
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
ויזואליזציה של HIV-1 Gag מחייב את הענק Unilamellar שלפוחיות (בוס) ממברנות
In This Article
Summary
We illustrate here an in vitro membrane binding assay in which interactions between HIV-1 Gag and lipid membranes are visually analyzed using YFP-tagged Gag synthesized in a wheat germ-based in vitro translation system and GUVs prepared by an electroformation technique.
Abstract
החלבון המבני של HIV-1, Pr55 Gag (או Gag), נקשר הממברנה בתאים במהלך תהליך ההרכבה וירוס. ממברנה עקדת Gag היא מהווה נדבך חיוני להיווצרות חלקיק וירוס, מאז פגם בתוצאות מחייב קרום Gag ליקוי חמור של ייצור החלקיקים ויראלי. כדי לקבל פרטים מכניסטית של אינטראקציות קרום שומנים Gag, שיטות חוץ גופייה מבוססת על תמ"ג, footprinting חלבון, תהודת plasmon פני שטח, צנטריפוגה הנפקת liposome, או חרוז שומני קרינת כריכה פותח עד כה. עם זאת, כל אחת משיטות במבחנה אלה יש מגבלות. כדי להתגבר על כמה מגבלות אלה ולספק גישה משלים לשיטות הוקמו בעבר, פתחנו assay במבחנה שבה אינטראקציות בין HIV-1 Gag וממברנות שומנים להתקיים בסביבה "דמוי תא". ב assay זה, Gag מחייב ממברנות שומנים בדם מנותח ויזואלית באמצעות YFP-taggeד Gag מסונתז ניבט חיטה מבוססת במערכת תרגום במבחנת GUVs שהוכן על ידי טכניקת electroformation. כאן נתאר את הרקע ואת הפרוטוקולים להשיג חלבונים Gag באורך מלא myristoylated וממברנות Guv הכרחי עבור assay וכדי לזהות Gag-בוס מחייב ידי מיקרוסקופ.
Introduction
סוג 1 הנגיף (HIV-1) הכשל החיסוני האנושי הוא וירוס אפוף כי מרכיב ב וניצנים מן הקרום פלזמה (PM) בסוגי תאים ביותר. הרכבה של חלקיקי הנגיף HIV-1 הוא מונע על ידי חלבון הליבה ויראלי 55 kDa שנקרא Pr55 Gag (GAG). Gag הוא מסונתז כמבשר polyprotein מורכב מארבעה תחומים מבניים גדולים, כלומר, מטריצה, קפסיד, nucleocapsid, ו P6, וכן שני פפטידים spacer SP1 ו- SP2. במהלך האסיפה, תחום מטריקס (MA) אחראי המיקוד של Gag לאתר ההרכבה, קפסיד (CA) תחום מתווכת אינטראקציות Gag-Gag, את nucleocapsid (NC) תחום מגייס RNA הגנומי ויראלי, וגרם מארח המתגייס P6 שסיוע scission חלקיק וירוס מן קרום הפלזמה. Gag גם הוא עובר שינוי שיתוף translational על ידי תוספת של חומצת שומן 14-חמצני או מחצית myristate בבית שלה N- הסופית.
ממברנה עקדת Gag היא תנאי הכרחי עבור הרכבת ויראלי, מאז מ 'utants כי הם פגומים קרום מחייב מצליח לייצר חלקיקי נגיף. אנחנו ואחרים הראינו כי קרום עקדת Gag מתווכת על ידי אותות bipartite בתוך תחום MA: מחצית myristate N-terminal שמתווכת אינטראקציות הידרופוביות עם bilayer השומנים מקבץ של שאריות בסיסיות בתוך תחום MA כפי שכונה באזור הבסיסי ביותר ( HBR) כי אינטראקציה עם שומנים חומצי על PM 1-4. מחקרים של אינטראקציות Gag-קרום באמצעות ביטוי אקטופי של IV 5-phosphatase polyphosphoinositide (5ptaseIV), אנזים המזרז הידרוליזה של PM-ספציפי phosphatidylinositol- פוספוליפידים חומצי (4,5) [PI (4,5) P 2] -bisphosphate אל פוספט phosphatidylinositol-4, בתאים הציע לוקליזציה Gag-PM מתווכת על ידי PI (4,5) P 2 3,5. עם זאת, במחקרים במבחנה מכוונים להבנת הפרטים מכניסטית יותר של Gag-PI (4,5) P 2 אינטראקציות הוכיחו להיות אתגר עבור מספר סיבות. ללמשל, טיהור באורך מלא myristoylated Gag HIV-1 עבור ניסויים ביוכימיים כבר קשה מבחינה טכנית לפחות בחלקו בשל הנטייה של Gag כדי לצבור במהלך טיהור. לפיכך, צורות קטועות של HIV-1 Gag, כגון MYR-MA או MYR-MA-CA, או בצורה בלתי myristoylated שמשו לעתים קרובות במחקרים המחייבים טיהור Gag (למשל, תהודה מגנטית גרעינית, footprinting חלבון, משטח plasmon תהודה 6-9). לחלופין, מצמיד בתגובות תרגום-שעתוק במבחנה שמש לייצר myristoylated באורך מלא HIV-1 Gag במחקרים ביוכימיים אחרים 1,2. בדרך כלל במערכת זו, פלסמיד קידוד Gag הוא עיבד ותרגם עם lysates תא אוקריוטים (למשל, lysates reticulocyte ארנב) כי הם נטולי כל קרומים הסלולר RNAs שליח אך מכילים את המכונות שעתוק ותרגום. לאחר התגובה, lysates תא המכיל Gag מעורבבים איתיmbranes לניתוח אינטראקציות Gag עם שומנים. בנוסף הקלות של הכנה באורך מלא myristoylated Gag, שיטות באמצעות מערכת תרגום שעתוק במבחנה יש יתרון כי Gag סינתזה ותגובות מחייב קרום עוקב מתרחשים אצל המילייה 'cytosol הדמוי איקריוטיים' כי עשויים לייצג מצבים פיסיולוגיים טובים יותר. מאפיין זה תרם מחקרים שהראו כי מולקולות RNA כבול לתחום MA להסדיר Gag מחייב שומנים חומצי באופן תחרותי 1,2,10-12. עם זאת, מאז את הסכום הכולל של חלבוני Gag שהושגו lysates התאים אלה אינם גבוהים, תיוג המטבולית של חלבונים עם חומצות אמינו radiolabeled הכרח הגילוי שלהן.
בהתאם לשיטה למדוד אינטראקציות Gag-שומנים, מגוון של הכנות קרום שמש. כל אחת משיטות אלו יש עוצמות ומגבלותיה. רוב המבחנים מבוססים NMR דורשים שימוש ליפידים עם acyl הקצררשתות המהוות מסיסים במים (למשל, C4- ו C8-PI (4,5) P 2) 6,8. בעוד שיטות תמ"ג לבדוק מחייב של Gag אל שומנים שיש שרשראות acyl ארוך המצוי בתאי מפותחים, הם שימשו רק עם MA myristoylated או nonmyristoylated עד כה 8,13. לחלופין, ליפוזומים שהוכן שומנים שיש שרשרות acyl אורך יליד שמש בשיטות ביוכימיות כגון הנפקת liposome או חרוז ליפוזום פלורסנט מבחני מחייב 2,3,10,14-16. עם זאת, ליפוזומים בשימוש מבחנים אלה יש קוטר קטן, ולכן יש הקרומים שלהם עקמומיות תלולה חיובית. לעומת זאת, בשלב המוקדם של מכלול חלקיקים בתאי HIV-1-נגועים, Gag נקשר לראש הממשלה, אשר כמעט מישוריים על הסקאלה של Gag, ובהמשך גורם עקמומיות שלילית במהלך ניצנים. לכן, ממברנות ליפוזום עם עקמומיות תלולה חיובית לא יכולות להיות דו-שכבה ליפידית אידיאלית ללמוד אינטראקציות שומנים-Gag. באשר ליפוזום שטויותמבחני ation, אזהרה נוספת היא פוטנציאל החשיפה של מתחמי Gag-שומנים כדי שיפוע סוכרוז היפרטוני במהלך צנטריפוגה עשוי להשפיע על התוצאה הניסיונית. כדי להקל על המגבלות האלה ולספק מערכת ניסיונית משלימים, מבחנים עבור Gag מחייב שלפוחית unilamellar ענקית (GUVs) פותחו בשנים האחרונות. GUVs הם שלפוחית bilayer השומנים יחיד אשר היקף בקטרים להאריך לכמה עשרות מיקרומטר. לפיכך, העקמומיות של הקרומים האלה דומה PM על הסקאלה של Gag. יתר על כן, בשל ממדיו הגדולים, המאפשר בדיקה ויזואלית תחת מיקרוסקופים אופטיים, קרום עקדת מתויג fluorescently או שכותרתו חלבוני Gag כדי שלפוחית אלה על ערבוב ניתן לקבוע בקלות ללא העיבוד הבא של מתחמי Gag-שומנים.
אנחנו כאן מתארים פרוטוקול ללמוד קרום HIV-1 Gag מחייב שימוש GUVs המתקבל בשיטה electroformation. שיטות שונות כגון לחות עדינה, הידרציה בסיוע ג'ל, מיקרופוןהטיסות הללו rofluidic, ו electroformation 17-22 שימשו להשיג GUVs. בשביל הפרוטוקול המתואר כאן, שיטת electroformation משמשת בעיקר בגלל יעילותה ביצירת GUVs עם שומנים חומצי וקלות היחסית של שימוש ללא הצורך של setups היקר. מאז להדמיה של Gag מחייבת כתב ניאון, חלבון פלואורסצנטי צהוב (YFP) מתווסף גנטית C- הסופית של Gag (Gag-YFP). חלבונים Gag-YFP מתקבלים על ידי תגובות שעתוק ותרגום במבחנה ב lysates נבט חיטה מבוסס על טכנולוגיה (CECF) זרימה רציפה רציף-החליפין. בטכנולוגיה זו, הן הסרת תוצרי לוואי מעכבות מהתגובות ואספקת מצעי תגובה ורכיבי אנרגית מושגות במנגנון המבוסס-דיאליזה. לקבלת התגובות האלה, קידוד פלסמיד Gag-YFP תחת שליטה של מקדם T7 משמש. מן הראוי לציין כי, כפי שראינו קודם, lysates נבט חיטה לתמוך myristoylation ללא רכיבים נוספים 23,24. באמצעות שיטה זו, ניתן היה להשיג כמויות מספיקות של באורך מלא myristoylated Gag-YFP להדמיה של Gag על ממברנות בוס 24. כאן אנו מתארים את הפרוטוקול שבה HIV-1 Gag מחייב PI (4,5) P 2 המכילים ממברנות Guv ניתן לבדוק ללא עיבוד שלאחר מכן ממושך בעקבות תגובות מחייבות ולהציע כי שיטה זו משלימה-קרום Gag קודם מבחני מחייב והוא יכול להיות להאריך עוד יותר להבין Gag-קרום HIV-1 מחייב.
Access restricted. Please log in or start a trial to view this content.
Protocol
יום 1: ביטוי של חלבונים Gag שימוש במערכת נבט חיטה Lysate מבוססי במבחנה תמלול-תרגום
1. הכנה של HIV-1 Gag
- הסר את כל ריאגנטים של ערכת CECF מסחרי נבט חיטה מן מקפיאים (lysates נבט חיטה מאוחסנים ב -80 ºC; חומרים כימיים אחרים ב -20 ºC). להפשיר אותם על הקרח ומערבבים את הרכיבים כפי שמוצג בטבלה 1.
| Mix-1 (תערובת האכלה) | |
| פתרון האכלה | 900 μl |
| חומצות אמינו | 80 μl |
| מתיונין | 20 μl |
| סה"כ | 1,000 μl |
| Mix-2 (תערובת Lysate) | |
| lysates wheatgerm | 15 μl |
| מים חינם RNase | 7 μl |
| חומצות אמינו | 4 μl |
| מתיונין | 1 μl |
| פלסמיד דנ"א במאגר TE (1 מיקרוגרם / μl) ** | 4 μl |
| מאגר תגובה | 15 μl |
| RNasin * | 4 μl |
| סה"כ | 50 μl |
טבלה 1: יצירות של תערובות נדרש לתגובות נבט חיטה.
הערה: אם הניסוי נועד לבחון את השפעת הסרת RNA על ידי RNase על Gag קרום מחייב, להחליף מעכבי ribonuclease עם מים RNase חינם. פלסמיד דנ"א צריך להיות מוגן מפני זיהומים על פי הוראה של היצרן. עם זאת, plasmids מוכן באמצעות קונבנציונאלי, אך לא רעלן פנימי ללא, ערכות בידוד פלסמיד שימשו בהצלחה. ההוראה של היצרן גם ממליצה להשתמש DNA מומס במים nuclease חינם. אם באמצעות DNA המרחפים TE [10 mM Tris-HCl (pH 7.4) המכיל 1 mM EDTA], להימנע משימוש ריכוזים נמוכים של דנ"א, מאז EDTA ב TE עשויים להשפיע על התשואות. פלסמידים מומס חיץ TE בטווח ריכוז של 0.8-1 מיקרוגרם / μl שימשו בהצלחה.
- ראשית להוסיף את תערובת ההאכלה (Mix-1) לתוך קלטת התגובה (תא שקוף). לאחר מכן מוסיפים את תערובת lysate (Mix-2) לתא אחר (אדום). אין להכניס כל בועות תוך הוספת התערובות לתאים שלהם, מאז שעלול להוביל חילופי יעילה של מרכיבי פתרון ולכן להפחית את היעילות של תגובה.
- מכסים לתאים עם הסרט הדבק מצורף לערכה.
- הכנס את קלטת התגובה לתוך מחזיק קלטת אינקובטוריםte האסיפה עבור 24 שעות ב 24 ºC במהירות טלטול מתמדת של 900 סל"ד בתוך Eppendorf thermomixer ר
יום 2: הכן GUVs ידי Electroformation קציר נבט חיטה Lysates
2. הכנת GUVs
- שומני Aliquot מומסים ממסים אורגניים כגון כלורופורם או תערובות של כלורופורם מתנול ב צלוחיות זכוכית עם פקקי הברגה מרופדים ספינות טפלון ועטפו עם Parafilm. טפל שומנים עם מזרקי זכוכית המילטון-סוג, או אם זמין, טיפי פלסטיק עמיד, כלורופורם. להוציא צלוחיות זכוכית המכילים ליפידים כדי לשמש להכנת Guv מהמקפיא -20 ºC.
- לאחר הצלוחיות equilibrated לטמפרטורת חדר, לוודא על ידי בדיקה ויזואלית כי השעיות השומנים ברורות. איכות שומנים, במיוחד שומנים חומצי, כגון phosphatidylserine Palmitoyl-oleoyl (POPS) ו PI (4,5) P 2, חשובה ניסויים מוצלחים. אין להשתמש שומנים כי הםב aliquots למשך זמן ארוך יותר מ -2 חודשים.
- טרום לחמם גוש חום לטמפרטורה הרצויה. טמפרטורה זו חייבת להיות לפחות 5 ºC גבוהה יותר מאשר טמפרטורת ההתכה (T מ) של שומני בדם (ים) עם T מ הגבוה ביותר.
- חשב את כמויות שומנים הדרושים על פי יחסי שומנים הרצויים הטוחנים. ללימוד HIV-1 Gag מחייב כאן, השתמש יחסי טוחנת הבאים כפי שמוצג בטבלה 2.
| תערובת של ליפידים | יחס טוחן | נפח (ב μl) | ריכוז מניות |
| POPC + פופס + חול | 46.6 + 23.3 + 30 | 19.74 + 10.18 + 6.57 | POPC, POPS, חול: 10 מ"ג / מ"ל |
| POPC + POPS + חול + Brain-PI (4,5) P 2 | 16,11 + 8,3 + 6,25 + 58,19 | POPC, POPS, חול: 10 מ"ג / מ"ל | |
| Brain-PI (4,5) P 2: 1 מ"ג / מ"ל |
טבלה 2: כרכים של שומנים שונים המשמשים להשיג GUVs.
- מוסיפים את בהיקפים הרצויים של שומנים לתוך בקבוקון זכוכית בורג הכתיר נקי.
- השתמש איטו מצופה שקופיות של ממדים 25 x 50 x 1.1 מ"מ 3 והתנגדות 70-100 Ω (כמתואר קטלוג). מניחים שתי שקופיות איטו מצופה על ספסל נקי. כל תא electroformation דורש שתי שקופיות זכוכית מצופות אינדיום-בדיל-תחמוצת (איטו). ודא שצד המוליך של שקופיות הזכוכית הוא פונה כלפי מעלה על ידי סימון ההתנגדות באמצעות מודד. זה צריך להראות שווי של כ -200 Ω. ודא פני השטח של השקופיות נקיים ידי מנגב בעדינות עם 70% אתנול באמצעות מגבונים ללא סיבים.
- נקה את המשטח החיצוני של מחט של מזרק על ידי ריnsing זה עם כלורופורם למקם את המזרק על הגוש החום.
- מקום שקופית זכוכית חדשה צופה איטו על הגוש החום עם צד מוליך פונה כלפי מעלה ולאפשר לו לאזן לטמפרטורה הרצויה בדרך כלל כ 2-3 דקות. אל תעשה שימוש חוזר בשקופיות זכוכית איטו מצופות עבור סיכון פוטנציאלי כי איטו-הציפוי על שקופיות הזכוכית עלול להינזק במהלך ניקוי נהלים.
- השתמש נפח של 40-50 μl של תערובת שומנים עבור כל שקופית עבור מדים ועם זאת מהיר מתפשטת של שומנים בשקופית זכוכית. התאם את הנפח הכולל עם ממיס אורגני מתאים כגון כלורופורם.
- מניחים מחצית הנפח הכולל של תערובת שומנים בשקופית זכוכית ומיד להפיץ את השומנים באמצעות המזרק ניקה (שלב 2.6) כדי משאירים ציפוי השומנים אחיד (הזזת המחט 2-3 פעמים ברחבי שקופיות). מורחים את הסרט השומנים בעדינות (אבל מהר מאוד), כדי למנוע כל נזק לציפוי איטו דק. לא אחיד מתפשט או מחוספס מתפשט עלולה להוביל תת אופטימליתתשואות או הטרוגניות חישובית של GUVs.
- לכן, להבטיח התפשטות אחידה על ידי בוחן סרט השומנים לאחר להתפשט לפני שתמשיך לשלב הבא. אם האטימות של הסרט מופיעות לא אחידות באופן מוגזם, שוב את ההליך באמצעות שקופית חדשה.
- מניחים את השקף מצופה בתוך צלחת פטרי עם הצד המצופה כלפי מעלה.
- חזור על שלבים 2.6-2.9 עבור שאר התערובת באמצעות אחר שקופית איטו מצופה.
- מניחים את צלחת פטרי המכילה גם את השקופיות בתא התייבשות ואקום קונבנציונאלי עבור 60-90 דקות כדי להסיר כל שמץ של ממיסים אורגניים.
- במהלך תהליך זה, להגדיר את החממה עד באותה הטמפרטורה כמו זה של הגוש החום (65 מעלות צלזיוס ברוב המקרים) וטרום-חם 300 מ"מ תמיסת סוכרוז בחממה.
- לאחר ייבוש, להביא את צלחת פטרי על הספסל במקום להחליק זכוכית על משטח נקי.
- לנקות בעדינות אטם PDMS (איור 1) באמצעות 70% אתנול ו משלים לו להתייבשtely.
- מניח אותו על שקופיות זכוכית מצופה שומנים כפי שמוצג באיור 1 בעדינות ובתקיפות ללחוץ כדי שצורות אטם חותם מים חזק. אשר בהעדר הפערים בין אטם PDMS ואת השקופיות. התוצאה היא היווצרות של תא רדוד שמכיל את תמיסת סוכרוז.

איור 1:. הפריסה של שקופית זכוכית מצופה איטו המשמש electroformation אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
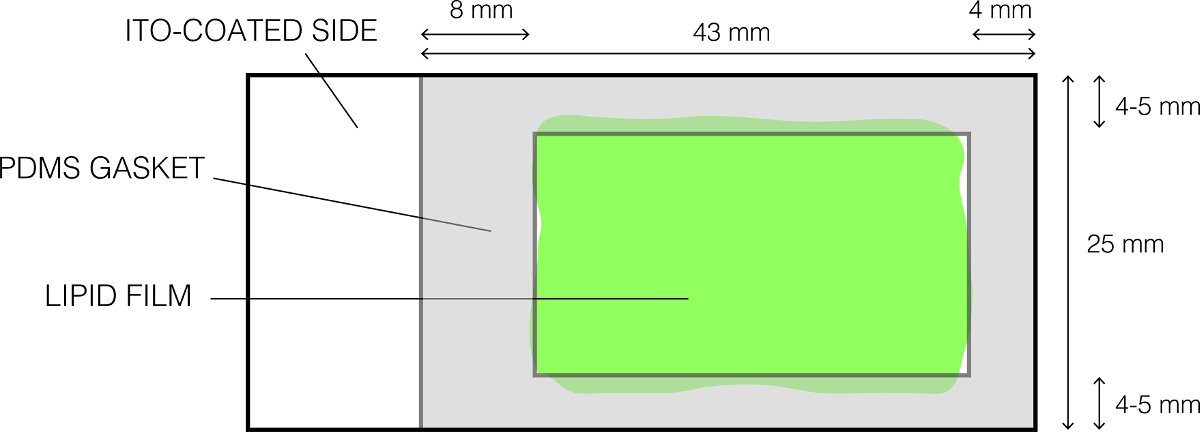{kind=link}
- ממלאים את החדר עם פתרון סוכרוז מחומם מראש ללא כל בועות.
- מניחים את השקף מצופים אחרים על העליונה כדי ליצור חותם חזק. ודא שאין בועות. ההרכבה הסופית אמורה להופיע כפי שמוצגת באיור 2. הדקי את usin הקאמריתקליפים קלסר גרם.

איור 2:. ייצוג סכמטי של תא electroformation (מבט מהצד) אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
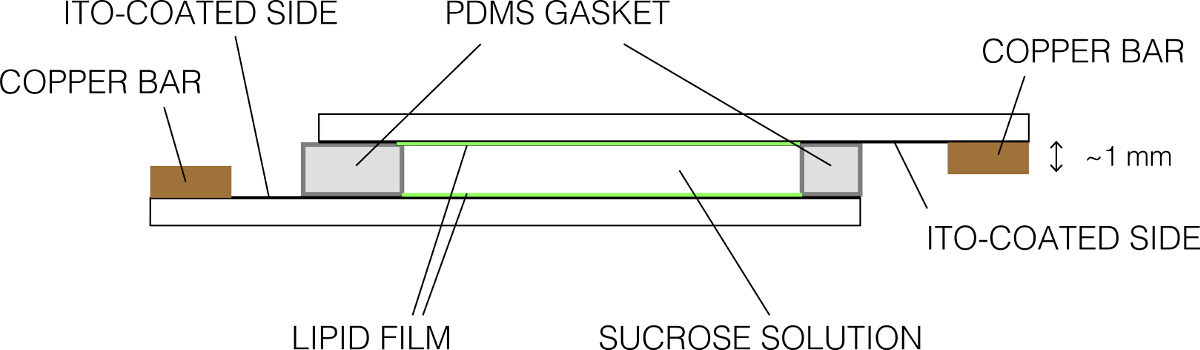{kind=link}
- הסר סוכרוז עודף ידי שאיפה ולנקות את השקופיות באמצעות מגבונים ללא סיבים. הדק את השקופיות לברות נחושת כדי שהצדדים המוליכים נמצאים בקשר אחיד עם הבר. ודא כי רק צד אחד מוליך של שקופיות הזכוכית נמצא בקשר עם בר נחושת אחד.
- חבר את הבר נחושת אל מוצא מחולל פונקציה באמצעות קליפים התנין. הנח את מכלול בתוך האינקובטור לאפשר הרכבה כדי לאזן את הטמפרטורה הרצויה.
- החל תדר גל סינוס של 10 הרץ ו פרש פוטנציאלים של 1 V עבור 90 דקות. ensurדואר שהמתח הוא 1 V ידי מודד כפי שמוצג באיור 3. משך התהליך במשך 90 דקות.

איור 3:. העמדות שבו אלקטרודות מן המודד ממוקם למדידת תדר וזרם אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
- לאחר 90 דקות, לאט להקטין את התדר עד 2 רץ ולהמשיך electroformation עוד 10 דקות. במהלך תקופה זו, קציר בתגובות תרגום במבחנה (שלבי 3.1-3.2).
- כבה את מחולל הפונקציה ולאפשר ההרכבה להתקרר לטמפרטורת חדר לאט (על-ידי כיבוי החממה ולהשאיר את דלת החממה נפתחה מעט).
- לאחר הקירור, לנתק את הכבלים מחוברים גנרטורמן המסילות. GUVs עשוי להיות מאוד שביר ולכן צריך להיות מטופלים בעדינות רבה. בזהירות להביא את המכלול לספסל.
- לפרק את החדר על ידי הסרת השקופית העליונה בעדינות בעזרת מלקחיים. קציר את ההשעיה סוכרוז המכיל GUVs באמצעות קצה לחתוך 1,000 μl ולהעביר צינור נקי. ידית הצינור בעדינות כדי למנוע שיבוש של GUVs.
- השתמש GUVs מיד. GUVs יציב במשך לפחות 90 דקות לאחר קציר כאשר לאחסן בטמפרטורת חדר.
3. תגובות תרגום חוץ גופית הקציר
- קציר את תגובות תרגום במבחנה המכילות חלבונים הרצוי מהאולם האדום באמצעות micropipette לתוך צינור.
- כדי להסיר חלבונים מצטברים, לסובב את lysates ב 16,200 XG במשך 15 דקות על בצנטריפוגה השולחן ובזהירות להעביר את supernatant לתוך צינור נוסף.
4. Guv Assay עקידת
- בעזרת קצה לחתוך, לערבב 51; l תגובות תרגום במבחנה המכילה חלבונים מתורגם ו -5 μl של GUVs בתוך שפופרת דגירה בטמפרטורת החדר למשך 2-3 דקות.
- צרף גיליון PDMS עם חור קטן (3-4 מ"מ קוטר) על פיסת כיסוי נקיה להבטיח חותם חזק על ידי בעדינות ובתקיפות מהדקים אותה אל coverslip.
- מניחים את התערובת 10 μl משלב 4.1 לתוך חור קטן.
- תמונת התערובת תחת מיקרוסקופ פלואורסצנטי עִלִית או confocal הפוך בטמפרטורת חדר.
הערה: כדי למנוע אידוי, לחדר ההדמיה יכול להיות מכוסה על ידי coverslip.
Access restricted. Please log in or start a trial to view this content.
תוצאות
שימוש בפרוטוקול לעיל, הכנו GUVs המורכב POPC + POPS + חול (יחס טוחן: 4.66: 2.33: 3). רכב זה נבחר כ לשקף את ריכוזי PS וכולסטרול של ראש הממשלה. יציב ויעיל מחייב של Gag נצפה רק כאשר המוח-PI (4,5) P 2 נכלל לתוך תערובת POPC + POPS + החול (POPC + POPS + חול + המוח-PI (4,5) P 2 [יחס טוחן :...
Access restricted. Please log in or start a trial to view this content.
Discussion
את assay מחייב Guv כמתואר לעיל מספק חלופה טובה בתרחישים בהם מבחני אינטראקציה חלבון-שומנים אחרים יש מגבלות שלהם. assay זה מאפשר לנו לבחון אינטראקציות בין Gag באורך מלא myristoylated ושומנים חומצי עם שרשרות acyl יליד באורך בהקשר השומנים bilayer ולעשות זאת בלי צנטריפוגה הנפקה ממושכת הדרך ?...
Access restricted. Please log in or start a trial to view this content.
Disclosures
We declare that we have no conflicting interests.
Acknowledgements
ברצוננו להודות מוחמד סלים, ג'ינג וו Krishnan Raghunathan לדיונים מועילים. אנו מודים גם פרייה Begani לסיוע במהלך הצילומים. עבודה זו נתמכת על ידי המכונים הלאומיים לבריאות מענקים R01 AI071727 (כדי AO) ו R01 GM110052 (כדי SLV).
Access restricted. Please log in or start a trial to view this content.
Materials
| Name | Company | Catalog Number | Comments |
| Digital Multimeter | Meterman | 30XR | A generic multimeter that can measure resistance and volts will serve the purpose. |
| Function Generator | Instek | GFG-8216A | A generic function generator that is capable of generating a sine wave at 10 Hz and 1 V is sufficient. |
| ITO coated glass slides | Delta Technologies, Loveland, CO | CG-90IN | |
| Incubator | Hoefer | Any incubator that can accurately maintain temperature will be sufficient | |
| Vacuum chamber | Nalgene | ||
| Thermomixer R | Eppendorf | 21516-166 | |
| Syringe | Hamilton | 80400 | Gauge 22S, Syringe number 702 |
| PDMS | Sylgard elastomer base kit, Dow-Corning | Sylgard, 184 | |
| RTS 100 Wheat Germ CECF Kit | BiotechRabbit, Berlin, Germany | BR1401001 | |
| DiD | Life Technologies, Carlsbad, CA | D7757 | |
| POPC | Avanti Polar Lipids, Alabaster, AL | 850457C | |
| POPS | Avanti Polar Lipids | 840034C | |
| Cholesterol | Avanti Polar Lipids | 700041P | |
| Brain-PI(4,5)P2 | Avanti Polar Lipids | 840046X |
References
- Chukkapalli, V., Inlora, J., Todd, G. C., Ono, A. Evidence in support of RNA-mediated inhibition of phosphatidylserine-dependent HIV-1 Gag membrane binding in cells. J Virol. 87 (12), 7155-7159 (2013).
- Chukkapalli, V., Oh, S. J., Ono, A. Opposing mechanisms involving RNA and lipids regulate HIV-1 Gag membrane binding through the highly basic region of the matrix domain. Proc Natl Acad Sci U S A. 107 (4), 1600-1605 (2010).
- Chukkapalli, V., Hogue, I. B., Boyko, V., Hu, W. S., Ono, A. Interaction between the human immunodeficiency virus type 1 Gag matrix domain and phosphatidylinositol-(4,5)-bisphosphate is essential for efficient gag membrane binding. J Virol. 82 (5), 2405-2417 (2008).
- Zhou, W., Parent, L. J., Wills, J. W., Resh, M. D. Identification of a membrane-binding domain within the amino-terminal region of human immunodeficiency virus type 1 Gag protein which interacts with acidic phospholipids. J Virol. 68 (4), 2556-2569 (1994).
- Ono, A., Ablan, S. D., Lockett, S. J., Nagashima, K., Freed, E. O. Phosphatidylinositol (4,5) bisphosphate regulates HIV-1 Gag targeting to the plasma membrane. Proc Natl Acad Sci U S A. 101 (41), 14889-14894 (2004).
- Saad, J. S., et al. Structural basis for targeting HIV-1 Gag proteins to the plasma membrane for virus assembly. Proc Natl Acad Sci U S A. 103 (30), 11364-11369 (2006).
- Shkriabai, N., et al. Interactions of HIV-1 Gag with assembly cofactors. Biochemistry. 45 (13), 4077-4083 (2006).
- Vlach, J., Saad, J. S. Trio engagement via plasma membrane phospholipids and the myristoyl moiety governs HIV-1 matrix binding to bilayers. Proc Natl Acad Sci U S A. 110 (9), 3525-3530 (2013).
- Anraku, K., et al. Highly sensitive analysis of the interaction between HIV-1 Gag and phosphoinositide derivatives based on surface plasmon resonance. Biochemistry. 49 (25), 5109-5116 (2010).
- Llewellyn, G. N., Grover, J. R., Olety, B., Ono, A. HIV-1 Gag associates with specific uropod-directed microdomains in a manner dependent on its MA highly basic region. J Virol. 87 (11), 6441-6454 (2013).
- Inlora, J., Chukkapalli, V., Derse, D., Ono, A. Gag localization and virus-like particle release mediated by the matrix domain of human T-lymphotropic virus type 1 Gag are less dependent on phosphatidylinositol-(4,5)-bisphosphate than those mediated by the matrix domain of HIV-1 Gag. J Virol. 85 (8), 3802-3810 (2011).
- Inlora, J., Collins, D. R., Trubin, M. E., Chung, J. Y., Ono, A. Membrane binding and subcellular localization of retroviral Gag proteins are differentially regulated by MA interactions with phosphatidylinositol-(4,5)-bisphosphate and RNA. MBio. 5 (6), e02202(2014).
- Valentine, K. G., et al. Reverse micelle encapsulation of membrane-anchored proteins for solution NMR studies. Structure. 18 (1), 9-16 (2010).
- Dick, R. A., Goh, S. L., Feigenson, G. W., Vogt, V. M. HIV-1 Gag protein can sense the cholesterol and acyl chain environment in model membranes. Proc Natl Acad Sci U S A. 109 (46), 18761-18766 (2012).
- Dick, R. A., Kamynina, E., Vogt, V. M. Effect of multimerization on membrane association of Rous sarcoma virus and HIV-1 MA proteins. J Virol. 87 (24), 13598-13608 (2013).
- Alfadhli, A., Still, A., Barklis, E. Analysis of human immunodeficiency virus type 1 matrix binding to membranes and nucleic acids. J Virol. 83 (23), 12196-12203 (2009).
- Rodriguez, N., Pincet, F., Cribier, S. Giant vesicles formed by gentle hydration and electroformation: a comparison by fluorescence microscopy. Colloids Surf B Biointerfaces. 42 (2), 125-130 (2005).
- Needham, D., McIntosh, T. J., Evans, E. Thermomechanical and transition properties of dimyristoylphosphatidylcholine/cholesterol bilayers. Biochemistry. 27 (13), 4668-4673 (1988).
- Darszon, A., et al. Reassembly of protein-lipid complexes into large bilayer vesicles: perspectives for membrane reconstitution. Proc Natl Acad Sci U S A. 77 (1), 239-243 (1980).
- Angelova, M. I., Dimitrov, D. S. Liposome electroformation. Faraday Discuss. Chem. Soc. (81), 303-311 (1986).
- Angelova, M. I., Soléau, S., Méléard, P. h, Faucon, F., Bothorel, P. Preparation of giant vesicles by external AC electric fields. Progr Colloid Polymer Sci. 89, 127-131 (1992).
- Stachowiak, J. C., et al. Unilamellar vesicle formation and encapsulation by microfluidic jetting. Proc Natl Acad Sci U S A. 105 (12), 4697-4702 (2008).
- Yamauchi, S., et al. The consensus motif for N-myristoylation of plant proteins in a wheat germ cell-free translation system. FEBS J. 277 (17), 3596-3607 (2010).
- Olety, B., Veatch, S. L., Ono, A. Phosphatidylinositol-(4,5)-Bisphosphate Acyl Chains Differentiate Membrane Binding of HIV-1 Gag from That of the Phospholipase Cdelta1 Pleckstrin Homology Domain. J Virol. 89 (15), 7861-7873 (2015).
- Morales-Penningston, N. F., et al. GUV preparation and imaging: minimizing artifacts. Biochim Biophys Acta. 1798 (7), 1324-1332 (2010).
- Veatch, S. L. Electro-formation and fluorescence microscopy of giant vesicles with coexisting liquid phases. Methods Mol Biol. 398, 59-72 (2007).
- Ayuyan, A. G., Cohen, F. S. Lipid peroxides promote large rafts: effects of excitation of probes in fluorescence microscopy and electrochemical reactions during vesicle formation. Biophys J. 91 (6), 2172-2183 (2006).
- Montes, L. R., et al. Electroformation of giant unilamellar vesicles from native membranes and organic lipid mixtures for the study of lipid domains under physiological ionic-strength conditions. Methods Mol Biol. 606, 105-114 (2010).
- Olety, B., Ono, A. Roles played by acidic lipids in HIV-1 Gag membrane binding. Virus Res. 193, 108-115 (2014).
- Keller, H., Krausslich, H. G., Schwille, P. Multimerizable HIV Gag derivative binds to the liquid-disordered phase in model membranes. Cell Microbiol. 15 (2), 237-247 (2013).
- Carlson, L. A., Hurley, J. H. In vitro reconstitution of the ordered assembly of the endosomal sorting complex required for transport at membrane-bound HIV-1 Gag clusters. Proc Natl Acad Sci U S A. 109 (42), 16928-16933 (2012).
- Gui, D., et al. A novel minimal in vitro system for analyzing HIV-1 Gag-mediated budding. J Biol Phys. 41 (2), 135-149 (2015).
- Prevost, C., et al. IRSp53 senses negative membrane curvature and phase separates along membrane tubules. Nat Commun. 6, 8529(2015).
Access restricted. Please log in or start a trial to view this content.
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved