
É necessária uma assinatura da JoVE para visualizar este conteúdo. Faça login ou comece sua avaliação gratuita.
Method Article
Visualização de HIV-1 Gag vinculação a gigante Unilamelares vesículas (GUV) Membranas
Neste Artigo
Resumo
We illustrate here an in vitro membrane binding assay in which interactions between HIV-1 Gag and lipid membranes are visually analyzed using YFP-tagged Gag synthesized in a wheat germ-based in vitro translation system and GUVs prepared by an electroformation technique.
Resumo
A proteína estrutural do VIH-1, Pr55 da mordaça (ou GAG), liga-se à membrana plasmática em células durante o processo de montagem do vírus. ligação de Gag membrana é um passo essencial para a formação de partículas de vírus, uma vez que um defeito no resultado de ligação à membrana da mordaça em grave comprometimento da produção de partículas virais. Para obter detalhes mecanísticos de interacções da membrana Gag-lipídicos, e métodos in vitro com base em RMN, a pegada de proteína, ressonância de plasma de superfície, a centrifugação dos lipossomas de flutuação, ou grânulo fluorescência ligação de lípidos têm sido desenvolvidos até agora. No entanto, cada um destes métodos in vitro tem as suas limitações. Para ultrapassar algumas destas limitações e proporcionar uma abordagem complementar com os métodos previamente estabelecidos, foi desenvolvido um ensaio in vitro em que as interacções entre o HIV-1 gag e membranas lipídicas ter lugar num ambiente de "célula-like". Neste ensaio, Gag ligação a membranas lipídicas é visualmente analisadas usando-YFP Tagged Gag sintetizado em um germe de trigo baseada no sistema de tradução in vitro e GUVs preparados por uma técnica da eletroformação. Aqui nós descrevemos os antecedentes e os protocolos para obtenção de proteínas da mordaça completos com ácido mirístico e membranas GUV necessárias para o ensaio e para detectar Gag-GUV ligação por microscopia.
Introdução
vírus da imunodeficiência humana tipo 1 (HIV-1) é um vírus com envelope que monta a e gomos a partir da membrana plasmática (PM) na maior parte dos tipos de células. A montagem do HIV-1 partículas de vírus é conduzida por a proteína de núcleo virai de 55 kDa chamado Pr55 Gag (GAG). Gag é sintetizado como um precursor de poliproteina composto por quatro domínios estruturais principais, isto é, matriz, da cápside, nucleocápside, e P6, bem como dois péptidos espaçadores SP1 e SP2. Durante a montagem, o (MA) de domínio matriz é responsável pelo direcionamento de mordaça para o local de montagem, o capsídeo (CA) de domínio medeia interações-Gag Gag, o (NC) de domínio nucleoc�side recruta RNA genômico viral, e fatores do hospedeiro p6 recrutas que auxiliam vírus cisão de partícula a partir da membrana plasmática. Gag também sofre uma modificação co-translacional pela adição de um ácido gordo de 14 carbonos ou radical miristato na sua extremidade N-terminal.
ligação de Gag membrana é um requisito essencial para a montagem viral, uma vez que mutants que são defeituosos na ligação à membrana não produzem partículas virais. Nós e outros autores mostraram que a ligação de gag é mediada por sinais bipartidos dentro do domínio MA membrana: a porção miristato de N-terminal que medeia interacções hidrófobas com a bicamada lipídica e um aglomerado de resíduos básicos dentro do domínio MA denominado como região altamente básica ( HBR) que interage com lípidos ácidas sobre o PM 1-4. Estudos de gag-membrana interacções utilizando a expressão ectópica de polyphosphoinositide 5-fosfatase IV (5ptaseIV), uma enzima que catalisa a hidrólise do fosfolípido phosphatidylinositol- ácida PM-específico (4,5) -bisphosphate [PI (4,5) P 2] a fosfatidilinositol-4-fosfato, em células sugeriram que Gag-AM localização é mediada por PI (4,5) P 2 3,5. No entanto, estudos in vitro visando a entender mais detalhes mecanicistas de Gag-PI (4,5) P 2 interações têm provado ser um desafio para um número de razões. Paraexemplo, a purificação de comprimento completo miristoilada Gag de HIV-1 por experiências bioquímicas tem sido tecnicamente difícil, pelo menos em parte, devido à tendência de Gag para agregar durante a purificação. Assim, as formas truncadas de HIV-1 da mordaça, tais como Myr-MA ou Myr-MA-CA, ou a forma não-miristoiladas têm sido frequentemente utilizada em estudos que necessitam de purificação da mordaça (por exemplo, ressonância magnética nuclear, a pegada de proteína, e a superfície ressonância de plasma de 6-9). Alternativamente, acoplados em reacções de transcrição-tradução in vitro têm sido utilizados para produzir de comprimento completo miristoilada Gag de HIV-1 em outros estudos bioquímicos 1,2. Tipicamente neste sistema, um plasmídeo Gag-codificação é transcrita e traduzida com lisados de células eucarióticas (por exemplo, os lisados de reticulócitos de coelho) que são desprovidas de quaisquer membranas celulares e RNAs mensageiros mas contêm os mecanismos de transcrição e tradução. Após a reacção, os lisados de células que contêm gag são misturados com membranes para análise das interações da mordaça com lipídios. Além da facilidade de preparação de comprimento completo miristoilada Gag, os métodos que utilizam o sistema de tradução in vitro de transcrição têm uma vantagem que a síntese de gag e reacções de ligação à membrana posterior ocorrer num meio 'eucariótica citosol-like ", que podem representar melhor as condições fisiológicas. Esta propriedade contribuiu para os estudos que mostraram que as moléculas de RNA ligadas ao domínio MA regular Gag ligação a lípidos ácidas de forma competitiva 1,2,10-12. No entanto, uma vez que a quantidade total de proteínas Gag obtidos nestes lisados de células não são elevadas, a marcação metabólica das proteínas com aminoácidos marcados radioactivamente é necessário para a sua detecção.
Dependendo do método para medir as interações Gag-lípidos, têm sido utilizados uma variedade de preparações de membrana. Cada um desses métodos tem seus pontos fortes e limitações. A maioria dos ensaios baseados-RMN requerem a utilização de lípidos com acilo curtocadeias que são solúveis em água (por exemplo, C4-C8-e PI (4,5) P 2) 6,8. Enquanto os métodos de RMN de teste de ligação de Gag aos lípidos que têm cadeias de acilo de comprimento encontradas nas células estão a ser desenvolvidos, eles foram usados apenas com MA com ácido mirístico ou nonmyristoylated, até agora, 8,13. Em alternativa, os lipossomas preparados a partir de lípidos que têm cadeias de acilo de comprimento nativos têm sido utilizados em métodos bioquímicos, tais como lipossomas ou flotação fluorescente talão lipossoma ensaios de ligação 2,3,10,14-16. No entanto, os lipossomas utilizados nestes ensaios têm pequenos diâmetros, e, assim, as suas membranas têm curvaturas acentuadas e positivos. Em contraste, durante a fase inicial de montagem de partículas em células de VIH-1 infectados, Gag se liga ao PM, que é praticamente planar na escala de Gag, e, subsequentemente, induz curvatura negativa durante a floração. Portanto, as membranas de lipossomas com curvatura acentuada e positivo pode não ser bicamadas lipídicas ideais para estudar as interações da mordaça-lipídico. Quanto Flot lipossomaensaios ation, outra ressalva potencial é que a exposição de complexos Gag-lipídicas para gradiente de sacarose hipertônica durante a centrifugação pode afetar o resultado experimental. Para atenuar estas limitações e proporcionar um sistema experimental complementar, para ensaios de ligação a GAG de gigantes vesículas unilamelares (GUVs) têm sido desenvolvidos nos últimos anos. GUVs são vesículas única bicamada lipídica cujos diâmetros se estendem a várias dezenas de micrómetros. Assim, a curvatura destas membranas se assemelha a PM na escala de Gag. Além disso, devido ao seu grande tamanho, que permite a inspeção visual sob microscópios ópticos, ligação à membrana proteínas da mordaça de fluorescência marcados ou rotulados com essas vesículas após a mistura pode ser facilmente determinada sem tratamento posterior dos complexos Gag-lipídicas.
Nós aqui descrevemos um protocolo para estudar HIV-1 Gag membrana ligação usando GUVs obtidos a partir de um método eletroformação. Vários métodos, tais como a hidratação suave, hidratação assistida-gel, microfonerofluidic de jacto, e eletroformação 17-22 têm sido usados para obter GUVs. Para o protocolo aqui descrito, o método da eletroformação é utilizado principalmente devido à sua eficiência na formação GUVs lípidos com ácidos e a sua relativa facilidade de utilização, sem a necessidade de instalações dispendiosas. Uma vez que a visualização de Gag necessita de um repórter fluorescente, amarela proteína fluorescente (YFP) está geneticamente adicionado ao C-terminal de Gag (gag-YFP). Proteínas gag-YFP são obtidos por reacções de transcrição in vitro e tradução em lisados de gérmen de trigo com base na tecnologia contínua de permuta de fluxo contínuo (CECF). Nesta tecnologia, tanto a remoção de produtos secundários inibidores das reacções e fornecimento de substratos de reacção e componentes de energia são alcançados de um mecanismo baseado em diálise. Para estas reacções, é usado um plasmídeo que codifica Gag-YFP sob o controlo de um promotor de T7. É de notar, como mostrado anteriormente, lisados de gérmen de trigo apoiar miristoila�o sem componentes adicionais 23,24. Usando este método, foi possível obter quantidades suficientes de comprimento completo miristoilada Gag-YFP para visualização de Gag em membranas GUV 24. Aqui nós descrevemos o protocolo com a qual o HIV-1 Gag ligação a PI (4,5) P 2 molecular contendo membranas GUV podem ser examinados sem processamento subsequente longa seguinte reacções de ligação e propor que este método complementa preexistente Gag-membrana ensaios de ligação e pode ser estendido para entender melhor o HIV-1 Gag-membrana vinculativo.
Access restricted. Please log in or start a trial to view this content.
Protocolo
Dia 1: Expressão de mordaça proteínas utilizando a transcrição in vitro-translation sistema baseado em Lysate germe de trigo
1. Preparação do HIV-1 Gag
- Remove todos os reagentes do kit comercial CECF de gérmen de trigo a partir de congeladores (lisados de gérmen de trigo são armazenadas a -80 ° C; outros reagentes, a -20 ºC). Descongelar-los em gelo e misturar os componentes como apresentados na Tabela 1.
| Mix-1 (alimentação mix) | |
| solução de alimentação | 900 ul |
| Aminoácidos | 80 ul |
| metionina | 20 ul |
| Total | 1.000 l |
| Mistura-2 (Lisado mix) | |
| lisados de gérmen de trigo | 15 ul |
| RNase água livre | 7 ul |
| Aminoácidos | 4 ul |
| metionina | 1 ul |
| O DNA de plasmídeo em tampão TE (1 ug / uL) ** | 4 ul |
| tampão de reacção | 15 ul |
| RNasin * | 4 ul |
| Total | 50 ul |
Tabela 1: Composições de misturas necessárias para reacções de gérmen de trigo.
Nota: Se o experimento foi concebido para examinar o efeito da remoção de ARN por ARNase de Gag de ligação à membrana, substituir inibidores de ribonuclease com água sem RNase. O DNA de plasmídeo deve ser livre de impurezas de acordo com as instruções do fabricante. No entanto, plasmids preparados utilizando técnicas convencionais, mas não, kits de isolamento de plasmídeos endotoxina-livres têm sido utilizados com sucesso. instruções do fabricante também recomenda o uso de DNA dissolvido em água livre de nuclease. Se utilizando ADN suspensas em TE [Tris-HCl a 10 (pH 7,4) contendo EDTA 1 mM], evitar a utilização de concentrações mais baixas de ADN, uma vez que o EDTA em TE pode afectar o rendimento. Os plasmídeos dissolvido em tampão TE a uma gama de concentrações de 0,8-1 mg / mL foram utilizadas com sucesso.
- Primeiro adicione a mistura de alimentação (Mix-1) na cassete de reacção (câmara transparente). Em seguida, adicione a mistura de lisado (Mix-2) para a outra câmara (vermelho). Não introduzir bolhas ao adicionar as misturas nas suas respectivas câmaras, uma vez que podem conduzir a troca ineficaz dos componentes da solução e, portanto, reduzir a eficiência da reacção.
- Cobrir as câmaras com a película adesiva fornecida com o kit.
- Insira a cassete de reação no compartimento de cassete e incubate o conjunto durante 24 horas a 24 ° C a uma velocidade de agitação constante de 900 rpm num Eppendorf Thermomixer R.
Dia 2: Preparar GUVs por eletroformação e colheita de germe de trigo Lysates
2. Preparação de GUVs
- Alíquotas lípidos dissolvidos em solventes inorgânicos tais como o clorofórmio ou misturas de clorofórmio e metanol em frascos de vidro com tampas de rosca forrada com forros de teflon e embrulhados com parafilme. Lidar com lipídios com seringas de vidro Hamilton-tipo, ou se disponível, pontas de plástico de clorofórmio-resistente. Retire os frascos de vidro contendo lípidos para ser usado para preparação de GUV a -20 ºC congelador.
- Uma vez que os frascos são equilibrados até à temperatura ambiente, certifique-se por inspecção visual que as suspensões lipídicas são claras. A qualidade de lípidos, em particular lípidos ácidos, tais como fosfatidilserina palmitoil-oleoil (POP) e PI (4,5) P 2, é importante para experiências bem sucedidas. Não use lipídios que sãoem alíquotas durante mais de 2 meses.
- Pré-aquecer num bloco de aquecimento para a temperatura desejada. Esta temperatura deve ser de pelo menos 5 ° C superior à temperatura de fusão (Tm) do lípido (s) com a mais elevada Tm.
- Calcular os volumes de lípidos necessários de acordo com os rácios molares desejados de lípidos. Para estudar o HIV-1 Gag de ligação aqui, utilizar as seguintes razões molares como mostrado na Tabela 2.
| mistura de lípidos | razão molar | Volume (em mL) | da concentração |
| POPC + POPS + Chol | 46,6 + 23,3 + 30 | 19,74 + 10,18 + 6,57 | POPC, POPS, Chol: 10 mg / ml |
| POPC + POPS + Chol + Brain-PI (4,5) P 2 | 16,11 + 8,3 + 6,25 + 58,19 | POPC, POPS, Chol: 10 mg / ml | |
| Brain-PI (4,5) P 2: 1 mg / mL |
Tabela 2: Volumes de diferentes lipídios utilizada para obter GUVs.
- Adicione os volumes desejados de lípidos em um frasco de vidro com tampa de rosca limpo.
- Lâminas de dimensões 25 x 50 x 1,1 mm3 e resistência 70-100 co (como descritos no catálogo) Uso de ITO revestido. Coloque duas lâminas ITO-revestidos de uma bancada limpa. Cada câmara eletroformação requer dois-tin-óxido de índio (ITO) lâminas de vidro -Revestido. Verificar que o lado condutora da lâmina de vidro é virada para cima, verificando a resistência utilizando um multímetro. Ele deve mostrar um valor de cerca de 200 Ω. Certifique-se a superfície das lâminas são limpos, limpando suavemente com 70% de etanol usando toalhetes sem fiapos.
- Limpar a superfície exterior de uma agulha de uma seringa por RInsing com clorofórmio e colocar a seringa no bloco de calor.
- Colocar uma nova lâmina de vidro revestido de ITO-no bloco de calor com o lado voltado para cima condutora e permitir que ele para equilibrar até à temperatura desejada geralmente cerca de 2-3 min. Não reutilize as lâminas de vidro ITO-revestidos para um risco potencial que o ITO-revestimento nas lâminas de vidro pode ser danificado durante procedimentos de limpeza.
- Use um volume de 40-50 mL da mistura de lípidos para cada slide para uniforme e ainda rápida divulgação dos lipídios sobre a lâmina de vidro. Ajustar o volume total com um solvente orgânico apropriado tal como clorofórmio.
- Coloque uma metade do volume total da mistura de lípidos na lâmina de vidro e espalhou imediatamente o lípido usando a seringa limpa (passo 2.6) para dar uma película lipidica uniforme (movendo a agulha 2-3 vezes em toda a corrediça). Espalhe o filme lipídico gentilmente (mas rapidamente) para evitar qualquer dano ao revestimento ITO fina. Não uniforme difundir ou espalhar áspera pode levar a sub-óptimarendimentos ou heterogeneidade computacional de GUVs.
- Portanto, certifique-se disseminação uniforme inspecionando o filme lipídico após espalhar antes de prosseguir para a próxima etapa. Se a opacidade do filme parece excessivamente não uniforme, refazer o procedimento usando uma nova lâmina.
- Colocar a lâmina revestido dentro de um prato de Petri com o lado revestido voltado para cima.
- Repita os passos de 2,6-2,9 para o resto da mistura utilizando uma outra lâmina de ITO revestido.
- Colocar a placa de petri contendo ambos os diapositivos numa câmara de vácuo convencional dessecação durante 60-90 minutos para remover quaisquer vestígios de solventes orgânicos.
- Durante este processo, definido na incubadora para a mesma temperatura que a do bloco de aquecimento (65 ° C na maior parte dos casos) e pré-aquecer a 300 mM de uma solução de sacarose na incubadora.
- Após a secagem, trazer o prato de petri para o banco e colocar a lâmina de vidro sobre uma superfície limpa.
- Suavemente limpar uma junta de PDMS (Figura 1) usando 70% de etanol e seca-lo completely.
- Colocá-lo sobre a lâmina de vidro revestida de lípido, como mostrado na Figura 1 e suavemente e pressione firmemente de modo a que as formas de vedação uma vedação estanque à água. Confirmar a ausência de lacunas entre a junta PDMS eo slide. Isto resulta na formação de uma câmara superficial que contém a solução de sacarose.

Figura 1:. O layout da lâmina de vidro ITO revestido usado para eletroformação Por favor clique aqui para ver uma versão maior desta figura.
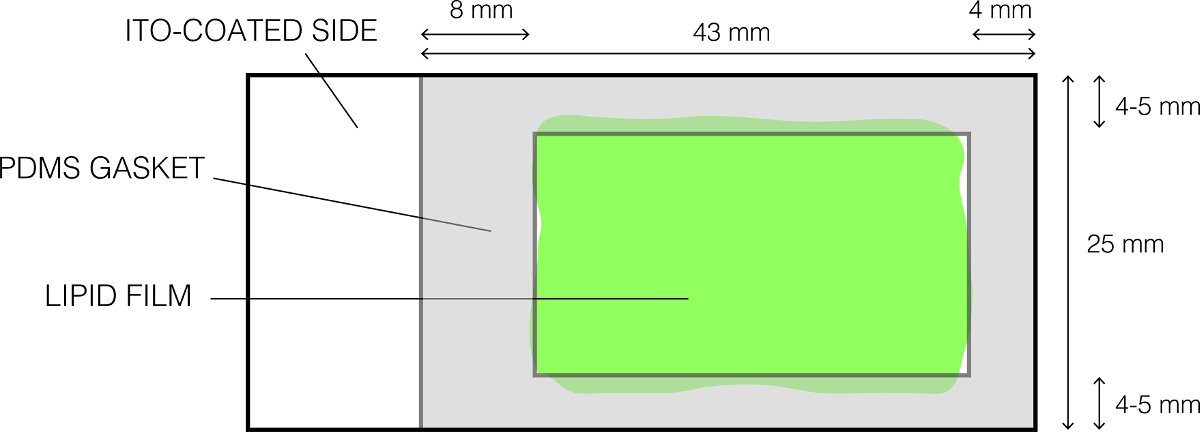{kind=link}
- Encha a câmara com solução de sacarose pré-aquecido, sem quaisquer bolhas.
- Coloque a outra lâmina revestida em cima para fazer um selo apertado. Certifique-se que não há bolhas. A montagem final deve aparecer como mostrado na Figura 2. Aperte o usin câmaraclips g Binder.

Figura 2:. Representação esquemática da câmara de eletroformação (vista lateral) Por favor clique aqui para ver uma versão maior desta figura.
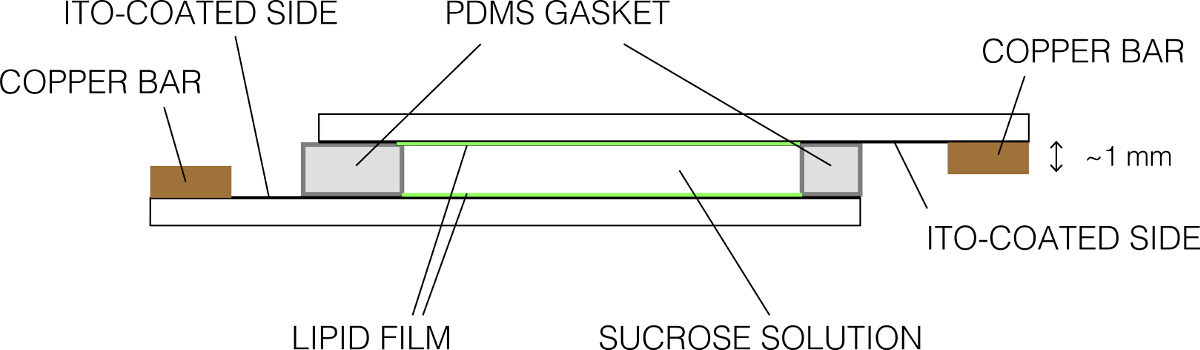{kind=link}
- Retire o excesso de sacarose por aspiração e limpar os slides usando toalhetes sem fiapos. Apertem os slides para a barra de cobre para que os lados condutores estão em contato uniforme com a barra. Certifique-se de que apenas um lado condutor da placa de vidro está em contacto com uma barra de cobre.
- Conectar-se a barra de cobre para a saída do gerador de função usando as garras jacaré. Coloque o conjunto dentro da incubadora para permitir que o conjunto para equilibrar com a temperatura desejada.
- Aplique uma freqüência de onda senoidal de 10 Hz e uma diferença de potencial de 1 V por 90 min. assegue que a tensão é 1 V por um multímetro conforme mostrado na Figura 3. Continuar o processo durante 90 min.

Figura 3:. As posições em que os eléctrodos do multímetro são colocados para medir a frequência e corrente Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
- Após 90 min, lentamente diminuir a frequência de 2 Hz e continuar eletroformação durante mais 10 min. Durante este tempo, a colheita em reacções de tradução in vitro (passos 3,1-3,2).
- Desligue o gerador de função e permitir que o conjunto arrefecer lentamente à temperatura ambiente (desligando a incubadora e deixando a porta da incubadora ligeiramente aberta).
- Após o resfriamento, desconecte os cabos do geradordos conjuntos deslizantes. GUVs pode ser muito frágil e, portanto, precisa ser manuseado com muito cuidado. trazer cuidadosamente o conjunto para o banco.
- Desmonte a câmara, removendo suavemente o cursor superior usando uma pinça. Colheita das suspensões contendo GUVs sacarose utilizando uma ponta ul corte 1.000 e transferir para um tubo limpo. Lidar com o tubo suavemente para evitar a interrupção de GUVs.
- Use as GUVs imediatamente. GUVs são estáveis durante pelo menos 90 minutos após a colheita, quando armazenados à temperatura ambiente.
3. Reacções colheita tradução in vitro
- Colher as reacções de tradução in vitro contendo proteínas desejadas a partir da câmara de vermelho utilizando uma micropipeta para um tubo.
- Para remover proteínas agregadas, girar os lisados a 16200 xg durante 15 min numa centrífuga de mesa e transferir cuidadosamente o sobrenadante para outro tubo.
4. ensaio de ligação GUV
- Usando uma ponta de corte, misturar 51; l de reacções de tradução in vitro contendo proteínas traduzidas e 5 ul de GUVs em um tubo e incubar à temperatura ambiente durante 2-3 minutos.
- Anexar uma folha de PDMS com um pequeno furo (3-4 mm de diâmetro) em uma lamela limpa e garantir a vedação estanque, pressionando-o suavemente e firmemente contra a lamela.
- Coloque a mistura de 10 ul a partir do passo 4.1 no pequeno orifício.
- Imagem a mistura sob um microscópio de fluorescência invertido confocal de epi ou à temperatura ambiente.
Nota: Para evitar a evaporação, a câmara de imagem pode ser coberto por uma lamela.
Access restricted. Please log in or start a trial to view this content.
Resultados
Usando o protocolo acima, preparamos GUVs compostos por POPC + + POPS Chol (razão molar: 4,66: 2,33: 3). Esta composição foi escolhida para aproximadamente reflectir as concentrações de colesterol e de PS PM. Robusto e eficiente ligação de Gag foi observada somente quando o cérebro-PI (4,5) P 2 foi incluída na mistura POPC + POPS + Chol-PI cerebral (4,5) P 2 [razão molar (POPC + POPS + Chol + : 4: 2: 3: 1], neste exemplo) (Figura 4, ...
Access restricted. Please log in or start a trial to view this content.
Discussão
O ensaio de ligação GUV como descrito acima fornece uma boa alternativa em situações onde outros ensaios de interação proteína-lipídica têm as suas limitações. Este ensaio nos permite examinar as interações entre ácido mirístico Gag de corpo inteiro e lipídios ácidos com cadeias acilo de comprimento nativa no contexto bicamada lipídica e fazê-lo sem longas centrifugação flotação por meio de alta densidade gradientes de sacarose ou outro processamento pós-obrigatório de Gag-lipídico complexos. O...
Access restricted. Please log in or start a trial to view this content.
Divulgações
We declare that we have no conflicting interests.
Agradecimentos
Nós gostaríamos de agradecer Mohammad Saleem, Jing Wu e Krishnan Raghunathan para discussões úteis. Agradecemos também Priya Begani de assistência durante as filmagens. Este trabalho é apoiado pelos Institutos Nacionais de Saúde concede R01 AI071727 (para AO) e R01 GM110052 (a SLV).
Access restricted. Please log in or start a trial to view this content.
Materiais
| Name | Company | Catalog Number | Comments |
| Digital Multimeter | Meterman | 30XR | A generic multimeter that can measure resistance and volts will serve the purpose. |
| Function Generator | Instek | GFG-8216A | A generic function generator that is capable of generating a sine wave at 10 Hz and 1 V is sufficient. |
| ITO coated glass slides | Delta Technologies, Loveland, CO | CG-90IN | |
| Incubator | Hoefer | Any incubator that can accurately maintain temperature will be sufficient | |
| Vacuum chamber | Nalgene | ||
| Thermomixer R | Eppendorf | 21516-166 | |
| Syringe | Hamilton | 80400 | Gauge 22S, Syringe number 702 |
| PDMS | Sylgard elastomer base kit, Dow-Corning | Sylgard, 184 | |
| RTS 100 Wheat Germ CECF Kit | BiotechRabbit, Berlin, Germany | BR1401001 | |
| DiD | Life Technologies, Carlsbad, CA | D7757 | |
| POPC | Avanti Polar Lipids, Alabaster, AL | 850457C | |
| POPS | Avanti Polar Lipids | 840034C | |
| Cholesterol | Avanti Polar Lipids | 700041P | |
| Brain-PI(4,5)P2 | Avanti Polar Lipids | 840046X |
Referências
- Chukkapalli, V., Inlora, J., Todd, G. C., Ono, A. Evidence in support of RNA-mediated inhibition of phosphatidylserine-dependent HIV-1 Gag membrane binding in cells. J Virol. 87 (12), 7155-7159 (2013).
- Chukkapalli, V., Oh, S. J., Ono, A. Opposing mechanisms involving RNA and lipids regulate HIV-1 Gag membrane binding through the highly basic region of the matrix domain. Proc Natl Acad Sci U S A. 107 (4), 1600-1605 (2010).
- Chukkapalli, V., Hogue, I. B., Boyko, V., Hu, W. S., Ono, A. Interaction between the human immunodeficiency virus type 1 Gag matrix domain and phosphatidylinositol-(4,5)-bisphosphate is essential for efficient gag membrane binding. J Virol. 82 (5), 2405-2417 (2008).
- Zhou, W., Parent, L. J., Wills, J. W., Resh, M. D. Identification of a membrane-binding domain within the amino-terminal region of human immunodeficiency virus type 1 Gag protein which interacts with acidic phospholipids. J Virol. 68 (4), 2556-2569 (1994).
- Ono, A., Ablan, S. D., Lockett, S. J., Nagashima, K., Freed, E. O. Phosphatidylinositol (4,5) bisphosphate regulates HIV-1 Gag targeting to the plasma membrane. Proc Natl Acad Sci U S A. 101 (41), 14889-14894 (2004).
- Saad, J. S., et al. Structural basis for targeting HIV-1 Gag proteins to the plasma membrane for virus assembly. Proc Natl Acad Sci U S A. 103 (30), 11364-11369 (2006).
- Shkriabai, N., et al. Interactions of HIV-1 Gag with assembly cofactors. Biochemistry. 45 (13), 4077-4083 (2006).
- Vlach, J., Saad, J. S. Trio engagement via plasma membrane phospholipids and the myristoyl moiety governs HIV-1 matrix binding to bilayers. Proc Natl Acad Sci U S A. 110 (9), 3525-3530 (2013).
- Anraku, K., et al. Highly sensitive analysis of the interaction between HIV-1 Gag and phosphoinositide derivatives based on surface plasmon resonance. Biochemistry. 49 (25), 5109-5116 (2010).
- Llewellyn, G. N., Grover, J. R., Olety, B., Ono, A. HIV-1 Gag associates with specific uropod-directed microdomains in a manner dependent on its MA highly basic region. J Virol. 87 (11), 6441-6454 (2013).
- Inlora, J., Chukkapalli, V., Derse, D., Ono, A. Gag localization and virus-like particle release mediated by the matrix domain of human T-lymphotropic virus type 1 Gag are less dependent on phosphatidylinositol-(4,5)-bisphosphate than those mediated by the matrix domain of HIV-1 Gag. J Virol. 85 (8), 3802-3810 (2011).
- Inlora, J., Collins, D. R., Trubin, M. E., Chung, J. Y., Ono, A. Membrane binding and subcellular localization of retroviral Gag proteins are differentially regulated by MA interactions with phosphatidylinositol-(4,5)-bisphosphate and RNA. MBio. 5 (6), e02202(2014).
- Valentine, K. G., et al. Reverse micelle encapsulation of membrane-anchored proteins for solution NMR studies. Structure. 18 (1), 9-16 (2010).
- Dick, R. A., Goh, S. L., Feigenson, G. W., Vogt, V. M. HIV-1 Gag protein can sense the cholesterol and acyl chain environment in model membranes. Proc Natl Acad Sci U S A. 109 (46), 18761-18766 (2012).
- Dick, R. A., Kamynina, E., Vogt, V. M. Effect of multimerization on membrane association of Rous sarcoma virus and HIV-1 MA proteins. J Virol. 87 (24), 13598-13608 (2013).
- Alfadhli, A., Still, A., Barklis, E. Analysis of human immunodeficiency virus type 1 matrix binding to membranes and nucleic acids. J Virol. 83 (23), 12196-12203 (2009).
- Rodriguez, N., Pincet, F., Cribier, S. Giant vesicles formed by gentle hydration and electroformation: a comparison by fluorescence microscopy. Colloids Surf B Biointerfaces. 42 (2), 125-130 (2005).
- Needham, D., McIntosh, T. J., Evans, E. Thermomechanical and transition properties of dimyristoylphosphatidylcholine/cholesterol bilayers. Biochemistry. 27 (13), 4668-4673 (1988).
- Darszon, A., et al. Reassembly of protein-lipid complexes into large bilayer vesicles: perspectives for membrane reconstitution. Proc Natl Acad Sci U S A. 77 (1), 239-243 (1980).
- Angelova, M. I., Dimitrov, D. S. Liposome electroformation. Faraday Discuss. Chem. Soc. (81), 303-311 (1986).
- Angelova, M. I., Soléau, S., Méléard, P. h, Faucon, F., Bothorel, P. Preparation of giant vesicles by external AC electric fields. Progr Colloid Polymer Sci. 89, 127-131 (1992).
- Stachowiak, J. C., et al. Unilamellar vesicle formation and encapsulation by microfluidic jetting. Proc Natl Acad Sci U S A. 105 (12), 4697-4702 (2008).
- Yamauchi, S., et al. The consensus motif for N-myristoylation of plant proteins in a wheat germ cell-free translation system. FEBS J. 277 (17), 3596-3607 (2010).
- Olety, B., Veatch, S. L., Ono, A. Phosphatidylinositol-(4,5)-Bisphosphate Acyl Chains Differentiate Membrane Binding of HIV-1 Gag from That of the Phospholipase Cdelta1 Pleckstrin Homology Domain. J Virol. 89 (15), 7861-7873 (2015).
- Morales-Penningston, N. F., et al. GUV preparation and imaging: minimizing artifacts. Biochim Biophys Acta. 1798 (7), 1324-1332 (2010).
- Veatch, S. L. Electro-formation and fluorescence microscopy of giant vesicles with coexisting liquid phases. Methods Mol Biol. 398, 59-72 (2007).
- Ayuyan, A. G., Cohen, F. S. Lipid peroxides promote large rafts: effects of excitation of probes in fluorescence microscopy and electrochemical reactions during vesicle formation. Biophys J. 91 (6), 2172-2183 (2006).
- Montes, L. R., et al. Electroformation of giant unilamellar vesicles from native membranes and organic lipid mixtures for the study of lipid domains under physiological ionic-strength conditions. Methods Mol Biol. 606, 105-114 (2010).
- Olety, B., Ono, A. Roles played by acidic lipids in HIV-1 Gag membrane binding. Virus Res. 193, 108-115 (2014).
- Keller, H., Krausslich, H. G., Schwille, P. Multimerizable HIV Gag derivative binds to the liquid-disordered phase in model membranes. Cell Microbiol. 15 (2), 237-247 (2013).
- Carlson, L. A., Hurley, J. H. In vitro reconstitution of the ordered assembly of the endosomal sorting complex required for transport at membrane-bound HIV-1 Gag clusters. Proc Natl Acad Sci U S A. 109 (42), 16928-16933 (2012).
- Gui, D., et al. A novel minimal in vitro system for analyzing HIV-1 Gag-mediated budding. J Biol Phys. 41 (2), 135-149 (2015).
- Prevost, C., et al. IRSp53 senses negative membrane curvature and phase separates along membrane tubules. Nat Commun. 6, 8529(2015).
Access restricted. Please log in or start a trial to view this content.
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados