
このコンテンツを視聴するには、JoVE 購読が必要です。 サインイン又は無料トライアルを申し込む。
Method Article
細菌でエチレン応答を研究するためのツールとして、エチレン放出化合物、2-クロロエチルホスホン酸を利用します
要約
The protocols outlined herein facilitate the convenient investigation of bacterial ethylene responses by utilizing 2-chloroethylphosphonic acid (CEPA). Ethylene is produced in situ through the decomposition of CEPA in an aqueous bacterial growth medium, circumventing the requirement for pure ethylene gas.
要約
Ethylene (C2H4) is a gaseous phytohormone that is involved in numerous aspects of plant development, playing a dominant role in senescence and fruit ripening. Exogenous ethylene applied during early plant development triggers the triple response phenotype; a shorter and thicker hypocotyl with an exaggerated apical hook. Despite the intimate relationship between plants and bacteria, the effect of exogenous ethylene on bacteria has been greatly overlooked. This is partly due to the difficulty of controlling gaseous ethylene within the laboratory without specialized equipment. 2-Chloroethylphosphonic acid (CEPA) is a compound that decomposes into ethylene, chlorine, and phosphate in a 1:1:1:1 molar ratio when dissolved in an aqueous medium of pH 3.5 or greater. Here we describe the use of CEPA to produce in situ ethylene for the investigation of ethylene response in bacteria using the fruit-associated, cellulose-producing bacterium Komagataeibacter xylinus as a model organism. The protocols described herein include both the verification of ethylene production from CEPA via the Arabidopsis thaliana triple response assay and the effects of exogenous ethylene on K. xylinus cellulose production, pellicle properties and colonial morphology. These protocols can be adapted to examine the effect of ethylene on other microbes using appropriate growth media and phenotype analyses. The use of CEPA provides researchers with a simple and efficient alternative to pure ethylene gas for the routine determination of bacterial ethylene response.
概要
オレフィン、エチレン(C 2 H 4)それは、石炭ガスランプを使用し、実験室で増殖させたエンドウ苗は、(胚軸)をステムする異常な形態を示したことが観察された場合、最初1901年に植物ホルモンとして発見されたが、より短い厚かったですそして、通常のエンドウの苗に比べて横に曲がりました。表現型は、後に三重反応1,2と呼ばれます。その後の研究は、エチレンが、そのような成長、ストレス応答、果実の成熟と老化3。 シロイヌナズナ 、植物生物学研究のためのモデル生物として多数の発生過程を調節する重要な植物ホルモンは、十分にエチレンへの対応に関して検討されているであることを実証しました。いくつかのエチレン応答変異体は暗成長A.で観察されたトリプル応答表現型を利用することによって単離されていますエチレン1,4,5の存在下でのシロイヌナズナの苗。植物におけるエチレン生産のための生合成前駆体は、1-Aでありますminocyclopropaneカルボン酸(ACC)6、一般三重応答表現型1,4,5を導く内因性エチレン産生を増加させるためにトリプル応答アッセイ中に使用されます。
エチレン応答は、広く植物で研究されていますが、細菌上の外因性エチレンの影響は非常に植物と細菌の密接な関係にもかかわらず、understudiedされます。ある研究では、特定のシュードモナス株は炭素とエネルギー7の唯一の供給源としてエチレンを使用して生き残ることができることを報告しました。しかし、2つだけの研究では、細菌は、エチレンに応答することを実証しました。最初の研究は、 緑膿菌 、Pの株フルオレッセンス 、P.プチダ 、およびP. syringaeの溶融アガロース純エチレンガス8で平衡化し、走化性緩衝液と混合したアガロースプラグアッセイを使用してエチレンに向かって走化性ました。しかし、我々の知る限り、何のフュルトはなされていません特殊な装置なしに実験室でガスを扱う困難に可能性の高い細菌のエチレン応答を、特徴付けるために純粋なエチレンガスを使用して、えーレポート。細菌のエチレン応答の第二の報告書は、エチレンが9 xylinusフルーツ関連細菌、Komagataeibacter(旧グルコン )にバクテリアセルロースの生産と影響を受けた遺伝子の発現を増加させたことを実証しました。この場合には、エチレン放出化合物は、2-クロロエチルホスホン酸(CEPA)は、純粋なエチレンガスまたは特殊な装置の必要性を迂回し、細菌増殖培地内でその場でエチレンを製造するために使用しました。
14 -塩基触媒、一次反応12を介してpHが3.5 10,11上記モル比1:CEPAは1でエチレンを生成します。 CEPAの劣化を確実にpHおよび温度13,14およびエチルの製造における結果と相関していますエン、塩化物およびリン酸。 CEPAは、ガス状のエチレンへの便利な代替手段でエチレンに対する細菌の応答を研究に興味を持って研究者に提供します。
次のプロトコルの全体的な目標は、CEPA分解を確実にするために、細菌のエチレン応答を研究するための簡単かつ効率的な方法を提供し、細菌増殖培地中CEPA分解からエチレン生産の生理的に適切なレベルの検証を含む、培養pHを分析する時に損なわれません細菌の増殖、および細菌の形態および表現型に対するエチレンの影響の評価。我々は、Kを使用して、これらのプロトコルを示しますxylinusは、しかしながら、これらのプロトコルは、適切な増殖培地を使用し、表現型解析することによって、他の細菌におけるエチレン応答を研究するために適合させることができます。
Access restricted. Please log in or start a trial to view this content.
プロトコル
1.化学物質
- 500mMのCEPA(144.49グラム/モル)、及び500mMのNaCl(58.44グラム/モル)と500 mMののNaH 2 PO 4・H 2 O(137.99グラム/モル)酸性化(pHの両方からなる溶液の溶液を調製します2.5)超純水又は0.1NのHClです。解決策が明確になるまでボルテックスを用いて混合します。
- 直列5 mMのと50mMのストックを得るために、同じ溶媒中で(10倍)の500mM溶液を希釈します。
- 超純水に、1 - アミノシクロプロパンカルボン酸(101.1グラム/モルACC)の10mM溶液を準備します。
- -20℃でストック溶液とストアのアリコートをフィルター滅菌します。
- フィルター滅菌セルラーゼを。 4°Cで保存のアリコート。
2 - クロロエチルホスホン酸分解2.検証エチレン生成:トリプル応答アッセイ
- 気相法を用いたシロイヌナズナ生態型コロンビアの種子を表面殺菌:
注意:次のステップはトンを生産有酸素塩素ガス。ドラフト内で種子消毒を実施します。- 種子消毒用の250ミリリットルのガラスビーカーに対応し、ヒュームフードに置くのに十分な深密閉可能な容器を得ます。
- A.を追加シロイヌナズナの種子マイクロチューブに、ラックにチューブを配置します。密閉可能な容器に開いたチューブを含むラックを配置します。
注:塩素ガスが低く、種子に浸透することを可能にするのに半分を完全に個々のチューブを埋めないでください。 - シール可能な容器の中で、市販の漂白剤の100ミリリットルを含む250ミリリットルのビーカーを置きます。慎重に漂白剤に濃HClの3ミリリットルを追加し、すぐにその蓋で容器を密閉。
注意:漂白剤およびHClを表面滅菌種子に作用する毒性塩素ガスを生成するように反応します。 - 換気フードで4時間塩素ガスの存在下で種子をインキュベートします。
注:長い殺菌時間が発芽効率が低下します。 - 滅菌後、Cを許容しますhlorineガスは、次いで、微小遠心管を封止し、少なくとも1時間、ドラフト内で発散します。廃棄する前に、少なくとも24時間換気フードに漂白剤とHClの混合物を残します。
注:滅菌種子をすぐに使用するために室温で保存してもよいです。 - 長期保存のために4℃で種子を保管してください。結露を防止するために、マイクロチューブを開ける前に室温に種子を持参してください。暗闇の中の種子を保管してください。
- 4セクターのペトリ皿(90×15ミリメートル)に寒天プレートを準備します。
- A.の成長培地110 mlで調製シロイヌナズナの苗:1×ムラシゲ・スクーグ(MS)基本培地15(4.33グラム/ L)(w / v)のスクロース、0.8%(w / v)の寒天1%を含有します。 NaOHでpHを6にMS培地を調整します。
- CEPA分解のための細菌増殖培地の100ミリリットルを準備します。シュラムとHestrin(SH)媒体16は、1.5%を含有する寒天(w / v)の。 NaOHでpHを7にSH培地を調整します。
- オートクレーブ処理とテンペによってメディアを殺菌55℃の水浴中、R。
- MS寒天を鍛えた後、滅菌フラスコに40ミリリットルを追加します。 A.の陽性対照増殖培地を調製するためにシロイヌナズナの苗は、10μMのACCの最終濃度を得るために、10 mMのACCの40μlのMS寒天40mlのアリコートを補います。
- セクター化ペトリ皿( 図1)の適切な象限に培地の5ミリリットルを加え、寒天を固化することができます。三連ですべてのプレートを準備します。
注:CEPAは、後のプロトコルでまで増殖培地に添加されていません。

図1:CEPA トリプル応答アッセイのために使用される寒天プレートのセットアップ概略図(陰性対照(A)、陽性対照(B)のための特定の象限、実験プレートを示していますC)。この図は、Augimeriとストラップ9から変更されている。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
- 階層化A.同期発芽を確認するためのシロイヌナズナの種子:
- 約50 A.を追加MSまたはMS + ACC寒天を含む各象限へのシロイヌナズナの種子。確保種子を均一に苗除去および分析を容易にするために配布されます。
- 4で暗所で種を含むプレートをインキュベート 3-4日間Cを°。
- 細菌の増殖培地上のCEPA(SH)を分解し、トリプル応答アッセイを実行します。
- 層化した後、2時間蛍光灯に種を公開します。
- 最終CEPA濃度を得るために、SH寒天( 図1のpH 7)を含む実験的なプレートの象限に500 mMのCEPAストック溶液10μlを広めます1 mMになります。
- 実験用フィルムでプレートをシールし、種子のための暗い環境を作成するためにホイルでカバーしています。
- 寒天側を下にして3日間23℃の暗所でプレートをインキュベートすることにより、種子を発芽。
プレートを積層することができるが、エチレンは空気より軽いので、陰性対照は、底部に配置する必要があります注意してください。
- トリプル応答アッセイデータの分析:
- 火炎滅菌鉗子で、解剖やデジタルUSB顕微鏡下での各治療やビューに対応するプレートから単一の苗を削除します。苗の底整列さをし、写真を確認してください。
- 各象限(生物学的複製あたり60苗と処理当たり180本の苗木)から30本の苗木を削除します。定規と黒の背景と写真で表面に合わせます。
- ImageJソフトウェア17を使用して複製苗の胚軸長(mm)を測定します。 * *ストレートをクリックしてドラッグすることでスケールを設定します10ミリメートルの長さを選択するためのツール。 「分析」タブの下に "設定スケール」を選択し、* *セグメント化されたツールを使用して10に既知の距離を設定し、クリック&ドラッグして距離を測定する胚軸とプレスMの長さを選択します。 Tukeyの多重比較検定で一方向ANOVAを使用して、生物学的複製の平均を比較。 p <0.05 の場合には差は有意であると考えています。
細菌増殖を通じてのpHの3解析
- 成長し、三連で定量化Komagataeibacter xylinusスターター文化:
- Kの単一コロニーを接種しますxylinus 0.2%(v / v)のろ過滅菌セルラーゼを補充したSH培地5ml液(pH 5)に変換します。 0.4から0.3のOD 600が (約72時間)に達するまで、150rpmで攪拌しながら30℃で培養をインキュベートします。
- 遠心分離により収穫スターター培養物(2,000×gで、4℃、10分)。無菌の0.85%NAC(w / v)の5mLでL溶液は、細胞を2回洗浄し、細胞ペレットを再懸濁します。細胞を氷上に保管してください。
- Petroff-Hausser計数チャンバーを用いて細胞を定量化します。
- pH値の分析のための培養を接種します:
- 箔の蓋付き12 500ミリリットルのフラスコに0.2%(v / v)のフィルター滅菌セルラーゼを補っSHブロス(pHは7)の150ミリリットルを追加します。 K.とのフラスコに接種10 5細胞/ mlの濃度でxylinusスターター培養物。 3スターター培養物を使用して、処理当たり3つの生物学的複製物を準備します。
- それぞれ5300μlの、50、または500 mMのCEPA原液0.01の最終CEPA濃度を得るために、0.1、および1.0 mMの、と補足培養。 CEPAを溶解するために使用される溶剤の300μlの未処理対照培養物を補足。
- しっかりフラスコに箔の蓋をテーピングすることにより、フラスコを密封し、150rpmで攪拌しながら30℃で14日間培養をインキュベートします。
- 毎日、asepticaLLY(10分; 4℃で2000×gで)遠心分離によって各フラスコ、ペレット細胞から5ミリリットルのサンプルを削除します。きれいなチューブに上清を移し、pHメーターを使用して、各生物学的複製のpHを測定します。
- 生物学的複製の平均pH値をグラフによって経時的データを分析します。
注:培養pHが3.5以下に低下しないことが重要です。 5の培養pHは大幅にCEPAからのエチレン放出を減少させ、そして3.5以下のpHは完全にエチレン放出を阻害します。 - 、塩素及びリン酸のレベルをコントロール0.01、0.1を使用して同一の実験を行い、それぞれ、5、50、および500 mMのストックを使用してのNaCl-のNaH 2 PO 4溶液の1.0 mmまで。
4.コロニー形態
- K.を育てます三重でxylinusスターター培養物:
- Kの単一コロニーを接種しますxylinus 0.2%(v / v)のろ過滅菌セルラーゼを補充したSH培地5ml液(pH 5)に変換します。 Incuba0.4から0.3のOD 600まで150rpmで撹拌しながら30℃でのテ培養物を(約72時間)に達します。
- 遠心分離により収穫スターター培養物(2,000×gで、4℃、10分)。滅菌生理食塩水5mlで、細胞を洗浄した後、細胞ペレットを再懸濁します。細胞を氷上に保管してください。
- SHの25ミリリットル、1.5%(w / v)の寒天で(pHが7)培地を含む24寒天プレートを準備します。
- 寒天が固化した後は、それぞれ、0.01、0.1、および1.0 mMの最終的なCEPA濃度を得るために、5、50、50μlの、および寒天上で500mMのCEPAの原液を広げます。無修正で構成され、溶媒を50μlの広がりを設定し、未処理、溶媒対照プレートは、試験化合物を溶解するために使用されます。
- Kのループフル(〜5μl)を有する単離コロニーのためのストリークプレートスターターカルチャーxylinus、パラフィンフィルムでそれらを封印。三連ですべてのプレートを接種します。 30℃で5日間培養します。
- Phot定性的デジタルUSB顕微鏡を使用し、20X倍率でographコロニーは、固体培地上でコロニーの形態とセルロースの生産を評価します。
注:セルロースは、コロニーのマージンに沿って濁った物質として表示されます。 - 、塩素及びリン酸のレベルをコントロール0.01、0.1を使用して同一の実験を行い、それぞれ、5、50、および500 mMのストックを使用してのNaCl-のNaH 2 PO 4溶液の1.0 mmまで。
5.ペリクルアッセイ
- 成長し、定量化K.三重でxylinusスターター培養物:
- 三連では、Kの単一コロニーを接種xylinus 0.2%(v / v)のろ過滅菌セルラーゼを補充したSH培地5ml液(pH 5)に変換します。 0.4から0.3のOD 600が (約72時間)に達するまで、150rpmで攪拌しながら30℃で培養をインキュベートします。
- 遠心分離により収穫スターター培養物(2,000×gで、4℃、10分)。滅菌生理食塩水5mlで、Tを洗います彼細胞を2回、細胞ペレットを再懸濁します。細胞を氷上に保管してください。
- Petroff-Hausser計数チャンバーを用いて細胞を定量化します。
- 24ウェルプレートに接種し、マスターミックスを準備します。
- それぞれ、0.01の最終CEPA濃度を得るために、5の120μlを、50、および500 mMのCEPAの株式とSH培地(pHは7)の60ミリリットルを補足0.1、および1.0 mMの。 CEPAを溶解するために使用される溶剤の120μlのSH培地(pHは7)の別の60ミリリットルを補足。ミックスする渦。
- 4 14ミリリットルのアリコートにCEPA含有培地の各60ミリリットルを分割します。 10 5細胞/ mlの濃度で、スターター培養物の生物学的反復、14 mlのアリコートのうち3つを接種します。セルロースの生産を防止するために、氷上で細胞とチューブを保管してください。
注:残りの14ミリリットルのアリコートを無菌の対照ウェルのために使用されます。
- 24ウェルプレートに接種します。
- 生物学的反復の3行を持つ独自のプレートに、各処理をテストし、無菌コントロールの行( 図2)。

図2:ペリクルアッセイおよび分析の株式CEPA補充pHが7 SH培地(60ミリリットル) に使用されるプロトコルを示すフローチャートは、3つの別々の生物学的複製接種および滅菌コントロール(14ミリリットルずつ)のために分注します。これらの培養物を24ウェルプレートに6テクニカルレプリケート(2ml)中に分注し、次いでパラフィンフィルムで封止されています。 30℃で7日間インキュベートした後、ペリクルを回収し、FT-IRにより湿重量、厚み、乾燥重量、および結晶化度を決定することを特徴とする。 この図の拡大版をご覧になるにはこちらをクリックしてください。
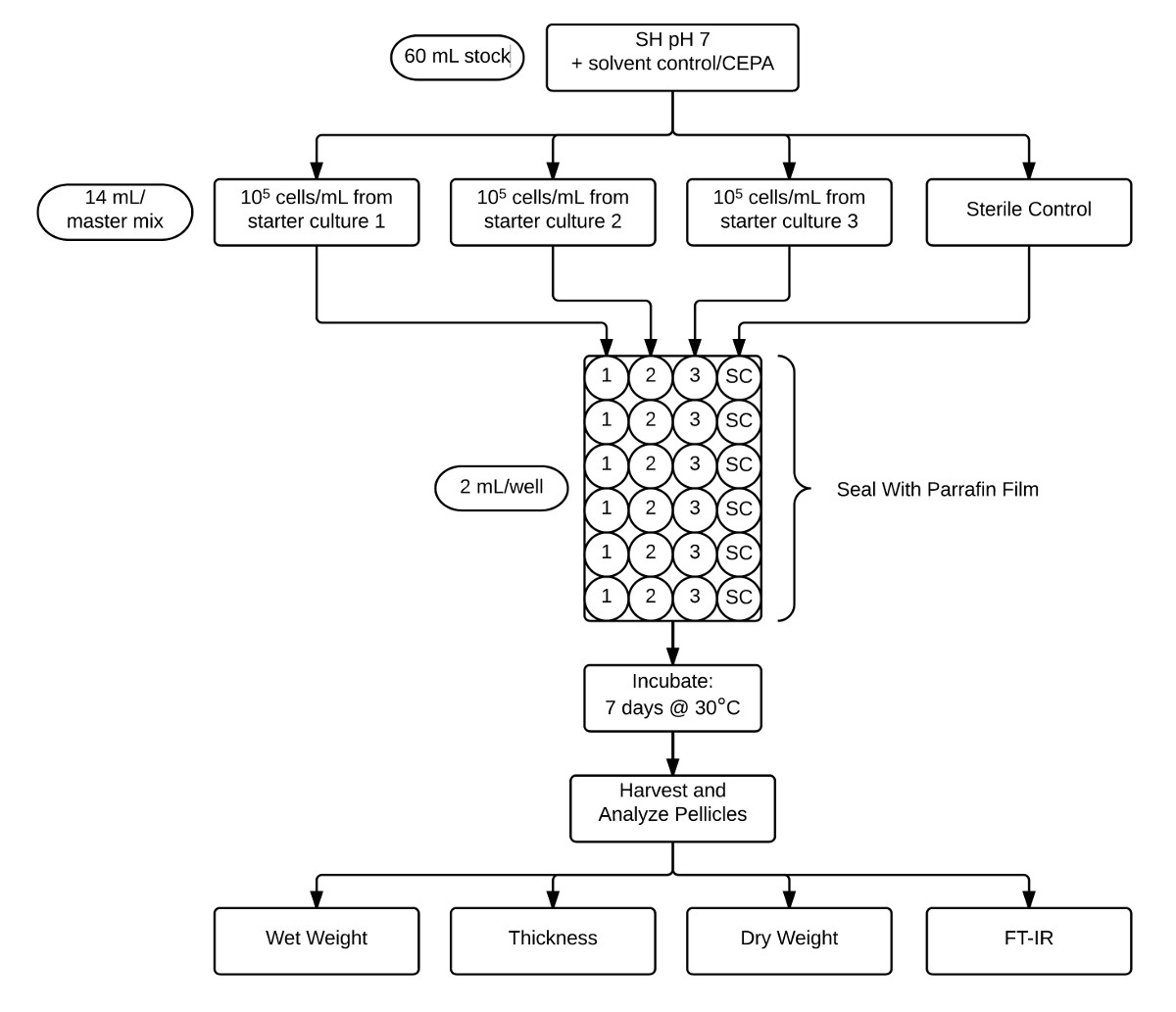{kind=link}
- 14ミリリットルマスターミックスを使用して、EACに2ミリリットルを追加します。滅菌した24ウェルプレートの6ウェルのH。 3つの生物学的複製物および滅菌コントロール( 図2)のために完了します。各治療のために繰り返します。
- パラフィンフィルムでシールプレートし、30℃で7日間、静的にインキュベートします。
- 収穫と測定ペリクル湿重量、厚さ、乾燥重量(セルロース収量)と結晶化度( 図2):
- 反対側のペリクルエッジを高め、鉗子で個々のペリクルを除去するために、ペリクルの片側を押し下げます。グリップを保持しつつ、その湿重量を決定するために計量前に、過剰な培地を除去するために、3秒間の新鮮なペーパータオル上に置きます。
- 高解像度デジタルカメラを使用して、サイドから定規と写真に隣接したペリクルを合わせます。
- ImageJソフトウェア17を使用して、左肩、右肩と各ペリクルの中央にペリクルの厚さを測定します。各生物学的replicatのためのすべての技術的反復の平均電子。
- 個別に6ウェルプレートのウェルにペリクルを移します。細胞を溶解するために20分間、80℃で0.1 N NaOHを12 mlのペリクルを扱います。
- NaOHを削除し、攪拌しながら24時間超純水で洗浄することによりペリクルを中和します。水ごとに6時間を変更します。
注:ペリクルは、洗浄工程の完了時に白でなければなりません。 - 一定重量になるまで48時間、50℃でシリコンマットとドライにペリクルを置きます。乾燥したら、マットから削除して、バクテリアセルロースの収率を決定するために分析的規模でペリクル重量を測定します。
- 4,000〜650センチメートルの範囲で赤外分光法(FT-IR)32スキャンを用い、分解能4cm -1フーリエ変換用いてペリクル結晶を分析-1。 1437 / A 895を使用して、結晶化度、CI(IR)を計算します。以前18に記載されているように 、「結晶性バンド」および「非晶質バンド」の吸光度比。
- 、塩素及びリン酸のレベルをコントロール0.01、0.1を使用して同一の実験を行い、それぞれ、5、50、および500 mMのストックを使用してのNaCl-のNaH 2 PO 4溶液の1.0 mmまで。
- ペリクルデータを分析します。
- ペリクル湿重量と乾燥重量との差を決定することによって、ペリクル水和を計算します。
- 統計分析のために、各生物学的複製のための単一の値を取得するためにすべての技術的反復の値を平均します。 Tukeyの多重比較検定で一方向ANOVAを用いて治療を比較してください。 p <0.05 の場合の違いは重要です。
- 未処理対照の割合としてデータを正規化し、生物学的複製の平均をプロットします。
Access restricted. Please log in or start a trial to view this content.
結果
C -トリプル応答アッセイによるSH培地(pHは7)におけるCEPAからのエチレン放出を検証するための模式的なプレートのセットアップを図1Aに示されています。ペリクルプロトコルを示すフロー・チャートは、 図2ダーク成長したAに示されていますシロイヌナズナの苗は、ACCの存在下およびSH培地(pHは7)上ではなく、?...
Access restricted. Please log in or start a trial to view this content.
ディスカッション
ここで説明する方法は、モデル生物、Kを用いて細菌エチレン応答の研究のためのCEPAからエチレンをその場生産に概説しますxylinus。エチレンは、CEPAは、純粋なエチレンガスや特殊な実験装置の必要性を否定すると3.5 10,11よりも高いpHを有する任意の水性媒体を補充することによって製造することができるように、この方法は非常に便利です。この方法は、?...
Access restricted. Please log in or start a trial to view this content.
開示事項
The authors have nothing to disclose.
謝辞
The authors thank Dr. Dario Bonetta for providing Arabidopsis thaliana seeds and for technical assistance in regards to the triple response assay, as well as Simone Quaranta for help with FT-IR. This work was supported by a Natural Sciences and Engineering Research Council of Canada Discovery Grant (NSERC-DG) to JLS, an Ontario Graduate Scholarship (OGS) to RVA, and a Queen Elizabeth II Graduate Scholarship in Science and Technology (QEII-GSST) to AJV.
Access restricted. Please log in or start a trial to view this content.
資料
| Name | Company | Catalog Number | Comments |
| 1-aminocyclopropane carboxylic acid (ACC) | Sigma | A3903 | Biosynthetic precursor of ethylene in plants |
| 4-sector Petri dish | Phoenix Biomedical | CA73370-022 | For testing triple response |
| Agar | BioShop | AGR001.1 | To solidify medium |
| Canon Rebel T1i DLSR camera | Canon | 3818B004 | For pictures of pellicles |
| Cellulase from Trichoderma reesei ATCC 26921 | Sigma | C2730 | Aqueous solution |
| Citric acid | BioShop | CIT002.500 | For SH medium |
| Commercial bleach | Life Brand | 57800861874 | Bleach for seed sterilization |
| Concentrated HCl | BioShop | HCL666.500 | Hydrochloric acid for pH adjustment |
| Digital USB microscope | Plugable | N/A | For pictures of colonies |
| Ethephon (≥96%; 2-chloroethylphosphonic acid) | Sigma | C0143 | Ethylene-releasing compound |
| Glucose | BioBasic | GB0219 | For SH medium |
| Komagataeibacter xylinus ATCC 53582 | ATCC | 53582 | Bacterial cellulose-producing alphaproteobacterium |
| Microcentrifuge tube | LifeGene | LMCT1.7B | 1.7 ml microcentrifuge tube |
| Murashige and Skoog (MS) basal medium | Sigma | M5519 | Arabidopsis thaliana growth medium |
| Na2HPO4·7H2O | BioShop | SPD579.500 | Sodium phosphate, dibasic heptahydrate for SH medium |
| NaCl | BioBasic | SOD001.1 | Sodium chloride for saline and control solution |
| NaH2PO4·H2O | BioShop | SPM306.500 | Sodium phosphate, monobasic monohydrate for control solution |
| NaOH | BioShop | SHY700.500 | Sodium hydroxide for pH adjustment |
| Paraffin film | Parafilm | PM996 | For sealing plates and flasks |
| Peptone (bacteriological) | BioShop | PEP403.1 | For SH medium |
| Petroff-Hausser counting chamber | Hausser scientific | 3900 | Bacterial cell counting chamber |
| Polyethersulfone sterilization filter 0.2 µm | VWR | 28145-501 | For sterilizing cellulase |
| Sucrose | BioShop | SUC600.1 | Sucrose for MS medium |
| Yeast extract | BioBasic | G0961 | For SH medium |
参考文献
- Guzmán, P., Ecker, J. R. Exploiting the triple response of Arabidopsis to identify ethylene-related mutants. Plant Cell. 2 (6), 513-523 (1990).
- Bakshi, A., Shemansky, J. M., Chang, C., Binder, B. M. History of research on the plant hormone ethylene. J. Plant Growth Regul. 34 (4), 809-827 (2015).
- Schaller, G. E. Ethylene and the regulation of plant development. BMC Biol. 10 (1), (2012).
- Hua, J., Sakai, H., et al. EIN4 and ERS2 are members of the putative ethylene receptor gene family in Arabidopsis. Plant Cell. 10 (8), 1321-1332 (1998).
- Bleecker, A. B., Estelle, M. A., Somerville, C., Kende, H. Insensitivity to ethylene conferred by a dominant Mutation in Arabidopsis thaliana. Science. 241 (4869), 1086-1089 (1988).
- Hamilton, A. J., Bouzayen, M., Grierson, D. Identification of a tomato gene for the ethylene-forming enzyme by expression in yeast. Proc. Natl. Acad. Sci. 88 (16), 7434-7437 (1991).
- Kim, J. Assessment of ethylene removal with Pseudomonas strains. J. Hazard. Mater. 131 (3), 131-136 (2006).
- Kim, H. E., Shitashiro, M., Kuroda, A., Takiguchi, N., Kato, J. Ethylene chemotaxis in Pseudomonas aeruginosa and other Pseudomonas species. Microbes Environ. 22 (2), 186-189 (2007).
- Augimeri, R. V., Strap, J. L. The phytohormone ethylene enhances bacterial cellulose production, regulates CRP/FNRKx transcription and causes differential gene expression within the cellulose synthesis operon of Komagataeibacter (Gluconacetobacter) xylinus ATCC 53582. Front. Microbiol. 6, 1459(2015).
- Zhang, W., Wen, C. K. Preparation of ethylene gas and comparison of ethylene responses induced by ethylene, ACC, and ethephon. Plant Physiol. Biochem. 48 (1), 45-53 (2010).
- Zhang, W., Hu, W., Wen, C. K. Ethylene preparation and its application to physiological experiments. Plant Signal. Behav. 5 (4), 453-457 (2010).
- Warner, H. L., Leopold, A. C. Ethylene evolution from 2-chloroethylphosphonic acid. Plant Physiol. 44 (1), 156-158 (1969).
- Biddle, E., Kerfoot, D. G. S., Kho, Y. H., Russell, K. E. Kinetic studies of the thermal decomposition of 2-chloroethylphosphonic acid in aqueous solution. Plant Physiol. 58 (5), 700-702 (1976).
- Klein, I., Lavee, S., Ben-Tal, Y. Effect of water vapor pressure on the thermal decomposition of 2-chloroethylphosphonic acid. Plant Physiol. 63 (3), 474-477 (1979).
- Murashige, T., Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 15 (3), 473-497 (1962).
- Schramm, M., Hestrin, S. Factors affecting production of cellulose at the air/liquid interface of a culture of Acetobacter xylinum. J. Gen. Microbiol. 11 (1), 123-129 (1954).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9 (7), 671-675 (2012).
- Ciolacu, D., Ciolacu, F., Popa, V. I. Amorphous cellulose-structure and characterization. Cellul. Chem. Technol. 45 (1), 13-21 (2011).
Access restricted. Please log in or start a trial to view this content.
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved