
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
Visualization of HIV-1 Gag Binding to Giant Unilamellar Vesicle (GUV) Membranes
In This Article
Summary
We illustrate here an in vitro membrane binding assay in which interactions between HIV-1 Gag and lipid membranes are visually analyzed using YFP-tagged Gag synthesized in a wheat germ-based in vitro translation system and GUVs prepared by an electroformation technique.
Abstract
The structural protein of HIV-1, Pr55Gag (or Gag), binds to the plasma membrane in cells during the virus assembly process. Membrane binding of Gag is an essential step for virus particle formation, since a defect in Gag membrane binding results in severe impairment of viral particle production. To gain mechanistic details of Gag-lipid membrane interactions, in vitro methods based on NMR, protein footprinting, surface plasmon resonance, liposome flotation centrifugation, or fluorescence lipid bead binding have been developed thus far. However, each of these in vitro methods has its limitations. To overcome some of these limitations and provide a complementary approach to the previously established methods, we developed an in vitro assay in which interactions between HIV-1 Gag and lipid membranes take place in a "cell-like" environment. In this assay, Gag binding to lipid membranes is visually analyzed using YFP-tagged Gag synthesized in a wheat germ-based in vitro translation system and GUVs prepared by an electroformation technique. Here we describe the background and the protocols to obtain myristoylated full-length Gag proteins and GUV membranes necessary for the assay and to detect Gag-GUV binding by microscopy.
Introduction
Human immunodeficiency virus type 1 (HIV-1) is an enveloped virus that assembles at and buds from the plasma membrane (PM) in most cell types. The assembly of HIV-1 virus particles is driven by the 55 kDa viral core protein called Pr55Gag (Gag). Gag is synthesized as a precursor polyprotein composed of four major structural domains, namely, matrix, capsid, nucleocapsid, and p6, as well as two spacer peptides SP1 and SP2. During assembly, the matrix (MA) domain is responsible for targeting of Gag to the assembly site, the capsid (CA) domain mediates Gag-Gag interactions, the nucleocapsid (NC) domain recruits viral genomic RNA, and p6 recruits host factors that aid virus particle scission from the plasma membrane. Gag also undergoes a co-translational modification by the addition of a 14-carbon fatty acid or myristate moiety at its N-terminus.
Membrane binding of Gag is an essential requirement for the viral assembly, since mutants that are defective in membrane binding fail to produce virus particles. We and others have shown that membrane binding of Gag is mediated by bipartite signals within the MA domain: the N-terminal myristate moiety that mediates hydrophobic interactions with the lipid bilayer and a cluster of basic residues within the MA domain termed as highly basic region (HBR) that interacts with acidic lipids on the PM1-4. Studies of Gag-membrane interactions using ectopic expression of polyphosphoinositide 5-phosphatase IV (5ptaseIV), an enzyme that catalyzes the hydrolysis of PM-specific acidic phospholipid phosphatidylinositol-(4,5)-bisphosphate [PI(4,5)P2] to phosphatidylinositol-4-phosphate, in cells suggested that Gag-PM localization is mediated by PI(4,5)P23,5 . However, in vitro studies aiming at understanding more mechanistic details of Gag-PI(4,5)P2 interactions have proven to be challenging for a number of reasons. For example, purification of full-length myristoylated HIV-1 Gag for biochemical experiments has been technically difficult at least in part due to the tendency of Gag to aggregate during purification. Hence, truncated forms of HIV-1 Gag, such as Myr-MA or Myr-MA-CA, or the non-myristoylated form have been frequently used in studies that necessitate Gag purification (e.g., nuclear magnetic resonance, protein footprinting, and surface plasmon resonance6-9). Alternatively, coupled in vitro transcription-translation reactions have been used to produce full-length myristoylated HIV-1 Gag in other biochemical studies1,2. Typically in this system, a Gag-encoding plasmid is transcribed and translated with eukaryotic cell lysates (e.g., rabbit reticulocyte lysates) that are devoid of any cellular membranes and messenger RNAs but contain the machinery for transcription and translation. After the reaction, cell lysates containing Gag are mixed with membranes for analysis of Gag interactions with lipids. In addition to the ease of preparing full-length myristoylated Gag, methods using the in vitro transcription translation system have an advantage that Gag synthesis and subsequent membrane binding reactions occur in an 'eukaryotic cytosol-like' milieu that may better represent physiological conditions. This property contributed to the studies that showed that RNA molecules bound to the MA domain regulate Gag binding to acidic lipids in a competitive manner1,2,10-12. However, since the total amount of Gag proteins obtained in these cell lysates are not high, metabolic labeling of proteins with radiolabeled amino acids is necessary for their detection.
Depending on the method to measure Gag-lipid interactions, a variety of membrane preparations have been used. Each of these methods has its strengths and limitations. Most NMR-based assays require the use of lipids with short acyl chains that are water-soluble (e.g., C4- and C8-PI(4,5)P2)6,8. While NMR methods to test binding of Gag to the lipids that have long acyl chains found in cells are being developed, they have been used only with myristoylated or nonmyristoylated MA thus far8,13. Alternatively, liposomes prepared from lipids that have native length acyl chains have been used in biochemical methods such as liposome flotation or fluorescent liposome bead binding assays2,3,10,14-16. However, liposomes used in these assays have small diameters, and thus their membranes have steep and positive curvatures. In contrast, during the early phase of particle assembly in HIV-1-infected cells, Gag binds to the PM, which is nearly planar on the scale of Gag, and subsequently induces negative curvature during budding. Therefore, liposome membranes with steep and positive curvature might not be ideal lipid bilayers to study Gag-lipid interactions. As for liposome flotation assays, another potential caveat is that exposure of Gag-lipid complexes to hypertonic sucrose gradient during centrifugation may affect the experimental outcome. To alleviate these limitations and provide a complementary experimental system, assays for Gag binding to giant unilamellar vesicles (GUVs) have been developed in recent years. GUVs are single lipid bilayer vesicles whose diameters extend to several tens of micrometers. Thus, the curvature of these membranes resembles the PM on the scale of Gag. Furthermore, due to its large size, which enables visual inspection under optical microscopes, membrane binding of fluorescently tagged or labeled Gag proteins to these vesicles upon mixing can be easily determined without subsequent processing of Gag-lipid complexes.
We here describe a protocol to study HIV-1 Gag membrane binding using GUVs obtained from an electroformation method. Various methods such as gentle hydration, gel-assisted hydration, microfluidic jetting, and electroformation17-22 have been used to obtain GUVs. For the protocol described here, the electroformation method is used primarily because of its efficiency in forming GUVs with acidic lipids and its relative ease of use without the need of expensive setups. Since visualization of Gag necessitates a fluorescent reporter, yellow fluorescent protein (YFP) is genetically added to the C-terminus of Gag (Gag-YFP). Gag-YFP proteins are obtained by in vitro transcription and translation reactions in wheat germ lysates based on the continuous exchange-continuous flow (CECF) technology. In this technology, both removal of inhibitory byproducts of the reactions and supply of reaction substrates and energy components are achieved in a dialysis-based mechanism. For these reactions, a plasmid encoding Gag-YFP under the control of a T7 promoter is used. Of note, as shown earlier, wheat germ lysates support myristoylation without additional components23,24. Using this method, it has been possible to obtain sufficient quantities of full-length myristoylated Gag-YFP for visualization of Gag on GUV membranes24. Here we describe the protocol with which HIV-1 Gag binding to PI(4,5)P2-containing GUV membranes can be examined without lengthy subsequent processing following binding reactions and propose that this method complements preexisting Gag-membrane binding assays and can be extended to further understand HIV-1 Gag-membrane binding.
Access restricted. Please log in or start a trial to view this content.
Protocol
Day 1: Expression of Gag Proteins Using the Wheat Germ Lysate-based In Vitro Transcription-translation System
1. Preparation of HIV-1 Gag
- Remove all the reagents of the commercial wheat germ CECF kit from freezers (wheat germ lysates are stored at -80 ºC; other reagents at -20 ºC). Thaw them on ice and mix the components as shown in Table 1.
| Mix-1 (Feeding mix) | |
| Feeding solution | 900 µl |
| Amino acids | 80 µl |
| Methionine | 20 µl |
| Total | 1,000 µl |
| Mix-2 (Lysate mix) | |
| Wheatgerm lysates | 15 µl |
| RNase free water | 7 µl |
| Amino acids | 4 µl |
| Methionine | 1 µl |
| Plasmid DNA in TE buffer (1 µg/µl)** | 4 µl |
| Reaction buffer | 15 µl |
| RNasin* | 4 µl |
| Total | 50 µl |
Table 1: Compositions of mixtures required for wheat germ reactions.
Note: If the experiment is designed to examine the effect of RNA removal by RNase on Gag membrane binding, replace ribonuclease inhibitors with RNase-free water. Plasmid DNA should be free from impurities according to the manufacturer's instruction. However, plasmids prepared using conventional, but not endotoxin-free, plasmid isolation kits have been successfully used. The manufacturer's instruction also recommends using DNA dissolved in nuclease-free water. If using DNA suspended in TE [10 mM Tris-HCl (pH 7.4) containing 1 mM EDTA], avoid using lower concentrations of DNA, since EDTA in TE may affect the yield. Plasmids dissolved in TE buffer at a concentration range of 0.8-1 µg/µl have successfully been used.
- First add the Feeding mixture (Mix-1) into the reaction cassette (transparent chamber). Then add the lysate mix (Mix-2) to the other chamber (red). Do not introduce any bubbles while adding the mixtures into their respective chambers, since that may lead to inefficient exchange of solution components and therefore reduce the efficiency of reaction.
- Cover the chambers with the adhesive film provided with the kit.
- Insert the reaction cassette into the cassette holder and incubate the assembly for 24 hr at 24 ºC at a constant shaking speed of 900 rpm in an Eppendorf thermomixer R.
Day 2: Prepare GUVs by Electroformation and Harvest Wheat Germ Lysates
2. Preparation of GUVs
- Aliquot lipids dissolved in inorganic solvents such as chloroform or mixtures of chloroform and methanol in glass vials with screw caps lined with Teflon liners and wrapped with Parafilm. Handle lipids with Hamilton-type glass syringes, or if available, chloroform-resistant plastic tips. Take out glass vials containing the lipids to be used for GUV preparation from the -20 ºC freezer.
- Once the vials are equilibrated to room temperature, make sure by visual inspection that the lipid suspensions are clear. The quality of lipids, particularly acidic lipids, such as palmitoyl-oleoyl phosphatidylserine (POPS) and PI(4,5)P2, is important for successful experiments. Do not use lipids that are in aliquots for longer than 2 months.
- Pre-warm a heat block to the desired temperature. This temperature must be at least 5 ºC higher than the melting temperature (Tm) of the lipid(s) with the highest Tm.
- Calculate the lipid volumes needed according to the desired molar lipid ratios. For studying HIV-1 Gag binding here, use the following molar ratios as shown in Table 2.
| Lipid mix | Molar ratio | Volume (in µl) | Stock concentration |
| POPC+POPS+Chol | 46.6+23.3+30 | 19.74+10.18+6.57 | POPC, POPS, Chol:10 mg/ml |
| POPC+POPS+Chol+Brain-PI(4,5)P2 | 40+20+30+10 | 16.11+8.3+6.25+58.19 | POPC, POPS, Chol:10 mg/ml |
| Brain-PI(4,5)P2:1 mg/ml |
Table 2: Volumes of different lipids used to obtain GUVs.
- Add the desired volumes of lipids into a clean screw-capped glass vial.
- Use ITO-coated slides of dimensions 25 x 50 x 1.1 mm3 and resistance 70-100 Ω (as described in the catalog). Place two ITO-coated slides on a clean bench. Each electroformation chamber requires two indium-tin-oxide (ITO)-coated glass slides. Verify that the conductive side of the glass slide is facing up by checking the resistance using a multimeter. It should show a value of approximately 200 Ω. Make sure the surface of the slides are clean by gently wiping with 70% ethanol using lint-free wipes.
- Clean the outer surface of a needle of a syringe by rinsing it with chloroform and place the syringe on the heat block.
- Place a new ITO-coated glass slide on the heat block with conductive side facing up and allow it to equilibrate to the desired temperature usually about 2-3 min. Do not reuse the ITO-coated glass slides for a potential risk that the ITO-coating on the glass slides might be damaged during cleaning procedures.
- Use a volume of 40-50 µl of the lipid mixture for each slide for uniform and yet quick spreading of the lipids on the glass slide. Adjust the total volume with an appropriate organic solvent such as chloroform.
- Place one half of the total volume of the lipid mixture on the glass slide and immediately spread the lipid using the cleaned syringe (step 2.6) to leave a uniform lipid film (moving the needle 2-3 times across the slide). Spread the lipid film gently (but quickly) to avoid any damage to the thin ITO coating. Non-uniform spreading or rough spreading may lead to sub-optimal yields or computational heterogeneity of GUVs.
- Therefore, ensure uniform spread by inspecting the lipid film after spread before proceeding to the next step. If the opacity of the film appears excessively non-uniform, redo the procedure using a new slide.
- Place the coated slide inside a Petri dish with the coated side facing up.
- Repeat Steps 2.6-2.9 for the rest of the mixture using another ITO-coated slide.
- Place the petri dish containing both the slides in a conventional vacuum desiccation chamber for 60-90 min to remove any trace of organic solvents.
- During this process, set the incubator to the same temperature as that of the heat block (65 ºC in most cases) and pre-warm 300 mM sucrose solution in the incubator.
- After drying, bring the petri dish to the bench and place the glass slide on a clean surface.
- Gently clean a PDMS gasket (Figure 1) using 70% ethanol and dry it completely.
- Place it on the lipid-coated glass slide as shown in Figure 1 and gently and firmly press so that the gasket forms a water tight seal. Confirm the absence of gaps between the PDMS gasket and the slide. This results in the formation of a shallow chamber that holds the sucrose solution.

Figure 1: The layout of the ITO-coated glass slide used for electroformation. Please click here to view a larger version of this figure.
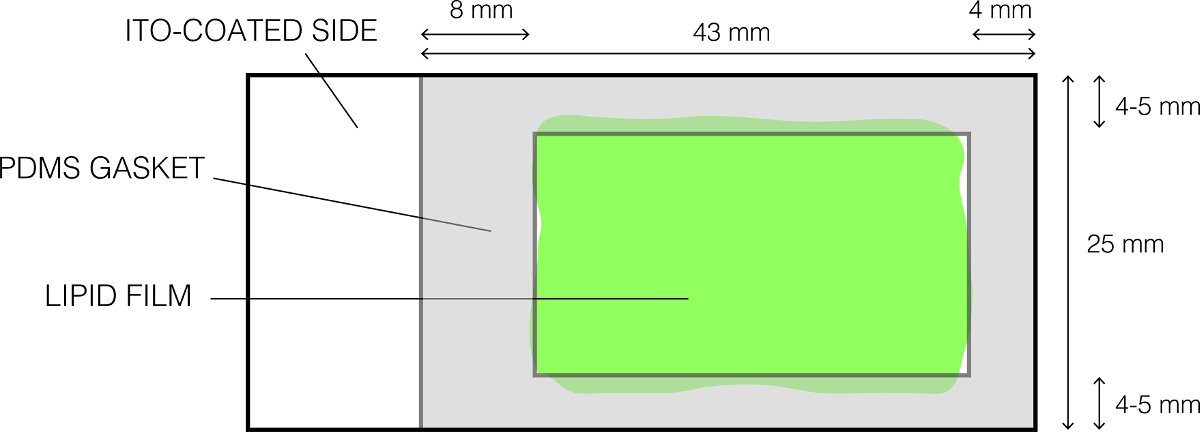{kind=link}
- Fill the chamber with pre-warmed sucrose solution without any bubbles.
- Place the other coated slide on top to make a tight seal. Ensure that there are no bubbles. The final assembly should appear as shown in Figure 2. Fasten the chamber using binder clips.

Figure 2: Schematic representation of the electroformation chamber (side view). Please click here to view a larger version of this figure.
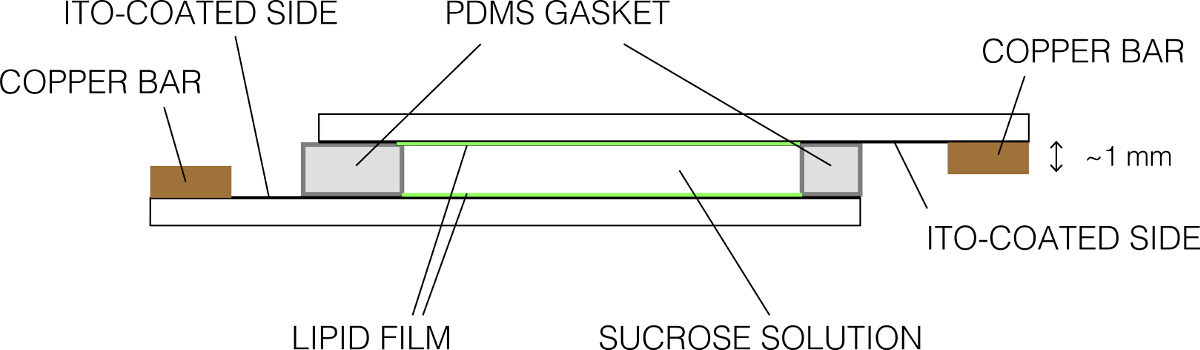{kind=link}
- Remove excess sucrose by aspiration and clean the slides using lint-free wipes. Fasten the slides to the copper bar so that the conductive sides are in uniform contact with the bar. Ensure that only one conductive side of the glass slide is in contact with one copper bar.
- Connect the copper bar to the function generator output using the alligator clips. Place the assembly inside the incubator to allow the assembly to equilibrate to the desired temperature.
- Apply a sine wave frequency of 10 Hz and a potential difference of 1 V for 90 min. Ensure that the voltage is 1 V by a multimeter as shown in Figure 3. Continue the process for 90 min.

Figure 3: The positions at which the electrodes from the multimeter are placed for measuring frequency and current. Please click here to view a larger version of this figure.
{kind=link}
- After 90 min, slowly decrease the frequency to 2 Hz and continue electroformation for another 10 min. During this time, harvest in vitro translation reactions (steps 3.1-3.2).
- Switch off the function generator and allow the assembly to cool down to room temperature slowly (by turning off the incubator and leaving the incubator door slightly opened).
- After cooling, disconnect the cables to the generator from the slide assemblies. GUVs may be very fragile and therefore need to be handled very gently. Carefully bring the assembly to the bench.
- Disassemble the chamber by gently removing the top slide using forceps. Harvest the sucrose suspension containing GUVs using a cut 1,000 µl tip and transfer to a clean tube. Handle the tube gently to prevent disruption of GUVs.
- Use the GUVs immediately. GUVs are stable for at least 90 min after harvesting when stored at room temperature.
3. Harvest In Vitro Translation Reactions
- Harvest the in vitro translation reactions containing desired proteins from the red chamber using a micropipette into a tube.
- To remove aggregated proteins, spin the lysates at 16,200 x g for 15 min on a tabletop centrifuge and carefully transfer the supernatant into another tube.
4. GUV Binding Assay
- Using a cut tip, mix 5 µl of in vitro translation reactions containing translated proteins and 5 µl of GUVs in a tube and incubate at room temperature for 2-3 min.
- Attach a PDMS sheet with a small hole (3-4 mm diameter) on a clean cover slip and ensure tight seal by gently and firmly pressing it against the coverslip.
- Place the 10 µl mixture from step 4.1 into the small hole.
- Image the mixture under an inverted epi- or confocal fluorescence microscope at room temperature.
Note: To prevent evaporation, the imaging chamber can be covered by a coverslip.
Access restricted. Please log in or start a trial to view this content.
Results
Using the above protocol, we prepared GUVs composed of POPC+POPS+Chol (molar ratio: 4.66:2.33:3). This composition was chosen to approximately reflect the PS and cholesterol concentrations of the PM. Robust and efficient binding of Gag was observed only when brain-PI(4,5)P2 was included into the POPC+POPS+Chol mixture (POPC+POPS+Chol+brain-PI(4,5)P2 [molar ratio: 4:2:3:1] in this example) (Figure 4, compare panels B and D with A and C). Gag-YFP bindi...
Access restricted. Please log in or start a trial to view this content.
Discussion
The GUV binding assay as described above provides a good alternative in scenarios where other protein-lipid interaction assays have their limitations. This assay allows us to examine interactions between myristoylated full-length Gag and acidic lipids with native-length acyl chains in the lipid bilayer context and do so without lengthy flotation centrifugation through high-density sucrose gradients or other post-binding processing of Gag-lipid complexes. Another advantage is that the behavior of Gag can be examined in a ...
Access restricted. Please log in or start a trial to view this content.
Disclosures
We declare that we have no conflicting interests.
Acknowledgements
We would like to thank Mohammad Saleem, Jing Wu and Krishnan Raghunathan for helpful discussions. We also thank Priya Begani for assistance during the filming. This work is supported by National Institutes of Health grants R01 AI071727 (to A.O.) and R01 GM110052 (to S.L.V).
Access restricted. Please log in or start a trial to view this content.
Materials
| Name | Company | Catalog Number | Comments |
| Digital Multimeter | Meterman | 30XR | A generic multimeter that can measure resistance and volts will serve the purpose. |
| Function Generator | Instek | GFG-8216A | A generic function generator that is capable of generating a sine wave at 10 Hz and 1 V is sufficient. |
| ITO coated glass slides | Delta Technologies, Loveland, CO | CG-90IN | |
| Incubator | Hoefer | Any incubator that can accurately maintain temperature will be sufficient | |
| Vacuum chamber | Nalgene | ||
| Thermomixer R | Eppendorf | 21516-166 | |
| Syringe | Hamilton | 80400 | Gauge 22S, Syringe number 702 |
| PDMS | Sylgard elastomer base kit, Dow-Corning | Sylgard, 184 | |
| RTS 100 Wheat Germ CECF Kit | BiotechRabbit, Berlin, Germany | BR1401001 | |
| DiD | Life Technologies, Carlsbad, CA | D7757 | |
| POPC | Avanti Polar Lipids, Alabaster, AL | 850457C | |
| POPS | Avanti Polar Lipids | 840034C | |
| Cholesterol | Avanti Polar Lipids | 700041P | |
| Brain-PI(4,5)P2 | Avanti Polar Lipids | 840046X |
References
- Chukkapalli, V., Inlora, J., Todd, G. C., Ono, A. Evidence in support of RNA-mediated inhibition of phosphatidylserine-dependent HIV-1 Gag membrane binding in cells. J Virol. 87 (12), 7155-7159 (2013).
- Chukkapalli, V., Oh, S. J., Ono, A. Opposing mechanisms involving RNA and lipids regulate HIV-1 Gag membrane binding through the highly basic region of the matrix domain. Proc Natl Acad Sci U S A. 107 (4), 1600-1605 (2010).
- Chukkapalli, V., Hogue, I. B., Boyko, V., Hu, W. S., Ono, A. Interaction between the human immunodeficiency virus type 1 Gag matrix domain and phosphatidylinositol-(4,5)-bisphosphate is essential for efficient gag membrane binding. J Virol. 82 (5), 2405-2417 (2008).
- Zhou, W., Parent, L. J., Wills, J. W., Resh, M. D. Identification of a membrane-binding domain within the amino-terminal region of human immunodeficiency virus type 1 Gag protein which interacts with acidic phospholipids. J Virol. 68 (4), 2556-2569 (1994).
- Ono, A., Ablan, S. D., Lockett, S. J., Nagashima, K., Freed, E. O. Phosphatidylinositol (4,5) bisphosphate regulates HIV-1 Gag targeting to the plasma membrane. Proc Natl Acad Sci U S A. 101 (41), 14889-14894 (2004).
- Saad, J. S., et al. Structural basis for targeting HIV-1 Gag proteins to the plasma membrane for virus assembly. Proc Natl Acad Sci U S A. 103 (30), 11364-11369 (2006).
- Shkriabai, N., et al. Interactions of HIV-1 Gag with assembly cofactors. Biochemistry. 45 (13), 4077-4083 (2006).
- Vlach, J., Saad, J. S. Trio engagement via plasma membrane phospholipids and the myristoyl moiety governs HIV-1 matrix binding to bilayers. Proc Natl Acad Sci U S A. 110 (9), 3525-3530 (2013).
- Anraku, K., et al. Highly sensitive analysis of the interaction between HIV-1 Gag and phosphoinositide derivatives based on surface plasmon resonance. Biochemistry. 49 (25), 5109-5116 (2010).
- Llewellyn, G. N., Grover, J. R., Olety, B., Ono, A. HIV-1 Gag associates with specific uropod-directed microdomains in a manner dependent on its MA highly basic region. J Virol. 87 (11), 6441-6454 (2013).
- Inlora, J., Chukkapalli, V., Derse, D., Ono, A. Gag localization and virus-like particle release mediated by the matrix domain of human T-lymphotropic virus type 1 Gag are less dependent on phosphatidylinositol-(4,5)-bisphosphate than those mediated by the matrix domain of HIV-1 Gag. J Virol. 85 (8), 3802-3810 (2011).
- Inlora, J., Collins, D. R., Trubin, M. E., Chung, J. Y., Ono, A. Membrane binding and subcellular localization of retroviral Gag proteins are differentially regulated by MA interactions with phosphatidylinositol-(4,5)-bisphosphate and RNA. MBio. 5 (6), e02202(2014).
- Valentine, K. G., et al. Reverse micelle encapsulation of membrane-anchored proteins for solution NMR studies. Structure. 18 (1), 9-16 (2010).
- Dick, R. A., Goh, S. L., Feigenson, G. W., Vogt, V. M. HIV-1 Gag protein can sense the cholesterol and acyl chain environment in model membranes. Proc Natl Acad Sci U S A. 109 (46), 18761-18766 (2012).
- Dick, R. A., Kamynina, E., Vogt, V. M. Effect of multimerization on membrane association of Rous sarcoma virus and HIV-1 MA proteins. J Virol. 87 (24), 13598-13608 (2013).
- Alfadhli, A., Still, A., Barklis, E. Analysis of human immunodeficiency virus type 1 matrix binding to membranes and nucleic acids. J Virol. 83 (23), 12196-12203 (2009).
- Rodriguez, N., Pincet, F., Cribier, S. Giant vesicles formed by gentle hydration and electroformation: a comparison by fluorescence microscopy. Colloids Surf B Biointerfaces. 42 (2), 125-130 (2005).
- Needham, D., McIntosh, T. J., Evans, E. Thermomechanical and transition properties of dimyristoylphosphatidylcholine/cholesterol bilayers. Biochemistry. 27 (13), 4668-4673 (1988).
- Darszon, A., et al. Reassembly of protein-lipid complexes into large bilayer vesicles: perspectives for membrane reconstitution. Proc Natl Acad Sci U S A. 77 (1), 239-243 (1980).
- Angelova, M. I., Dimitrov, D. S. Liposome electroformation. Faraday Discuss. Chem. Soc. (81), 303-311 (1986).
- Angelova, M. I., Soléau, S., Méléard, P. h, Faucon, F., Bothorel, P. Preparation of giant vesicles by external AC electric fields. Progr Colloid Polymer Sci. 89, 127-131 (1992).
- Stachowiak, J. C., et al. Unilamellar vesicle formation and encapsulation by microfluidic jetting. Proc Natl Acad Sci U S A. 105 (12), 4697-4702 (2008).
- Yamauchi, S., et al. The consensus motif for N-myristoylation of plant proteins in a wheat germ cell-free translation system. FEBS J. 277 (17), 3596-3607 (2010).
- Olety, B., Veatch, S. L., Ono, A. Phosphatidylinositol-(4,5)-Bisphosphate Acyl Chains Differentiate Membrane Binding of HIV-1 Gag from That of the Phospholipase Cdelta1 Pleckstrin Homology Domain. J Virol. 89 (15), 7861-7873 (2015).
- Morales-Penningston, N. F., et al. GUV preparation and imaging: minimizing artifacts. Biochim Biophys Acta. 1798 (7), 1324-1332 (2010).
- Veatch, S. L. Electro-formation and fluorescence microscopy of giant vesicles with coexisting liquid phases. Methods Mol Biol. 398, 59-72 (2007).
- Ayuyan, A. G., Cohen, F. S. Lipid peroxides promote large rafts: effects of excitation of probes in fluorescence microscopy and electrochemical reactions during vesicle formation. Biophys J. 91 (6), 2172-2183 (2006).
- Montes, L. R., et al. Electroformation of giant unilamellar vesicles from native membranes and organic lipid mixtures for the study of lipid domains under physiological ionic-strength conditions. Methods Mol Biol. 606, 105-114 (2010).
- Olety, B., Ono, A. Roles played by acidic lipids in HIV-1 Gag membrane binding. Virus Res. 193, 108-115 (2014).
- Keller, H., Krausslich, H. G., Schwille, P. Multimerizable HIV Gag derivative binds to the liquid-disordered phase in model membranes. Cell Microbiol. 15 (2), 237-247 (2013).
- Carlson, L. A., Hurley, J. H. In vitro reconstitution of the ordered assembly of the endosomal sorting complex required for transport at membrane-bound HIV-1 Gag clusters. Proc Natl Acad Sci U S A. 109 (42), 16928-16933 (2012).
- Gui, D., et al. A novel minimal in vitro system for analyzing HIV-1 Gag-mediated budding. J Biol Phys. 41 (2), 135-149 (2015).
- Prevost, C., et al. IRSp53 senses negative membrane curvature and phase separates along membrane tubules. Nat Commun. 6, 8529(2015).
Access restricted. Please log in or start a trial to view this content.
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved