
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
ניצול במגרש משחררים אתילן, 2-Chloroethylphosphonic חומצה, ככלי לחקר תגובה אתילן בחיידקים
In This Article
Summary
The protocols outlined herein facilitate the convenient investigation of bacterial ethylene responses by utilizing 2-chloroethylphosphonic acid (CEPA). Ethylene is produced in situ through the decomposition of CEPA in an aqueous bacterial growth medium, circumventing the requirement for pure ethylene gas.
Abstract
Ethylene (C2H4) is a gaseous phytohormone that is involved in numerous aspects of plant development, playing a dominant role in senescence and fruit ripening. Exogenous ethylene applied during early plant development triggers the triple response phenotype; a shorter and thicker hypocotyl with an exaggerated apical hook. Despite the intimate relationship between plants and bacteria, the effect of exogenous ethylene on bacteria has been greatly overlooked. This is partly due to the difficulty of controlling gaseous ethylene within the laboratory without specialized equipment. 2-Chloroethylphosphonic acid (CEPA) is a compound that decomposes into ethylene, chlorine, and phosphate in a 1:1:1:1 molar ratio when dissolved in an aqueous medium of pH 3.5 or greater. Here we describe the use of CEPA to produce in situ ethylene for the investigation of ethylene response in bacteria using the fruit-associated, cellulose-producing bacterium Komagataeibacter xylinus as a model organism. The protocols described herein include both the verification of ethylene production from CEPA via the Arabidopsis thaliana triple response assay and the effects of exogenous ethylene on K. xylinus cellulose production, pellicle properties and colonial morphology. These protocols can be adapted to examine the effect of ethylene on other microbes using appropriate growth media and phenotype analyses. The use of CEPA provides researchers with a simple and efficient alternative to pure ethylene gas for the routine determination of bacterial ethylene response.
Introduction
האתילן אולפינים (C 2 H 4) התגלה לראשונה כהורמון צמח בשנת 1901 כאשר נצפתה כי השתילים אפונה, שגודלו במעבדה כי השתמשו במנורות גז פחם, הציג מורפולוגיה לא תקינה אשר נובעת (hypocotyls) היו קצרים, עבה והתכופף הצידה לעומת שתילי אפונה נורמלים; פנוטיפ מאוחר יותר כינה את 1,2 בתגובה המשולש. מחקרים מאוחרים יותר הראו כי אתילן הוא phytohormone חיוני המווסת תהליכים התפתחותיים רבים כגון צמיחה, תגובת הלחץ, הבשלת הפרי והזדקנות 3. ארבידופסיס thaliana, אורגניזם מודל למחקר בביולוגיה צמח, נחקרה גם לגבי תגובתה אתילן. מוטנטים בתגובת אתילן כמה בודדו על ידי ניצול פנוטיפ בתגובה המשולש שנצפה א כהה המבוגר שתילי thaliana בנוכחות אתילן 1,4,5. מבשר biosynthetic לייצור אתילן בצמחים הוא 1-אחומצת קרבוקסילית minocyclopropane (ACC) 6 והוא נפוץ במהלך assay בתגובה המשולש להגדיל ייצור אתילן אנדוגני שמוביל 1,4,5 פנוטיפ בתגובה המשולש.
למרות התגובה אתילן הוא למד נרחב בצמחים, ההשפעה של אתילן אקסוגניים על חיידקים אילתר בהרבה למרות מקירבת חיידקים עם צמחים. מחקר אחד דיווח כי זנים מסוימים Pseudomonas יכולים לשרוד באמצעות אתילן כמקור בלעדי של פחמן ואנרגיה 7. עם זאת, רק שני מחקרים הראו כי חיידקים להגיב אתילן. המחקר הראשון הראה כי זנים של Pseudomonas aeruginosa, פ fluorescens, פ putida, ופ ' syringae היו chemotactic כלפי אתילן באמצעות assay התקע agarose שבו agarose מותכת עורבב עם חיץ chemotaxis equilibrated בגז אתילן טהור 8. עם זאת, למיטב ידיעתנו, לא חל Furthדוחות אה באמצעות גז אתילן טהור לאפיין בתגובת אתילן חיידקים, ככל הנראה בשל המתקשים להתמודד עם גזים במעבדה ללא ציוד מיוחד. הדו"ח השני של תגובת אתילן חיידקים הראה כי גדל אתילן ייצור תאי חיידקי ביטוי גנים מושפע חיידק הפרות קשורים, Komagataeibacter (לשעבר Gluconacetobacter) xylinus 9. במקרה זה, במתחם שחרור אתילן, חומצה 2-chloroethylphosphonic (CEPA) שימש ליצירת אתילן באתרם בתוך המדיום התפתחות חיידקים, תוך עקיפת הצורך גז אתילן טהור או ציוד מיוחד.
CEPA מייצרת אתילן ביחס של 1: 1 יחס טוחן מעל pH 3.5 10,11 באמצעות תגובת בסיס-זרז, מסדר הראשון 12 - 14. השפלתו של CEPA מתואם חיובית עם pH וטמפרטורה 13,14 ותוצאות בייצור אתילפנטן, כלוריד ופוספט. CEPA מספקת לחוקרים המעוניינים ללמוד תגובות חיידקים אתילן עם אלטרנטיבה נוחה אתילן גזים.
המטרה הכללית של הפרוטוקולים הבאים היא לספק שיטה פשוטה ויעילה ללמוד בתגובה אתילן בקטריאלי כולל אימות של רמות רלוונטי מבחינה פיזיולוגית של ייצור אתילן מן הפירוק CEPA במדיום התפתחות חיידקים, ניתוח של pH תרבות על מנת להבטיח CEPA הפירוק אינו פגום במהלך התפתחות חיידקים, והערכת ההשפעה של אתילן על מורפולוגיה פנוטיפ חיידקים. אנו מדגימים פרוטוקולים אלה באמצעות ק xylinus, לעומת זאת, יכולים להיות מותאם הפרוטוקולים הללו ללמוד בתגובה אתילן בחיידקים אחרים על ידי שימוש במדיום הגידול המתאים פנוטיפ המנתח.
Access restricted. Please log in or start a trial to view this content.
Protocol
1. כימיקלים
- הכן פתרון של 500 מ"מ CEPA (144.49 g / mol), ופתרון המורכבת משני 500 mM NaCl (58.44 g / mol) ו -500 מ"מ לאא 2 PO 4 · H 2 O (137.99 g / mol) ב acidified (pH 2.5) מים טהורים או 0.1 N HCl. מערבבים בעזרת מערבולת עד הפתרונות ברורים.
- סדרתי לדלל (10x) הפתרונות 500 מ"מ באותו ממס להשיג 5 מ"מ ו -50 מ"מ מניות.
- הכן פתרון 10 מ"מ של חומצה קרבוקסילית 1-aminocyclopropane (ACC; 101.1 g / mol) במים טהורים.
- מסנן לעקר פתרונות המניות aliquots ולאחסן ב -20 מעלות צלזיוס.
- מסנן לעקר cellulase. aliquots חנות ב 4 מעלות צלזיוס.
2. הפקת אתילן אימות מ- 2-Chloroethylphosphonic חומצה פירוק: Assay תגובה טריפל
- משטח לעקר ארבידופסיס thaliana זרעי אקוטיפ קולומביה שימוש בשיטת החמקן פאזיים:
זהירות: השלב הבא מייצרת tגז כלור oxic. לנהל עיקור זרע במנדף.- השג מיכל סגר עמוק מספיק כדי להכיל כוס זכוכית 250 מ"ל עבור עיקור זרע ולמקם אותו במנדף.
- להוסיף א זרעים thaliana לצינור microcentrifuge ובמקום הצינור בתוך ארון תקשורת. מניח את המתלה המכילה את הצינור הפתוח לתוך מכל הסגר.
הערה: אין למלא צינורות בודדים יותר מחצי מיכל לאפשר גז כלור לחדור את הזרעים הנמוכים. - מניחים כוס 250 מ"ל המכילה 100 מ"ל של אקונומיקה מסחרי במיכל רב סגר. בזהירות להוסיף 3 מ"ל של HCl מרוכזת אל אקונומיקה ומיד לאטום את מיכל עם מכסה שלה.
זהירות: Bleach ו HCl להגיב להפיק גז כלור רעיל הפועל משטח לעקר את הזרעים. - דגירת הזרעים בנוכחות גז כלור עבור 4 שעות ב fumehood.
הערה: פעמי עיקור ארוכות תפחתנה יעילות נביטה. - לאחר העיקור, לאפשר גגז hlorine לפרוק את במנדף לפחות שעה 1 אז לאטום את הצינור microcentrifuge. השאירו את אקונומיקה תערובת HCl ב fumehood למשך 24 שעות לפחות לפני השלכת.
הערה: זרעים מפוסטרים ניתן לאחסן בטמפרטורת חדר לשימוש מיידי. - שמור זרעים ב 4 ° C עבור אחסון לטווח ארוך. תביאו זרעים לטמפרטורת החדר לפני פתיחת צינור microcentrifuge כדי למנוע התעבות. זרעי חנות בחושך.
- להכין צלחות אגר במנות 4 הסקטור פטרי (90 x 15 מ"מ):
- הכן 110 מ"ל של מדיום הגידול עבור א thaliana שתילים: Murashige 1x ו Skoog (MS) בינוני בסיס 15 (4.33 גר '/ ל') המכיל 1% (w / v) סוכרוז 0.8% (w / v) אגר. התאם המדיום MS ל- pH 6 עם NaOH.
- כן 100 מיליליטר של מדיום גידול חיידקי פירוק CEPA: שרם Hestrin (SH) בינוני 16 המכיל 1.5% (w / v) אגר. התאם המדיום SH ל- pH 7 עם NaOH.
- לעקר תקשורת ידי מעוקר ו בטמפהr באמבט מים C ° 55.
- לאחר אגר MS יש מזג, להוסיף 40 מ"ל בבקבוק סטרילי. כדי להכין את מדיום גידול בקרה החיובי עבור א שתילי thaliana, להשלים את aliquot 40 מ"ל של אגר MS עם 40 μl של 10 מ"מ ACC כדי לקבל ריכוז סופי של 10 מיקרומטר ACC.
- הוסף 5 מ"ל של מדיום עד הרביעים המתאים של צלחות פטרי סקטוריאלית (איור 1) ולאפשר אגר כדי לחזק. הכן את כל הצלחות בשלושה עותקים.
הערה: CEPA לא מתווסף את מדיום הגידול עד מאוחר בפרוטוקול.

איור 1:. התקנה של צלחות אגרו לשמש עבור assay בתגובה המשולש עם CEPA סכימטי ממחיש את הרביעים ספציפיים השליטה השלילית (א '), בקרה חיובית (B), וצלחות ניסוי (C). נתון זה יש הבדל בין Augimeri ו רצועת 9. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
{kind=link}
- לרבד א זרעים thaliana כדי להבטיח נביטה סינכרוני:
- להוסיף כחמישים א זרעים thaliana על בכל רבע המכיל אגר MS או MS + ACC. ודא זרעים מופצים באופן שווה כדי להקל שתיל הסרה וניתוח.
- דגירה צלחות המכילות זרעים בחושך ב 4 ° C במשך 3-4 ימים.
- לפרק CEPA על בקטריאלי צמיחה בינוני (SH) ולבצע Assay תגובה טריפל:
- לאחר ריבוד, לחשוף את הזרעים לאור ניאון עבור שעה 2.
- מורחים 10 μl של פתרון המניות 500 מ"מ CEPA על הרביעים של הצלחות ניסיוני המכיל אגר SH (pH 7; איור 1) כדי להשיג ריכוז CEPA הסופישל 1 מ"מ.
- חותם את הצלחות עם סרט מעבדה ומכסים בנייר כסף כדי ליצור סביבה בחושך במשך הזרעים.
- לנבוט זרעים על ידי דוגרי צלחות בחושך ב 23 מעלות צלזיוס במשך 3 ימים עם הצד אגר למטה.
הערה: צלחות יכולים להערם, אבל שולטת שלילי צריך להיות ממוקם בחלק התחתון מאז אתילן קל מהאוויר.
- לנתח נתוני Assay תגובת טריפל:
- עם מלקחי להבת מעוקרים, להסיר שתילי יחיד מתוך צלחת המתאימה לכל אחד טיפול ונוף תחת לנתח או מיקרוסקופ USB הדיגיטלי. ודא התחתון של השתילים מיושרים ולצלם.
- הסר 30 שתילים מכל רביע (60 שתילים לכל לשכפל ביולוגי 180 שתילים לכל טיפול). יישר על משטח עם רקע שחור ולצלם עם סרגל.
- מדוד את אורך hypocotyl (מ"מ) של שתילים לשכפל באמצעות ImageJ תוכנה 17. הגדר את קנה המידה על ידי לחיצה וגרירה של * ישר *כלי לבחור באורך של 10 מ"מ. בחר "סולם גדר" תחת הלשונית "לנתח" ולהגדיר את המרחק הידוע עד 10. שימוש * הבדיקה המחולקת * הכלי, לחץ וגרור כדי לבחור את אורך M hypocotyl ולחץ למדידת מרחק. השווה את האמצעי ביולוגי משכפל באמצעות ANOVA חד כיווני עם מבחן ההשוואה הנפוץ של Tukey. הבדלים נחשבים משמעותי אם p <0.05.
3. ניתוח של pH לאורך התפתחות חיידקים
- לגדול ולכמת Komagataeibacter תרבויות Starter xylinus בשלושה עותקים:
- לחסן מושבה אחת של ק xylinus לתוך 5 מ"ל של מדיום SH (pH 5) בתוספת 0.2% (v / v) cellulase מעוקר-מסנן. דגירה תרבויות על 30 מעלות צלזיוס עם תסיסה ב 150 סל"ד עד OD 600 של 0.3 עד 0.4 הוא הגיע (כ -72 שעות).
- תרבויות המתנע קציר על ידי צנטריפוגה (2,000 XG; 4 מעלות צלזיוס; 10 דקות). עם 5 מ"ל סטרילי 0.85% (w / v) NACפתרון l, לשטוף את התאים פעמיים resuspend התא גלולה. שמור את התאים על הקרח.
- לכמת תאים באמצעות תא ספירת Petroff-אוסר.
- לחסן תרבויות עבור ניתוח של pH:
- להוסיף 150 מ"ל של מרק SH (pH 7) בתוספת 0.2% (v / v) cellulase מעוקרות מסנן עד שתים עשרה 500 מ"ל צלוחיות עם מכסים בנייר כסף. לחסן צלוחיות עם ק תרבויות המתנע xylinus בריכוז של 10 5 תאים / מ"ל. שימוש תרבויות השלוש המתנע, להכין שלושה ביולוגים משכפל לכל טיפול.
- מוסף תרבויות עם 300 μl של 5, 50, או 500 מ"מ CEPA פתרונות המניות להשיג ריכוזים CEPA הסופי של 0.01, 0.1 ו -1.0 מ"מ, בהתאמה. מוסף תרבויות קבוצת הביקורת עם 300 μl של הממס משמש כדי לפזר CEPA.
- חותם את צלוחיות ידי בחוזקה מקליטה את המכסים בנייר כסף כדי צלוחיות דגירה תרבויות במשך 14 ימים ב 30 מעלות צלזיוס עם תסיסה ב 150 סל"ד.
- כל יום, aseptically להסיר 5 מיליליטר דגימות מכל תאי בקבוק גלול על ידי צנטריפוגה (2,000 XG; 4 מעלות צלזיוס; 10 דקות). העבר supernatants לצינור נקי למדוד את רמת החומציות של כל לשכפל ביולוגי באמצעות מד pH.
- לנתח את נתוני מנות עת על ידי גרפים ה- pH הממוצע של המשכפל הביולוגי.
הערה: חשוב כי pH התרבות לא לרדת מתחת 3.5; חומציות תרבות של 5 מפחית שחרור אתילן משמעותי מן CEPA, ו- pH מתחת 3.5 מעכב שחרור אתילן לחלוטין. - כדי לשלוט על רמות כלור פוספט, עורכים ניסוי זהה באמצעות 0.01, 0.1, ו -1.0 מ"מ של הפתרון NaCl-לאא 2 PO 4 באמצעות 5, 50, ו 500 מניות מ"מ, בהתאמה.
4. מורפולוגיה המושבה
- לגדול ק תרבויות Starter xylinus בשלושה עותקים:
- לחסן מושבה אחת של ק xylinus לתוך 5 מ"ל של מדיום SH (pH 5) בתוספת 0.2% (v / v) cellulase מעוקר-מסנן. אינקובטוריםte תרבויות על 30 מעלות צלזיוס עם תסיסה ב 150 סל"ד עד OD 600 של 0.3 עד 0.4 הוא הגיע (כ -72 שעות).
- תרבויות המתנע קציר על ידי צנטריפוגה (2,000 XG; 4 מעלות צלזיוס; 10 דקות). עם 5 מ"ל של תמיסת מלח סטרילית, לשטוף את התאים ולאחר מכן resuspend גלולה התא. שמור את התאים על הקרח.
- הכן 24 צלחות אגר המכיל 25 מ"ל של SH (pH 7) בינוני עם 1.5% (w / v) אגר.
- לאחר אגר יש הקרושה, להפיץ 50 μl של 5, 50, ו 500 המ"מ CEPA פתרונות מניות על אגרת להשיג ריכוזי CEPA סופיים של 0.01, 0.1 ו -1.0 מ"מ, בהתאמה. הגדרת לוחות קבוצת ביקורת ממסים, אשר מבוססים על לא תיקון והפצת 50 μl של הממס המשמשים לפזר את תרכובות הבדיקה.
- לוחות Streak עבור מושבות מבודדות עם לולאה-מלא (~ 5 μl) של ק תרבות המתנע xylinus ולאטום אותם עם סרט פרפין. לחסן את כל הצלחות בשלושה עותקים. דגירת צלחות במשך חמישה ימים ב 30 מעלות צלזיוס.
- אוֹרִיָהמושבות ograph בהגדלה 20X באמצעות מיקרוסקופ USB דיגיטלי איכותי להעריך ייצור מורפולוגיה תאי קולוניאלי על בינוני מוצק.
הערה: תאית מופיע כחומר מעורפל לאורך השוליים של המושבה. - כדי לשלוט על רמות כלור פוספט, עורכים ניסוי זהה באמצעות 0.01, 0.1, ו -1.0 מ"מ של הפתרון NaCl-לאא 2 PO 4 באמצעות 5, 50, ו 500 מניות מ"מ, בהתאמה.
5. מבחנים קרומיים
- לגדול ולכמת ק תרבויות Starter xylinus בשלושה עותקים:
- בשלושה עותקים, לחסן מושבה אחת של ק xylinus לתוך 5 מ"ל של מדיום SH (pH 5) בתוספת 0.2% (v / v) cellulase מעוקר-מסנן. דגירה תרבויות על 30 מעלות צלזיוס עם תסיסה ב 150 סל"ד עד OD 600 של 0.3 עד 0.4 הוא הגיע (כ -72 שעות).
- תרבויות המתנע קציר על ידי צנטריפוגה (2,000 XG; 4 מעלות צלזיוס; 10 דקות). עם 5 מ"ל של תמיסת מלח סטרילית, לשטוף tהוא התאים פעמיים ו resuspend התא גלולה. שמור את התאים על הקרח.
- לכמת תאים באמצעות תא ספירת Petroff-אוסר.
- כן מאסטר תערובות כדי לחסן צלחות 24 גם:
- מוסף 60 מ"ל של מדיום SH (pH 7) עם 120 μl של 5, 50, ו 500 מניות מ"מ CEPA להשיג ריכוזים CEPA הסופי של 0.01, 0.1 ו -1.0 מ"מ, בהתאמה. מוסף עוד 60 מ"ל של מדיום SH (pH 7) עם 120 μl של הממס משמש כדי לפזר CEPA. וורטקס לערבב.
- מחלקים כל 60 מ"ל של מדיום המכיל CEPA לארבע aliquots 14 מ"ל. לחסן שלושה aliquots 14 מ"ל עם משכפל ביולוגי של תרבות המתנע בריכוז של 10 5 תאים / מ"ל. שמור צינורות עם תאים על קרח כדי למנוע ייצור תאי.
הערה: aliquot 14 המ"ל הנותר ישמש עבור בארות שליטת סטרילי.
- לחסן צלחות 24 גם:
- בדוק כל טיפול בצלחת משלה עם שלוש שורות של משכפל ביולוגישורה של בקרות סטרילית (איור 2).

איור 2:. זרימה תרשים הממחיש את פרוטוקול המשמש assay קרומית וניתוח Stock בינוני SH pH 7 CEPA-השלימו (60 מ"ל) הוא aliquoted במשך שלושה חיסונים לשכפל ביולוגי נפרד שליטה סטרילי (14 מ"ל כל אחד). תרבויות אלו מכן aliquoted לשש משכפל טכנית (2 מ"ל) לתוך צלחת 24 היטב ואחר כך חתמו בזריקת הסרט פרפין. לאחר דגירה למשך 7 ימים ב 30 מעלות צלזיוס, pellicles נקצר ומאופיין על ידי קביעת משקל רטוב, עובי, משקל יבש, ואת crystallinity ידי FT-IR. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו
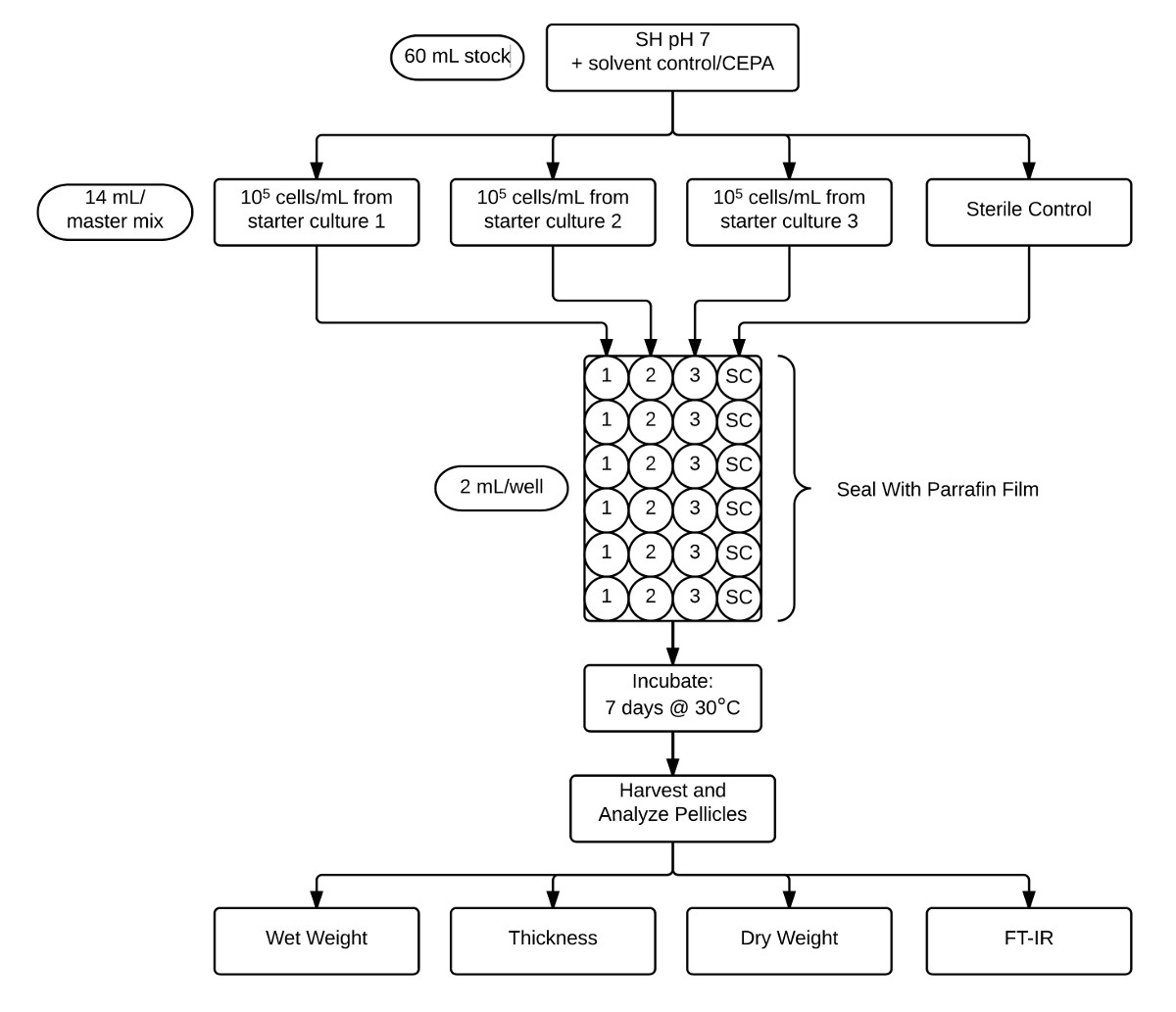{kind=link}
- תוך שימוש בתמהיל מאסטר 14 מ"ל, להוסיף 2 מ"ל לתוך EACשעות של שש בארות צלחת 24 גם סטרילי. השלם לשלושה הביולוגים משכפל ובקרה סטרילית (איור 2). חזור על הפעולה עבור כל טיפול.
- חותם צלחות עם סרט פרפין דגירה סטטי למשך 7 ימים ב 30 מעלות צלזיוס.
- קציר מדוד קרומי רטוב משקל, עובי, משקל יבש (תשואת צלולוזה) ו crystallinity (איור 2):
- לדכא צד אחד של קרומי לרומם את הקצה הקרומית היריבה ולהסיר pellicles פרט עם מלקחיים. תוך שמירה על אחיזה, ומניח אותם על מגבת נייר טריה במשך 3 שניות כדי להסיר בינוני עודף לפני השקילה כדי לקבוע משקולות הרטובות שלהם.
- יישר pellicles סומך סרגל ולצלם מהצד באמצעות מצלמה דיגיטלית ברזולוציה גבוהה.
- באמצעות ImageJ תוכנת 17, למדוד את העובי קרומי על הכתף השמאלית, כתף ימין והמרכז של כל קרומי. ממוצע כל המשכפל הטכני לכל replicat הביולוגידואר.
- בנפרד להעביר את pellicles לתוך הבארות של צלחת 6 באר. פנקו pellicles עם 12 מ"ל של 0.1 N NaOH על 80 מעלות צלזיוס למשך 20 דקות כדי lyse התאים.
- הסר את NaOH ולנטרל pellicles על ידי שטיפה עם מים טהורים למשך 24 שעות עם תסיסה. החלפת מים כל 6 שעות.
הערה: Pellicles צריך להיות לבן עם השלמת שלב הכביסה. - pellicles מקום על מחצלות סיליקון ויבש על 50 מעלות צלזיוס למשך 48 שעות כדי משקל קבוע. לאחר יבש, להסיר מן המחצלות ולמדוד משקולות קרומיות בקנה מידה אנליטית לקבוע תשואה תאית חיידקים.
- לנתח crystallinity קרומית באמצעות פורייה-transform ספקטרוסקופיה אינפרא אדום (FT-IR) באמצעות 32 סריקות ברזולוציה של 4 ס"מ -1 בטווח של 4,000 עד 650 ס"מ -1. חישוב המדד crystallinity, CI (IR), באמצעות 1437 / A 895; היחס הספיג של "להקת הגבישים" ו- "להקה אמורפי" כפי שתואר לעיל 18.
- כדי לשלוט על רמות כלור פוספט, עורכים ניסוי זהה באמצעות 0.01, 0.1, ו -1.0 מ"מ של הפתרון NaCl-לאא 2 PO 4 באמצעות 5, 50, ו 500 מניות מ"מ, בהתאמה.
- לנתח קרומי נתונים:
- חישוב הידרציה קרומית על ידי קביעת ההבדל בין משקל הקרומי רטוב משקל יבש.
- ממוצע הערכים של כל המשכפל הטכני על מנת להשיג ערך יחיד עבור כל לשכפל ביולוגי לניתוח סטטיסטי. השוואת טיפולים באמצעות ANOVA חד כיווני עם מבחן ההשוואה הנפוץ של Tukey. ההבדלים הם משמעותיים אם p <0.05.
- לנרמל את הנתונים כמו האחוז שולט מטופל ואת עלילת האמצעי ביולוגי משכפל.
Access restricted. Please log in or start a trial to view this content.
תוצאות
התקנת צלחת סכמטי עבור אימות שחרור אתילן CEPA במדיום SH (pH 7) על ידי assay בתגובה המשולש מוצגת באיור 1 א - ג. זרם-תרשים המתאר את הפרוטוקול הקרומי מוצג באיור 2. Dark-מבוגר א שתילי thaliana להפגין את הפנוטיפ בתגובה המשולשת (קצר ורחב...
Access restricted. Please log in or start a trial to view this content.
Discussion
השיטות שתוארו כאן מתארות את בייצור באתרו של אתילן CEPA לחקר תגובת אתילן חיידקים באמצעות האורגניזם המודל, ק xylinus. שיטה זו מאוד שימושי כמו אתילן יכול להיות מיוצר על ידי השלמת כל בתווך מימי כי יש pH גדול מ 3.5 10,11 עם CEPA מבטל את הצורך גז אתילן טהור או ציוד מעבדה מ?...
Access restricted. Please log in or start a trial to view this content.
Disclosures
The authors have nothing to disclose.
Acknowledgements
The authors thank Dr. Dario Bonetta for providing Arabidopsis thaliana seeds and for technical assistance in regards to the triple response assay, as well as Simone Quaranta for help with FT-IR. This work was supported by a Natural Sciences and Engineering Research Council of Canada Discovery Grant (NSERC-DG) to JLS, an Ontario Graduate Scholarship (OGS) to RVA, and a Queen Elizabeth II Graduate Scholarship in Science and Technology (QEII-GSST) to AJV.
Access restricted. Please log in or start a trial to view this content.
Materials
| Name | Company | Catalog Number | Comments |
| 1-aminocyclopropane carboxylic acid (ACC) | Sigma | A3903 | Biosynthetic precursor of ethylene in plants |
| 4-sector Petri dish | Phoenix Biomedical | CA73370-022 | For testing triple response |
| Agar | BioShop | AGR001.1 | To solidify medium |
| Canon Rebel T1i DLSR camera | Canon | 3818B004 | For pictures of pellicles |
| Cellulase from Trichoderma reesei ATCC 26921 | Sigma | C2730 | Aqueous solution |
| Citric acid | BioShop | CIT002.500 | For SH medium |
| Commercial bleach | Life Brand | 57800861874 | Bleach for seed sterilization |
| Concentrated HCl | BioShop | HCL666.500 | Hydrochloric acid for pH adjustment |
| Digital USB microscope | Plugable | N/A | For pictures of colonies |
| Ethephon (≥96%; 2-chloroethylphosphonic acid) | Sigma | C0143 | Ethylene-releasing compound |
| Glucose | BioBasic | GB0219 | For SH medium |
| Komagataeibacter xylinus ATCC 53582 | ATCC | 53582 | Bacterial cellulose-producing alphaproteobacterium |
| Microcentrifuge tube | LifeGene | LMCT1.7B | 1.7 ml microcentrifuge tube |
| Murashige and Skoog (MS) basal medium | Sigma | M5519 | Arabidopsis thaliana growth medium |
| Na2HPO4·7H2O | BioShop | SPD579.500 | Sodium phosphate, dibasic heptahydrate for SH medium |
| NaCl | BioBasic | SOD001.1 | Sodium chloride for saline and control solution |
| NaH2PO4·H2O | BioShop | SPM306.500 | Sodium phosphate, monobasic monohydrate for control solution |
| NaOH | BioShop | SHY700.500 | Sodium hydroxide for pH adjustment |
| Paraffin film | Parafilm | PM996 | For sealing plates and flasks |
| Peptone (bacteriological) | BioShop | PEP403.1 | For SH medium |
| Petroff-Hausser counting chamber | Hausser scientific | 3900 | Bacterial cell counting chamber |
| Polyethersulfone sterilization filter 0.2 µm | VWR | 28145-501 | For sterilizing cellulase |
| Sucrose | BioShop | SUC600.1 | Sucrose for MS medium |
| Yeast extract | BioBasic | G0961 | For SH medium |
References
- Guzmán, P., Ecker, J. R. Exploiting the triple response of Arabidopsis to identify ethylene-related mutants. Plant Cell. 2 (6), 513-523 (1990).
- Bakshi, A., Shemansky, J. M., Chang, C., Binder, B. M. History of research on the plant hormone ethylene. J. Plant Growth Regul. 34 (4), 809-827 (2015).
- Schaller, G. E. Ethylene and the regulation of plant development. BMC Biol. 10 (1), (2012).
- Hua, J., Sakai, H., et al. EIN4 and ERS2 are members of the putative ethylene receptor gene family in Arabidopsis. Plant Cell. 10 (8), 1321-1332 (1998).
- Bleecker, A. B., Estelle, M. A., Somerville, C., Kende, H. Insensitivity to ethylene conferred by a dominant Mutation in Arabidopsis thaliana. Science. 241 (4869), 1086-1089 (1988).
- Hamilton, A. J., Bouzayen, M., Grierson, D. Identification of a tomato gene for the ethylene-forming enzyme by expression in yeast. Proc. Natl. Acad. Sci. 88 (16), 7434-7437 (1991).
- Kim, J. Assessment of ethylene removal with Pseudomonas strains. J. Hazard. Mater. 131 (3), 131-136 (2006).
- Kim, H. E., Shitashiro, M., Kuroda, A., Takiguchi, N., Kato, J. Ethylene chemotaxis in Pseudomonas aeruginosa and other Pseudomonas species. Microbes Environ. 22 (2), 186-189 (2007).
- Augimeri, R. V., Strap, J. L. The phytohormone ethylene enhances bacterial cellulose production, regulates CRP/FNRKx transcription and causes differential gene expression within the cellulose synthesis operon of Komagataeibacter (Gluconacetobacter) xylinus ATCC 53582. Front. Microbiol. 6, 1459(2015).
- Zhang, W., Wen, C. K. Preparation of ethylene gas and comparison of ethylene responses induced by ethylene, ACC, and ethephon. Plant Physiol. Biochem. 48 (1), 45-53 (2010).
- Zhang, W., Hu, W., Wen, C. K. Ethylene preparation and its application to physiological experiments. Plant Signal. Behav. 5 (4), 453-457 (2010).
- Warner, H. L., Leopold, A. C. Ethylene evolution from 2-chloroethylphosphonic acid. Plant Physiol. 44 (1), 156-158 (1969).
- Biddle, E., Kerfoot, D. G. S., Kho, Y. H., Russell, K. E. Kinetic studies of the thermal decomposition of 2-chloroethylphosphonic acid in aqueous solution. Plant Physiol. 58 (5), 700-702 (1976).
- Klein, I., Lavee, S., Ben-Tal, Y. Effect of water vapor pressure on the thermal decomposition of 2-chloroethylphosphonic acid. Plant Physiol. 63 (3), 474-477 (1979).
- Murashige, T., Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 15 (3), 473-497 (1962).
- Schramm, M., Hestrin, S. Factors affecting production of cellulose at the air/liquid interface of a culture of Acetobacter xylinum. J. Gen. Microbiol. 11 (1), 123-129 (1954).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9 (7), 671-675 (2012).
- Ciolacu, D., Ciolacu, F., Popa, V. I. Amorphous cellulose-structure and characterization. Cellul. Chem. Technol. 45 (1), 13-21 (2011).
Access restricted. Please log in or start a trial to view this content.
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved