
需要订阅 JoVE 才能查看此. 登录或开始免费试用。
Method Article
利用乙烯释放化合物,2-氯乙基膦酸,作为一种工具来研究细菌乙烯反应
摘要
The protocols outlined herein facilitate the convenient investigation of bacterial ethylene responses by utilizing 2-chloroethylphosphonic acid (CEPA). Ethylene is produced in situ through the decomposition of CEPA in an aqueous bacterial growth medium, circumventing the requirement for pure ethylene gas.
摘要
Ethylene (C2H4) is a gaseous phytohormone that is involved in numerous aspects of plant development, playing a dominant role in senescence and fruit ripening. Exogenous ethylene applied during early plant development triggers the triple response phenotype; a shorter and thicker hypocotyl with an exaggerated apical hook. Despite the intimate relationship between plants and bacteria, the effect of exogenous ethylene on bacteria has been greatly overlooked. This is partly due to the difficulty of controlling gaseous ethylene within the laboratory without specialized equipment. 2-Chloroethylphosphonic acid (CEPA) is a compound that decomposes into ethylene, chlorine, and phosphate in a 1:1:1:1 molar ratio when dissolved in an aqueous medium of pH 3.5 or greater. Here we describe the use of CEPA to produce in situ ethylene for the investigation of ethylene response in bacteria using the fruit-associated, cellulose-producing bacterium Komagataeibacter xylinus as a model organism. The protocols described herein include both the verification of ethylene production from CEPA via the Arabidopsis thaliana triple response assay and the effects of exogenous ethylene on K. xylinus cellulose production, pellicle properties and colonial morphology. These protocols can be adapted to examine the effect of ethylene on other microbes using appropriate growth media and phenotype analyses. The use of CEPA provides researchers with a simple and efficient alternative to pure ethylene gas for the routine determination of bacterial ethylene response.
引言
烯烃乙烯(C 2 H 4)首次被发现在1901年的植物激素被观察到,豌豆苗,在使用煤气灯一个实验室中生长,显示出不正常的形态,其中茎(胚轴)为较短时,较厚的和弯曲相比普通豌豆苗横盘整理;表型后称为三重反应1,2。随后的研究表明,乙烯是一种调节多种发育过程如生长,应激反应,果实成熟衰老3。 拟南芥植物生物学研究模式生物的重要激素,已在关于其对乙烯反应得到很好的研究。几个乙烯反应突变体已经被分离通过利用在黑暗生长的答中观察到的三联反应表型拟南芥秧苗在乙烯1,4,5的存在。生物合成的前体为乙烯生产厂是1-一个minocyclopropane羧酸(ACC)6和三重反应测定期间通常用于增加内源性乙烯产量,导致三联响应表型1,4,5。
虽然乙烯反应被广泛研究在植物中,外源乙烯对细菌的效果大大充分研究,尽管细菌与植物的密切联系。一项研究报告指出某些假单胞菌株可以使用乙烯作为碳源和能源7的唯一来源生存。然而,只有两个研究已经证明,细菌乙烯反应。第一项研究表明, 铜绿假单胞菌 ,P的菌株荧光假单胞菌 ,P.恶臭 ,和P.丁香使用琼脂糖塞测定,其中熔融的琼脂糖用纯乙烯气8平衡的趋化性缓冲液混合分别朝向乙烯趋化。但是,据我们所知,没有出现过菲尔特采用纯乙烯气体呃报告表征细菌乙烯反应,可能是由于处理实验室气体没有专业设备的困难。细菌乙烯反应的第二份报告表明,乙烯增加细菌纤维素生产的影响基因表达的水果相关的细菌,Komagataeibacter(原Gluconacetobacter)xylinus 9。在这种情况下,乙烯-释放化合物,2-氯乙基膦酸(CEPA)用于细菌生长介质内就地产生乙烯,绕过用于纯乙烯气体或专门的设备的需要。
- 14通过一碱催化的,一级反应12 1的摩尔比高于pH 3.5 10,11:CEPA在1产生乙烯。 CEPA的降解在生产乙酯与pH和温度13,14和结果正相关烯,氯和磷酸盐。 CEPA提供兴趣研究细菌反应,乙烯与方便的替代乙烯气体的研究。
以下协议的总体目标是提供一种简单而有效的方法来研究细菌乙烯反应,并且包括从安排分解细菌生长培养基中生产乙烯的生理相关水平的验证,培养物pH的分析,以确保CEPA分解期间不受损细菌生长,和的乙烯对细菌形态和表型的影响的评估。我们演示使用K.这些协议xylinus,然而,这些协议可以适于通过使用适当的生长培养基中和表型分析,以研究在其它细菌乙烯反应。
Access restricted. Please log in or start a trial to view this content.
研究方案
1.化学品
- 制备500mM的安排(144.49克/摩尔)的溶液中,并同时包含500 mM氯化钠(58.44克/摩尔)中的溶液和500毫的NaH 2 PO 4·H 2 O(137.99克/摩尔)中酸化(pH 2.5)超纯水或0.1N HCl中。混合使用一个旋涡,直到解决方案是明确的。
- 连续稀释(10倍)的500mM的溶液在相同的溶剂,得到的5mM和50mM的股票。
- 制备的1-氨基环丙烷甲酸(ACC;101.1克/摩尔)的10mM溶液在超纯水。
- 过滤消毒在-20°C股票的解决方案和存储等分。
- 过滤消毒纤维素酶。商店等分在4℃。
从2-氯乙基酸分解2.验证乙烯生产:三重反应分析
- 表面消毒拟南芥使用气相法生态型哥伦比亚种子:
注意:以下步骤产生Ť好氧氯气。进行种子消毒在通风橱中。- 获得的可密封容器足够深以容纳种子灭菌的250ml玻璃烧杯中,并将其放置在通风橱中。
- A.添加拟南芥种子离心管,并将管在机架中。将装有开管到密封的容器架。
注意:不要填写个人管在半满,使氯气渗透到较低的种子。 - 放置装有在可密封容器100毫升市售漂白剂的250毫升烧杯中。小心加入3毫升浓盐酸于漂白,并立即封住其盖子的容器。
注意:漂白和HCl反应而产生,其作用进行表面消毒的种子有毒的氯气。 - 孵育在氯气存在下的种子在通风柜4小时。
注:较长的消毒时间会降低发芽效率。 - 灭菌后,以便Chlorine气体在通风柜泄至少1小时,然后密封该离心管中。丢弃之前离开的漂白剂和盐酸混合物,在通风柜至少24小时。
注意:灭菌的种子可在室温下保存立即使用。 - 保持种子在4℃下长期贮存。打开微量离心管,以防止冷凝之前带来种子至室温。商店的种子在黑暗中。
- 在4个部门的培养皿(90×15毫米)准备琼脂平板:
- 准备A.110毫升生长培养基拟南芥苗:1×Murashige和含有1%斯库格(MS)的基础培养基15(4.33克/升)(重量/体积)蔗糖和0.8%(重量/体积)的琼脂。调整用NaOH的MS培养基至pH 6。
- 制备100毫升CEPA分解细菌生长培养基:施拉姆和Hestrin含16 1.5%(SH)培养基(重量/体积)的琼脂。调整用NaOH的SH培养基至pH 7。
- 通过高压灭菌和消毒坦佩媒体R IN 55℃水浴。
- 一旦MS琼脂回火,加40毫升的无菌瓶中。为了准备A.阳性对照生长培养基拟南芥幼苗,补充40毫升一份的MS琼脂的40微升10毫ACC,得到10μM的ACC的终浓度。
- 5ml培养基添加到的扇形培养皿中( 图1)相应的象限,并允许在琼脂固化。准备好所有板块一式三份。
注意:CEPA不被添加到生长培养基中,直到在后面的协议。

图 1: 用于与CEPA的三重反应实验琼脂平板上的安装示意图说明具体为阴性对照组(A),阳性对照(B)的象限,和实验板(C)。这个数字已经从Augimeri和带9修改。 请点击此处查看本图的放大版本。
{kind=link}
- 分层A.拟南芥种子保证同步萌发:
- 加入大约50 A.拟南芥种子到包含MS或MS + ACC琼脂每个象限。确保种子均匀分布以促进苗去除和分析。
- 孵育含板种子在黑暗中在4 °下3-4天。
- 分解CEPA对细菌生长培养基(SH)和执行三重反应分析:
- 分层后,暴露的种子,以荧光灯2小时。
- 蔓延10微升的500mM CEPA原液在含SH琼脂试验板的象限(pH值为7; 图1),以获得最终CEPA浓度1毫米。
- 与密封实验薄膜板材和铝箔盖创造的种子黑暗的环境。
- 通过在23℃下与琼脂面朝下在黑暗中温育板3天发芽种子。
注意:板可堆叠,但阴性对照应放置在底部,因为乙烯是比空气轻。
- 分析三重反应分析数据:
- 用火焰灭菌的镊子,从对应于解剖或数字的USB显微镜下的每个处理和视图板除去单苗。确保苗的底部对齐,并且照片。
- 取下每个象限30苗(每生物重复60苗每处理180苗)。对齐表面上的黑色背景和照片用尺子。
- 测量使用ImageJ软件17重复苗的下胚轴长度(mm)。通过点击并拖动*直设置比例*工具选择为10毫米的长度。选择"设置刻度"的"分析"选项卡下的和已知的距离设置为10使用*分段*工具,单击并拖动来选择胚轴和按M键的长度测量距离。比较使用单向ANOVA和Tukey多重比较检验生物学重复的平均值。差异被认为是显著如果p <0.05。
整个细菌生长pH值为3分析
- 一式三份成长和量化Komagataeibacter xylinus发酵剂:
- 接种K的单菌落xylinus到5ml的SH培养基(pH为5),补充了0.2%(体积/体积)过滤灭菌纤维素酶。孵育在30℃下以150rpm搅拌培养物,直到OD达到0.3〜0.4 600(约72小时)。
- 通过离心收获发酵剂(2,000 xg离心; 4℃; 10分钟)。用5毫升的无菌0.85%(重量/体积)NACL溶液,洗涤细胞两次,重悬细胞沉淀。保持细胞在冰上。
- 量化使用佩特罗夫,豪塞尔计数室细胞。
- 接种培养pH值分析:
- 增加150个10ml补充有0.2%(V / V)过滤灭菌纤维素到十二500毫升烧瓶用铝箔盖的SH培养基(pH 7)的。与接种瓶K. xylinus起始培养以10 5个细胞/ ml的浓度。用三个发酵剂,每次准备治疗三次生物学重复。
- 补充培养物用300μl的5,50或500mM的安排储备溶液,以获得最终CEPA浓度的0.01,0.1和1.0毫米。补充用300μl用于溶解CEPA溶剂的未处理的对照培养物。
- 通过紧密胶带箔盖的烧瓶密封烧瓶中并在150rpm下搅拌,在30℃孵育培养物14天。
- 每一天,asepticaLLY通过离心除去从每个烧瓶中并沉淀格5 1ml样品(2,000 xg离心; 4℃; 10分钟)。转移上清液至一个干净的试管,并测量使用pH计各生物复制的pH。
- 通过作图的生物学重复的均值pH值分析时程数据。
注意:重要的是,培养物的pH不低于3.5; 5个培养物pH显著降低乙烯释放从安排,和pH值低于3.5完全抑制乙烯释放。 - 为了控制氯和磷酸盐水平,执行使用0.01,0.1相同的实验,并用5,50和500毫米的股票,分别的NaCl的NaH 2 PO 4溶液的1.0mM的。
4.菌落形态
- 成长K. xylinus发酵剂培养物一式三份:
- 接种K的单菌落xylinus到5ml的SH培养基(pH为5),补充了0.2%(体积/体积)过滤灭菌纤维素酶。 Incuba德培养在30℃下以150rpm搅拌直到0.3至0.4的OD 600达到(约72小时)。
- 通过离心收获发酵剂(2,000 xg离心; 4℃; 10分钟)。用5毫升的无菌盐水的,洗细胞,然后重悬细胞沉淀。保持细胞在冰上。
- 制备(pH 7)中介质24琼脂平板含有25毫升的SH为1.5%(重量/体积)的琼脂。
- 一旦琼脂凝固,散布50微升的5,50和500上的琼脂毫安排储备溶液分别获得0.01,0.1和1.0毫最终CEPA浓度。建立未处理和溶剂对照板,其中包括没有修改和50微升的溶剂的扩频用于溶解的测试化合物。
- 为K的一个循环满(〜5微升)分离的菌落的Streak平板xylinus启动文化和石蜡膜密封。接种所有板块一式三份。在30℃下孵育板五天。
- 辐透使用数字USB显微镜和质量在放大20倍ograph殖民地评估固体培养基上菌落形态和纤维素的生产。
注:纤维素表现为沿菌落边缘朦胧物质。 - 为了控制氯和磷酸盐水平,执行使用0.01,0.1相同的实验,并用5,50和500毫米的股票,分别的NaCl的NaH 2 PO 4溶液的1.0mM的。
5.薄皮测定
- 成长和量化K. xylinus发酵剂培养物一式三份:
- 一式三份,接种K的单菌落xylinus到5ml的SH培养基(pH为5),补充了0.2%(体积/体积)过滤灭菌纤维素酶。孵育在30℃下以150rpm搅拌培养物,直到OD达到0.3〜0.4 600(约72小时)。
- 通过离心收获发酵剂(2,000 xg离心; 4℃; 10分钟)。用5毫升无菌盐水,冲洗Ť他细胞两次,重悬细胞沉淀。保持细胞在冰上。
- 量化使用佩特罗夫,豪塞尔计数室细胞。
- 准备预混接种24孔板:
- 补充60毫升的SH培养基(pH为7)与120微升的5,50和500毫CEPA股以获得0.01,0.1和1.0毫最终CEPA浓度分别。补充另60毫升SH培养基(pH为7)与120微升用于溶解CEPA溶剂的。旋涡混合。
- 划分各60毫升含有CEPA培养基成4个14毫升分装。以10 5个细胞/ ml的浓度接种三连起子培养的生物学重复14毫升等分试样。保持与管细胞在冰上以防止纤维素的生产。
注:其余14毫升一份将用于无菌对照孔。
- 接种24孔板:
- 测试在其自己的板各处理与生物学重复的三排,并无菌控件的行( 图2)。

图2:示出了用于防护薄膜组件测定和分析库存CEPA补充的pH为7 SH培养基(60毫升) 的协议流程图等分为三个独立的生物复制接种和无菌控制(各14ml)中。然后,这些培养物等分至六个技术重复(2ml)中于24孔板,然后用石蜡膜密封。在30℃下培养7天后,菌膜收获和特点是由FT-IR确定湿重,厚度,干重,和结晶度。 请点击此处查看该图的放大版本
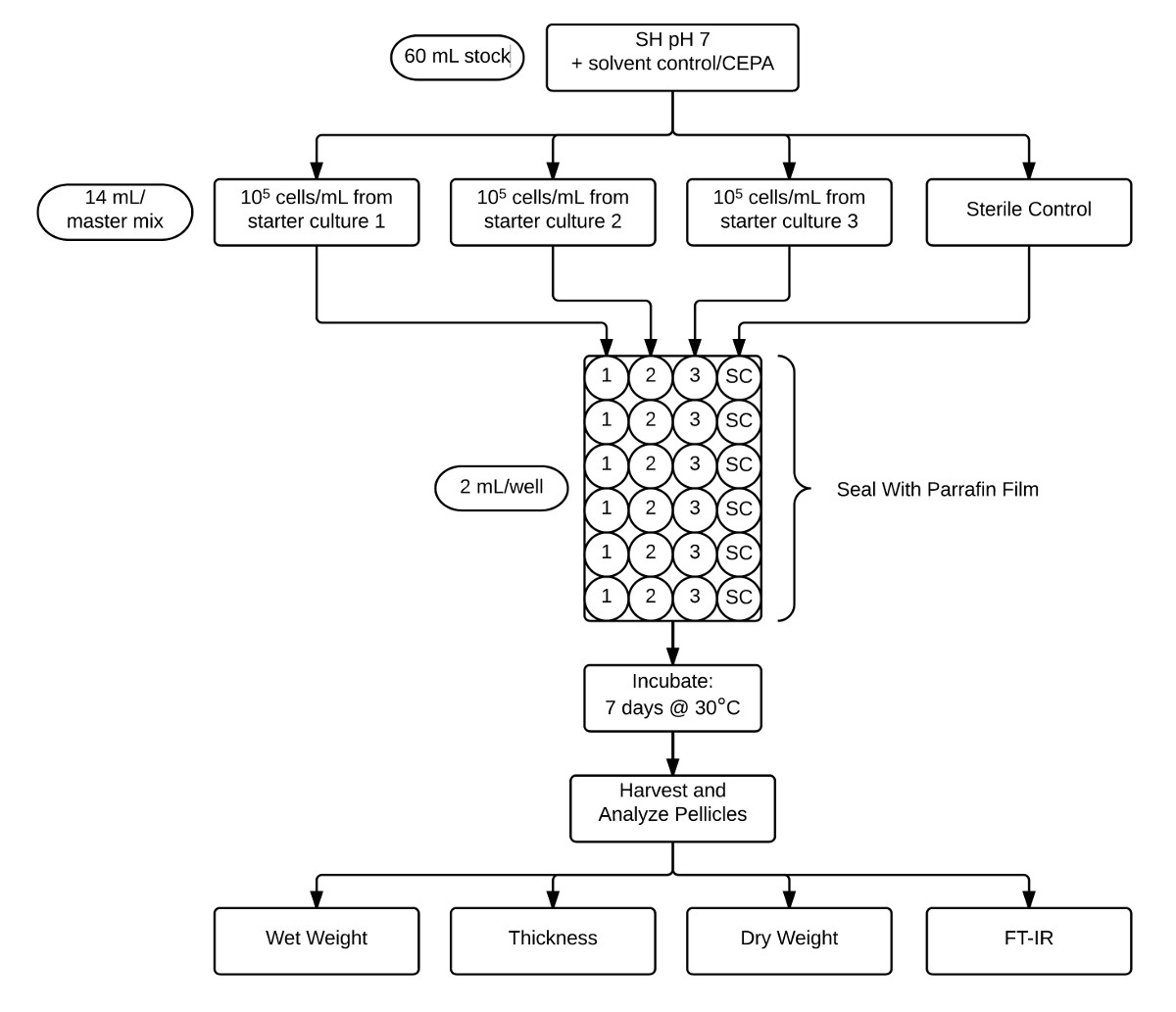{kind=link}
- 使用14毫升主结构,加2ml到EAC无菌24孔板六个孔小时。完成三个生物学重复和无菌控制( 图2)。重复每个处理。
- 用石蜡膜密封板,并在30℃下静态孵育7天。
- 采收和测量薄皮湿重,厚度,干重(纤维素产量)和结晶度( 图2):
- 压下薄膜的一侧提升对方薄膜边缘并移除个人药膜用钳子。在保持抓地力,把他们放在新鲜的纸巾在3秒内称重,以确定其湿重之前删除多余的网上平台。
- 相邻对齐使用高分辨率数码相机从侧面一把尺子和照片药膜。
- 使用ImageJ软件17,测量左肩,右肩各薄膜的薄膜中心厚度。平均所有技术复制每个生物replicat即
- 单独的菌膜转移到6孔板的孔中。在80℃下用12ml 0.1N NaOH中的治疗药膜20分钟以裂解细胞。
- 取出NaOH和用超纯水清洗与搅拌24小时中和药膜。换水每隔6小时。
注意:药膜应该是洗涤步骤完成后白色。 - 上硅垫地方药膜和在50℃下干燥48小时至恒重。干燥后,从垫取出,并测量薄膜上的权重分析的规模,以确定细菌纤维素产量。
- 使用傅立叶变换红外光谱使用32次扫描(FT-IR)和4cm的分辨率-1 4000至650厘米范围内分析薄膜结晶-1。计算结晶度指数,CI(IR),采用了1437 / A 895;在"晶体乐队"和"非晶带"的吸收比前面所描述的18。
- 为了控制氯和磷酸盐水平,执行使用0.01,0.1相同的实验,并用5,50和500毫米的股票,分别的NaCl的NaH 2 PO 4溶液的1.0mM的。
- 分析数据的Pellicle:
- 通过确定薄膜湿重和干重之差计算防护膜水合。
- 平均所有技术重复的值,以获得用于统计分析每个生物复制一个值。比较使用单向ANOVA和Tukey多重比较检验处理。差异显著如果p <0.05。
- 正常化的数据作为未处理的对照组的百分比和绘制生物学重复的平均值。
Access restricted. Please log in or start a trial to view this content.
结果
示意图板设置从CEPA乙烯解放SH培养基(pH值为7)经三重反应实验验证如图1A - ℃。示出了防护薄膜组件的协议的流程图在图2中黑暗生长的A中示出拟南芥幼苗展现的三重反应表型(以一种夸张的顶钩较短和较厚的下胚轴)中ACC的存在下,并在通过CEPA的SH培养基上分解产生乙烯(pH 7)中的存在,但未经处理的条件下,不
Access restricted. Please log in or start a trial to view this content.
讨论
此处所描述的方法使用的模式生物,K勾勒出在从CEPA 就地产生乙烯的细菌乙烯反应的研究xylinus。这种方法是非常有用的乙烯可以通过补充有pH值大于3.5 10,11-用CEPA否定为纯乙烯气或专门的实验室设备的需要的任何含水介质来制备。这种方法不限定于学习CEPA衍生乙烯对细菌的效果,但也可以适用于研究在真核生物乙烯反应。重要的是平行的对照实验,用磷酸盐和?...
Access restricted. Please log in or start a trial to view this content.
披露声明
The authors have nothing to disclose.
致谢
The authors thank Dr. Dario Bonetta for providing Arabidopsis thaliana seeds and for technical assistance in regards to the triple response assay, as well as Simone Quaranta for help with FT-IR. This work was supported by a Natural Sciences and Engineering Research Council of Canada Discovery Grant (NSERC-DG) to JLS, an Ontario Graduate Scholarship (OGS) to RVA, and a Queen Elizabeth II Graduate Scholarship in Science and Technology (QEII-GSST) to AJV.
Access restricted. Please log in or start a trial to view this content.
材料
| Name | Company | Catalog Number | Comments |
| 1-aminocyclopropane carboxylic acid (ACC) | Sigma | A3903 | Biosynthetic precursor of ethylene in plants |
| 4-sector Petri dish | Phoenix Biomedical | CA73370-022 | For testing triple response |
| Agar | BioShop | AGR001.1 | To solidify medium |
| Canon Rebel T1i DLSR camera | Canon | 3818B004 | For pictures of pellicles |
| Cellulase from Trichoderma reesei ATCC 26921 | Sigma | C2730 | Aqueous solution |
| Citric acid | BioShop | CIT002.500 | For SH medium |
| Commercial bleach | Life Brand | 57800861874 | Bleach for seed sterilization |
| Concentrated HCl | BioShop | HCL666.500 | Hydrochloric acid for pH adjustment |
| Digital USB microscope | Plugable | N/A | For pictures of colonies |
| Ethephon (≥96%; 2-chloroethylphosphonic acid) | Sigma | C0143 | Ethylene-releasing compound |
| Glucose | BioBasic | GB0219 | For SH medium |
| Komagataeibacter xylinus ATCC 53582 | ATCC | 53582 | Bacterial cellulose-producing alphaproteobacterium |
| Microcentrifuge tube | LifeGene | LMCT1.7B | 1.7 ml microcentrifuge tube |
| Murashige and Skoog (MS) basal medium | Sigma | M5519 | Arabidopsis thaliana growth medium |
| Na2HPO4·7H2O | BioShop | SPD579.500 | Sodium phosphate, dibasic heptahydrate for SH medium |
| NaCl | BioBasic | SOD001.1 | Sodium chloride for saline and control solution |
| NaH2PO4·H2O | BioShop | SPM306.500 | Sodium phosphate, monobasic monohydrate for control solution |
| NaOH | BioShop | SHY700.500 | Sodium hydroxide for pH adjustment |
| Paraffin film | Parafilm | PM996 | For sealing plates and flasks |
| Peptone (bacteriological) | BioShop | PEP403.1 | For SH medium |
| Petroff-Hausser counting chamber | Hausser scientific | 3900 | Bacterial cell counting chamber |
| Polyethersulfone sterilization filter 0.2 µm | VWR | 28145-501 | For sterilizing cellulase |
| Sucrose | BioShop | SUC600.1 | Sucrose for MS medium |
| Yeast extract | BioBasic | G0961 | For SH medium |
参考文献
- Guzmán, P., Ecker, J. R. Exploiting the triple response of Arabidopsis to identify ethylene-related mutants. Plant Cell. 2 (6), 513-523 (1990).
- Bakshi, A., Shemansky, J. M., Chang, C., Binder, B. M. History of research on the plant hormone ethylene. J. Plant Growth Regul. 34 (4), 809-827 (2015).
- Schaller, G. E. Ethylene and the regulation of plant development. BMC Biol. 10 (1), (2012).
- Hua, J., Sakai, H., et al. EIN4 and ERS2 are members of the putative ethylene receptor gene family in Arabidopsis. Plant Cell. 10 (8), 1321-1332 (1998).
- Bleecker, A. B., Estelle, M. A., Somerville, C., Kende, H. Insensitivity to ethylene conferred by a dominant Mutation in Arabidopsis thaliana. Science. 241 (4869), 1086-1089 (1988).
- Hamilton, A. J., Bouzayen, M., Grierson, D. Identification of a tomato gene for the ethylene-forming enzyme by expression in yeast. Proc. Natl. Acad. Sci. 88 (16), 7434-7437 (1991).
- Kim, J. Assessment of ethylene removal with Pseudomonas strains. J. Hazard. Mater. 131 (3), 131-136 (2006).
- Kim, H. E., Shitashiro, M., Kuroda, A., Takiguchi, N., Kato, J. Ethylene chemotaxis in Pseudomonas aeruginosa and other Pseudomonas species. Microbes Environ. 22 (2), 186-189 (2007).
- Augimeri, R. V., Strap, J. L. The phytohormone ethylene enhances bacterial cellulose production, regulates CRP/FNRKx transcription and causes differential gene expression within the cellulose synthesis operon of Komagataeibacter (Gluconacetobacter) xylinus ATCC 53582. Front. Microbiol. 6, 1459(2015).
- Zhang, W., Wen, C. K. Preparation of ethylene gas and comparison of ethylene responses induced by ethylene, ACC, and ethephon. Plant Physiol. Biochem. 48 (1), 45-53 (2010).
- Zhang, W., Hu, W., Wen, C. K. Ethylene preparation and its application to physiological experiments. Plant Signal. Behav. 5 (4), 453-457 (2010).
- Warner, H. L., Leopold, A. C. Ethylene evolution from 2-chloroethylphosphonic acid. Plant Physiol. 44 (1), 156-158 (1969).
- Biddle, E., Kerfoot, D. G. S., Kho, Y. H., Russell, K. E. Kinetic studies of the thermal decomposition of 2-chloroethylphosphonic acid in aqueous solution. Plant Physiol. 58 (5), 700-702 (1976).
- Klein, I., Lavee, S., Ben-Tal, Y. Effect of water vapor pressure on the thermal decomposition of 2-chloroethylphosphonic acid. Plant Physiol. 63 (3), 474-477 (1979).
- Murashige, T., Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 15 (3), 473-497 (1962).
- Schramm, M., Hestrin, S. Factors affecting production of cellulose at the air/liquid interface of a culture of Acetobacter xylinum. J. Gen. Microbiol. 11 (1), 123-129 (1954).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9 (7), 671-675 (2012).
- Ciolacu, D., Ciolacu, F., Popa, V. I. Amorphous cellulose-structure and characterization. Cellul. Chem. Technol. 45 (1), 13-21 (2011).
Access restricted. Please log in or start a trial to view this content.
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。