
Method Article
ניתוח מותנה של rRNA 2 '-או-מתילציה
In This Article
Summary
כאן אנו מציגים פרוטוקול עבור הפצילות תלויי-RNA של רנ א. זה מאפשר ניתוח מהיר ותלוי האתר של RNA 2 '-או-מתילציה. גישה זו יכולה לשמש להערכה ראשונית או מרכזית של פעילות הסנחנה.
Abstract
תיבת המדריך C/D נוקלאוולאר RNAs (נחלות) לזרז 2 '-O-מתילציה של ריבוזומאז ו-RNA גרעיני קטן. עם זאת, מספר רב של שנחורה ב eukaryotes גבוה יותר עשוי promiscuously הכיר מינים אחרים RNA ו 2 '-O-מתיונין מטרות מרובות. כאן, אנו מספקים מדריך צעד אחר צעד עבור ניתוח מהיר ולא יקר של האתר הספציפי 2 '-O-מתילציה באמצעות שיטה מבוססת היטב העסקת DNA olig, באופן שימוש קצר הנקרא DNAzymes. שברי ה-DNA הללו מכילים רצפים קטליטיים אשר קליב RNA בתנוחות הסכמה ספציפיות, כמו גם זרועות הומולוגיה משתנה המכוונים למטרות של מטרות RNA שלה. פעילות דנזולי מעכבות ב -2-' הו-מתילציה של הנוקלאוטיד הסמוך לאתר המחשוף ברנ א. לפיכך, דנזאמס, מוגבל רק על ידי הקונצנזוס של הרצף ביקע, הם כלים מושלמים לניתוח מהיר של השנחא-תיווך 2 '-או-מתילציה. ניתחנו את הsnR13-ו-snR47-מונחים 2 '-O-מתילציה של ה-RNA של ה-25 הריבוזומאז ב סכביטוסיאה , כדי להדגים את הפשטות של הטכניקה ולספק פרוטוקול מפורט לגבי הצורך התלוי בשיטת Dנזיימה.
Introduction
שינויים RNA לשחק תפקידים חשובים בוויסות של ביטוי גנים. Rna 2 '-O-מתילציה ו פסבדו, אשר מונחים על ידי תיבת C/D ו תיבת H/ACA nucleolar קטן rnas (שנחולים) בהתאמה, להגן על RNA מפני השפלה וייצוב מבנים מסדר גבוה שלהם1,2,3 . מטרות שנחנה זוהו בעיקר RNAs (rRNA) ו-RNAs גרעינית קטנה (snRNAs). עם זאת, ב eukaryotes גבוה יותר, יש פוטנציאל מאות של שנחנה עם פונקציות לא מוקצה וחלקם עשויים לזהות rnas מרובים1,4,5,6,7. לכן, שיטות המאפשרות זיהוי וניתוח של שינויי שינויים מודרכים הם כלים חשובים במנגנוני גילוי המסדירים תהליכים סלולאריים.
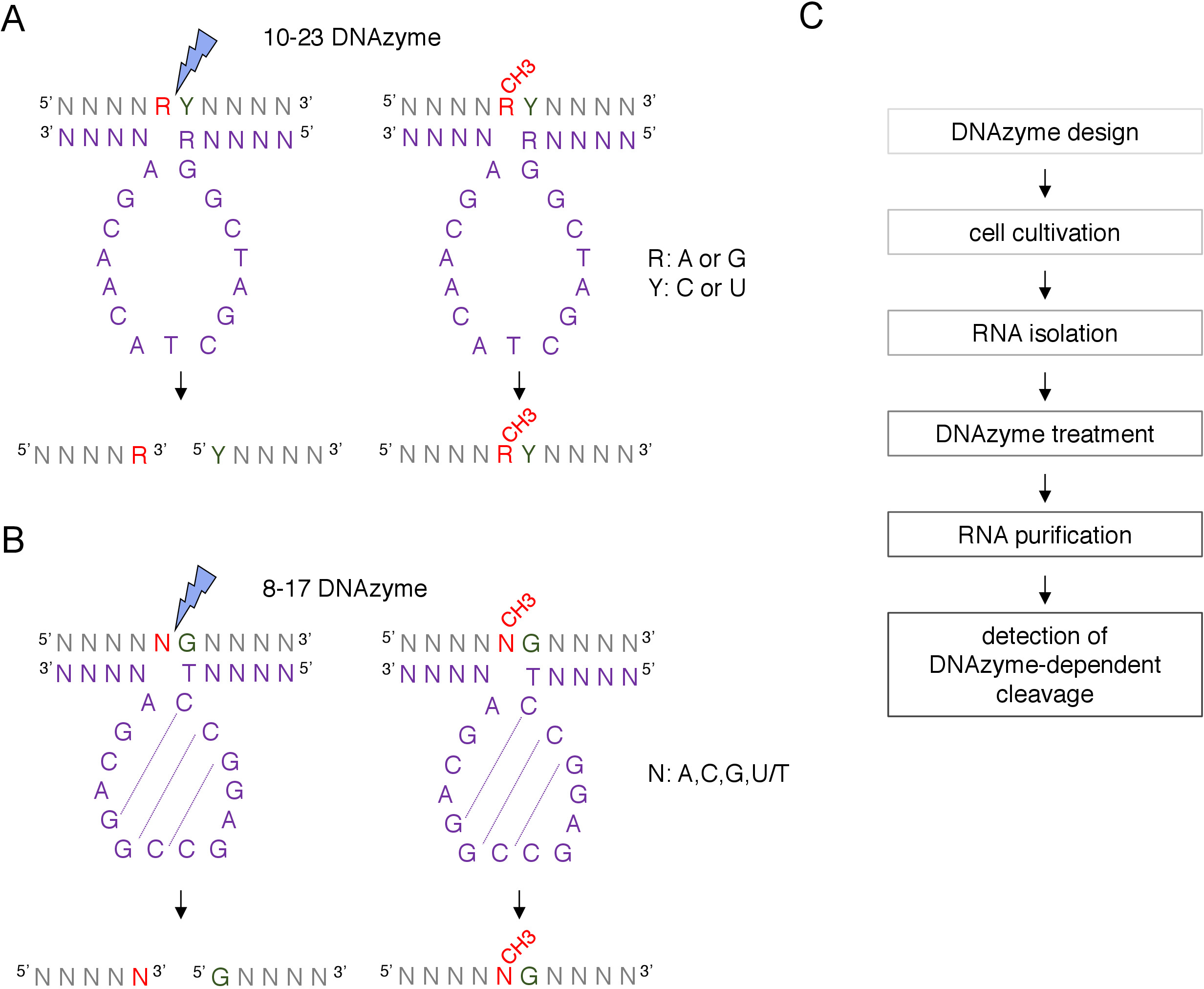
בתיבת C/D שננחנה-מונחה באתר 2 '-O-מתילציה ניתן לזהות ביולוגי ומאושר ניסויים על ידי טכניקות רבות, כולל RNase H בבימויו של מחשוף, או ספציפי לאתר ושיטות הגנום-רחב, אשר מעסיקים שעתוק הפוכה באמצעות הריכוז של נוקלאוטידים נמוכים (dntps)8,9,10,11. טכניקות אלה רגישות מאוד, אבל גם מפרך ויקר, לכן, לא יכול להיות מתאים הבדיקה הראשונית או מהירה. אחת השיטות הפשוטות והנמוכות ביותר כדי לזהות 2 '-O-מתילציה אתרים הם מחשוף RNA התלוי ברנ א12. DNAzymes הם קצרים, חד-תקועים, מולקולות DNA פעיל מזרז המסוגל לחשוף את המחשוף של ה-RNA בתנוחות מסוימות. הם מורכבים רצף ליבה שימור ומזרז פעיל ו 5 ' ו 3 ' זרועות קשירה מורכב רצפי משתנים שנועדו לhybridize על ידי ווטסון-קריק שיוך בסיס ליעד RNA (איור 1). לפיכך, הזרועות 5 ' ו-3 מספקים את רצף הקטליטי לאתר ה-RNA הספציפי. המחשוף התלוי בדנזמי מעוכב על ידי 2 '-O-מתילציה של הנוקלאוטיד ממוקם ישירות במעלה הזרם של המחשוף12,13. הדבר מאפשר כלים מעשיים מאוד לניתוח של אתרי רנ א או הידוע ב-RNA 2 מתילציה.
שני סוגים של DNAzymes משמשים עבור שינויי RNA ניתוח12. הרצף הפעיל של 10-23 דנזאלי (איור 1A) מורכב מ-15 נוקלאוטידים (5 ' RGGCTAGCTACAACGA3 ') אשר יוצרים לולאה סביב RNA purine-pyrimidine (RY) dinucleotide ומזרז את המחשוף בין שני נוקלאוטידים. ה-RNA פורין (R) אינו מזווג בסיס עם הדנזאני ושני המתילציה מעכבים את המחשוף. הזרועות הכבילה של 10-23. בדרך כלל 10-15 נוקלאוטידים ארוכים הכיתה השנייה של דנזלי, 8-17 דנזיימס (איור 1B) מכילות 14-נוקלאוטיד (5 ' TCCGAGCCGGACGA3 '). נוקלאוטידים C2, c3 ו-g4 זוגות עם C9 G10 ו-g11 ויוצרים מבנה קצר לולאה. 8-17 dnazymes קליב בזרם RNA של כל גואנין כי הוא מזווג באופן מושלם עם תימין הראשון מן הרצף הפעיל dנזיימה. ה-RNA נוקלאוטיד במעלה הזרם של הגואנין אינו מזווג בסיס עם דנזלי והשניים שלו-או-מתילציה מכהים את המחשוף. 8-17 DNAzymes דורשים זרועות הומולוגיה ארוכות יותר של כ -20 נוקלאוטידים כדי להפנות את הקשר לרצף המסוים שלו.
כאן, אנו מספקים פרוטוקול צעד אחר צעד לניתוח של 2 '-O-מתילציה של rrna ב סכביתאז cerevisiae ס באמצעות 10-23 ו 8-17 dnazyme-גישות תלויות12,13 (איור 1c). פרוטוקול זה ניתן להתאים בקלות עבור אורגניזמים אחרים ומינים RNA ומועסק עבור ניתוח מהיר, ראשוני או מרכזי של RNA ספציפי לאתר 2 '-O-מתילציה.
Protocol
1. זנים, מדיה, מאגר מתכונים
- להכין מדיה שמרים (S. cerevisiae ס) כפי שמפורט להלן: YP (1% w/v תמצית שמרים, 2% w/v פאיוולוגית), ו גלוקוז ו גלקטוז מניות ב 20% w/v.
- הכנת הנתרן אצטט (NaAc)-EDTA (AE) מאגר כמפורט כאן: 50 mM NaAc pH 5.3 ו 10 מ"מ EDTA.
- הכינו 10-23 מאגר הדגירה 4x של Dנזלי מפורט כמפורט כאן: 24 mM Tris pH 8.0, 60 mM הנאל ו 10-23 מאגר התגובה 4x של Dנזאלי: 200 mM טריס pH 8.0 ו 600 מ"מ הנאל.
- הכינו 8-17 Dנזאלי 2x מאגר התגובה כמפורט כאן: 200 mM KCl, 800 mM הנאקל, 100 mM HEPES pH 7.0, 15 מ"מ MgCl2, ו 15 מ"מ mgcl2.
- הכינו מאגר מגבים 10x כמפורט להלן: 200 מ"מ בלבד, 50 mM NaAc, 1 מ"מ EDTA; pH 7.0 ו-1.5 x לדוגמה מאגר מיקוד: 50% v/v הטופס, 20% v/v פורמלדהיד, 1.5 x מאגר מגבים.
- השג זנים של ס. cerevisiae ס , BY4741 (מאטה His3Δ1 leu2Δ0 met15Δ0 ura3Δ0); GAL1:: SNR13 (כמו BY4741 אבל GAL1:: SNR13: KANmX); GAL1:: SNR47 (כמו BY4741 אך GAL1:: SNR47: HIS3mX). כל זן שמרים אחרים יכול לשמש ניתוח זה.
2. עיצוב מעצבים
- מצא את רצף הריבית או האתר של מתילציה באמצעות מסד נתונים מתאים. עבור S. cerevisiae ס מטרות נחנה, השתמש שמרים מסדי נתונים: http://people.biochem.umass.edu/fournierlab/snornadb/mastertable.php14
- כדי למצוא את אתר מתילציה של עניין, למשל, snR13 תלוי באתר, בחר "snR13" ולרשום את המיקום של נוקלאוטיד שונה (למשל, snR13-מונחה A2281 ב-25 rrna).
- למצוא רצפים במעלה ובמורד הזרם של נוקלאוטיד שונה באמצעות מסד נתונים המתאים. עבור S. cerevisiae, השתמש במסד הנתונים של הגנום הסכקוע: https://www.yeastgenome.org/
- חפש את שם הגן היעד לדוגמה, RDN25 (בקידוד 25S rRNA).
- מתוך "רצף" הכרטיסייה, בחר 10-15 נוקלאוטידים במעלה הזרם (5 ' זרוע) ו במורד הזרם (3 ' זרוע) של האתר מתילציה כאשר באמצעות מעבד 10-23 dנזלי ו 20 נוקלאוטידים במעלה הזרם (5 ' זרוע) ו במורד הזרם (3 ' זרוע) של האתר מתילציה עבור 8-17 dנז
- ליצור רצפים משלימים של 5 ' ו 3 ' זרועות.
- אגף 10-23 או 8-17 הרצף הקטליטי עם רצפים משלימים של 5 ' ו-3 ' זרועות.
- תזמיני ממני בתור דנ א רגיל. של הדנ א מהספק
3. S. cerevisiae ס תנאי צמיחה
הערה: S. cerevisiae ס BY4741 נגזרות הזנים שימשו, שבו הביטוי של SNR13 או SNR47 שנחורה מונע מיזם inducible GAL1 . כדי לזרז או לעכב את הסינתזה שלהם, לגדל תאים על בינוני המכיל גלקטוז (GAL1תלויי שעתוק) או גלוקוז (שעתוק תלויGAL1). כפקד, השתמש זן סוג פראי (BY4741) גדל או על גלקטוז או גלוקוז.
- לגדל זנים שמרים במדיום המתאים התנאים. כדי לנתח GAL1:: SNR13 ו GAL1:: SNR47 זנים כמו גם זן מסוג פראי האיזוגניים, לגדול תאים 50 ML של מדיית YP עם 2% גלוקוז (YPD) או גלקטוז (ypgal) ב 30 ° c לשלב האקספוננציאלי האמצעי.
- התאים צנטריפוגה ב 1,000 x g, עבור 3 דקות ב 4 ° c.
- . התעלם מהעל-חושית ושמור את החבילות
- הקפאת כדורי תא בחנקן נוזלי ואחסן אותם ב-80 ° c.
זהירות: חנקן נוזלי עלול לגרום. לכוויות מקריוגניים חמורות תמיד לובשים בגדי הגנה ומפעילים אמצעי זהירות.
הערה: ניתן לאחסן את כדורי התא ב-80 ° c עד חודש. הפרוטוקול יכול להיות מושהה כאן במידת הצורך.
4. רנ א בידוד15
הערה: השתמש בשיטה המתאימה ביותר כדי לבודד RNA. עבור שמרים S. cerevisiae ס, חם-פנול הוצאת RNA ניתן להשתמש.
- להוסיף 1 מ ל של מים קרים קרח, להשהות מחדש את כדורי והעברת תאים מחדש כדי 1.5 mL מיקרו צינורות.
- צנטריפוגה ב 20,000 x g עבור 10 s ב 4 ° c ולהסיר את הסופרנטאנט.
- הוסף 400 μL של מאגר AE והשהה מחדש את התאים.
הערה: השלבים 4.4-4.15 מתבצעים בטמפרטורת החדר, אלא אם נכתב אחרת. - הוסף 40 μL של 10% SDS ו 400 μL של חומצה פנול (pH 4.5).
זהירות: פנול רעיל ויש לטפל. בו מתחת למכסה המנוע תמיד ללבוש חלוק מעבדה, כפפות מגן, ומשקפיים כאשר עובדים עם פנול. היפטר מפסולת בהתאם לתקנות המוסדיים. - מערבבים היטב על ידי vortexing עבור 20 s.
- מודקון ב 65 ° c עבור 10 דקות. כל 2 דקות, בעדינות לפתוח ולסגור את הצינור כדי לשחרר את הלחץ ולהפוך את הצינור 2-3 פעמים כדי לערבב את השלבים.
- להעביר את הצינורות ל-80 ° c ו דגירה עבור 10 דקות.
- להפשיר את הצינורות על הספסל ואת צנטריפוגה ב 20,000 x g, עבור 5 דקות בטמפרטורת החדר.
- העבר את השלב העליון לצינור חדש המכיל 400 μL חומצה פנול: כלורופורם: isoamyl אלכוהול (25:24:1). אין לשבש את השלב הפנימי.
זהירות: כלורופורם הוא רעיל ויש לטפל מתחת למכסה המנוע. תמיד ללבוש חלוק מעבדה, כפפות מגן, ומשקפיים כאשר עובדים עם כלורופורם. היפטר מפסולת בהתאם לתקנות המוסדיות. - מערבבים היטב על ידי vortexing עבור 30 s ו צנטריפוגה ב 20,000 x g עבור 10 דקות בטמפרטורת החדר.
- העבר את השלב העליון (~ 400 μL) לצינורית חדשה המכילה 400 כלורופורם μL.
- מערבבים היטב על ידי vortexing עבור 30 s ו צנטריפוגה ב 20,000 x g עבור 5 דקות בטמפרטורת החדר.
- העבר את השלב העליון (~ 300-350 μL) לצינור חדש המכיל 1 מ ל של אטוח ו 40 μL של 7.5 M אמוניום אצטט (NH4AC). מערבבים על ידי היפוך הצינור כמה פעמים.
- מ80 מעלות צלזיוס עבור 2 מעלות או לילה ב-20 ° c.
הערה: ניתן להשהות כאן את התהליך. - צנטריפוגה ב 20,000 x g, עבור 10 דקות ב 4 ° c. קטן, גלולה לבנים RNA יהיה גלוי בתחתית הצינור.
- מסירים את האטואה על ידי מלטף כדי למנוע הפרעה הגלולה.
- הוסף 1 מ"ל של 70% אטוח ו צנטריפוגה ב 20,000 x g עבור 5 דקות בטמפרטורת החדר.
- הסר 70% אטוע על ידי ליטוף.
- צנטריפוגה ב 20,000 x g עבור 15 s ולהסיר את אטוח הנותרים עם 2-20 μl הצינורות.
- להשאיר את הצינור פתוח על הספסל 5 דקות לייבש את הגלולה RNA.
הערה: גלולה RNA משנה את צבעו מלבן לשקוף כאשר יבש. - השהה מחדש את גלולה RNA ב 30 μL של RNase/DNase-H חינם2O, להעביר את הצינור מיד על הקרח למדוד את הריכוז RNA על מיקרוספקטרוסקופיה.
- הקפא את הדגימות ב-20 ° c.
הערה: רנ א ניתן לאחסן ב-20 ° c עד 1 לחודש ו-80 ° צ' עד 1 עד שנה. ההליך יכול להיות מושהה כאן או המשיך ישירות לשלב הבא.
5. מינון העיכול
-
10-23 מינון העיכול
- בשנת 1.5 mL צינורות להכין מיקס דגירה על ידי שילוב של 5 μg של RNA, 200 pmol של 10-23 DNAzyme (2 μL של 100 הפתרון מניות mM) ו-2.5 μL של מאגר דגירה של 4x 10-23 בנפח כולל של 10 μL. שמרו על שפופרות קרח.
- להעביר את צינורות לחסום חום יבש להגדיר ב 95 ° c ו דגירה עבור 3 דקות.
- להעביר את הצינורות מיד על הקרח ו-דגירה עבור 5 דקות.
- הסתובב לזמן קצר והחזר את הצינורות בחזרה לקרח.
- הוסף 20 U של RNase מעכב (למשל, 0.5 μL הריבוlock RNase מעכב).
- מניחים את הצינורות בבלוק חום יבש להגדיר 25 ° c ו דגירה עבור 10 דקות.
- בינתיים, להכין תערובת תגובה ב-1.5 μL שפופרת על ידי שילוב של 5 μL של 4x 10-23 מאגר התגובה עם 4 μL של 300 מ"מ MgCl2 ו-1 Μl H2O. מניחים את הצינור בבלוק יבש מוגדר כדי 37 ° c.
- העבר את ערבוב הדגירה לבלוק חום יבש להגדיר עבור 37 ° צ' ולהוסיף 10 μL של ערבוב מראש מחומם.
- מודיית את התגובה בשעה 37. מעלות צלזיוס לשעה אחת
- להעביר את הצינורות על הקרח ולהמשיך שלב 5.3.1.
-
8-17 מינון העיכול
- הכנת מיקרוtube 1.5 mL עם 5 μg של RNA בנפח כולל של 6 μL.. שמור על השפופרת על הקרח
- להכין מיקרוtube 1.5 mL עם 400 pmol של 8-17 DNAzyme (4 μL מתוך מניות 100 mM). . תשאיר את השפופרת על הקרח
- להעביר את צינורות לחסום חום יבש להגדיר עבור 95 ° c ו דגירה עבור 2 דקות.
- הזיזו את דגימת ה-RNA על הקרח.
- הסתובב בתוך הצינור עם Dנזמי עבור 5 s ו דגירה ב 25 ° c עבור 10 דקות.
- באותו זמן, להכין שפופרת 1.5 mL עם 10 μL של 2x 8-17 מאגר התגובה ואת הדגירה ב 25 ° c.
- הכינו תערובת תגובה על ידי הוספת 10 μL של מאגר התגובה מראש 2x מחומם לצינור עם DNAzyme.
- העבר 14 μL של תערובת התגובה לצינור עם RNA ולהוסיף 20 U של מעכב RNase.
- מודג את התגובה ב -25 ° c עבור 2 h.
- להעביר את הצינור על הקרח ולהמשיך לטיהור RNA (שלב 5.3.1).
-
רנ א-טיהור
- הוסף 350 μL של מים ו 400 μL של כלורופורם לצינור התגובה, לערבב גם על ידי vortexing עבור 30 s ו צנטריפוגה ב 20,000 x g עבור 5 דקות בטמפרטורת החדר.
- העבר את השלב העליון (~ 300-350 μL) לצינורית חדשה המכילה 1 מ ל של אטוח, 40 μL של 7.5 M NH4AC ו 1 μl של הגליקוגן (10 μg/μl). מערבבים על ידי היפוך הצינור כמה פעמים.
- מ80 מעלות צלזיוס עבור 2 מעלות או לילה ב-20 ° c.
הערה: ניתן להשהות כאן את התהליך. - חזור על הצעדים מ 4.15 עד 4.21.
- להשעות מחדש את הגלולה RNA ב 10 μL של RNase/DNase-H חינם2O ולהעביר את הצינורות מיד על הקרח.
- הקפא את הדגימות ב-20 ° c.
הערה: רנ א ניתן לאחסן ב-20 ° c עד חודש ו-80 ° c עד שנה. ההליך יכול להיות מושהה כאן או להמשיך לאלקטרופורזה RNA.
6. רנ א אלקטרופורזה
- רסס את ציוד האלקטרופורזה (טנק, מגש, מסרק) עם 1% SDS, לעזוב 15 דקות ולשטוף עם הרבה של ddH2O.
- מתמוסס 1.5 g של agarose ב 127.5 מ ל של ddH2O על ידי לחמם אותו במיקרוגל.
- הוסף 15 מ ל של מגבים 10x ו 7.5 mL של 37% פורמלדהיד לפתרון agarose (הנפח הכולל הוא 150 mL).
זהירות: פורמלדהיד רעיל ויש לטפל בו מתחת למכסה המנוע. תמיד ללבוש חלוק מעבדה, כפפות מגן, ומשקפיים כאשר עובדים עם פורמלדהיד. היפטר מפסולת בהתאם לתקנות המוסדיות. - הוסף כמות מתאימה של כתם ג'ל הבחירה לפתרון agarose (למשל, 15 μL של SYBR בטוח ג'ל DNA כתם). מערבבים היטב ויוצקים את הצמח על מגש.
- הכנס מסרק לתוך הג מיד.
- . תשאיר את זה ל 45 דקות מתחת למכסה המנוע מכסים את המגש ברדיד אלומיניום כאשר משתמשים בכתם ג'ל רגיש לאור.
- הכן 600 mL של מאגר מגבים 1x.
-
הכנה לדוגמא RNA
- ב שפופרת 1.5 mL, לשלב 10 μL של המדגם RNA מתעכל ומטוהרים, 5 μL של מאגר מדגם Denaturing רפתקאות ו 0.5 μL של 6x טעינת צבען.
זהירות: , הטופסרעל רעיל. ויש לטפל בו מתחת למכסה המנוע תמיד ללבוש חלוק מעבדה, כפפות מגן, ומשקפיים כאשר עובדים עם הטופסלמיד. היפטר מפסולת בהתאם לתקנות המוסדיות. - מודטה ה-RNA דגימות ב 70 ° c עבור 5 דקות. להעביר את הדגימות על קרח. דגירה של 5 דקות.
- התנדנד למטה לזמן קצר לפני טעינת ג'ל.
- ב שפופרת 1.5 mL, לשלב 10 μL של המדגם RNA מתעכל ומטוהרים, 5 μL של מאגר מדגם Denaturing רפתקאות ו 0.5 μL של 6x טעינת צבען.
- לשים את הג במיכל האלקטרופורזה ולמלא עם מאגר מגבים 1x. טען את הנפח כולו של כל מדגם (15 μL) על ג'ל. הפעל ב 80 V עד ברומאופנול כחול מגיע 2/3 של אורך ג'ל.
- התמונה ג'ל באמצעות צמידה מתאים לזהות את הכתם ג'ל שנבחר (למשל, הינע UV).
תוצאות
שירות המחשוף התלוי בניתוח של שינויי rRNA הוצג לאחרונה בהקשר של התבגרות שנחונות13. הטיפול התלוי בדנזלי שימש כדי להראות כי חוסר של 5 '-end טרום העיבוד התהליך משפיע על 2 '-O-מתילציה רמות של 25S ו-18 rRNA ב S. cerevisiae ס13.
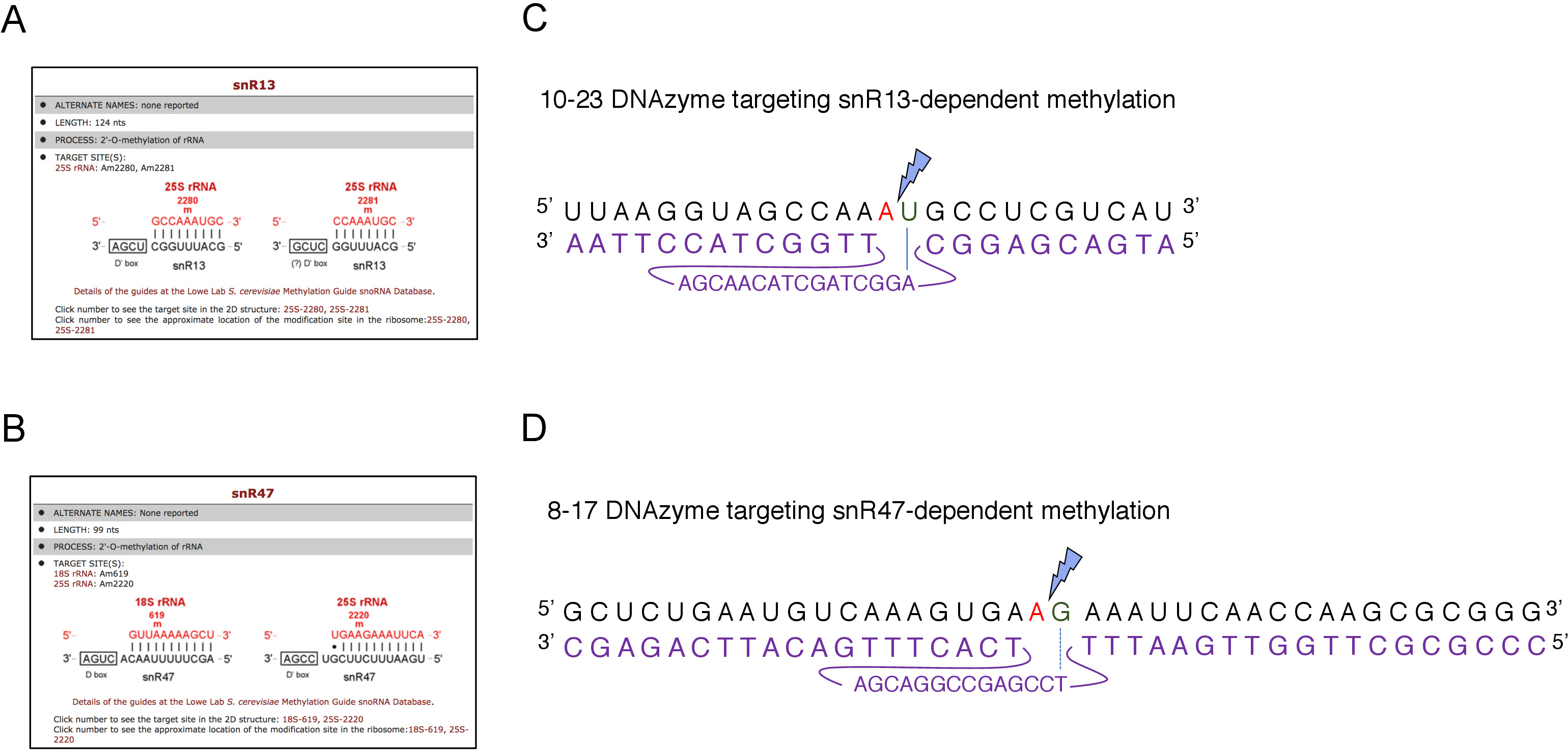
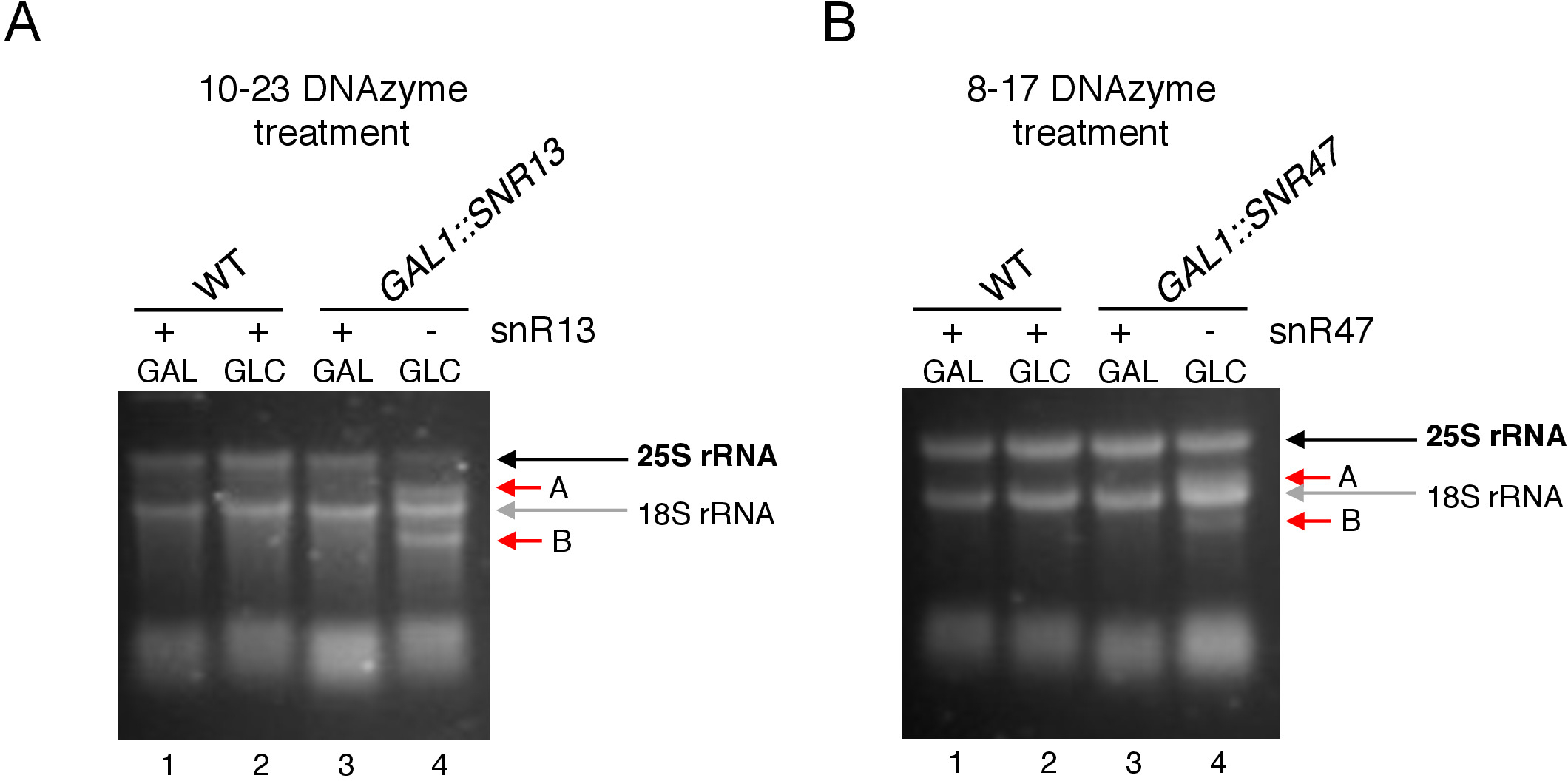
כאן, השתמשנו מערכת תמלול inducible Na להפגין את האפקטיביות ואת הפשטות של הטכניקה. תיבת C/D snR13 מדריכים מתילציה בשתי עמדות ב-25 rrna, כולל אדנין 2281 (איור 2a). הנוקלאוטיד הזה אחריו uracil, אשר מהווה את ההסכמה הקונצנזוס (RY) להיות בעלי הסכמה על ידי 10-23 DNAzyme. תיבת C/D snR47 גם מדריכים מתילציה של שני נוקלאוטידים ב-25 rRNA (איור 2B). האדנין במיקום 2220 מלווה בשאריות גואנין, ואפשר לקבל את החומר הזה על ידי מ8-17 דנזאלי. על מנת לזרז או לעכב את הסינתזה של snR13 או snR47, הוספת את inducible GAL1 יזם במעלה של snR13 או snR47 גנים ותאים מעובדים בינונית המכילה גלקטוז (GAL1תלוי שעתוק) או גלוקוז (שעתוק תלויGAL1). לאחר מכן, RNA מבודד מ GAL1:: SNR13 תאים היו מודבטים עם 10-23 DNAzyme שנועד לקליב Rrna 25 באתר תלוי SNR13, בין נוקלאוטידים 2281 ו 2282 (איור 2c). RNA מ GAL1:: SNR47 המתח טופל עם 8-17 DNAzyme מיקוד האתר תלוי SNR47 בין נוקלאוטידים 2220 ו 2221 (איור 2d). כפקד, רנ א מן פראי סוג BY4741 הזנים גדל על גלקטוז או גלוקוז היה מודבטים עם שני הצדדים. אלקטרופורזה של ה-RNA המטופל בדנזאלי חשף כי 25S rRNA הופק מ GAL1:: SNR13 ו GAL1:: SNR47 זנים הגדלים על גלקטוז (גל) נותרו שלמים (איור 3 א,ב; נתיבים 3). לעומת זאת, RNA מבודד מ GAL1:: SNR13 ו GAL1:: SNR47 תאים הגדלים על גלוקוז (glc) היה מתעכל על ידי dnazymes בהתאמה (איור 3a, B; נתיבים 4). בשני המקרים, הלהקה rRNA של 25 ירד ו 5 ' ו 3 ' קיצוץ מוצרי מחשוף (A ו-B) נצפו. הדבר מצביע על כך ב- GAL1:: SNR13 ו- GAL1:: SNR47 זנים, ה-25s rrna היה 2 '-O-מתיליום ב SNR13-או SNR47-אתרים מודרכים כאשר גלקטוז שימש כמקור פחמן ושנחורה אלה ביטאו. העדר rRNA מתילציה כאשר snR13 או snR47 הביטוי היה נכבה על גלוקוז מותר על המחשוף תלוי Dנזיימה. לא הייתה מידה של עיכול RNA לדגימות מסוג פראי (איור 3 א,ב; נתיבים 1 ו-2), כביטוי snR13 ו-snR47 הוא גלקטוז/גלוקוז עצמאי במאמץ זה. לכן, rRNA היה מוקשה בדרך כלל ועמיד כל כך לפעילות DNAzymes.
בסך הכל, הניסוי שלנו מראה כי פעילות המחשוף של 10-23 (איור 3A) ו 8-17 (איור 3a) בקורלציה עם היעדר תיבת C/D snR13 או snR47, בבירור המציין כי שנחרות אלה אחראים 25 rrna 2 '-O-מתילציה באתרים מסוימים.

איור 1: DNAzymes ומצעים RNA שלהם. (א) 10-23 דנזיימס קליב א-פירימיטין (רי) רנ א. R ב-RNA אינו משויך לדנזמי, ואילו Y משלים את הבסיס R בדנזיימי. מתילציה של הפורצין (R) ב-RNA מדכא את המחשוף התלוי בדנזיימי. (ב) 8-17 דנזולימס במעלה ה-RNA של הזרם של גואנין, שאינו מזווג באופן מושלם עם התימין הראשון ברצף הקטליטי של דנזאלי. הנוקלאוטיד שלפני גואן-טין אינו משויך והתילציה שלו מגינה מפני המחשוף התלוי בדנזמי. רנ א מוצג באפור (מלבד אתר מתילציה), Dנזיימה מוצג בסגול. N = כל נוקלאוטיד, R = purine: אדצין או גואנין, Y = pyrimidine: ציטוסינוס או uracil; CH3-מציין RNA מתילציה. זיווג בסיסי בתוך הרצפים הפעילים של DNAzyme מסומן בקווים מנוקדים. ברק כחול מסמן את אתר המחשוף. (ג) תרשים זרימה המציג את השלבים של ניתוח תלוי-באמצעות Dנזלי. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: התמקדות באתרי מתילציה snR13-ו-snR47 התלויים ב-25S rRNA. (א, ב) צילומי מסך הדפדפןמראהsnR13 תלויי (a) ו-snR47 תלויי (B) אתרי מתילציה ב-25s rrna (C) 25s ברצף של מתילציה התלויים snR13 (A2281) ו 10-23 dנזלי (מוצג בסגול) נועד לקליב RNA בין A2281 ו-U2282. 2281 לא מזווג עם dנזלי בעוד U2282 טפסים זוג עם הנוקלאוטיד הראשון מתוך הרצף הפעיל של dנזיימה (מסומן בקו כחול). ברק כחול מסמן את המחשוף. (ד) 25s הרצף rrna המקיף snR47 באתר מתילציה תלוי (A2220) ו 8-17 dnazyme (המוצג בסגול) נועד קליב RNA בין2220 ו-G2221. 2220 הוא לא הוכלא עם dנזלי בעוד G2221 מזווג באופן מושלם עם תימין (מסומן בקו מקווקו). ברק כחול מסמן את אתר המחשוף. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3: ניתוח ספציפי לאתר 2 '-O-מתילציה של 25S rRNA באמצעות 10-23 ו 8-17 DNAzyme הנתון התלוי. (א) אנליזה של מתילציה snR13 התלויים ב -25, באמצעות 10-23 דנזיימי. (ב) ניתוח מתילציה של 25 snR47 התלויים בשימוש ב-8-17 Dנזלי. RNA היה מכתים את הצמח בג. מוצרי המחשוף A ו-B מסומנים בחיצים אדומים. WT = זן מסוג פראי; גל = גלקטוז, GLC = גלוקוז. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
Discussion
העיכול התלוי בדנזמי יכול לשמש כשיטה פשוטה ומהירה לניתוח ה-RNA הספציפי לאתר 2-O-מתילציה12,13. כאשר הנוקלאוטיד במעלה הזרם של אתר המחשוף אינו מתילזה. בניגוד לגישות אחרות, כולל העיכול rnase H בבימויו, השפלה בסיסית או תמלול הפוכה בריכוז נוקלאוטידים נמוך ואחריו PCR כמותי או רצף8,10,11 ,16, הגישה dנזלי דורש דנ א פשוט olig, וריאגנטים בסיסי הנמצאים בכל מעבדה ביולוגיה מולקולרית. יתרה מזאת, ניתן להשתמש ב-DNAzymes בדרך דומה לניתוח של RNA מתווך באמצעות תיבת H/ACA שנחנה12, מה שהופך אותם לכלים מגוונים בלימוד מטרות השנחנים.
הגישות התלויות בדנזבי מוגבלות רק באמצעות מחשוף הקונצנזוס באתר ברצף17. 10-23 DNAzymes ניתן להשתמש כדי לנתח 2 '-O-מתילציה רק במיקום R של הדיקלקסאות, בעוד 8-17 DNAzymes להכיר את השינוי של נוקלאוטיד הממוקם במעלה הזרם של גואנין. כתוצאה מכך, שינויים כמו 2 '-O-מתילציה של הנוקלאוטיד הראשון של dinucleotides גואנין-אדנין (GA), אדנין-אדנין (AA), pyrimidine-אדנין (YA) ו pyrimidine-pyriine (YY) לא ניתן לנתח. כמו-כן, יש לשקול את היעילות הנמוכה של המחשוף התלוי ב-DNAzyme12 . למרות שחלק מהאתרים מסוימים כמעט לחלוטין (איור 3b), מספר האנשים הרבים מעכל רק חלקית את המטרות שלהם (איור 3b). היעילות עשויה להיות תלויה ברצף שמקיף את אתר המחשוף. לדוגמה, אזורי RNA עם מתיחות של אותו נוקלאוטיד עשויים להשפיע על המיקום הנכון של הרצף הפעיל של Dנזיימה. יתר על כן, אזורי RNA היוצרים מבנה משני חזק עלול לhybridize מחדש ולדכא את הכריכה של דנזיימי לרצף היעד. כדי להתגבר על בעיות אלה, מחזורים של חימום וקירור של 10-23 DNAzyme ומצע ה-RNA שלה ניתן להחיל18.
השתמשנו בגישה של הדנזאלי לחקור 2 '-או-מתילציה של rRNA. אחד יכול גם להשתמש בטכניקה זו כדי לנתח שינויים אחרים RNA, כגון N6-methyladenosine19. רנ א ריבוזומלית, בשל השפע שלה, ניתן לנתח על ידי אלקטרופורזה ומוצרי המחשוף ניתן לדמיין תחת אור UV. עם זאת, זה לא ישים עבור RNAs שופע פחות כמו RNA פולימראז השני שנוצר קידוד RNAs (mRNA) ו-RNAs לא קידוד (ncRNA). RNAs אלה בדרך כלל לא ניתן להבחין ישירות על-ידי כתמים RNA בתוך הצמח או הפוליאקרילאמיד ג'ל. במקרים כאלה, מחשוף התלוי בפני הכתמים הצפוניים, מזוהה בעקיפין על ידי ה-pcr/PCR כמותי או מנותח על-ידי PCR כמותי עם פולימות (למשל, KlenTaq DNA פולימראז) המסוגל להפלות 2-O-מתילאז-RNA מ רנ א בלתימתילבן 20,21.
Disclosures
. למחברים אין מה לגלות
Acknowledgements
אנו מודים למאיה וילסון ולאנאיקה לייני לקריאה הקריטית של כתב היד. עבודה זו נתמכה על ידי מלגת סר הנרי דייל במשותף על ידי האמון ברוך הבא ואת החברה המלכותית (200473/Z/16/Z).
Materials
| Name | Company | Catalog Number | Comments |
| Chemicals | |||
| Acid phenol | SIGMA | P4682 | |
| Agarose | VWR | A2114 | |
| Ammonium acetate | SIGMA | A1542 | |
| Chlorophorm | Fisher scientific | 10293850 | |
| DNase/RNase free water | Fischer Scientific | 10526945 | |
| DNAzyme | Integrated DNA Technology | Custom oligo DNA | |
| EDTA | SIGMA | E9884 | |
| Ethanol Absolute | Fisher scientific | 10437341 | |
| Formaldehyde | Sigma | F8775 | |
| Formamide | sigma | F9037 | |
| Galactose | SIGMA | G0750 | |
| Gel Loading Dye | Thermo Fisher Scientific | R0611 | |
| Glucose | SIGMA | G7021 | |
| Glycogen | Thermo Fisher Scientific | R0561 | |
| HEPES | SIGMA | H3375 | |
| Isoamyl | SIGMA | W205702 | |
| KCl | SIGMA | P9333 | |
| MgCl2 | SIGMA | M8266 | |
| MnCl2 | SIGMA | 244589 | |
| MOPS | SIGMA | M1254 | |
| NaCl | SIGMA | S7653 | |
| Oxoid Peptone Bacteriological | Thermo Fisher Scientific | LP0037 | |
| Oxoid Yeast Extract Powder | Thermo Fisher Scientific | LP0021 | |
| RiboLock RNase Inhibitor (40 U/µL) | Thermo Fisher Scientific | EO0382 | |
| SDS | SIGMA | 74255 | |
| Sodium acetate trihydrate | SIGMA | S8625 | |
| SYBR Safe DNA Gel Stain | Thermo Fisher Scientific | S33102 | |
| Tris base | SIGMA | TRIS-RO | |
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| 1.5 mL microtubes | Sarstedt | ||
| 152VR5C01M -80°C freezer | Thermo Fisher Scientific | ||
| 250 mL Erlenmeyer flasks | Cole-Parmer | ||
| 50 mL conical tubes | Sarstedt | ||
| Combicup VX200 vortex | Appleton Woods | ||
| DS-11 microspectrophotometer | Denovix | ||
| Electrophoresis chamber (20 cm tray) | SIGMA | ||
| FiveEasy F20 pH meter | Appleton Woods | ||
| Gel documentation system | Syngene | ||
| Heraeus Fresco 21 micro centrifuge | Fisher Scientific | ||
| Megafuge 8R centrifuge with rotator suitable for 50 mL conical tubes | Fisher Scientific | ||
| Mini Fuge Plus mini centrifuge | Starlab | ||
| Mixer HC thermal block | Starlab | ||
| OLS26 Shaking Water Bath | Grant | ||
| PowerPac power supplier | BioRad |
References
- Dieci, G., Preti, M., Montanini, B. Eukaryotic snoRNAs: a paradigm for gene expression flexibility. Genomics. 94 (2), 83-88 (2009).
- Watkins, N. J., Bohnsack, M. T. The box C/D and H/ACA snoRNPs: key players in the modification, processing and the dynamic folding of ribosomal RNA. Wiley Interdisciplinary Review RNA. 3 (3), 397-414 (2012).
- Kufel, J., Grzechnik, P. Small Nucleolar RNAs Tell a Different Tale. Trends in Genetics. , (2018).
- Li, T., Zhou, X., Wang, X., Zhu, D., Zhang, Y. Identification and characterization of human snoRNA core promoters. Genomics. 96 (1), 50-56 (2010).
- Jorjani, H., et al. An updated human snoRNAome. Nucleic Acids Research. 44 (11), 5068-5082 (2016).
- Hubbard, T. J., et al. Ensembl 2009. Nucleic Acids Research. 37 (Database issue), D690-D697 (2009).
- Makarova, J. A., Kramerov, D. A. SNOntology: Myriads of novel snoRNAs or just a mirage?. BMC Genomics. 12, 543 (2011).
- Yu, Y. T., Shu, M. D., Steitz, J. A. A new method for detecting sites of 2'-O-methylation in RNA molecules. RNA. 3 (3), 324-331 (1997).
- Decatur, W. A., Liang, X. H., Piekna-Przybylska, D., Fournier, M. J. Identifying effects of snoRNA-guided modifications on the synthesis and function of the yeast ribosome. Methods in Enzymology. 425, 283-316 (2007).
- Dong, Z. W., et al. RTL-P: a sensitive approach for detecting sites of 2'-O-methylation in RNA molecules. Nucleic Acids Research. 40 (20), e157 (2012).
- Birkedal, U., et al. Profiling of ribose methylations in RNA by high-throughput sequencing. Angewandte Chemie International Edition, England. 54 (2), 451-455 (2015).
- Buchhaupt, M., Peifer, C., Entian, K. D. Analysis of 2'-O-methylated nucleosides and pseudouridines in ribosomal RNAs using DNAzymes. Analytical Biochemistry. 361 (1), 102-108 (2007).
- Grzechnik, P., et al. Nuclear fate of yeast snoRNA is determined by co-transcriptional Rnt1 cleavage. Nature Communication. 9 (1), 1783 (2018).
- Piekna-Przybylska, D., Decatur, W. A., Fournier, M. J. New bioinformatic tools for analysis of nucleotide modifications in eukaryotic rRNA. RNA. 13 (3), 305-312 (2007).
- Schmitt, M. E., Brown, T. A., Trumpower, B. L. A rapid and simple method for preparation of RNA from Saccharomyces cerevisiae. Nucleic Acids Research. 18 (10), 3091-3092 (1990).
- Maden, B. E. Mapping 2'-O-methyl groups in ribosomal RNA. Methods. 25 (3), 374-382 (2001).
- Santoro, S. W., Joyce, G. F. A general purpose RNA-cleaving DNA enzyme. Proceedings of the National Academy of Science, U. S. A. 94 (9), 4262-4266 (1997).
- Hengesbach, M., Meusburger, M., Lyko, F., Helm, M. Use of DNAzymes for site-specific analysis of ribonucleotide modifications. RNA. 14 (1), 180-187 (2008).
- Sednev, M. V., et al. N(6) -Methyladenosine-Sensitive RNA-Cleaving Deoxyribozymes. Angewandte Chemie International Edition, England. 57 (6), 15117-15121 (2018).
- Aschenbrenner, J., Marx, A. Direct and site-specific quantification of RNA 2'-O-methylation by PCR with an engineered DNA polymerase. Nucleic Acids Research. 44 (8), 3495-3502 (2016).
- Lee, K. W., Bogenhagen, D. F. Assignment of 2'-O-methyltransferases to modification sites on the mammalian mitochondrial large subunit 16 S ribosomal RNA (rRNA). Journal of Biological Chemistry. 289 (36), 24936-24942 (2014).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved