
このコンテンツを視聴するには、JoVE 購読が必要です。 サインイン又は無料トライアルを申し込む。
Method Article
巨大単層ベシクル(GUV)膜への結合HIV-1のGagの可視化
要約
We illustrate here an in vitro membrane binding assay in which interactions between HIV-1 Gag and lipid membranes are visually analyzed using YFP-tagged Gag synthesized in a wheat germ-based in vitro translation system and GUVs prepared by an electroformation technique.
要約
HIV-1の構造タンパク質、Pr55 のGag(またはギャグ)は、ウイルスの組立プロセス中に細胞内で原形質膜に結合します。ギャグの膜結合は、ウイルス粒子産生の重度の障害でギャグ膜結合結果の欠陥以来、ウイルス粒子形成に必須の工程です。ギャグ脂質膜との相互作用のメカニズムの詳細は、NMRに基づくインビトロ方法を得るために、タンパク質フットプリント、表面プラズモン共鳴、リポソーム浮遊遠心分離、又は蛍光脂質ビーズ結合は、これまで開発されてきました。しかし、これらのin vitroの方法の各々は、その限界を有します。これらの制限の一部を克服し、以前に確立された方法に相補的なアプローチを提供するために、我々は、 インビトロアッセイを開発したHIV-1 Gagおよび脂質膜との相互作用」とは、細胞のような」環境で行われます。このアッセイでは、脂質膜に結合したGag目視YFP-taggeを用いて分析しますDのギャグは、電鋳技術により調製インビトロ翻訳システムとGUVs に小麦ベースの胚芽で合成します。ここでは、アッセイのために必要なミリストイル化完全長Gagタンパク質とGUV膜を得るために、およびGag-GUVは顕微鏡によって結合を検出するために、背景とプロトコルについて説明します。
概要
ヒト免疫不全ウイルス1型(HIV-1)は、組み立ておよび大部分の細胞型における原形質膜(PM)から芽エンベロープウイルスです。 HIV-1ウイルス粒子のアセンブリは、Pr55 ギャグ (GAG)と呼ばれる55 kDaのウイルスコアタンパク質によって駆動されます。ギャグは、4つの主要な構造ドメイン、すなわち、マトリックス、カプシド、ヌクレオカプシド、およびP6、ならびに2のスペーサーペプチドSP1およびSP2で構成される前駆体ポリタンパク質として合成されます。組み立て中、マトリックス(MA)ドメインは、組立現場にギャグのターゲティングのために責任がある、カプシド(CA)ドメインはギャグ・ギャグの相互作用を媒介する、ヌクレオカプシド(NC)ドメインは、ウイルスゲノムRNAを募集し、p6の新兵の宿主因子を助けます原形質膜からのウイルス粒子の切断。 GAGはまた、N末端に14炭素脂肪酸またはミリスチン部分の添加により、共翻訳後修飾を受けます。
ギャグの結合膜はメートル以来、ウイルスのアセンブリのために不可欠な要件でありますウイルス粒子を産生しない膜結合に欠陥があるutants。我々と他の人はギャグの結合膜はMAドメイン内の二部の信号によって媒介されることが示されている:脂質二重層や高塩基性領域(と呼ばMAドメイン内の塩基性残基のクラスターとの疎水性相互作用を媒介N末端ミリスチン酸部分PM 1-4上の酸性脂質と相互作用HBR)。ポリリン酸化イノシチド5-ホスファターゼIV(5ptaseIV)、PM-特定の酸性リン脂質ホスファチジルの加水分解を触媒する酵素の異所性発現を使用してギャグ-膜の相互作用の研究(4,5)-bisphosphate [PI(4,5)P 2]ホスファチジルイノシトール-4-ホスフェートに、細胞中のGag-PMの局在をPI(4,5)P 2 3,5によって媒介されることを示唆しました。しかし、in vitroでの研究はギャグ-PI(4,5)Pの複数のメカニズムの詳細を理解することを目指して2相互作用は、多くの理由のために挑戦することが証明されています。ために生化学的実験のために、例えば、完全長ミリストイル化HIV-1の精製ギャグが原因精製中に凝集するギャグの傾向に少なくとも部分的には技術的に困難でした。したがって、このようなマイア-MAまたはマイア-MA-CA、または非ミリストイル化形態のようなHIV-1のGagの切断型は、しばしばギャグ精製( 例えば 、核磁気共鳴、タンパク質フットプリント、および表面を必要とする研究で使用されていますプラズモン共鳴6-9)。あるいは、 インビトロ転写-翻訳反応において結合された他の生化学的研究1,2-における全長ミリストイル化HIV-1のGagを生成するために使用されてきました。典型的には、このシステムでは、ギャグをコードするプラスミドは、任意の細胞膜とメッセンジャーRNAを欠いているが、転写および翻訳のための機械が含まれている真核生物の細胞溶解物( 例えば 、ウサギ網状赤血球溶解物)で転写および翻訳されます。反応後、ギャグを含む細胞溶解物を私と一緒に混合されます脂質とギャグの相互作用を分析するためのmbranes。ギャグミリストイル全長の調製の容易さに加えて、in vitro転写翻訳系を使用する方法は、GAG合成およびその後の膜結合反応は、より良い生理的条件を表すことができる「真核生物の細胞質ゾルのような」環境で発生するという利点を有します。このプロパティは、MAドメインに結合したRNA分子は競合的1,2,10-12に酸性脂質に結合するギャグを調節することを示した研究に貢献しました。これらの細胞溶解物で得られたGagタンパク質の合計量は高くないので、放射性標識アミノ酸を有するタンパク質の代謝標識は、それらの検出のために必要です。
ギャグ - 脂質相互作用を測定する方法に依存して、膜調製物の様々な使用されてきました。これらの各メソッドは、その長所と短所があります。ほとんどのNMRベースのアッセイは、短いアシルと脂質の使用を必要とします水溶性である鎖( 例えば 、C4-およびC8-PI(4,5)P 2)6,8。細胞に見られる長いアシル鎖を有する脂質へのギャグの結合を試験するためのNMR法が開発されているが、それらはこれまで8,13のみミリストイルまたはnonmyristoylated MAで使用されています。あるいは、天然の長さのアシル鎖を有する脂質から調製されたリポソームは、リポソーム浮遊または蛍光リポソームビーズ結合アッセイ2,3,10,14-16などの生化学的方法に使用されてきました。しかしながら、これらのアッセイにおいて使用されるリポソームは、小さな直径を有し、従って、それらの膜は、急な正の曲率を有しています。対照的に、HIV-1感染細胞における粒子アセンブリの初期段階の間、ギャグほぼたGagの規模で平坦であるPMに結合し、続いて出芽中負の曲率を誘導します。そのため、急な正の曲率を有するリポソーム膜はギャグ - 脂質相互作用を研究するための理想的な脂質二重層ではないかもしれません。リポソームFLOTについてエーションアッセイ、別の潜在的な注意点は、実験結果に影響を与える可能性が遠心分離中高張ショ糖勾配にギャグ脂質複合体の露出があります。これらの制限を緩和し、補完的な実験システムを提供すること、ギャグ巨大単層小胞(GUVs)への結合についてのアッセイは、近年開発されてきました。 GUVsは、その直径が数十μmまで延びる単一の脂質二重層小胞です。したがって、これらの膜の曲率は、ギャグの規模でPMに似ています。また、原因光学顕微鏡下で目視検査を可能にし、その大きなサイズに、混合時にこれらの小胞への蛍光タグ付きまたは標識Gagタンパク質を結合膜が容易にギャグ - 脂質複合体のその後の処理を行わずに決定することができます。
ここではエレクトロフォーメーション法から得られたGUVsを使用して結合HIV-1のGag膜を研究するためのプロトコルについて説明します。このような穏やかな水和、ゲルアシスト水和、マイクなどの様々な方法rofluidic噴射、及び電鋳17-22 GUVsを取得するために使用されてきました。ここで説明されたプロトコルのために、エレクトロフォーメーション法は、高価なセットアップを必要とせずに、酸性脂質およびその使用比較的容易にGUVsを形成する際に、その効率の主に使用されます。ギャグの可視化は、蛍光レポーターを必要とするので、黄色蛍光タンパク質(YFP)は、遺伝的にギャグ(GAG-YFP)のC末端に付加されています。ギャグ-YFPタンパク質は連続交換-連続フロー(CECF)技術に基づいた小麦胚芽溶解物におけるインビトロ転写および翻訳反応によって得られます。この技術では、反応は、反応基質およびエネルギー成分の供給の阻害副産物の両方の除去は、透析に基づく機構で達成されます。これらの反応のために、T7プロモーターの制御下のGag-YFPをコードするプラスミドを用いています。先に示したように注目すべきは、小麦胚芽溶解物は、追加のコンポーネント2なしミリストイル化をサポート3,24。この方法を使用して、GUV膜24上のGagの可視化のためのGag-YFPミリストイル全長の十分な量を得ることができました。ここでは、PIに結合したGag(4,5)P 2を含むGUV膜が結合反応、次の長い以降の処理を行うことなく検査し、この方法はギャグ-膜結合アッセイを既存の補完とすることができることを提案することができるHIV-1でのプロトコルを記述しますさらに、HIV-1のGag-膜結合を理解するように拡張。
Access restricted. Please log in or start a trial to view this content.
プロトコル
1日目:小麦胚芽溶解物ベースのin vitro転写・翻訳システムを用いたGagタンパク質の発現
HIV-1のGagの調製
- 冷凍庫から商業小麦胚芽CECFキットの全ての試薬を取り外します(小麦胚芽溶解物を-80ºCで保存されている。-20ºCで他の試薬)。氷の上で解凍し、 表1に示す成分を混合。
| ミックス-1(摂食ミックス) | |
| 供給溶液 | 900μlの |
| アミノ酸 | 80μlの |
| メチオニン | 20μlの |
| 合計 | 千μlの |
| ミックス-2(溶解物ミックス) | |
| 小麦胚芽溶解物 | 15μlの |
| RNaseを含まない水 | 7μlの |
| アミノ酸 | 4μlの |
| メチオニン | 1μlの |
| TE緩衝液中のプラスミドDNA(1μgの/μl)を** | 4μlの |
| 反応緩衝液 | 15μlの |
| RNasin * | 4μlの |
| 合計 | 50μlの |
表1:コムギ胚芽反応に必要な混合物の組成物。
注:この実験は、GAG結合膜上のRNaseによるRNA除去の効果を調べるために設計されている場合は、RNaseフリー水でリボヌクレアーゼ阻害剤を交換してください。プラスミドDNAを、製造業者の指示に従って不純物から自由でなければなりません。しかし、Plasmids従来のではなく、エンドトキシンフリーを用いて調製し、プラスミド単離キットが正常に使用されてきました。製造元の指示はまた、ヌクレアーゼフリー水に溶解したDNAを使用することをお勧めします。 TE [1mMのEDTAを含む10mMトリス-HCl(pH7.4)]中に懸濁したDNAを使用する場合、TEにおけるEDTAの歩留まりに影響を与える可能性があるため、DNAの低濃度の使用を避けます。 0.8から1μgの/μlの濃度範囲でのTE緩衝液に溶解したプラスミドは、成功裡に使用されています。
- 最初の反応カセット(透明室)に供給混合物(ミックス-1)を追加します。その後、他のチャンバ(赤)に溶解液ミックス(ミックス-2)を追加します。それぞれの室に混合物を添加しながらそれは溶液成分の非効率的な交流につながるため、反応の効率を低下させる可能性があるため、任意の気泡を導入しないでください。
- キットに付属の接着フィルム付き室をカバーしています。
- カセットホルダーとincubaに反応カセットを挿入しますエッペンドルフサーモミキサーRに900rpmでの一定の振とう速度で24ºCで24時間のアセンブリをTE
2日目:電鋳と収穫小麦胚芽ライセートによってGUVsを準備
GUVsの調製
- このようなスクリューキャップテフロンライナーでライニングし、パラフィルムで包まれたガラスバイアル中のクロロホルムとメタノールのクロロホルムまたはこれらの混合物などの無機溶媒に溶解したアリコート脂質。ハミルトン型ガラスシリンジ、または利用可能な場合、クロロホルム-性プラスチックのヒントと脂質を扱います。 -20ºCの冷凍庫からGUVの調製のために使用される脂質を含有するガラスバイアルを取り出し。
- バイアルを室温に平衡化されると、脂質懸濁液が明確であることを目視により確認してください。脂質の品質、このようなパルミトイル-オレオイルホスファチジルセリン(POPS)とPI(4,5)P 2として特に酸性脂質は、成功した実験のために重要です。ある脂質を使用しないでくださいより長い2ヶ月アリコートインチ
- 所望の温度に加熱ブロックを事前に温めます。この温度は、最も高いT mを有する脂質(複数可)の融解温度(T m)よりも少なくとも5ºC高くなければなりません。
- 所望のモル脂質比に応じて必要な脂質量を計算します。 表2に示すように、ここで結合HIV-1のGagを研究するために、以下のモル比を使用します。
| 脂質ミックス | モル比 | 容量(μL中) | ストック濃度 |
| POPC + POPS +チョル | 46.6 + 23.3 + 30 | 19.74 + 10.18 + 6.57 | POPC、POPS、チョル:10 mg / mlで |
| POPC + POPS +チョル+脳-PI(4,5)P 2 | 16.11 + 8.3 + 6.25 + 58.19 | POPC、POPS、チョル:10 mg / mlで | |
| 脳-PI(4,5)P 2:1mg / mlの |
表2:異なる脂質のボリュームはGUVsを得るために使用されます。
- きれいなスクリューキャップガラスバイアル中の脂質の所望のボリュームを追加します。
- 寸法25×50×1.1ミリメートル3と抵抗70から100Ω(カタログに記載されているように)の使用ITO被覆スライド。クリーンベンチ上の2つのITO被覆スライドを置きます。各エレクトロフォーメーションチャンバーは、二つのインジウムスズ酸化物(ITO)被覆ガラススライドを必要とします。スライドガラスの導電側はマルチメータを使用して抵抗をチェックすることにより、上を向いていることを確認します。これは、約200Ωの値を表示する必要があります。スライドの表面をやさしくリントフリーワイプを用いて70%エタノールで拭いてきれいであることを確認します。
- 里によって注射器の針の外面を清掃してくださいクロロホルムでそれをnsingし、ヒートブロック上に注射器を置きます。
- 上を向いて導電側をヒートブロック上に新しいITO被覆ガラススライドを置き、それは通常、約2〜3分所望の温度に平衡化することができます。ガラススライド上のITOコーティングが手続きを洗浄中に破損する可能性がある潜在的なリスクのためのITO被覆ガラススライドを再利用しないでください。
- 均一でスライドガラス上の脂質の広がりはまだ迅速なため、各スライドの脂質混合物の40〜50マイクロリットルの容量を使用してください。クロロホルムなどの適当な有機溶媒との総容量を調整します。
- スライドガラス上の脂質混合物の総体積の半分を置き、すぐに均一な脂質フィルム(スライド全体に針2-3回の移動)を残すために洗浄シリンジ(ステップ2.6)を使用して、脂質を広げます。薄いITOコーティングへの損傷を避けるために穏やかに脂質フィルムを広げて(ただし迅速に)。不均一な拡散またはサブ最適につながる可能性が広がるラフ利回りまたはGUVsの計算異質。
- そのため、後の次のステップに進む前に広がる脂質フィルムを検査することにより、均一な広がりを確保します。フィルムの不透明度が過度に不均一に表示された場合は、新しいスライドを使用して手順をやり直し。
- コーティングされた側を上に向けてペトリ皿の内側コートスライドを置きます。
- 別のITO被覆スライドを用いて混合物の残りの手順を2.6から2.9を繰り返します。
- 有機溶剤の痕跡を除去するために、60から90分間、従来の真空乾燥室でスライドの両方を含むペトリ皿を置きます。
- このプロセスの間に、ヒートブロック(ほとんどの場合、65ºC)とプリ暖かい300mMのインキュベーター中のスクロースの溶液と同じ温度にインキュベーターを設定します。
- 乾燥させた後、ベンチにペトリ皿を持参し、清潔な表面上にスライドガラスを配置します。
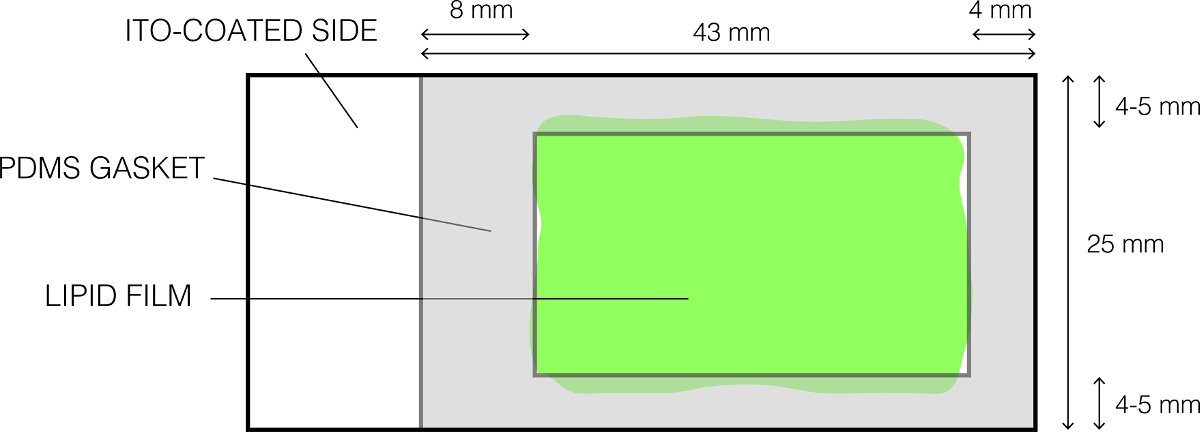
- 静かに70%エタノールとドライそれのcompleを使用して、PDMSガスケット( 図1)をきれいにtely。
- 図1に示すように、脂質コーティングされたガラススライド上に置き、ゆっくりとしっかりとガスケットは水密シールを形成するように押してください。 PDMSガスケットとスライドとの間にギャップが存在しないことを確認してください。これは、蔗糖液を保持している浅い室が形成されます。

図1:電鋳のために使用されるITO被覆ガラススライドのレイアウトは、 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
- 気泡のない予熱したショ糖溶液でチャンバーを埋めます。
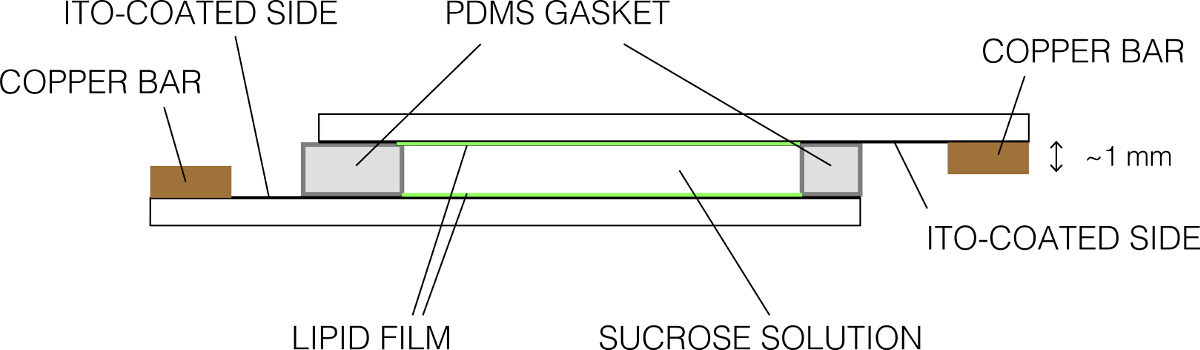
- 気密シールを作るために上に他のコーティングされたスライドを配置します。気泡がないことを確認してください。 図2に示すように、最終的な組み立てが表示されます。チャンバーusinを固定しますGバインダークリップ。

図2:エレクトロフォーメーション室(側面図)の概略図 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
- 吸引により過剰のスクロースを取り外し、リントフリーワイプを使用してスライドを清掃してください。導電性の側面では、バーと均一に接触するように銅バーにスライドを固定します。ガラススライドの一方のみ導電側は1つの銅棒に接触していることを確認してください。
- ワニ口クリップを使用して、関数発生器の出力に銅バーを接続します。アセンブリが所望の温度に平衡化することを可能にするインキュベーターの内側にアセンブリを配置します。
- 10ヘルツの正弦波周波数と90分間、1 Vの電位差を適用します。 Ensur図3に示すように、電圧をマルチメータにより1 Vであるeが。90分間の処理を続行します。

図3:マルチメーターの電極は周波数と電流を測定するために配置された位置 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
- 90分後、ゆっくりと2 Hzまでの周波数を減少させ、さらに10分間電鋳を続けます。この間、 インビトロ翻訳反応における収穫は(3.1から3.2ステップ)。
- 関数発生器のスイッチをオフにし、アセンブリは(インキュベーターをオフにすると、わずかに開いたインキュベータのドアを残して)ゆっくりと室温まで冷却することができます。
- 冷却した後、発電機にケーブルを外しスライドアセンブリから。 GUVsは非常に壊れやすいこと、したがって、非常に穏やかに処理する必要があります。慎重にベンチにアセンブリをもたらします。
- 優しく鉗子を使用して、トップスライドを除去することにより、チャンバーを分解します。きれいなチューブにカット千μlの先端と転送を使用して、スクロースを含む懸濁液GUVsを収穫。 GUVsの破壊を防止するために、穏やかにチューブを扱います。
- すぐにGUVsを使用してください。室温で保存した場合GUVs収穫後少なくとも90分間安定です。
3.収穫インビトロ翻訳反応
- チューブにマイクロピペットを用いて赤チャンバーから所望のタンパク質を含むインビトロ翻訳反応を収穫。
- 、凝集したタンパク質を除去卓上遠心機で15分間、16,200×gで溶解物をスピンし、慎重に別のチューブに上清を転送します。
4. GUV結合アッセイ
- 切断されたチップを用いて、5を混在1;チューブに翻訳されたタンパク質とGUVsの5μLを含むin vitro翻訳反応のリットルと2-3分間室温でインキュベートします。
- きれいなカバースリップ上の小さな穴(3-4ミリの直径)とPDMSシートを取り付け、静かに、しっかりとカバーガラスに対してそれを押すことによって、気密シールを確保します。
- 小さな穴にステップ4.1から10μlの混合物を配置します。
- 画像室温で反転エピまたは共焦点蛍光顕微鏡下で混合物。
注:蒸発を防止するために、撮像チャンバは、カバースリップで覆うことができます。
Access restricted. Please log in or start a trial to view this content.
結果
上記のプロトコルを使用して、我々は、POPC + POPS +チョルから成るGUVsを用意し(モル比:4.66:2.33:3)。この組成物は、約PMのPSおよびコレステロール濃度を反映するために選ばれました。ギャグの堅牢かつ効率的な結合は、脳-PI(4,5)P 2は、POPC + POPS + Cholの混合物(POPC + POPS +チョル+脳-PI(4,5)P 2 [モル比に含まれていた場合にのみ観察されまし?...
Access restricted. Please log in or start a trial to view this content.
ディスカッション
上記のようにGUV結合アッセイは、他のタンパク質 - 脂質相互作用アッセイは、その限界を持っているシナリオでは良い代替手段を提供します。このアッセイは、私たちは、脂質二重層の文脈における天然の長さのアシル鎖を有するミリストイル化完全長Gagおよび酸性脂質間の相互作用を検討およびGag-脂質の高密度ショ糖勾配または他のポスト結合処理により長い浮上遠心分離することな...
Access restricted. Please log in or start a trial to view this content.
開示事項
We declare that we have no conflicting interests.
謝辞
私たちは便利な議論のためのモハンマドサリーム、ジン呉とクリシュナンRaghunathanに感謝したいと思います。また、撮影中に支援するためのプリヤBeganiに感謝します。この作品は、国立衛生研究所R01 AI071727(AO)と(SLV)はR01 GM110052を付与することによってサポートされています。
Access restricted. Please log in or start a trial to view this content.
資料
| Name | Company | Catalog Number | Comments |
| Digital Multimeter | Meterman | 30XR | A generic multimeter that can measure resistance and volts will serve the purpose. |
| Function Generator | Instek | GFG-8216A | A generic function generator that is capable of generating a sine wave at 10 Hz and 1 V is sufficient. |
| ITO coated glass slides | Delta Technologies, Loveland, CO | CG-90IN | |
| Incubator | Hoefer | Any incubator that can accurately maintain temperature will be sufficient | |
| Vacuum chamber | Nalgene | ||
| Thermomixer R | Eppendorf | 21516-166 | |
| Syringe | Hamilton | 80400 | Gauge 22S, Syringe number 702 |
| PDMS | Sylgard elastomer base kit, Dow-Corning | Sylgard, 184 | |
| RTS 100 Wheat Germ CECF Kit | BiotechRabbit, Berlin, Germany | BR1401001 | |
| DiD | Life Technologies, Carlsbad, CA | D7757 | |
| POPC | Avanti Polar Lipids, Alabaster, AL | 850457C | |
| POPS | Avanti Polar Lipids | 840034C | |
| Cholesterol | Avanti Polar Lipids | 700041P | |
| Brain-PI(4,5)P2 | Avanti Polar Lipids | 840046X |
参考文献
- Chukkapalli, V., Inlora, J., Todd, G. C., Ono, A. Evidence in support of RNA-mediated inhibition of phosphatidylserine-dependent HIV-1 Gag membrane binding in cells. J Virol. 87 (12), 7155-7159 (2013).
- Chukkapalli, V., Oh, S. J., Ono, A. Opposing mechanisms involving RNA and lipids regulate HIV-1 Gag membrane binding through the highly basic region of the matrix domain. Proc Natl Acad Sci U S A. 107 (4), 1600-1605 (2010).
- Chukkapalli, V., Hogue, I. B., Boyko, V., Hu, W. S., Ono, A. Interaction between the human immunodeficiency virus type 1 Gag matrix domain and phosphatidylinositol-(4,5)-bisphosphate is essential for efficient gag membrane binding. J Virol. 82 (5), 2405-2417 (2008).
- Zhou, W., Parent, L. J., Wills, J. W., Resh, M. D. Identification of a membrane-binding domain within the amino-terminal region of human immunodeficiency virus type 1 Gag protein which interacts with acidic phospholipids. J Virol. 68 (4), 2556-2569 (1994).
- Ono, A., Ablan, S. D., Lockett, S. J., Nagashima, K., Freed, E. O. Phosphatidylinositol (4,5) bisphosphate regulates HIV-1 Gag targeting to the plasma membrane. Proc Natl Acad Sci U S A. 101 (41), 14889-14894 (2004).
- Saad, J. S., et al. Structural basis for targeting HIV-1 Gag proteins to the plasma membrane for virus assembly. Proc Natl Acad Sci U S A. 103 (30), 11364-11369 (2006).
- Shkriabai, N., et al. Interactions of HIV-1 Gag with assembly cofactors. Biochemistry. 45 (13), 4077-4083 (2006).
- Vlach, J., Saad, J. S. Trio engagement via plasma membrane phospholipids and the myristoyl moiety governs HIV-1 matrix binding to bilayers. Proc Natl Acad Sci U S A. 110 (9), 3525-3530 (2013).
- Anraku, K., et al. Highly sensitive analysis of the interaction between HIV-1 Gag and phosphoinositide derivatives based on surface plasmon resonance. Biochemistry. 49 (25), 5109-5116 (2010).
- Llewellyn, G. N., Grover, J. R., Olety, B., Ono, A. HIV-1 Gag associates with specific uropod-directed microdomains in a manner dependent on its MA highly basic region. J Virol. 87 (11), 6441-6454 (2013).
- Inlora, J., Chukkapalli, V., Derse, D., Ono, A. Gag localization and virus-like particle release mediated by the matrix domain of human T-lymphotropic virus type 1 Gag are less dependent on phosphatidylinositol-(4,5)-bisphosphate than those mediated by the matrix domain of HIV-1 Gag. J Virol. 85 (8), 3802-3810 (2011).
- Inlora, J., Collins, D. R., Trubin, M. E., Chung, J. Y., Ono, A. Membrane binding and subcellular localization of retroviral Gag proteins are differentially regulated by MA interactions with phosphatidylinositol-(4,5)-bisphosphate and RNA. MBio. 5 (6), e02202(2014).
- Valentine, K. G., et al. Reverse micelle encapsulation of membrane-anchored proteins for solution NMR studies. Structure. 18 (1), 9-16 (2010).
- Dick, R. A., Goh, S. L., Feigenson, G. W., Vogt, V. M. HIV-1 Gag protein can sense the cholesterol and acyl chain environment in model membranes. Proc Natl Acad Sci U S A. 109 (46), 18761-18766 (2012).
- Dick, R. A., Kamynina, E., Vogt, V. M. Effect of multimerization on membrane association of Rous sarcoma virus and HIV-1 MA proteins. J Virol. 87 (24), 13598-13608 (2013).
- Alfadhli, A., Still, A., Barklis, E. Analysis of human immunodeficiency virus type 1 matrix binding to membranes and nucleic acids. J Virol. 83 (23), 12196-12203 (2009).
- Rodriguez, N., Pincet, F., Cribier, S. Giant vesicles formed by gentle hydration and electroformation: a comparison by fluorescence microscopy. Colloids Surf B Biointerfaces. 42 (2), 125-130 (2005).
- Needham, D., McIntosh, T. J., Evans, E. Thermomechanical and transition properties of dimyristoylphosphatidylcholine/cholesterol bilayers. Biochemistry. 27 (13), 4668-4673 (1988).
- Darszon, A., et al. Reassembly of protein-lipid complexes into large bilayer vesicles: perspectives for membrane reconstitution. Proc Natl Acad Sci U S A. 77 (1), 239-243 (1980).
- Angelova, M. I., Dimitrov, D. S. Liposome electroformation. Faraday Discuss. Chem. Soc. (81), 303-311 (1986).
- Angelova, M. I., Soléau, S., Méléard, P. h, Faucon, F., Bothorel, P. Preparation of giant vesicles by external AC electric fields. Progr Colloid Polymer Sci. 89, 127-131 (1992).
- Stachowiak, J. C., et al. Unilamellar vesicle formation and encapsulation by microfluidic jetting. Proc Natl Acad Sci U S A. 105 (12), 4697-4702 (2008).
- Yamauchi, S., et al. The consensus motif for N-myristoylation of plant proteins in a wheat germ cell-free translation system. FEBS J. 277 (17), 3596-3607 (2010).
- Olety, B., Veatch, S. L., Ono, A. Phosphatidylinositol-(4,5)-Bisphosphate Acyl Chains Differentiate Membrane Binding of HIV-1 Gag from That of the Phospholipase Cdelta1 Pleckstrin Homology Domain. J Virol. 89 (15), 7861-7873 (2015).
- Morales-Penningston, N. F., et al. GUV preparation and imaging: minimizing artifacts. Biochim Biophys Acta. 1798 (7), 1324-1332 (2010).
- Veatch, S. L. Electro-formation and fluorescence microscopy of giant vesicles with coexisting liquid phases. Methods Mol Biol. 398, 59-72 (2007).
- Ayuyan, A. G., Cohen, F. S. Lipid peroxides promote large rafts: effects of excitation of probes in fluorescence microscopy and electrochemical reactions during vesicle formation. Biophys J. 91 (6), 2172-2183 (2006).
- Montes, L. R., et al. Electroformation of giant unilamellar vesicles from native membranes and organic lipid mixtures for the study of lipid domains under physiological ionic-strength conditions. Methods Mol Biol. 606, 105-114 (2010).
- Olety, B., Ono, A. Roles played by acidic lipids in HIV-1 Gag membrane binding. Virus Res. 193, 108-115 (2014).
- Keller, H., Krausslich, H. G., Schwille, P. Multimerizable HIV Gag derivative binds to the liquid-disordered phase in model membranes. Cell Microbiol. 15 (2), 237-247 (2013).
- Carlson, L. A., Hurley, J. H. In vitro reconstitution of the ordered assembly of the endosomal sorting complex required for transport at membrane-bound HIV-1 Gag clusters. Proc Natl Acad Sci U S A. 109 (42), 16928-16933 (2012).
- Gui, D., et al. A novel minimal in vitro system for analyzing HIV-1 Gag-mediated budding. J Biol Phys. 41 (2), 135-149 (2015).
- Prevost, C., et al. IRSp53 senses negative membrane curvature and phase separates along membrane tubules. Nat Commun. 6, 8529(2015).
Access restricted. Please log in or start a trial to view this content.
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved