
Zum Anzeigen dieser Inhalte ist ein JoVE-Abonnement erforderlich. Melden Sie sich an oder starten Sie Ihre kostenlose Testversion.
Method Article
Visualisierung von HIV-1 Gag Bindung an Riesen unilamellare Vesikeln (GUV) Membranes
In diesem Artikel
Zusammenfassung
We illustrate here an in vitro membrane binding assay in which interactions between HIV-1 Gag and lipid membranes are visually analyzed using YFP-tagged Gag synthesized in a wheat germ-based in vitro translation system and GUVs prepared by an electroformation technique.
Zusammenfassung
Das Strukturprotein von HIV-1, Pr55 Gag (oder Gag), bindet an die Plasmamembran in Zellen während der Virus - Montageprozesses. Membranbindung von Gag ist ein wesentlicher Schritt für die Viruspartikelbildung, da ein Defekt in Gag Membranbindungs führt zu einer starken Beeinträchtigung der viralen Partikelproduktion. Zu gewinnen sind mechanistischen Details Gag-Lipid - Membran - Wechselwirkungen in vitro - Verfahren basiert auf NMR, Protein Fußabdrucks, Oberflächen - Plasmonresonanz, Liposom - Flotation Zentrifugation oder Fluoreszenz Lipid Wulst Bindung so weit entwickelt. Jedoch jede dieser in - vitro - Verfahren hat seine Grenzen. Um einige dieser Beschränkungen und bieten einen komplementären Ansatz zu den bisher etablierten Methoden zu überwinden, entwickelten wir ein in - vitro - Test , bei dem Interaktionen zwischen HIV-1 Gag und Lipidmembranen finden in einem "zellähnlichen" Umfeld. In diesem Test wird die Bindung an Gag Lipidmembranen visuell analysiert YFP-Tagged Gag in einem Weizen synthetisierten Keim-basierten in vitro - Translationssystem und GUVs durch eine Elektroformation Technik hergestellt. Hier beschreiben wir den Hintergrund und die Protokolle myristoylierten voller Länge Gag Proteinen und GUV Membranen, die für den Assay zu erhalten, und Gag-GUV Bindung durch Mikroskopie zu detektieren.
Einleitung
Human Immunodeficiency Virus Typ 1 (HIV-1) ist ein umhülltes Virus, das an und Knospen aus der Plasmamembran (PM) in den meisten Zelltypen assembliert. Die Montage von HIV-1 - Viruspartikel wird durch das 55 kDa viralen Kernprotein genannt Pr55 Gag (gag) angetrieben. Gag wird als ein Vorläufer-Polyproteins synthetisiert besteht aus vier Hauptstrukturdomänen, nämlich, matrix, Capsid, Nucleocapsid und p6 sowie zwei Distanz Peptide SP1 und SP2. Bei der Montage ist die Matrix (MA) Domäne verantwortlich für die Ausrichtung der Gag an den Montageort, das Kapsid (CA) Domäne Gag-Gag-Interaktionen vermittelt, rekrutiert der Nucleocapsid (NC) Domäne virale genomische RNA und p6 Rekruten Wirtsfaktoren, dass die Hilfe Viruspartikel scission aus der Plasmamembran. Gag erfährt auch eine co-translationale Modifikation durch die Zugabe einer 14-Kohlenstoff-Fettsäure oder Myristat-Rest an seinem N-Terminus.
Membranbindung von Gag ist eine wesentliche Voraussetzung für die virale Montage, da mutants, die defekt in der Membran sind verbindlich versagen Viruspartikel zu erzeugen. Wir und andere haben gezeigt, dass die Membranbindung von Gag dargestellt durch bipartite Signale innerhalb der MA-Domäne vermittelt wird: die N-terminalen Myristat-Einheit, die mit der Lipid-Doppelschicht und einem Cluster basischer Reste innerhalb der MA-Domäne bezeichnet als stark basische Region hydrophober Wechselwirkungen vermittelt ( HBR) , die mit sauren Lipiden auf der PM 1-4 zusammenwirkt. Studien der Gag-Membran - Wechselwirkungen unter Verwendung ektopische Expression von Polyphosphoinositid 5-phosphatase IV (5ptaseIV), ein Enzym, das die Hydrolyse von PM-spezifische saure Phospholipid phosphatidylinositol- (4,5) -bisphosphate [PI (4,5) P 2] katalysiert zu Phosphatidylinositol-4-phosphat, schlug in Zellen , die Gag-PM Lokalisierung von PI (4,5) P 2 3,5 vermittelt wird. Jedoch, mit dem Ziel in - vitro - Studien auf das Verständnis mehr mechanistischen Details Gag-PI (4,5) P 2 Wechselwirkungen haben bewiesen , für eine Reihe von Gründen in Frage zu stellen. FürBeispiel Reinigung von voller Länge myristoyliert HIV-1 Gag für biochemische Experimente hat sich zumindest teilweise aufgrund der Tendenz von Gag technisch schwierig während der Reinigung zu aggregieren. Daher verkürzte Formen von HIV-1 Gag, wie Myr-MA oder Myr-MA-CA, oder das nicht-myristoylierten Form häufig in Untersuchungen verwendet wurden , die Gag - Reinigung ( zum Beispiel Kernspinresonanz, Protein footprinting erfordern und Oberflächen Plasmonresonanz 6-9). Alternativ gekoppelt ist , in vitro Transkriptions-Translations - Reaktionen verwendet wurden , 1,2 voller Länge myristoyliert HIV-1 Gag in andere biochemische Studien herzustellen. Typischerweise in diesem System wird ein Gag-kodierenden Plasmid wird mit eukaryotische Zelllysate (zB Kaninchen - Retikulozyten - Lysaten) , die frei von jeglicher Zellmembranen und messenger RNAs transkribiert und translatiert , sondern die Maschinen für die Transkription und Translation enthalten. Nach der Reaktion enthält Gag Zelllysate werden bei mir gemischtmbranes zur Analyse von Gag Wechselwirkungen mit Lipiden. Zusätzlich zu der Leichtigkeit , mit voller Länge zur Herstellung myristoyliert Gag, Methoden der in vitro Transkriptions - Translations - System verwenden , haben den Vorteil , dass Gag - Synthese und nachfolgende Membranbindungsreaktionen in einem "eukaryotischen Cytosol-like 'Milieu auftreten , die eine bessere physiologische Bedingungen darstellen. Diese Eigenschaft trug den Studien zu , die zeigten , dass RNA - Moleküle an die MA - Domäne gebunden regulieren Gag in kompetitiver Weise 1,2,10-12 zu saure Lipide zu binden. Da jedoch in diesen Zellysaten die Gesamtmenge der Gag-Proteine erhalten werden, nicht hoch ist, ist die metabolische Markierung von Proteinen mit radiomarkierten Aminosäuren für deren Nachweis notwendig.
Je nach Methode Gag-Lipid-Wechselwirkungen zu messen, eine Vielzahl von Membranpräparationen verwendet wurden. Jede dieser Methoden hat ihre Stärken und Schwächen. Die meisten NMR-basierte Assays erfordern die Verwendung von Lipiden mit kurzen acylKetten , die wasserlösliche (zB C4- und C8-PI (4,5) P 2) 6,8 sind. Während NMR Methoden Bindung von Gag zu den Lipiden zu testen , die in Zellen gefunden langen Acylketten aufweisen entwickelt werden, haben sie bisher nur mit 8,13 myristoylierten oder nonmyristoylated MA verwendet. Alternativ Liposomen aus Lipiden , die native Länge Acylketten haben wurden in biochemischen Methoden wie Liposomen Flotation oder fluoreszierende Liposom Perle Bindungsassays 2,3,10,14-16 verwendet. Jedoch in diesen Assays verwendeten Liposomen haben kleine Durchmesser, und somit deren Membranen haben steile und positive Krümmungen. Im Gegensatz dazu ist während der frühen Phase der Partikelanordnung in HIV-1-infizierten Zellen bindet Gag an die PM, die nahezu auf der Skala von Gag planar und induziert anschließend negative Krümmung während Knospung. Daher Liposomenmembranen mit steilen und positive Krümmung vielleicht nicht ideal Lipid-Doppelschichten zu Gag-Lipid-Wechselwirkungen untersuchen. Wie für Liposom flichtation Assays ist eine weitere mögliche Einschränkung, dass die Exposition von Gag-Lipid-Komplexen zu hypertonischen Saccharosegradienten während der Zentrifugation die experimentellen Ergebnisse beeinflussen können. Um diese Einschränkungen zu mildern und ein komplementäres experimentelles System bereitstellen, Assays für die Bindung an Gag giant unilamellaren Vesikeln (GUV), wurden in den letzten Jahren entwickelt. GUVs sind einzelne Lipid-Doppelschicht-Vesikel, deren Durchmesser erstrecken sich bis zu mehreren zehn Mikrometern. Somit gleicht die Krümmung dieser Membranen der PM auf der Skala von Gag. Außerdem ist es aufgrund seiner großen Größe, die unter optischen Mikroskopen Sichtprüfung ermöglicht, Binde- Membran von fluoreszenz getaggt oder markiert Gag-Proteine dieser Vesikel beim Mischen kann leicht ohne nachfolgende Verarbeitung Gag-Lipid-Komplexe bestimmt werden.
Wir beschreiben hier ein Protokoll HIV-1 Gag Membran zu untersuchen Bindung mit GUVs von einem Elektroformation Verfahren erhalten. Verschiedene Methoden wie sanfte Feuchtigkeit, Gel-unterstützte Trink, microfluidic Jetting und Elektroformation 17-22 wurden verwendet GUVs zu erhalten. Für das hier beschriebene Protokoll wird die Elektroformation Verfahren in erster Linie wegen seiner Effizienz bei der Bildung GUVs mit sauren Lipiden und seine relative Einfachheit der Verwendung, ohne die Notwendigkeit von teuren Aufbauten verwendet. Da die Visualisierung von Gag ein fluoreszierendes Reporter erfordert, ist genetisch gelb fluoreszierendes Protein (YFP) an den C-Terminus von Gag (Gag-YFP) zugegeben. Gag-YFP Proteine werden durch in vitro Transkriptions- und Translationsreaktionen in wheat germ Lysaten erhalten auf der Basis der kontinuierlichen Austausch kontinuierlichen Strömung (CECF) Technologie. In dieser Technologie wird sowohl die Entfernung von inhibitorischen Nebenprodukte der Reaktionen und Zufuhr von Reaktionssubstraten und Energiekomponenten sind in einer Dialysebasierten Mechanismus erreicht. Bei diesen Reaktionen wird ein Plasmid, kodierend Gag-YFP unter der Kontrolle eines T7-Promotors verwendet. Bemerkenswert ist , wie bereits gezeigt, Weizenkeim Lysaten unterstützen Myristoylation ohne zusätzliche Komponenten 23,24. Myristoyliert Gag-YFP zur Visualisierung von Gag auf GUV Membranen 24 Mit dieser Methode war es möglich , ausreichende Mengen an in voller Länge zu erhalten. Hier beschreiben wir das Protokoll , mit dem HIV-1 Gag zu PI - Bindung (4,5) P 2 -haltigen GUV Membranen können ohne langwierige nachfolgende Verarbeitung folgende Bindungsreaktionen untersucht und schlagen vor , dass diese Methode bereits existierenden Gag-Membran - Bindungstests ergänzt und kann sein erweitert, um weitere Bindung HIV-1 Gag-Membran zu verstehen.
Access restricted. Please log in or start a trial to view this content.
Protokoll
Tag 1: Expression von Gag - Proteinen Mit dem Weizenkeim - Lysat-basierten In - vitro - Transkriptions-Translations - System
1. Herstellung von HIV-1 Gag
- Entfernen Sie alle Reagenzien des kommerziellen Weizenkeim CECF Kit von Gefrierschränken (Weizenkeim Lysate bei -80 ° C gelagert werden, andere Reagenzien bei -20 ° C). Tauen Sie sie auf Eis und mischen Sie die Komponenten wie in Tabelle 1 gezeigt.
| Mix-1 (Feeding mix) | |
| Feeding-Lösung | 900 ul |
| Aminosäuren | 80 ul |
| Methionine | 20 ul |
| Gesamt | 1000 ul |
| Mix-2 (Lysate mix) | |
| wheatgerm Lysate | 15 ul |
| RNase freies Wasser | 7 ul |
| Aminosäuren | 4 ul |
| Methionine | 1 ul |
| Plasmid DNA in TE-Puffer (1 & mgr; g / & mgr; l) ** | 4 ul |
| Reaktionspuffer | 15 ul |
| RNasin * | 4 ul |
| Gesamt | 50 ul |
Tabelle 1: Zusammensetzungen von Mischungen für Weizenkeim Reaktionen erforderlich.
Hinweis: Wenn das Experiment entwickelt, um die Wirkung von RNA-Entfernung durch RNase auf Gag Membranbindung, ersetzen Ribonuklease-Inhibitoren mit RNase-freiem Wasser zu untersuchen. Plasmid DNA sollte entsprechend den Anweisungen des Herstellers frei von Verunreinigungen sein. Jedoch plasmids hergestellt unter Verwendung konventioneller, aber nicht Endotoxin-frei, Plasmidisolation Kits erfolgreich eingesetzt wurden. Die Anweisungen des Herstellers empfiehlt auch DNA in Nuklease-freies Wasser gelöst werden. Bei der Verwendung von DNA in TE suspendiert [10 mM Tris-HCl (pH 7,4), enthaltend 1 mM EDTA], vermeiden niedrigere Konzentrationen von DNA verwendet, da EDTA in TE die Ausbeute auswirken kann. Plasmide, in TE-Puffer 0,8-1 ug / ul in einem Konzentrationsbereich von erfolgreich verwendet worden sind, gelöst.
- Zuerst fügen Sie die Futtermischung (Mix-1) in die Reaktionskassette (transparente Kammer). Dann fügen Sie das Lysat (Mix-2) in die andere Kammer (rot). Führen Sie keine Blasen, während die Gemische in ihre jeweiligen Kammern Zugabe, da diese zu einer ineffizienten Austausch von Lösungskomponenten führen kann und damit die Effizienz der Reaktion zu verringern.
- Decken Sie die Kammern mit der Klebefolie mit dem Kit zur Verfügung gestellt.
- Legen Sie die Reaktionskassette in den Kassettenhalter und Incubate die Baugruppe für 24 h bei 24 ºC bei einer konstanten Schüttelgeschwindigkeit von 900 Upm in einer Eppendorf-Thermomixer R.
Tag 2: Bereiten Sie GUVs von Elektroformation und Ernte Weizenkeim Lysate
2. Herstellung von GUVs
- Aliquotieren Lipide in anorganischen Lösungsmitteln gelöst, wie Chloroform oder Mischungen aus Chloroform und Methanol in Glasfläschchen mit Schraubverschluss ausgekleidet mit Teflon-Liner und mit Parafilm umwickelt. Griff Lipide mit Hamilton-Typ Glasspritzen, oder falls vorhanden, Chloroform beständige Kunststoffspitzen. Nehmen Glasphiolen die Lipide enthalten, für GUV Zubereitung aus der -20 ºC Gefrierschrank verwendet werden.
- Sobald die Fläschchen auf Raumtemperatur ins Gleichgewicht gebracht werden, stellen Sie sicher durch visuelle Inspektion, die die Lipidsuspensionen klar sind. Die Qualität von Lipiden, insbesondere saure Lipide wie Palmitoyl-Oleoyl Phosphatidylserin (POPS) und PI (4,5) P 2, ist wichtig für die erfolgreiche Experimente. Verwenden Sie keine Lipide, die sindin Aliquots für länger als 2 Monate.
- Vorwärmen einen Wärmeblock auf die gewünschte Temperatur. Diese Temperatur muß mindestens 5 ºC höher ist als die Schmelztemperatur (T m) des Lipid (e) mit der höchsten T m liegen.
- Berechne die Lipidmengen benötigt entsprechend den gewünschten molaren Lipid-Verhältnissen. Für das Studium hier HIV-1 Gag Bindung, verwenden Sie die folgenden Molverhältnisse wie in Tabelle 2 dargestellt.
| Lipid - Mix | Molverhältnis | Volumen (in ul) | Auf Konzentration |
| POPC + POPS + Chol | 46,6 + 23,3 + 30 | 19,74 + 10,18 + 6,57 | POPC, POPS, Chol: 10 mg / ml |
| POPC + POPS + Chol + Brain-PI (4,5) P 2 | 16.11 + 8.3 + 6.25 + 58.19 | POPC, POPS, Chol: 10 mg / ml | |
| Brain-PI (4,5) P 2: 1 mg / ml |
Tabelle 2: Volumina verschiedener Lipide GUVs zu erhalten.
- Fügen Sie die gewünschten Mengen von Lipiden in einem sauberen Schraubdeckelröhrchen Glasfläschchen.
- Verwendung ITO-beschichtete Objektträger mit den Abmessungen 25 x 50 x 1,1 mm 3 und Widerstand 70-100 Ω (wie im Katalog beschrieben). Legen Sie zwei ITO-beschichtete Folien auf eine saubere Bank. Jeder Elektroformation Kammer erfordert zwei Indium-Zinn-Oxid (ITO) beschichteten Glasplättchen. Stellen Sie sicher, dass die leitende Seite des Glasobjektträger wird durch Prüfen der Widerstand mit einem Multimeter nach oben zeigen. Es sollte einen Wert von ca. 200 Ω zeigen. Achten Sie darauf, die Oberfläche der Folien sauber sind, indem sie mit 70% Ethanol mit fusselfreien Tüchern sanft abwischen.
- Reinigen der Außenfläche einer Nadel einer Spritze durch rinsing es mit Chloroform und legen Sie die Spritze auf dem Wärmeblock.
- Legen Sie eine neue ITO-beschichtete Glasobjektträger auf dem Wärmeblock mit leitenden Seite nach oben und lassen Sie es auf die gewünschte Temperatur in der Regel ca. 2-3 min äquilibrieren. Nicht über die ITO-beschichtete Glasobjektträger für ein potentielles Risiko wiederzuverwenden, dass die ITO-Schicht auf den Glasobjektträger können bei Reinigungsverfahren beschädigt werden.
- Verwenden Sie ein Volumen von 40 bis 50 & mgr; l der Lipidmischung für jede Folie für eine einheitliche und dennoch schnelle Ausbreitung der Lipide auf dem Glasträger. Stellen Sie das Gesamtvolumen mit einem geeigneten organischen Lösungsmittel, wie Chloroform.
- Legen Sie eine Hälfte des Gesamtvolumens der Lipidmischung auf dem Glasträger und sofort die Lipid verteilt die gereinigte Spritze (Schritt 2.6) ein einheitliches Lipidfilm zu verlassen (Bewegen der Nadel 2-3 mal in der Folie). Verbreiten Sie den Lipidfilm sanft (aber schnell) eine Beschädigung der dünnen ITO-Beschichtung zu vermeiden. Uneinheitliche Verbreitung oder grobe Verbreitung zu suboptimalen führen kannErträge oder Rechen Heterogenität der GUVs.
- Daher gewährleisten gleichmäßige Verteilung durch die Lipidfilm nach Ausbreitung kontrollieren, bevor mit dem nächsten Schritt fortfahren. Wenn die Trübung der Folie übermäßig ungleichmäßige erscheint, wiederholen Sie die Prozedur eine neue Folie verwendet wird.
- Legen Sie die beschichteten Objektträger in einer Petrischale mit der beschichteten Seite nach oben zeigt.
- Wiederholen Sie die Schritte 2,6-2,9 für den Rest der Mischung eine andere ITO-beschichteten Objektträger verwendet wird.
- Legen Sie die Petrischale, die sowohl die Folien in einem herkömmlichen Vakuumtrocknungskammer für 60-90 min zu entfernen jede Spur von organischen Lösungsmitteln.
- Während dieses Prozesses, stellen Sie den Inkubator auf die gleiche Temperatur wie die des Wärmeblock (65 ºC in den meisten Fällen) und vorwärmen 300 mM Saccharose-Lösung in den Inkubator.
- Nach dem Trocknen bringen die Petrischale auf die Bank und legen Sie den Glasobjektträger auf eine saubere Oberfläche.
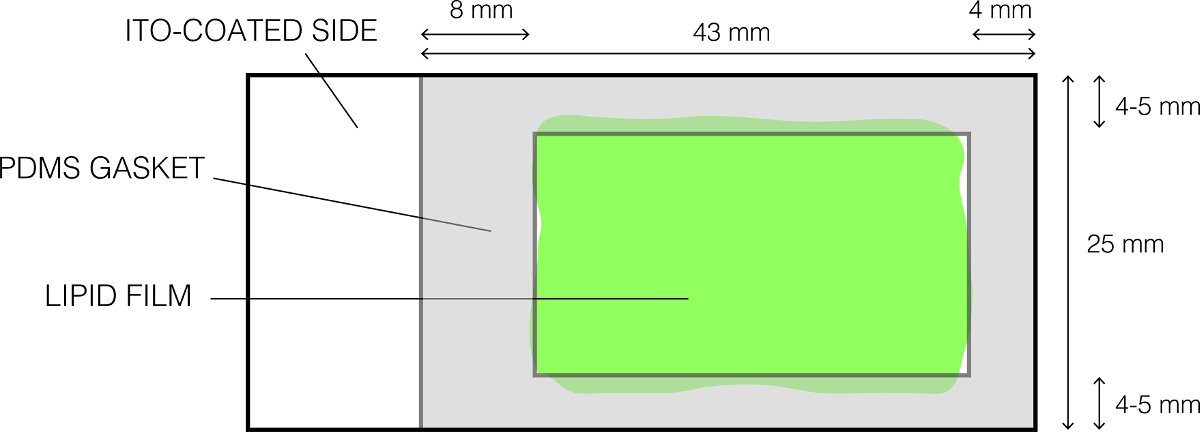
- Reinigen Sie vorsichtig mit einem PDMS - Dichtung (Abbildung 1) unter Verwendung von 70% igem Ethanol und trocken es compledig.
- Legen Sie sie auf die Lipid - beschichteten Glasobjektträger , wie in Abbildung 1 gezeigt und sanft und fest drücken , so dass die Dichtung bildet eine wasserdichte Abdichtung. Bestätigen Sie die Abwesenheit von Lücken zwischen der PDMS-Dichtung und der Folie. Dies resultiert in der Bildung einer flachen Kammer, die das Sucrose-Lösung hält.

Abb . 1: Das Layout der ITO-beschichtete Glasobjektträger für Elektroformation verwendet Bitte hier klicken , um eine größere Version dieser Figur zu sehen.
{kind=link}
- Füllen Sie die Kammer mit vorgewärmten Saccharoselösung ohne Blasen.
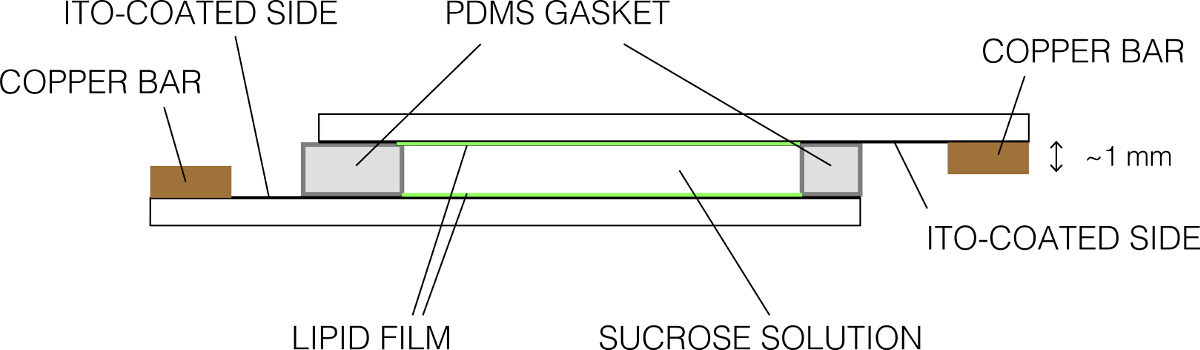
- Legen Sie die andere beschichtete Folie auf oben, um eine dichte Abdichtung zu machen. Stellen Sie sicher, dass es keine Blasen gibt. Die Endmontage sollte wie folgt aussehen in Abbildung 2. Befestigen Sie die Kammer using Bindemittel-Clips.

Abbildung 2:. Schematische Darstellung der Elektroformation Kammer (Seitenansicht) Bitte hier klicken , um eine größere Version dieser Figur zu sehen.
{kind=link}
- Entfernen Sie überschüssige Saccharose durch Absaugen und Reinigen Sie die Folien mit fusselfreien Tüchern. Befestigen Sie die Folien an die Kupferschiene, so dass die leitenden Seiten in gleichmäßigen Kontakt mit der Bar sind. Stellen Sie sicher, dass nur eine leitfähige Seite des Glasobjektträger in Kontakt mit einem Kupferstab ist.
- Verbinden Sie die Kupferschiene an den Funktionsgenerator Ausgang die Krokodilklemmen mit. Platzieren Sie die Baugruppe im Inneren des Inkubators die Montage zu ermöglichen, auf die gewünschte Temperatur äquilibrieren.
- Anwenden eines Sinuswellenfrequenz von 10 Hz und einer Potentialdifferenz von 1 V für 90 min. Ensure , die die Spannung V 1 mit einem Multimeter ist wie in Figur 3 gezeigt. Fortsetzung des Prozesses für 90 min.

Abbildung 3:. Die Positionen , an denen die Elektroden vom Multimeter zur Messung von Frequenz und Strom gesetzt werden Bitte hier klicken , um eine größere Version dieser Figur zu sehen.
{kind=link}
- Nach 90 min, langsam verringern die Frequenz 2 Hz und weiter Elektroformation für weitere 10 min. Während dieser Zeit der Ernte in vitro - Translationsreaktionen (Schritte 3,1-3,2).
- Schalten Sie den Funktionsgenerator aus und lassen Sie die Baugruppe abkühlen langsam auf Raumtemperatur (durch den Inkubator Abschalten und Verlassen der Inkubator Tür leicht geöffnet).
- Nach dem Abkühlen ziehen Sie die Kabel an den Generatorvon den Ausziehschienen. GUVs kann sehr zerbrechlich sein und müssen daher sehr vorsichtig behandelt werden. bringen Sie vorsichtig die Montage auf der Bank.
- Bauen Sie die Kammer, indem Sie vorsichtig die obere Rutsche mit einer Pinzette entfernen. Ernten Sie die Saccharose enthaltende Suspension GUVs mit einem Schnitt 1.000 ul Spitze und den Transfer in ein sauberes Röhrchen. Behandeln Sie das Rohr vorsichtig Unterbrechung der GUVs zu verhindern.
- Verwenden Sie die GUVs sofort. GUVs sind für mindestens 90 min nach der Ernte stabil, wenn sie bei Raumtemperatur gelagert.
3. Ernte In - vitro - Translationsreaktionen
- Ernte der in vitro - Translationsreaktionen enthält gewünschten Proteine aus dem roten Kammer einer Mikropipette in ein Rohr verwendet wird .
- Um aggregierte Proteine zu entfernen, drehen Sie die Lysate bei 16.200 × g für 15 min auf einer Tischzentrifuge und vorsichtig den Überstand in ein anderes Röhrchen übertragen.
4. GUV-Bindungsassay
- Unter Anwendung eines Cut Spitze, mischen 51; l von in vitro Reaktionen Übersetzung übersetzt Proteine und 5 ul GUVs in einem Rohr und inkubiere bei Raumtemperatur für 2-3 min enthält.
- Bringen Sie ein PDMS Blatt mit einem kleinen Loch (3-4 mm Durchmesser) auf einem sauberen Deckglas und gewährleisten dichte Abdichtung durch sanft und fest gegen die Deck drücken.
- Legen Sie die 10 & mgr; l Mischung aus Schritt 4.1 in das kleine Loch.
- Bild das Gemisch unter einem inversen epi- oder konfokalen Fluoreszenzmikroskop bei Raumtemperatur.
Hinweis: Um die Verdampfung zu verhindern, kann die Bilderzeugungsraum durch eine Deckglas abgedeckt.
Access restricted. Please log in or start a trial to view this content.
Ergebnisse
Unter Verwendung des obigen Protokolls stellten wir GUVs bestehend aus POPC + POPS + Chol (Molverhältnis: 4,66: 2,33: 3). Diese Zusammensetzung wurde auf etwa reflektieren die PS und Cholesterin Konzentrationen des PM ausgewählt. Robuste und effiziente Bindung von Gag wurde nur beobachtet , wenn Gehirn-PI (4,5) P 2 in die POPC + POPS + Chol - Gemisch (POPC + POPS + Chol + brain-PI (4,5) P 2 [Molverhältnis eingeschlossen wurde : 4: 2: 3: 1] in diesem Beispie...
Access restricted. Please log in or start a trial to view this content.
Diskussion
Der GUV-Bindungstest, wie oben beschrieben stellt eine gute Alternative in Szenarien, in denen andere Protein-Lipid-Interaktion Tests haben ihre Grenzen. Dieser Test ermöglicht es uns, mit nativen Länge Acylketten in der Lipiddoppelschicht Kontext und zwar ohne langwierige Flotation Zentrifugation durch mit hoher Dichte Saccharosegradienten oder andere post-Bindung Verarbeitung von Gag-Lipid-Wechselwirkungen zwischen myristoylierten voller Länge Gag und saure Lipide zu untersuchen Komplexe. Ein weiterer Vorteil ist ,...
Access restricted. Please log in or start a trial to view this content.
Offenlegungen
We declare that we have no conflicting interests.
Danksagungen
Wir möchten, dass Mohammad Saleem, Jing Wu und Krishnan Raghunathan für hilfreiche Diskussionen. Wir danken auch Priya Begani für die Unterstützung während der Dreharbeiten. Diese Arbeit wird von National Institutes of Health gewährt R01 AI071727 (AO) und R01 GM110052 (SLV) unterstützt.
Access restricted. Please log in or start a trial to view this content.
Materialien
| Name | Company | Catalog Number | Comments |
| Digital Multimeter | Meterman | 30XR | A generic multimeter that can measure resistance and volts will serve the purpose. |
| Function Generator | Instek | GFG-8216A | A generic function generator that is capable of generating a sine wave at 10 Hz and 1 V is sufficient. |
| ITO coated glass slides | Delta Technologies, Loveland, CO | CG-90IN | |
| Incubator | Hoefer | Any incubator that can accurately maintain temperature will be sufficient | |
| Vacuum chamber | Nalgene | ||
| Thermomixer R | Eppendorf | 21516-166 | |
| Syringe | Hamilton | 80400 | Gauge 22S, Syringe number 702 |
| PDMS | Sylgard elastomer base kit, Dow-Corning | Sylgard, 184 | |
| RTS 100 Wheat Germ CECF Kit | BiotechRabbit, Berlin, Germany | BR1401001 | |
| DiD | Life Technologies, Carlsbad, CA | D7757 | |
| POPC | Avanti Polar Lipids, Alabaster, AL | 850457C | |
| POPS | Avanti Polar Lipids | 840034C | |
| Cholesterol | Avanti Polar Lipids | 700041P | |
| Brain-PI(4,5)P2 | Avanti Polar Lipids | 840046X |
Referenzen
- Chukkapalli, V., Inlora, J., Todd, G. C., Ono, A. Evidence in support of RNA-mediated inhibition of phosphatidylserine-dependent HIV-1 Gag membrane binding in cells. J Virol. 87 (12), 7155-7159 (2013).
- Chukkapalli, V., Oh, S. J., Ono, A. Opposing mechanisms involving RNA and lipids regulate HIV-1 Gag membrane binding through the highly basic region of the matrix domain. Proc Natl Acad Sci U S A. 107 (4), 1600-1605 (2010).
- Chukkapalli, V., Hogue, I. B., Boyko, V., Hu, W. S., Ono, A. Interaction between the human immunodeficiency virus type 1 Gag matrix domain and phosphatidylinositol-(4,5)-bisphosphate is essential for efficient gag membrane binding. J Virol. 82 (5), 2405-2417 (2008).
- Zhou, W., Parent, L. J., Wills, J. W., Resh, M. D. Identification of a membrane-binding domain within the amino-terminal region of human immunodeficiency virus type 1 Gag protein which interacts with acidic phospholipids. J Virol. 68 (4), 2556-2569 (1994).
- Ono, A., Ablan, S. D., Lockett, S. J., Nagashima, K., Freed, E. O. Phosphatidylinositol (4,5) bisphosphate regulates HIV-1 Gag targeting to the plasma membrane. Proc Natl Acad Sci U S A. 101 (41), 14889-14894 (2004).
- Saad, J. S., et al. Structural basis for targeting HIV-1 Gag proteins to the plasma membrane for virus assembly. Proc Natl Acad Sci U S A. 103 (30), 11364-11369 (2006).
- Shkriabai, N., et al. Interactions of HIV-1 Gag with assembly cofactors. Biochemistry. 45 (13), 4077-4083 (2006).
- Vlach, J., Saad, J. S. Trio engagement via plasma membrane phospholipids and the myristoyl moiety governs HIV-1 matrix binding to bilayers. Proc Natl Acad Sci U S A. 110 (9), 3525-3530 (2013).
- Anraku, K., et al. Highly sensitive analysis of the interaction between HIV-1 Gag and phosphoinositide derivatives based on surface plasmon resonance. Biochemistry. 49 (25), 5109-5116 (2010).
- Llewellyn, G. N., Grover, J. R., Olety, B., Ono, A. HIV-1 Gag associates with specific uropod-directed microdomains in a manner dependent on its MA highly basic region. J Virol. 87 (11), 6441-6454 (2013).
- Inlora, J., Chukkapalli, V., Derse, D., Ono, A. Gag localization and virus-like particle release mediated by the matrix domain of human T-lymphotropic virus type 1 Gag are less dependent on phosphatidylinositol-(4,5)-bisphosphate than those mediated by the matrix domain of HIV-1 Gag. J Virol. 85 (8), 3802-3810 (2011).
- Inlora, J., Collins, D. R., Trubin, M. E., Chung, J. Y., Ono, A. Membrane binding and subcellular localization of retroviral Gag proteins are differentially regulated by MA interactions with phosphatidylinositol-(4,5)-bisphosphate and RNA. MBio. 5 (6), e02202(2014).
- Valentine, K. G., et al. Reverse micelle encapsulation of membrane-anchored proteins for solution NMR studies. Structure. 18 (1), 9-16 (2010).
- Dick, R. A., Goh, S. L., Feigenson, G. W., Vogt, V. M. HIV-1 Gag protein can sense the cholesterol and acyl chain environment in model membranes. Proc Natl Acad Sci U S A. 109 (46), 18761-18766 (2012).
- Dick, R. A., Kamynina, E., Vogt, V. M. Effect of multimerization on membrane association of Rous sarcoma virus and HIV-1 MA proteins. J Virol. 87 (24), 13598-13608 (2013).
- Alfadhli, A., Still, A., Barklis, E. Analysis of human immunodeficiency virus type 1 matrix binding to membranes and nucleic acids. J Virol. 83 (23), 12196-12203 (2009).
- Rodriguez, N., Pincet, F., Cribier, S. Giant vesicles formed by gentle hydration and electroformation: a comparison by fluorescence microscopy. Colloids Surf B Biointerfaces. 42 (2), 125-130 (2005).
- Needham, D., McIntosh, T. J., Evans, E. Thermomechanical and transition properties of dimyristoylphosphatidylcholine/cholesterol bilayers. Biochemistry. 27 (13), 4668-4673 (1988).
- Darszon, A., et al. Reassembly of protein-lipid complexes into large bilayer vesicles: perspectives for membrane reconstitution. Proc Natl Acad Sci U S A. 77 (1), 239-243 (1980).
- Angelova, M. I., Dimitrov, D. S. Liposome electroformation. Faraday Discuss. Chem. Soc. (81), 303-311 (1986).
- Angelova, M. I., Soléau, S., Méléard, P. h, Faucon, F., Bothorel, P. Preparation of giant vesicles by external AC electric fields. Progr Colloid Polymer Sci. 89, 127-131 (1992).
- Stachowiak, J. C., et al. Unilamellar vesicle formation and encapsulation by microfluidic jetting. Proc Natl Acad Sci U S A. 105 (12), 4697-4702 (2008).
- Yamauchi, S., et al. The consensus motif for N-myristoylation of plant proteins in a wheat germ cell-free translation system. FEBS J. 277 (17), 3596-3607 (2010).
- Olety, B., Veatch, S. L., Ono, A. Phosphatidylinositol-(4,5)-Bisphosphate Acyl Chains Differentiate Membrane Binding of HIV-1 Gag from That of the Phospholipase Cdelta1 Pleckstrin Homology Domain. J Virol. 89 (15), 7861-7873 (2015).
- Morales-Penningston, N. F., et al. GUV preparation and imaging: minimizing artifacts. Biochim Biophys Acta. 1798 (7), 1324-1332 (2010).
- Veatch, S. L. Electro-formation and fluorescence microscopy of giant vesicles with coexisting liquid phases. Methods Mol Biol. 398, 59-72 (2007).
- Ayuyan, A. G., Cohen, F. S. Lipid peroxides promote large rafts: effects of excitation of probes in fluorescence microscopy and electrochemical reactions during vesicle formation. Biophys J. 91 (6), 2172-2183 (2006).
- Montes, L. R., et al. Electroformation of giant unilamellar vesicles from native membranes and organic lipid mixtures for the study of lipid domains under physiological ionic-strength conditions. Methods Mol Biol. 606, 105-114 (2010).
- Olety, B., Ono, A. Roles played by acidic lipids in HIV-1 Gag membrane binding. Virus Res. 193, 108-115 (2014).
- Keller, H., Krausslich, H. G., Schwille, P. Multimerizable HIV Gag derivative binds to the liquid-disordered phase in model membranes. Cell Microbiol. 15 (2), 237-247 (2013).
- Carlson, L. A., Hurley, J. H. In vitro reconstitution of the ordered assembly of the endosomal sorting complex required for transport at membrane-bound HIV-1 Gag clusters. Proc Natl Acad Sci U S A. 109 (42), 16928-16933 (2012).
- Gui, D., et al. A novel minimal in vitro system for analyzing HIV-1 Gag-mediated budding. J Biol Phys. 41 (2), 135-149 (2015).
- Prevost, C., et al. IRSp53 senses negative membrane curvature and phase separates along membrane tubules. Nat Commun. 6, 8529(2015).
Access restricted. Please log in or start a trial to view this content.
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten