de deux composants silicium, l'abeille montre encore PER et peut être conditionné (par exemple., en utilisant de l'eau sucrée).

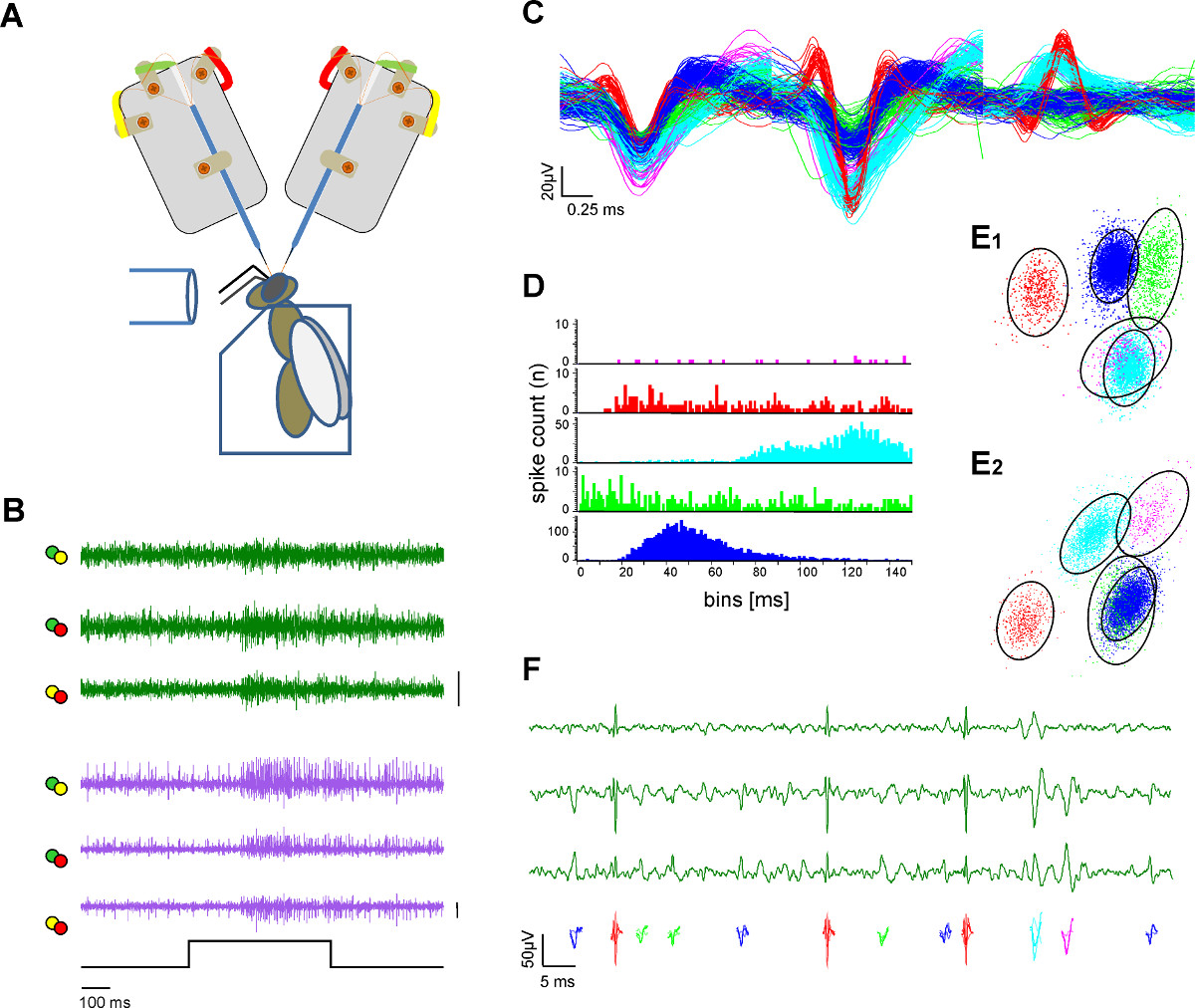
Figure 3. Extracellulaire enregistrement à deux voies neurales et l'extraction d'une seule unité (pic de tri). A ) Schéma du dispositif expérimental. L'abeille est obsédé dans un support en plexiglas. stimulation de l'odeur est fourni par l'intermédiaire d'un tube de verre. Deux tiges d'électrodes enregistrent du cerveau de l'abeille exposée. B) des enregistrements simultanés de la l-et m-ALT unités de raccordement (vert et traces violettes) montrant réponses excitateurs sur les deux voies à une stimulation de 500 odeur ms de miel dans une solution d'eau à une concentration 1:100 à 33 ° C. Chaque ligne tracée représente les canaux différenciés que l'étiquette à code de couleur des électrodes pattes à souder dans A. Bar:. 50 mV C) Après les procédures de pointes de tri des potentiels d'action simples sont classés et codés par couleur. Superposition des unités triées illustre la séparation des formes d'ondes. D) Transitoire intervalle histogramme indique la qualité de l'unité de séparation triées comme preuve de tri de pointe adéquate. Noter qu'il n'y a pas de pic à l'intérieur de la période réfractaire de l'unité. E) Deux vues (E1, E2) à partir deun groupement 3D de l'unité de tri avec analyse en composantes principales qui indique la distance entre les unités triées à l'autre. Les cercles indiquent l'2,5 fois la distance de Mahalanobis qui ressemble à la SD dans l'espace et indique une différenciation significative des grappes dans le composant principal espace F) Couleur unités codées représentent les potentiels visibles dans un grossissement de trois canaux de l'un enregistrement des voies d'action. S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
{kind=link}

Figure 4. Visualisation de fin d'enregistrement et la reconstruction 3D de la position d'enregistrement. A) Vue de projection de ortho-tranches le long des axes z avec l'alignement de l'intensité maximale d'un antéroet le remblayage rétrograde des neurones de projection uniglomerular saillie de la AL à la MB et LH. Le traceur intracellulaire Microruby (tetramethylrhodamin dextran) a été inséré dans le AL après les expériences d'enregistrement. Glomérules teinté à l'intérieur de la vue AL preuve coloration PN bon. B) Projection de tranches ortho long des axes z avec l'alignement de l'intensité maximale d'une coloration des deux électrodes avec Alexa HYDRAZIDE 488 indiquant le placement des électrodes pour le m-ALT (E1, flèche ) et l-ALT (E2, flèche). Le traceur Alexa 488 Hydrazide migre vers le tissu entourant l'électrode et colore le site d'insertion de l'électrode. Remarque, la coloration de premier plan dans l'AL est un artefact superficielle. C) 3D Reconstitution des cellules cibles colorées (SNP) et le site d'insertion de l'électrode de A, B (côté droit) avec un aperçu schématique du système olfactif des abeilles (à gauche côté) avec l'indication de la L-et m-ALT trajectoires. Remarque, ne uniglomertracts culier PN sont présentés. AN: nerf antennaire, AL: lobe antennaire, LH: corne latérale, MB: corps de champignon, E1, E2: sites d'insertion d'électrode, m-ALT: médial antennaire lobe voies, l-ALT: latéral antennaire lobe voies, c: caudale, r: rostrale, m: interne, l:. latéral S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
{kind=link}

Figure 5. Exemple d'odeur concentration codage dans la voie olfactive double. A) parcelles de chaleur illustrent la réponse d'un seul l-ALT (vert) et unique m-ALT PN (violet) acquises simultanément à partir d'une abeille individuelle en réponse à la substance odorante hexanal à l'augmentation des concentrations d'odeurs (de 1: 10 -6 à 1:100). Chaque ligne est une moyenne de dix stim d'essailation. La cadence de tir est montré que le changement d'intensité relative qui est le taux d'allumage par rapport à l'activité spontanée soustrait. B) réponse de la population de la latence de 11 l et 13 m-ALT unités de raccordement à partir de 7 abeilles enregistrées. Dans chaque abeille les unités de raccordement des deux voies ont été enregistrées simultanément. La latence de la réponse de la population présente un temps de latence en diminuant avec l'augmentation de la concentration d'odeur dans les unités de raccordement de deux voies. Le début de la réponse d'odeur à l'antenne à 99 ms a été enregistré via électroantennogrammes et est soustrait des latences de réponse PN. Notez que les concentrations les plus faibles réponses sont trop faibles pour les mesures de latence et, par conséquent, ont été exclues réponse. C) de la population cadence de tir de mêmes unités de raccordement que dans B. Avec l'augmentation de la concentration de l'odeur de la force de réaction augmente. Le l-ALT montre une force de la réponse forte. En B, C la moyenne et écart-type sont donnés.

Figure 6. Comparaison de l'activité de la population à deux étapes de traitement ultérieures long parcours olfactif de l'abeille. Données ont été enregistrées dans 20 des animaux qui ont été stimulées avec 1-hexanol et le 2-octanone. A) Chaque ligne représente la fausse couleur codée de la cadence de tir moyenne d'une projection neurone (PN) calculé sur 10 répétitions d'odeur de 1-hexanol. présentation de l'odeur commence au temps 0 et a duré trois secondes. b) montre la même que dans A), mais pour le corps de champignon neurones extrinsèques (EN). La matrice représentée en A) peut être vue comme un vecteur de population PN pendant la stimulation des odeurs avec le 1-hexanol. Nous avons calculé le même genre de vecteur de la population lors de la stimulation de l'odeur avec le 2-octanone et utilisé les deux vecteurs dans une analyse en composantes principales (ACP) en gardant la dimension temporelle. C) Les trois premières composantes principales (PC1, 2 et 3) ont ététracée par rapport à l'autre pour illustrer la séparation de l'odeur dans l'ensemble de l'activité PN à la sortie du lobe antennaire. Le temps avant l'apparition odeur est indiqué en noir. Activité pendant trois secondes stimulation avec 1-hexanol est représenté en bleu. L'activité pendant la stimulation avec le 2-octanol est indiquée en rouge. En outre, nous montrons 1,5 secondes (post) de l'activité après l'odeur d'un ensemble de 1-hexanol (bleu clair) et 2-octanonen (rose). Notez que le niveau d'ensemble PN, les deux odeurs évoquant des trajectoires très différentes s'installer dans un «point fixe», qui survit à l'ensemble de la période de stimulation de l'odeur. Seulement après odeur compenser les trajectoires se déplacent vers l'activité de base sans stimulation olfactive. D) La même analyse a été effectuée au niveau d'ensemble FR représentatif de l'activité à la sortie du corps de champignon. Par rapport à l'activité de PN odeurs évoquent une trajectoire moins distinctes. En outre, un «point fixe» n'est pas observable. Les initialement odeur trajectoires induites porcelainermingle avec l'activité de la ligne de base bien que l'odeur est toujours présente. Seul le décalage odeur évoqué une trajectoire supplémentaire.
. Film 1 Heure résolu évaluation d'une odeur induite trajectoire après analyse en composante principale d'un vecteur PN-population (à gauche) et un vecteur de la population FR. (à droite; cp. Figure 6) Les parties supérieures sont les trois premières composantes principales (PC1, 2 et 3) en fonction de l'autre. Les panneaux inférieurs illustrent l'évaluation de PC1, 2 et 3 au cours du temps. stimulation de l'odeur est marquée par la barre grise. Tous les panneaux ont été synchronisés. Notez que l'activité de la population FR commence un peu avant l'activité de la population PN, un phénomène qui semble être contraintuitive mais peut s'expliquer par la connectivité et les propriétés des couches impliquées, dont il est question plus haut 1.
Discussion
Cet article démontre la production et l'utilisation de la voie micro fils-électrodes multiples conçus sur mesure. Les électrodes décrites sont adaptés pour l'enregistrement des deux seule unité et l'activité de la population qui est particulièrement utile pour les mesures de latence et d'autres propriétés de réponse temporelle de différents neurones et différentes neuropils dans un seul échantillon (pour plus de détails voir 1,2,25). En outre, nous avons montré comment mettre en œuvre de façon permanente les fils-électrodes micro pour permettre des enregistrements stables à long terme en se comportant abeilles qui durent pendant des heures jusqu'à jours.
Enregistrements multi-unitaires extracellulaires sont devenus un outil favorable pour atteindre une haute résolution temporelle combinée avec l'information spatiale. Dans notre cas, ce sont des voies neuronales parallèles 2 ou deux neuropils différentes 1. Neurones multiples peuvent être enregistrés et analysés au niveau du neurone unique en parallèle et à haute résolution temporelle. Dossier Multi-unitréunions ont été appliquées en premier chez les mammifères 38 et plus tard aussi chez les insectes 39-41. Progrès considérables ont été accomplis dans le développement et l'amélioration des techniques d'enregistrement multi-canaux extracellulaires 42,43. C'est, par exemple, comprend le développement de nouvelles électrodes 44 ou de nouvelles pointes de tri et de regroupement des algorithmes 45. Méthodes générales de techniques d'enregistrement multi-unitaires extracellulaires sont bien décrites 46-48. Les électrodes auto construit présentés dans cette vidéo peuvent en outre être adaptées en ajoutant plus de microfils par électrode ou les micro fils peuvent être tordus pour gagner des distances constantes mesurables entre les pointes. Les deux procédures seraient, toutefois, conduire à la diminution de la flexibilité et augmentation de l'épaisseur de l'électrode.
Par rapport à des sondes de silicium couramment utilisés pour les enregistrements extracellulaires chez les insectes beaucoup plus comme la mite de faucon, criquets et blattes 40,49 - 51 micro-électrodes métalliques décrits sont plus petites, flexibles et peuvent facilement faire face à des mouvements du cerveau potentiels et, par conséquent, peuvent être utilisés de manière fiable dans les petits insectes sociaux comme les abeilles et les fourmis qui montrent un répertoire comportemental beaucoup plus large. La plupart des sondes en silicone ont tige pointue comme structures de coupe axones et les tissus nerveux le long de leur chaîne d'insertion, tandis que les fils micro décrites sont ronds, souples et plus petites et sont donc moins nocifs pour le tissu environnant qui est un avantage évident si l'objectif est d'étudier à long plasticité terme dans un animal intact et de se comporter. Un autre avantage de micro électrodes à fil est leur production à faible coût et la facilité de manipulation. Au lieu de nettoyer soigneusement une sonde de silicone cher les fils d'électrode sont fraîchement coupés avant l'insertion du cerveau et, par conséquent, les problèmes de manque de congestion. En outre, il est possible d'utiliser plus d'un micro fil-électrode dans la même préparation, soit inséré dans différentes neuropiles 1 osecteurs de r 2 que nous montrent ici. Cette approche est particulièrement favorable pour analyser et comparer les aspects temporels comme les latences et les interactions réponse à différents niveaux de traitement neural.
Nous sommes conscients du fait qu'un signal enregistré extracellulaire ne reflète pas l'activité de la cellule unique en soi. Il est toujours d'un composé de l'activité de tension autour de la pointe de l'électrode. Pour localiser la source du signal de la différence de deux micro-canaux voisins de fil à l'intérieur une électrode est toujours calculée. Ainsi, la source pour les signaux de pointes utilisées pour extraire l'activité d'une seule unité était toujours très proche de l'un ou l'autre canal d'électrode résultant en des formes d'onde de pic faciles à distinguer. Signaux de plus loin, comme l'activité musculaire ou l'activité de neuropils voisins, atteignent les deux électrodes en même temps évoquant des formes et des amplitudes comparables et seront rejetées par cette procédure. En utilisant la technique modèle correspondant de Spike2,nous sommes très confiants d'obtenir une activité d'une seule unité, ce qui n'est pas la même, mais très proche de l'activité des neurones unique. Cependant, la question de la pointe tri pourrait être évité en utilisant des techniques d'enregistrement intracellulaire.
Enregistrements de cellules uniques soit avec des électrodes pointus ou pipettes de patch permettent une connaissance approfondie sur les propriétés physiologiques d'un seul neurone. Toutefois, en raison de la petite taille des neurones d'insectes et de leurs axones (p. ex., Moins de 1 um pour abeilles unités de raccordement 52) Enregistrements seulement à court terme sont gérables. En outre, les enregistrements intracellulaires pourraient être invasive et pourraient nuire à la cellule qui est peut-être une autre raison pour les limitations temporelles. In vivo des enregistrements intracellulaires chez les insectes survivent rarement une heure. Une fenêtre de temps qui était suffisant pour le travail de pionnier de Martin Marteau 53 qui a enregistré intracellulaire à partir d'un seul neurone identifié, la ventrale non apparié maxilar neurone n ° 1 (VUMmx1). Il ne pouvait lson activité encre directement à la voie de la récompense. Juliane Mauelshagen 54 inscrit intracellulaire de l'activité d'un champignon corps extrinsèque neurone identifié, le pédicule neurone extrinsèque # 1 (PE1) pendant le conditionnement classique. Le même neurone était dans le centre de Menzel et Manz 55 quand ils ont trouvé LTP après stimulation électrique des cellules de Kenyon. Cependant, Okada et ses collègues 56 peuvent utiliser le modèle de dopage intracellulaire bien caractérisée (des pointes doubles et triples) pour l'identification de la PE1 pendant les enregistrements extracellulaires. Après tout une combinaison des deux méthodes, des enregistrements intracellulaires de neurones identifiés et des enregistrements extracellulaires à long terme pourraient être un outil puissant pour les enquêtes futures.
Cependant, en utilisant des électrodes pointus pour enregistrer plusieurs cellules (unités) simultanément à différents niveaux de traitement plus longues heures jusqu'à jours pour analyser leurs relations temporelles de réponse et / ou même des changements plastique is presque impossible.
Avec la première imagerie calcique approches de l'abeille 57,58 utilisant des colorants sensibles au calcium de l'analyse de la répartition spatiale des réponses d'odeurs étaient accessibles 59-62. Cependant, dans de nombreux cas, les colorants sensibles de calcium doivent être introduites dans le tissu de cerveau par des manipulations envahissantes qui limitent encore la durée de vie de l'abeille et les propriétés intrinsèques des cellules analysées. Ce problème est résolu dans d'autres organismes modèles comme la drosophile en utilisant des capteurs de calcium génétiquement introduites 63,64. Cependant, en général, des capteurs de calcium peuvent introduire d'autres limitations, car ils peuvent agir comme des tampons de calcium susceptibles d'influer sur les propriétés temporelles de réponses d'odeurs. Enregistrements intracellulaires simultanés combinés avec l'imagerie de calcium ou approches informatiques peuvent prouver la bonne résolution temporelle de l'imagerie traite 65,66. Cependant, la résolution temporelle du processus de formation d'image lui-même est rather limitée. Systèmes d'acquisition optiques utilisent généralement CCD-imagerie avec une résolution temporelle de 5-20 Hz 67, bien que 2-Photon Imaging pourrait être en mesure d'acquérir des séquences rapides 68. Cependant, le taux d'échantillonnage de plus en plus va toujours avec une perte de résolution spatiale. En outre, les colorants sensibles au calcium utilisés dans l'abeille à un blanchiment, ce qui réduit également le temps d'acquisition 69.
Comparé à d'autres techniques d'enregistrement physiologiques chez les insectes nos électrodes souples micro-fil multi-canal d'assurer l'accès de longue date à l'unité unique et de la population de l'activité neuronale en comporter abeilles.
Nous avons démontré l'utilisation de deux de ces électrodes à différents étages de traitement dans le même animal, ce qui facilite l'analyse des aspects de codage temporelles entre les différents sites d'enregistrement. En fonction de la problématique de recherche et le modèle d'insecte la méthode de base de la construction de l'électrode démontré ici est facilement s'étendentmesure et / ou qui peut être adapté. Par exemple, il est concevable d'utiliser plus de trois fils isolés pour produire des électrodes à canaux multiples. En outre, le nombre de sites d'enregistrement peut être étendu et en observant les aspects temporels de plus de deux voies ou neuropils est réalisable. Notre espoir est que cette méthode va inspirer de nombreux scientifiques et contribuera positivement à la compréhension du traitement neuronal sophistiqué petits cerveaux.
Déclarations de divulgation
Les auteurs déclarent qu'ils n'ont aucun intérêt financier concurrents.
Remerciements
The authors would like to thank Isabelle Reus for establishment of tracing the electrode insertion side, Tobias Rosenbaum for LabView programming, Anneke Meyer for data analyzes and helpful discussions. We thank Randolf Menzel for discussion and practical help during early stage of electrode development. Furthermore we thank Brian Smith for postdoctoral association to MS-B. This work was supported by the Deutsche Forschungsgemeinschaft (DFG, SPP 1392, Ro1177/5-2) to WR.
matériels
| Name | Company | Catalog Number | Comments |
| Paraffin oil | Fluka | 76235 | |
| Odors | Sigma Aldrich | ||
| PBS | pH 7.2 | ||
| 4% Formaldehyde | ThermoScientific | 28908 | Methanol free |
| Triton X | BioChemica | A1388 | |
| Methylsalicylate | Roth | 4529.1 | |
| Tetramethylrhodamin dextran, 10,000 MW (Microruby) | Invitrogen | D7162 | keep dark |
| Alexa 488 hydrazide | Invitrogen | A-10436 | keep dark |
| Alexa 568 hydrazide | Invitrogen | A-10437 | keep dark |
| Bee Ringer Solution | see 2 | ||
| Polyurethane-coated copper wire | Elektrisola | 15µm diameter & P155 insulation | |
| Dental Wax | Densply Detrey | 64103015S1 | moderate melting point |
| Dental Wax | Flexaponal | 124-202-00 | low-melting Wax |
| KWIK SIl | WPI | 03L | |
| 18 Pin Socket | Conrad Electronic | 189634-62 | |
| Hot melting glue | Conrad Electronic | 827673 | |
| soldering needle | Conrad Electronics | 830283 | 12 V |
| Soldering terminal lug | Conrad Electronic | 531901 | |
| Glaselectrodes | WPI | 1B100F-3 | |
| Minutien Pins | Fine Science Tools | 26002-20 | V2A 0.2 x 12 mm |
| switchable headstage | Tucer Davis Technologies | SH16 | |
| Headstage connection module | NPI | INT-03M | |
| Amplifier Module | NPI | PDA-2F | |
| Data Acquisition boards | National Instruments | NI-6123, Ni-6143 | |
| Acquisition Software | National Instruments | Lab View 8.2 | custom design |
| Spike-Sorting | CED | Spike 2 v7.11 | |
| Matlab | Mathworks | R2008B | |
| Micromanipulator | Leitz | manual | |
| AG-wires | WPI | AGT05100 | |
| Confocal laser scanning microscope | Leica | TCS SP2 AOBS | |
| AMIRA | Mercury Computer Systems | 2/5/2000 |
Références
- Strube-Bloss, M. F., Herrera-Valdez, M. a., Smith, B. H. Ensemble response in mushroom body output neurons of the honey bee outpaces spatiotemporal odor processing two synapses earlier in the antennal lobe. PLoS ONE. 7 (11), (2012).
- Brill, M. F., Rosenbaum, T., Reus, I., Kleineidam, C. J., Nawrot, M. P., Rössler, W. Parallel processing via a dual olfactory pathway in the honeybee. The Journal of neuroscience the official journal of the Society for Neuroscience. 33 (6), 2443-2456 (2013).
- Menzel, R. The honeybee as a model for understanding the basis of cognition. Nature Reviews Neuroscience. 13 (11), 758-768 (2012).
- Sandoz, J. Behavioral and neurophysiological study of olfactory perception and learning in honeybees. Frontiers in Systems Neuroscience. 5, (2011).
- Giurfa, M. Cognition with few neurons: higher-order learning in insects. Trends in Neurosciences. 36 (5), 1-10 (2013).
- Menzel, R., Giurfa, M. Cognitive architecture of a mini-brain: the honeybee. Trends in cognitive sciences. 5 (2), 62-71 (2001).
- Hildebrand, J. G., Shepherd, G. M. Mechanisms of olfactory discrimination: converging evidence for common principles across phyla. Annual Review of Neuroscience. 20, 595-631 (1997).
- Wilson, R. I., Mainen, Z. F. Early events in olfactory processing. Annu. Rev. Neurosci. 29, 163-201 (2006).
- Streinzer, M., Kelber, C., Pfabigan, S., Kleineidam, C. J., Spaethe, J. Sexual dimorphism in the olfactory system of a solitary and a eusocial bee species. The Journal of comparative neurology. 521 (12), (2013).
- Nishino, H., Nishikawa, M., Mizunami, M., Yokohari, F. Functional and topographic segregation of glomeruli revealed by local staining of antennal sensory neurons in the honeybee Apis mellifera. The Journal of comparative neurology. 515 (2), 161-180 (2009).
- Schneider, D. Elektrophysiologische Untersuchungen von Chemo- und Mechanorezeptoren der Antenne des Seidenspinners Bombyx mori L. Zeitschrift für Vergleichende Physiologie. 40 (1), 8-41 (1957).
- Menzel, R., Rybak, J. Antennal lobe of the honeybee. Handbook of brain microcircuits. , 427-432 (2011).
- Girardin, C. C., Kreissl, S., Galizia, C. G. Inhibitory connections in the honeybee antennal lobe are spatially patchy. Journal of neurophysiology. 109 (2), 332-343 (2013).
- Meyer, A., Galizia, C. G. Elemental and configural olfactory coding by antennal lobe neurons of the honeybee (Apis mellifera). Journal of comparative physiology. A, Neuroethology, sensory, neural, and behavioral. 198 (2), 159-171 (2012).
- Abel, R., Rybak, J., Menzel, R. Structure and response patterns of olfactory interneurons in the honeybee, Apis mellifera. The Journal of comparative neurology. 437 (3), 363-383 (2001).
- Kirschner, S., Kleineidam, C. J., Zube, C., Rybak, J., Grünewald, B., Rössler, W. Dual olfactory pathway in the honeybee, Apis mellifera. The Journal of comparative neurology. 499 (6), 933-952 (2006).
- Galizia, C. G., Rössler, W. Parallel olfactory systems in insects: anatomy and function. Annual review of entomology. 55 (August), 399-420 .
- Ito, K., Shinomiya, K., et al. A coordinated nomenclature system for the insect brain. Neuron. 81 (4), 755-765 (2014).
- Rybak, J. The digital honey bee brain atlas. Honeybee Neurobiology and Behavior. , 125-140 (2012).
- Rössler, W., Brill, M. F. Parallel processing in the honeybee olfactory pathway: structure, function, and evolution. Journal of comparative physiology. A, Neuroethology, sensory, neural, and behavioral. 199 (11), (2013).
- Mobbs, P. The brain of the honeybee Apis mellifera. I. The connections and spatial organization of the mushroom bodies. Philosophical transactions of the Royal Society of London. Series B, Biological. 298 (1091), 309-354 (1982).
- Strausfeld, N. J. Organization of the honey bee mushroom body: representation of the calyx within the vertical and gamma lobes. The Journal of comparative neurology. 450 (1), 4-33 (2002).
- Witthöft, W. Absolute Anzahl und Verteilung der Zellen im Hirn der Honigbiene. Zeitschrift für Morphologie der Tiere. 61 (1), 160-184 (1967).
- Rybak, J., Menzel, R. Anatomy of the mushroom bodies in the honey bee brain: the neuronal connections of the alpha-lobe. The Journal of Comparative Neurology. 465, 444-465 (1993).
- Strube-Bloss, M. F., Nawrot, M. P., Menzel, R. Mushroom body output neurons encode odor-reward associations. The Journal of neuroscience : the official journal of the Society for Neuroscience. (8), 3129-3140 (2011).
- Haddad, R., Lanjuin, A., Madisen, L., Zeng, H., Murthy, V. N., Uchida, N. Olfactory cortical neurons read out a relative time code in the olfactory bulb. Nature neuroscience. (May), 1-11 (2013).
- Martin, J. P., Beyerlein, A., et al. The neurobiology of insect olfaction: Sensory processing in a comparative context. Progress in neurobiology. 95 (3), 427-447 (2011).
- Nawrot, M. P. Dynamics of sensory processing in the dual olfactory pathway of the honeybee. Apidologie. 43 (3), 269-291 (2012).
- Farkhooi, F., Froese, A., Muller, E., Menzel, R., Nawrot, M. P. Cellular adaptation facilitates sparse and reliable coding in sensory pathways. PLoS computational biology. 9 (10), e1003251 (2013).
- Saha, D., Leong, K., Katta, N., Raman, B. Multi-unit recording methods to characterize neural activity in the locust (Schistocerca americana) olfactory circuits. Journal of visualized experiments JoVE. (71), (2013).
- Saha, D., Leong, K., Li, C., Peterson, S., Siegel, G., Raman, B. A spatiotemporal coding mechanism for background-invariant odor recognition. Nature neuroscience. 16 (12), 1-13 (2013).
- Mizunami, M., Okada, R., Li, Y., Strausfeld, N. J. Mushroom Bodies of the Cockroach Activity and Identities of Neurons. Journal of Comparative Neurology. 519 (July), 501-519 (1998).
- Okada, R., Ikeda, J., Mizunami, M. Sensory responses and movement-related activities in extrinsic neurons of the cockroach mushroom bodies. Journal of Comparative Physiology A: Sensory, Neural, and Behavioral Physiology. 185 (2), 115-129 (1999).
- Haehnel, M., Froese, A., Menzel, R. In vivo Ca2+ imaging of mushroom body neurons during olfactory learning in the honey bee. Journal of visualized experiments JoVE. (30), (2009).
- Felsenberg, J., Gehring, K. B., Antemann, V., Eisenhardt, D. Behavioural pharmacology in classical conditioning of the proboscis extension response in honeybees (Apis mellifera). Journal of visualized experiments JoVE. (47), (2011).
- Rehder, V. Quantification of the honeybee’s proboscis reflex by electromyographic recordings. Journal of Insect Physiology. 33 (7), 501-507 (1987).
- Serrano, E., Nowotny, T., Levi, R., Smith, B. H., Huerta, R. Gain control network conditions in early sensory coding. PLoS computational biology. 9 (7), e1003133 (2013).
- Hubel, D. H., Wiesel, T. N. Receptive fields and functional architecture of monkey striate cortex. The Journal of physiology. 195 (1), 215-243 (1968).
- Christensen, T. A., Pawlowski, V. M., Lei, H., Hildebrand, J. G. Multi-unit recordings reveal context-dependent modulation of synchrony in odor-specific neural ensembles. Nature neuroscience. 3 (9), 927-931 (2000).
- Byers, K. J. R. P., Sanders, E., Riffell, J. A. Identification of olfactory volatiles using gas chromatography-multi-unit recordings (GCMR) in the insect antennal lobe. Journal of visualized experiments JoVE. (72), e4381 (2013).
- Perez-Orive, J., Mazor, O., Turner, G. C., Cassenaer, S., Wilson, R. I., Laurent, G. Oscillations and sparsening of odor representations in the mushroom body. Science. 297 (5580), 359-365 (2002).
- Stevenson, I. H., Kording, K. P. How advances in neural recording affect data analysis. Nature. 14 (2), 139-142 (2011).
- Buzsáki, G. Large-scale recording of neuronal ensembles. Nature neuroscience. 7 (5), 446-451 (2004).
- Viventi, J., Kim, D. -. H., et al. Flexible, foldable, actively multiplexed, high-density electrode array for mapping brain activity in vivo. Nature neuroscience. 14 (12), 1599-1605 (2011).
- Pouzat, C., Mazor, O., Laurent, G. Using noise signature to optimize spike-sorting and to assess neuronal classification quality. Journal of neuroscience methods. 122 (1), 43-57 (2002).
- Lewicki, M. S. A review of methods for spike sorting: the detection and classification of neural action potentials. Network. 9 (4), R53-R78 (1998).
- Quian Quiroga, R., Panzeri, S. Extracting information from neuronal populations: information theory and decoding approaches). Nature reviews. Neuroscience. 10 (3), 173-185 (2009).
- Einevoll, G. T., Franke, F., Hagen, E., Pouzat, C., Harris, K. D. Towards reliable spike-train recordings from thousands of neurons with multielectrodes. Current opinion in neurobiology. 22 (1), 11-17 (2012).
- Riffell, J. a., Lei, H., Abrell, L., Hildebrand, J. G. Neural basis of a pollinator’s buffet: olfactory specialization and learning in Manduca sexta. Science. 164 (6), 877-892 (2013).
- Bender, J. a., Pollack, A. J., Ritzmann, R. E. Neural activity in the central complex of the insect brain is linked to locomotor changes. Current biology : CB. 20 (10), 921-926 (2010).
- Perez-Orive, J., Bazhenov, M., Laurent, G. Intrinsic and circuit properties favor coincidence detection for decoding oscillatory input. The Journal of neuroscience the official journal of the Society for Neuroscience. 24 (26), 6037-6047 (2004).
- Rybak, J. . Die strukturelle Organisation der Pilzkörper und synaptische Konnektivität protocerebraler Interneuronen im Gehrin der Honigbiene, Apis mellifera.: eine licht- und elektronenmikroskopische Studie. , (1994).
- Hammer, M. An identified neuron mediates the unconditioned stimulus in associative olfactory learning in honeybees. Nature. 366, 59-63 (1993).
- Mauelshagen, J. Neural correlates of olfactory learning paradigms in an identified neuron in the honeybee brain. Journal of neurophysiology. 69, 609-625 (1993).
- Menzel, R., Manz, G. Neural plasticity of mushroom body-extrinsic neurons in the honeybee brain. The Journal of experimental biology. 208 (22), 4317-4332 (2005).
- Okada, R., Rybak, J., Manz, G., Menzel, R. Learning-related plasticity in PE1 and other mushroom body-extrinsic neurons in the honeybee brain). The Journal of neuroscience: the official journal of the Society for Neuroscience. 27 (43), 11736-11747 (2007).
- Joerges, J., Küttner, A., Galizia, C. G., Menzel, R. Representation of odours and odour mixtures visualized in the honeybee brain. Nature. 387, 285-288 (1997).
- Galizia, C. G., Joerges, J., Küttner, A., Faber, T., Menzel, R. A semi-in-vivo preparation for optical recording of the insect brain. Journal of neuroscience. 76 (1), 61-69 (1997).
- Galizia, C. G., Sachse, S., Rappert, A., Menzel, R. The glomerular code for odor representation is species specific in the honeybee Apis mellifera. Nature. 2 (5), 473-478 (1999).
- Sandoz, J. -. C. Odour-evoked responses to queen pheromone components and to plant odours using optical imaging in the antennal lobe of the honey bee drone Apis mellifera L. The Journal of experimental biology. 209 (18), 3587-3598 (2006).
- Fernandez, P. C., Locatelli, F. F., Person-Rennell, N., Deleo, G., Smith, B. H. Associative conditioning tunes transient dynamics of early olfactory processing. The Journal of neuroscience the official journal of the Society for Neuroscience. 29 (33), 10191-10202 (2009).
- Locatelli, F. F., Fernandez, P. C., et al. Nonassociative plasticity alters competitive interactions among mixture components in early olfactory processing. European Journal of Neuroscience. 37 (1), (2013).
- Silbering, A. F., Bell, R., Galizia, C. G., Benton, R. Calcium imaging of odor-evoked responses in the Drosophila antennal lobe. Journal of visualized experiments JoVE. (61), 1-7 (2012).
- Strutz, A., Völler, T., Riemensperger, T., Fiala, A., Sachse, S. Calcium imaging of neural activity in the olfactory system of Drosophila. Genetically Encoded Functional Indicators. 72, 43-70 (2012).
- Galizia, C. G., Kimmerle, B. Physiological and morphological characterization of honeybee olfactory neurons combining electrophysiology, calcium imaging and confocal microscopy. Journal of Comparative Physiology A: Neuroethology, Sensory, Neural, and Behavioral Physiology. 190 (1), 21-38 (2004).
- Helmchen, F., Waters, J. Ca2+ imaging in the mammalian brain in vivo. European journal of pharmacology. 447 (2-3), 119-129 (2002).
- Stierle, J. S., Galizia, C. G., Szyszka, P. Millisecond stimulus onset-asynchrony enhances information about components in an odor mixture. Journal of Neuroscience. 33 (14), 6060-6069 (2013).
- Haase, A., Rigosi, E., et al. In-vivo two-photon imaging of the honey bee antennal lobe. Biomedical optics express. 2 (1), 131-138 (2010).
- Becker, P. L., Fay, F. S. Photobleaching of fura-2 and its effect on determination of calcium concentrations. The American journal of physiology. 253 (4 pt 1), C613-C618 (1987).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.