
É necessária uma assinatura da JoVE para visualizar este conteúdo. Faça login ou comece sua avaliação gratuita.
Method Article
Utilizando o composto libertador de etileno, ácido 2-cloroetilfosfônico, como uma ferramenta para estudar a resposta do etileno em bactérias
Neste Artigo
Resumo
The protocols outlined herein facilitate the convenient investigation of bacterial ethylene responses by utilizing 2-chloroethylphosphonic acid (CEPA). Ethylene is produced in situ through the decomposition of CEPA in an aqueous bacterial growth medium, circumventing the requirement for pure ethylene gas.
Resumo
Ethylene (C2H4) is a gaseous phytohormone that is involved in numerous aspects of plant development, playing a dominant role in senescence and fruit ripening. Exogenous ethylene applied during early plant development triggers the triple response phenotype; a shorter and thicker hypocotyl with an exaggerated apical hook. Despite the intimate relationship between plants and bacteria, the effect of exogenous ethylene on bacteria has been greatly overlooked. This is partly due to the difficulty of controlling gaseous ethylene within the laboratory without specialized equipment. 2-Chloroethylphosphonic acid (CEPA) is a compound that decomposes into ethylene, chlorine, and phosphate in a 1:1:1:1 molar ratio when dissolved in an aqueous medium of pH 3.5 or greater. Here we describe the use of CEPA to produce in situ ethylene for the investigation of ethylene response in bacteria using the fruit-associated, cellulose-producing bacterium Komagataeibacter xylinus as a model organism. The protocols described herein include both the verification of ethylene production from CEPA via the Arabidopsis thaliana triple response assay and the effects of exogenous ethylene on K. xylinus cellulose production, pellicle properties and colonial morphology. These protocols can be adapted to examine the effect of ethylene on other microbes using appropriate growth media and phenotype analyses. The use of CEPA provides researchers with a simple and efficient alternative to pure ethylene gas for the routine determination of bacterial ethylene response.
Introdução
O etileno olefina (C 2 H 4) foi descoberto pela primeira vez como uma hormona vegetal, em 1901, quando foi observado que as mudas de ervilha, cultivadas em laboratório que utilizadas lâmpadas de gás de carvão, exibiu uma morfologia anormal em que hastes (hipocótilos) eram mais curtos, mais espessa e inclinou-se para os lados em relação às plantas de ervilha normais; um fenótipo mais tarde denominado a 1,2 resposta tripla. Estudos posteriores demonstraram que o etileno é um hormônio vegetal vital que regula vários processos de desenvolvimento como o crescimento, a resposta ao estresse, o amadurecimento dos frutos e senescência 3. Arabidopsis thaliana, um organismo modelo para a investigação biologia vegetal, tem sido bem estudada em relação à sua resposta ao etileno. Mutantes de resposta Vários etileno foram isolados, explorando o fenótipo triplo resposta observada em dark-grown A. mudas thaliana na presença de etileno 1,4,5. O precursor biossintético para a produção de etileno em plantas é 1-Aácido minocyclopropane carboxílico (ACC) e 6 é comumente utilizado durante o ensaio de resposta tripla para aumentar a produção endógena de etileno que conduz ao fenótipo triplo 1,4,5 resposta.
Embora a resposta ao etileno é amplamente estudada em plantas, o efeito do etileno exógeno em bactérias é muito pouco estudada, apesar da estreita associação de bactérias com plantas. Um estudo relatou que certas estirpes de Pseudomonas pode sobreviver usando etileno como única fonte de carbono e energia 7. No entanto, apenas dois estudos demonstraram que as bactérias responder ao etileno. O primeiro estudo mostrou que as estirpes de Pseudomonas aeruginosa, P. fluorescens, P. putida, e P. syringae foram quimiotáctica para etileno utilizando um ensaio de tampão de agarose no qual agarose fundida foi misturada com um tampão de quimiotaxia equilibrada com etileno puro gás 8. No entanto, a nosso conhecimento, não houve nenhum further relatórios que utilizam gás de etileno puro para caracterizar a resposta de etileno bacteriana, provavelmente devido à dificuldade de manipulação de gases no laboratório sem equipamento especializado. O segundo relatório de resposta ao etileno bacteriana demonstrou que o etileno aumento da produção de celulose bacteriana e expressão do gene influenciado na bactéria associada à fruta, Komagataeibacter (anteriormente Gluconacetobacter) xylinus 9. Neste caso, o composto libertador de etileno, ácido 2-cloroetilfosfônico (CEPA) foi utilizado para a produção de etileno in situ no interior do meio de crescimento bacteriano, evitando a necessidade de gás de etileno puro ou equipamento especializado.
CEPA produz etileno numa proporção de 1: 1 molar de pH acima de 3,5 através de um 10,11 catalisada por base, a reacção de primeira ordem 12-14. A degradação do CEPA é positivamente correlacionada com o pH e temperatura 13,14 e resulta na produção de acetatoeno, cloreto e fosfato. CEPA fornece aos pesquisadores interessados em estudar respostas bacterianas etileno com uma alternativa conveniente para o etileno gasoso.
O objectivo global dos seguintes protocolos é fornecer um método simples e eficaz para estudar a resposta de etileno bacteriana e inclui a validação de níveis f isiologicamente relevantes de produção de etileno a partir de decomposição CEPA em meio de crescimento bacteriano, análise de pH da cultura para garantir CEPA decomposição não é prejudicada durante crescimento bacteriano, e avaliação do efeito do etileno na morfologia bacteriana e fenótipo. Nós demonstramos esses protocolos usando K. xylinus, no entanto, estes protocolos podem ser adaptados para estudar a resposta ao etileno nas outras bactérias, utilizando o meio de crescimento apropriado e analisa fenótipo.
Access restricted. Please log in or start a trial to view this content.
Protocolo
1. Produtos químicos
- Prepara-se uma solução de CEPA mM 500 (144,49 g / mol), e uma solução que consiste em ambos NaCl 500 mM (58,44 g / mol) e 500 mM de NaH 2 PO 4? H 2 O (137,99 g / mol) em acidificada (pH 2.5) de água ultra-pura ou HCl 0,1 N. Misture usando um vortex até que as soluções são claras.
- Seriadamente diluído (10x) as soluções 500 mM no mesmo solvente para se obter a 5 mM e 50 mM de stocks.
- Prepara-se uma solução de 10 mM de 1-aminociclopropano-carboxílico (ACC; 101,1 g / mol) em água ultra-pura.
- Filtro-esterilizar soluções de reserva e alíquotas armazenar a -20 ° C.
- Filtrar-esterilizar celulase. alíquotas armazenar a 4 ° C.
2. Verificação de produção de eteno a partir de 2-cloroetilfosfônico Ácido decomposição: ensaio de resposta Triplo
- Surface-esterilizar Arabidopsis thaliana Sementes Ecótipo Columbia Usando a fase de vapor Método:
CUIDADO: O passo seguinte produz tgás de cloro ico. Realizar a esterilização de sementes em um exaustor.- Obter um recipiente lacrado profundo o suficiente para acomodar uma proveta de vidro de 250 ml para esterilização de sementes e colocá-lo em um exaustor.
- Adicionar A. thaliana sementes para um tubo de microcentrífuga e colocar o tubo em um rack. Colocar o suporte com o tubo aberto para o recipiente lacrado.
Nota: Não encha tubos individuais mais da metade completo para permitir que o gás cloro para penetrar as sementes mais baixos. - Coloque um copo de 250 ml contendo 100 ml de lixívia comercial no recipiente selável. Adiciona-se cuidadosamente 3 mL de HCl concentrado para a água sanitária e selar imediatamente o recipiente com a sua tampa.
CUIDADO: Bleach e HCl reagem para produzir gás tóxico de cloro que actua à superfície-esterilizar as sementes. - Incubam-se as sementes na presença de cloro gasoso durante 4 horas na fumehood.
Nota: Os tempos de esterilização mais longos irá reduzir a eficiência de germinação. - Após a esterilização, permitir que o chlorine gás de ventilação, na hotte durante pelo menos 1 h, em seguida selar o tubo de microcentrífuga. Deixe a água sanitária e mistura de HCl na fumehood por pelo menos 24 horas antes de descartar.
Nota: As sementes esterilizadas podem ser armazenadas à temperatura ambiente para uso imediato. - Manter as sementes a 4 ° C para armazenamento a longo prazo. Traga sementes à temperatura ambiente antes de abrir o tubo de microcentrífuga para evitar a condensação. sementes armazenar no escuro.
- Prepare placas de ágar em 4 sectoriais placas de petri (90 x 15 mm):
- Preparar 110 ml de meio de crescimento de A. thaliana mudas: 1x Murashige e Skoog (MS), meio basal de 15 (4,33 g / L) contendo 1% (w / v) de sacarose e 0,8% (w / v) de agar. Ajustar o meio MS a pH 6 com NaOH.
- Preparação de 100 ml de meio de crescimento bacteriano para a decomposição CEPA: Schramm e Hestrin (SH) meio 16 contendo 1,5% (w / v) de agar. Ajustar o meio SH-se até pH 7 com NaOH.
- Esterilizar media por autoclavagem e temper em um banho de água a 55 ° C.
- Uma vez que a MS tem de agar temperado, adicionar 40 ml a um frasco estéril. Para preparar o meio de crescimento de controlo positivo para A. thaliana mudas, complementar a 40 ml de aliquota de agar MS com 40 ul de 10 mM ACC para obter uma concentração final de 10 uM ACC.
- Adicionar 5 ml de meio para os quadrantes adequadas das placas de Petri sectored (Figura 1) e deixar o agar solidificar. Prepare todas as placas em triplicado.
Nota: CEPA não é adicionado ao meio de crescimento, até mais tarde no protocolo.

Figura 1:. Configuração de placas de agar utilizadas para o ensaio de resposta tripla com CEPA Um esquema ilustra os quadrantes específicos para o controlo negativo (A), controlo positivo (B), e as placas experimentais (C). Este valor foi modificado a partir Augimeri e Alça 9. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
- Stratify A. thaliana Sementes para garantir Germinação Synchronous:
- Adicionar aproximadamente cinquenta A. sementes thaliana em cada quadrante que contém o MS ou MS + ACC agar. Garantir sementes são distribuídas uniformemente para facilitar a remoção e análise de mudas.
- Incubar as placas contendo as sementes no escuro a 4 ° C durante 3-4 dias.
- Decompor CEPA em Meio de Crescimento Bacteriano (SH) e executar Triplo resposta do ensaio:
- Após a estratificação, expor as sementes à luz fluorescente durante 2 h.
- Espalhe 10 ul da solução de estoque 500 mM CEPA para os quadrantes de as placas experimentais contendo ágar SH (pH 7; Figura 1) para se obter uma concentração final CEPAde 1 mM.
- Selar as placas com filme de laboratório e cobrir com folha para criar um ambiente escuro para as sementes.
- Germinar sementes de placas por incubação no escuro a 23 ° C durante 3 dias com o lado de agar para baixo.
Nota: As placas podem ser empilhadas, mas os controlos negativos devem ser colocados na parte inferior uma vez que o etileno é mais leve do que o ar.
- Analise Triplo resposta do ensaio Dados:
- Com uma pinça esterilizada por chama, retire uma única plântula de uma placa correspondente a cada tratamento e vista sob uma dissecação ou um microscópio USB digital. Assegurar a parte inferior das plântulas são alinhados e fotografia.
- Retirar 30 mudas de cada quadrante (60 plântulas por repetição biológica e 180 mudas por tratamento). Alinhar sobre uma superfície com um fundo preto e uma fotografia com uma régua.
- Meça o comprimento do hipocótilo (mm) de mudas em replicado usando software ImageJ 17. Defina a escala clicando e arrastando o * Liso *ferramenta para seleccionar um comprimento de 10 mm. Selecione "Scale Set" sob a guia "Analisar" e defina a distância conhecida a 10. Usando a ferramenta * segmentado *, clique e arraste para selecionar o comprimento do hipocótilo e pressione M para medir a distância. Comparar as médias de repetições biológicas usando um one-way ANOVA com teste de comparação múltipla de Tukey. As diferenças são considerados significativos se p <0,05.
3. Análise do pH ao longo do crescimento bacteriano
- Cresça e quantificar Komagataeibacter culturas xylinus de arranque em triplicado:
- Inocular uma única colônia de K. xylinus em 5 ml de meio SH (pH 5) suplementada com 0,2% (v / v) de celulase, esterilizada por filtração. Incubar as culturas a 30 ° C com agitação a 150 rpm até uma DO600 de 0,3 a 0,4 é atingida (cerca de 72 hr).
- Colheita de arranque culturas por centrifugação (2000 xg; 4 ° C; 10 min). Com 5 ml estéril a 0,85% (w / v) NaCL solução, lavar as células duas vezes e ressuspender o sedimento celular. Manter as células no gelo.
- Quantificar células usando uma câmara de contagem Petroff-Hausser.
- Análise para inocular culturas de pH:
- Adicionar 150 ml de caldo SH (pH 7) suplementado com 0,2% (v / v) de celulase, esterilizada por filtro de doze frascos de 500 ml com tampas de alumínio. Inocular frascos com K. xylinus culturas de arranque, a uma concentração de 5 a 10 células / ml. Usando as três culturas de arranque, preparar três repetições biológicas por tratamento.
- culturas suplementadas com 300 ul da 5, 50, ou 500 mM de soluções de reserva CEPA para se obter concentrações finais CEPA de 0,01, 0,1, e 1,0 mM, respectivamente. Suplemento as culturas de controlo não tratadas com 300 ul de o solvente utilizado para dissolver CEPA.
- Selar os frascos por firmemente gravando as tampas de folha para os frascos e incubar as culturas durante 14 dias a 30 ° C com agitação a 150 rpm.
- A cada dia, aseptically remover amostras de 5 ml de cada frasco e pelotas de células por centrifugação (2000 xg; 4 ° C; 10 min). Transfira os sobrenadantes para um tubo limpo e medir o pH de cada réplica biológica utilizando um medidor de pH.
- Analisar os dados em tempo-curso representando graficamente o pH médio das repetições biológicas.
Nota: É importante que o pH da cultura não caia abaixo de 3,5; um pH cultura de 5 reduz significativamente a libertação de etileno a partir de CEPA, e um pH abaixo de 3,5 inibe completamente a libertação de etileno. - Para controlar os níveis de cloro e de fosfato, realizar uma experiência idêntica utilizando 0,01, 0,1, e 1,0 mM de 4 a solução de NaCl-se NaH 2 PO utilizando as 5, 50, e os stocks a 500 mm, respectivamente.
4. Colony Morfologia
- Crescer K. Culturas xylinus de arranque, em triplicado:
- Inocular uma única colônia de K. xylinus em 5 ml de meio SH (pH 5) suplementada com 0,2% (v / v) de celulase, esterilizada por filtração. Incubate culturas a 30 ° C com agitação a 150 rpm até uma DO600 de 0,3 a 0,4 é atingida (cerca de 72 hr).
- Colheita de arranque culturas por centrifugação (2000 xg; 4 ° C; 10 min). Com 5 ml de solução salina estéril, lavar as células e em seguida ressuspender o sedimento celular. Manter as células no gelo.
- Prepare a 24 placas de agar contendo 25 ml de SH (pH 7) com 1,5% médios (w / v) de agar.
- Depois de o agar ter solidificado, espalhar 50 ul da 5, 50, e 500 mM de soluções de reserva CEPA sobre o agar para se obter concentrações finais CEPA de 0,01, 0,1, e 1,0 mM, respectivamente. Defina-se placas de controlo não tratadas e de solventes, que consistem em nenhuma alteração e de espalhamento de 50 ul de o solvente utilizado para dissolver os compostos de teste.
- Placas de registo contínuo para colónias isoladas com um loop-cheia (~ 5 mL) de K. xylinus cultura inicial e feche-os com filme de parafina. Inocular todas as placas em triplicado. Incubar as placas durante cinco dias a 30 ° C.
- Photcolónias OGRAFIA com ampliação de 20x usando um microscópio USB digital e qualitativamente avaliar a morfologia e celulose produção colonial em meio sólido.
Nota: Celulose aparece como uma substância turva ao longo da margem da colónia. - Para controlar os níveis de cloro e de fosfato, realizar uma experiência idêntica utilizando 0,01, 0,1, e 1,0 mM de 4 a solução de NaCl-se NaH 2 PO utilizando as 5, 50, e os stocks a 500 mm, respectivamente.
5. Os ensaios película
- Cresça e quantificar K. Culturas xylinus de arranque, em triplicado:
- Em triplicado, inocular uma única colônia de K. xylinus em 5 ml de meio SH (pH 5) suplementada com 0,2% (v / v) de celulase, esterilizada por filtração. Incubar as culturas a 30 ° C com agitação a 150 rpm até uma DO600 de 0,3 a 0,4 é atingida (cerca de 72 hr).
- Colheita de arranque culturas por centrifugação (2000 xg; 4 ° C; 10 min). Com 5 ml de solução salina estéril, lavar tele células duas vezes e ressuspender o sedimento celular. Manter as células no gelo.
- Quantificar células usando uma câmara de contagem Petroff-Hausser.
- Prepare misturas principais para inocular placas de 24 poços:
- Suplemento 60 ml de meio SH (pH 7) com 120 uL de a 5, 50, e 500 mM de stocks CEPA para se obter concentrações finais CEPA de 0,01, 0,1, e 1,0 mM, respectivamente. Suplemento mais 60 ml de meio SH (pH 7) com 120 ul de o solvente utilizado para dissolver CEPA. Vortex para misturar.
- Dividir cada 60 ml de meio contendo CEPA em quatro alíquotas de 14 ml. Inocular três das alíquotas de 14 ml com réplicas biológicas da cultura de arranque a uma concentração de 5 a 10 células / ml. Manter os tubos com as células em gelo para evitar a produção de celulose.
Nota: Os restantes 14 ml de alíquota irá ser utilizado para poços de controlo estéreis.
- Inocular placas de 24 poços:
- Teste cada tratamento em seu próprio prato com três fileiras de repetições biológicas euma linha de controlos estéreis (Figura 2).

Figura 2:. Que ilustra o protocolo utilizado para o ensaio e análise da película CEPA suplementado com meio SH pH 7 (60 ml) Fluxograma é dividido em alíquotas em três inoculações em replicado biológica separados e um controlo estéril (14 ml cada). Estas culturas são então divididas em alíquotas em seis repetições técnicas (2 ml) em uma placa de 24 poços e, em seguida, selados com parafilme. Após a incubação durante 7 dias a 30 ° C, películas são colhidas e caracteriza-se por determinar o peso molhado, espessura, peso seco, e cristalinidade por FT-IR. Por favor clique aqui para ver uma versão maior desta figura
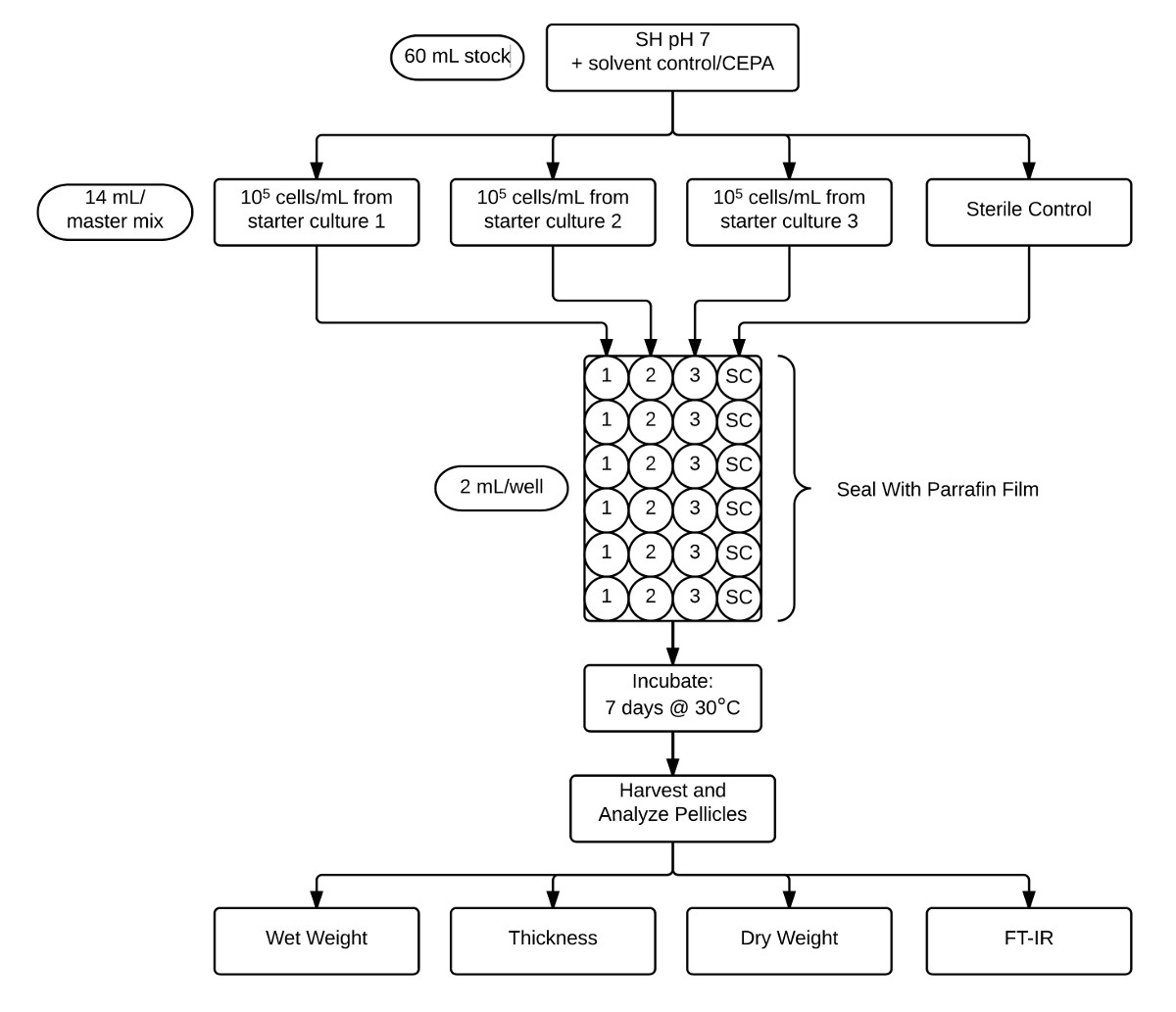{kind=link}
- Usando a 14 ml mistura principal, adicione 2 ml em each de seis poços de uma placa de 24 cavidades estéreis. Completas para os três repetições biológicas e controle estéril (Figura 2). Repetir para cada tratamento.
- placas de vedação com parafilme e incubar estaticamente durante 7 dias a 30 ° C.
- Colheita e Medida Película Wet peso, espessura, peso seco (Rendimento celulose) e a cristalinidade (Figura 2):
- Deprimir a um lado da película para elevar a extremidade da película oposta e remover películas individuais com uma pinça. Apesar de manter aderência, coloque-os sobre toalha de papel fresco durante 3 segundos para remover o excesso médio antes da pesagem para determinar os seus pesos molhados.
- Alinhar películas adjacentes a uma régua e uma fotografia do lado usando uma câmera digital de alta resolução.
- Usando o software ImageJ 17, medir a espessura da película sobre o ombro esquerdo, ombro direito e no centro de cada película. Média de todas as repetições técnicas para cada Replicat biológicae.
- Individualmente transferir as películas para as cavidades de uma placa de 6 poços. Tratar películas com 12 ml de NaOH 0,1 N a 80 ° C durante 20 min para lisar as células.
- Remover o NaOH e neutralizar películas por lavagem com água ultra-pura, durante 24 h com agitação. Mudar a água a cada 6 horas.
Nota: películas deve ser de cor branca após a conclusão da etapa de lavagem. - películas lugar nas esteiras de silício e seco a 50 ° C durante 48 horas até peso constante. Depois de seco, retire do esteiras e medir pesos película em escala analítica para determinar o rendimento de celulose bacteriana.
- Analise película cristalinidade usando Fourier-transform infrared spectroscopy (FT-IR) utilizando 32 varrimentos e uma resolução de 4 cm-1 na gama de 4000 a 650 cm1. Calcular o índice de cristalinidade, CI (IR), usando A 1437 / A 895; a relação de absorção da banda "cristalina" e "banda amorfa" como descrito anteriormente 18.
- Para controlar os níveis de cloro e de fosfato, realizar uma experiência idêntica utilizando 0,01, 0,1, e 1,0 mM de 4 a solução de NaCl-se NaH 2 PO utilizando as 5, 50, e os stocks a 500 mm, respectivamente.
- Analise Pellicle Dados:
- Calcule a hidratação da película, determinando a diferença entre pellicle peso úmido e peso seco.
- Calcular a média dos valores de todas as repetições técnicas para se obter um único valor para cada réplica biológica para análise estatística. Compare tratamentos usando um one-way ANOVA com o teste de comparação múltipla de Tukey. As diferenças são significativas se p <0,05.
- Normalizar os dados como a percentagem de controlos não tratados e traçar os meios de repetições biológicas.
Access restricted. Please log in or start a trial to view this content.
Resultados
Uma configuração de placa esquemática para verificação da libertação de etileno CEPA em meio SH (pH 7) pelo ensaio de resposta tripla é mostrado na Figura 1A - C. Um fluxograma que ilustra o protocolo da película é mostrada na Figura 2. escuro cultivadas A. thaliana mudas exibem o fenótipo resposta tripla (mais curto e mais espessa do hipocótilo com um gancho apical exagerado) na presença da ACC e na presença de et...
Access restricted. Please log in or start a trial to view this content.
Discussão
Os métodos aqui descritos descrevem a produção in situ de etileno a partir de CEPA para o estudo da resposta ao etileno bacteriana usando o organismo modelo, K. xylinus. Este método é muito útil como o etileno pode ser produzido, completando qualquer meio aquoso que tem um pH maior do que 3,5 com 10,11 CEPA eliminando a necessidade de gás de etileno puro ou equipamentos de laboratório especializado. Este método não se limita ao estudo dos efeitos do etileno derivado de CEPA em bact...
Access restricted. Please log in or start a trial to view this content.
Divulgações
The authors have nothing to disclose.
Agradecimentos
The authors thank Dr. Dario Bonetta for providing Arabidopsis thaliana seeds and for technical assistance in regards to the triple response assay, as well as Simone Quaranta for help with FT-IR. This work was supported by a Natural Sciences and Engineering Research Council of Canada Discovery Grant (NSERC-DG) to JLS, an Ontario Graduate Scholarship (OGS) to RVA, and a Queen Elizabeth II Graduate Scholarship in Science and Technology (QEII-GSST) to AJV.
Access restricted. Please log in or start a trial to view this content.
Materiais
| Name | Company | Catalog Number | Comments |
| 1-aminocyclopropane carboxylic acid (ACC) | Sigma | A3903 | Biosynthetic precursor of ethylene in plants |
| 4-sector Petri dish | Phoenix Biomedical | CA73370-022 | For testing triple response |
| Agar | BioShop | AGR001.1 | To solidify medium |
| Canon Rebel T1i DLSR camera | Canon | 3818B004 | For pictures of pellicles |
| Cellulase from Trichoderma reesei ATCC 26921 | Sigma | C2730 | Aqueous solution |
| Citric acid | BioShop | CIT002.500 | For SH medium |
| Commercial bleach | Life Brand | 57800861874 | Bleach for seed sterilization |
| Concentrated HCl | BioShop | HCL666.500 | Hydrochloric acid for pH adjustment |
| Digital USB microscope | Plugable | N/A | For pictures of colonies |
| Ethephon (≥96%; 2-chloroethylphosphonic acid) | Sigma | C0143 | Ethylene-releasing compound |
| Glucose | BioBasic | GB0219 | For SH medium |
| Komagataeibacter xylinus ATCC 53582 | ATCC | 53582 | Bacterial cellulose-producing alphaproteobacterium |
| Microcentrifuge tube | LifeGene | LMCT1.7B | 1.7 ml microcentrifuge tube |
| Murashige and Skoog (MS) basal medium | Sigma | M5519 | Arabidopsis thaliana growth medium |
| Na2HPO4·7H2O | BioShop | SPD579.500 | Sodium phosphate, dibasic heptahydrate for SH medium |
| NaCl | BioBasic | SOD001.1 | Sodium chloride for saline and control solution |
| NaH2PO4·H2O | BioShop | SPM306.500 | Sodium phosphate, monobasic monohydrate for control solution |
| NaOH | BioShop | SHY700.500 | Sodium hydroxide for pH adjustment |
| Paraffin film | Parafilm | PM996 | For sealing plates and flasks |
| Peptone (bacteriological) | BioShop | PEP403.1 | For SH medium |
| Petroff-Hausser counting chamber | Hausser scientific | 3900 | Bacterial cell counting chamber |
| Polyethersulfone sterilization filter 0.2 µm | VWR | 28145-501 | For sterilizing cellulase |
| Sucrose | BioShop | SUC600.1 | Sucrose for MS medium |
| Yeast extract | BioBasic | G0961 | For SH medium |
Referências
- Guzmán, P., Ecker, J. R. Exploiting the triple response of Arabidopsis to identify ethylene-related mutants. Plant Cell. 2 (6), 513-523 (1990).
- Bakshi, A., Shemansky, J. M., Chang, C., Binder, B. M. History of research on the plant hormone ethylene. J. Plant Growth Regul. 34 (4), 809-827 (2015).
- Schaller, G. E. Ethylene and the regulation of plant development. BMC Biol. 10 (1), (2012).
- Hua, J., Sakai, H., et al. EIN4 and ERS2 are members of the putative ethylene receptor gene family in Arabidopsis. Plant Cell. 10 (8), 1321-1332 (1998).
- Bleecker, A. B., Estelle, M. A., Somerville, C., Kende, H. Insensitivity to ethylene conferred by a dominant Mutation in Arabidopsis thaliana. Science. 241 (4869), 1086-1089 (1988).
- Hamilton, A. J., Bouzayen, M., Grierson, D. Identification of a tomato gene for the ethylene-forming enzyme by expression in yeast. Proc. Natl. Acad. Sci. 88 (16), 7434-7437 (1991).
- Kim, J. Assessment of ethylene removal with Pseudomonas strains. J. Hazard. Mater. 131 (3), 131-136 (2006).
- Kim, H. E., Shitashiro, M., Kuroda, A., Takiguchi, N., Kato, J. Ethylene chemotaxis in Pseudomonas aeruginosa and other Pseudomonas species. Microbes Environ. 22 (2), 186-189 (2007).
- Augimeri, R. V., Strap, J. L. The phytohormone ethylene enhances bacterial cellulose production, regulates CRP/FNRKx transcription and causes differential gene expression within the cellulose synthesis operon of Komagataeibacter (Gluconacetobacter) xylinus ATCC 53582. Front. Microbiol. 6, 1459(2015).
- Zhang, W., Wen, C. K. Preparation of ethylene gas and comparison of ethylene responses induced by ethylene, ACC, and ethephon. Plant Physiol. Biochem. 48 (1), 45-53 (2010).
- Zhang, W., Hu, W., Wen, C. K. Ethylene preparation and its application to physiological experiments. Plant Signal. Behav. 5 (4), 453-457 (2010).
- Warner, H. L., Leopold, A. C. Ethylene evolution from 2-chloroethylphosphonic acid. Plant Physiol. 44 (1), 156-158 (1969).
- Biddle, E., Kerfoot, D. G. S., Kho, Y. H., Russell, K. E. Kinetic studies of the thermal decomposition of 2-chloroethylphosphonic acid in aqueous solution. Plant Physiol. 58 (5), 700-702 (1976).
- Klein, I., Lavee, S., Ben-Tal, Y. Effect of water vapor pressure on the thermal decomposition of 2-chloroethylphosphonic acid. Plant Physiol. 63 (3), 474-477 (1979).
- Murashige, T., Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 15 (3), 473-497 (1962).
- Schramm, M., Hestrin, S. Factors affecting production of cellulose at the air/liquid interface of a culture of Acetobacter xylinum. J. Gen. Microbiol. 11 (1), 123-129 (1954).
- Schneider, C. A., Rasband, W. S., Eliceiri, K. W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9 (7), 671-675 (2012).
- Ciolacu, D., Ciolacu, F., Popa, V. I. Amorphous cellulose-structure and characterization. Cellul. Chem. Technol. 45 (1), 13-21 (2011).
Access restricted. Please log in or start a trial to view this content.
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados