
Method Article
הקרנה אסטרטגית ואפיון של הקומפלקס החזותי של החלבון GPCR-mini-G של התגבשות מוצלחת
In This Article
Summary
דו ח זה מתאר הקרנה של דטרגנטים שונים להכנת GPCR החזותית, הרודופסין ומתחם המורכב שלו עם mini-Go. השיטות הביוכימי המאפיינים את איכות הקומפלקס בשלבים שונים במהלך הטיהור מומחש. פרוטוקול זה ניתן להכליל למערכות חלבון ממברנה אחרים עבור המחקרים העתידיים שלהם במבנה.
Abstract
המפתח לקביעת מבני גביש של מכלולי חלבון ממברנה הוא איכות המדגם לפני התגבשות. בפרט, בחירת חומרי הניקוי היא קריטית, כי היא משפיעה על יציבות ומונוטוני של המתחם. לאחרונה קבענו את מבנה הגביש של מצב פעיל של העין הרודוסין משולב לחלבון G מהונדס, mini-Go, ב 3.1 Å פתרון. כאן, אנו מפרטים את ההליך למיטוב הכנתמורכבות הרודוסין-מיני-ג'י. מדינה אפלה הייתה מוכנה בחומרי ניקוי קלאסיים וneopentyl גליקול (NPG), ואחריו מבנה מורכב עם מיני-G מתחת לחשיפה לאור. היציבות של הרודופסין הוערך על ידי הספקטרוסקופיית אולטרה-סגולה (UV-VIS), העוקבת אחר החוקה לתוך הרודופסין של ליגסין רגיש לאור, 9-cis ברשתית. השימוש האוטומטי בגודל כרומטוגרפיה (SEC) שימש כדי לאפיין אתהמורכבות של הרודופסין ומתחם המידוסין-מיני-ג'י. SDS-פוליאקרילאמיד אלקטרופורזה (SDS-PAGE) אישר את היווצרות הקומפלקס על ידי זיהוי של 1:1 היחס הטוחנת בין הרודופסין ו-mini-Go לאחר להכתים את ג'ל עם coomassie כחול. לאחר אימות כל הנתונים האנליטיים הללו, הצלחנו לבטל את הניקוי המתאים ולהמשיך בתכשיר הניקוי הטוב ביותר להכנה והתגבשות בקנה מידה גדול. בעיה נוספת עלתה בטרוגניות של N-הגליסיזילציה. באופן הטרוליאני-ביטוי בהדופסין נצפה ב-SDS-PAGE כדי להיות בעלי שני אוכלוסיות שונות משנת N-גליסיתיים, שכנראה היו מעכבת את crystallogenesis. לכן נבדקו אנזימים שונים של דגליקוזילציה, ואנדוגליסידאז F1 (EndoF1) הפיקה את הרודופסין עם מינים בודדים של N-הגליסיציה. עם צינור אסטרטגי זה לאפיון איכות החלבון, הכנת מתחם הרודוסין-mini-Go היה מיטבי כדי לספק את מבנה הגביש. זה היה רק מבנה הגביש השלישי של מורכב GPCR – G חלבון. גישה זו יכולה גם להיות כללית עבור חלבונים ממברנה אחרים מתחמי שלהם כדי להקל על הכנה לדוגמה ונחישות מבנה.
Introduction
קביעת מבנים גביש של חלבונים הממברנה והתסביכים שלהם תמיד היה מאתגר בשל קשיים בהשגת קריסטלים היטב משחק. בניגוד לחלבונים מסיסים, חלבונים ממברנות אינטגרליים מהווים גרעין הידרופובי המתפרס על קרום התא. כדי להסיר חלבונים ממברנה מקרום התא לתוך מאגר מימית, חומרי ניקוי יש להשתמש כדי ליצור מיכל אבקת חלבון, ובכך להחליף את השומנים סביב הליבה הידרופובי של חלבונים ממברנה. יציבות, פעילות ויושרה של חלבונים ממברנה תלויים ישירות על תכונות כימיות ומבניות של אבקת הניקוי1, ואת התכונות של ניקוי גם לקבוע את הגודל של micelle. מיכל ניקוי גדול עשוי להיסגר את משטחי ההידרופיפילית של חלבון ממברנה קטן, ובכך מניעת התגבשות בשל חוסר המגע הגבישי בעת שימוש בשיטת דיפוזיה אדים. מיכל אבקת כביסה קטן הוא יתרון עבור קריסטלוגרפיה, אבל שרשרת שקלים קצרים בדרך כלל ולכן להוביל לחוסר יציבות וצבירה של חלבון הממברנה. לכן, לפני התגבשות, הליך נוסף ניקוי ההקרנה היא הכרחית, בדרך כלל מיקוד ניקוי קצר שעדיין לשמור על יציבות החלבון.
G חלבון מצמידים קולטנים (GPCRs) הם חלבונים ממברנה אינטגרלי המכיל שבעה טרנסקרום a-helices. GPCRs קיים בשני מדינות עיקריות, או מדינה לא פעילה התייצב על ידי אגוניסטים הופכי או אנטוניסטים, או מדינה פעילה המאוגדת אגוניסט והתייצב על-ידי חלבון G, למרות שסביר להניח כי ריבוי מדינות קיימות בין שני הקיצוניויות האלה. קביעת מבנה של GPCRs בתחילה התמקדה מדינות לא פעיל הקשורים אגוניסטים הופכי ו האנטוניסטים בשל היציבות הגבוהה שלהם מאשר מדינות פעיל2. כאשר GPCRs מופעלים על גבי כריכת אגוניסט, הקולטנים הם דינמיים מאוד, וצורות שסוע מארעיות על פני הפנים cytoplasmic של קולטן עבור צימוד חלבון G. הוא חשב כי דינמיות זו היא הסיבה מדוע אגוניסט מאוגד GPCRs הם לעתים קרובות יותר יציבה מאשר מצב לא פעיל. לכן, זה הופך להיות חיוני על המסך עבור חומרי ניקוי המתאימים למצב הקונמובי של הקולטן תחת המחקר, כי סביר להניח כי הניקוי מתון יהיה צורך ללמוד מדינה פעילה בהשוואה למצב לא פעיל.
בדו ח זה, אנו משתמשים ב-gpcr החזותית, בשור הרודוסין3, והקומפלקס שלועם חלבון mini -G4,5 לניסויים בסינון חומרי ניקוי, המייצגים את המצב הלא-פעיל והמצב הפעיל, בהתאמה. הקרנת חומרי הניקוי התמקדה בחומרי הניקוי הקלאסיים ובדטרגנטים הneopentyl גליקול (NPG). בהקשר זה, כביסה קלאסית בנויה מקבוצה של ראש סוכר ושרשרת אלקלאני, בעוד שחומרי הניקוי מסוג npg מכילים שני חומרי ניקוי קלאסיים זהים אשר מותתים על-ידי פחמן מקווארי בממשק בין סוכרים ושרשראות האלאלאני6,7,8.
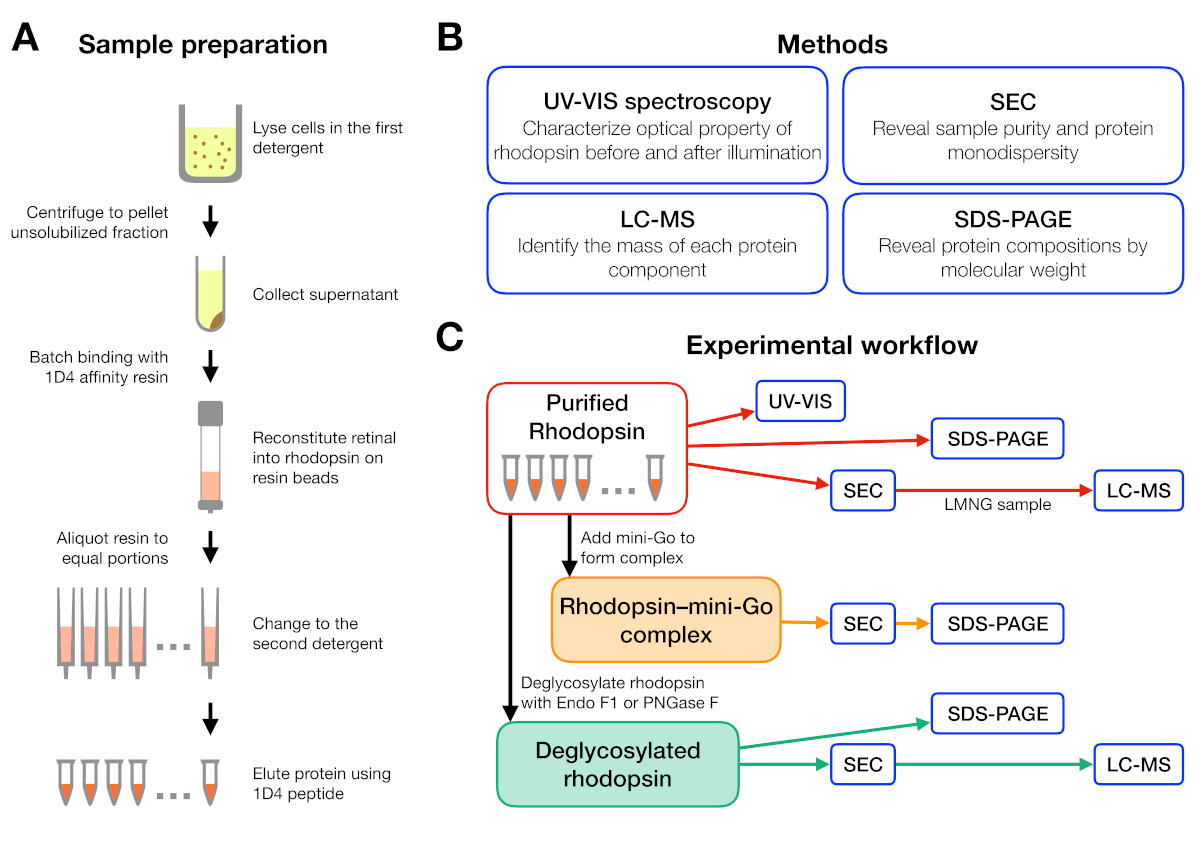
זרימת עבודה ניסיונית תוכננה החל מטיהור של הרודופסין בדטרגנטים שונים, ולאחריה היווצרותמורכבות הרודוסין-מיני-ג'י ומסתיימת באפיון הקומפלקס באמצעות מספר שיטות (איור 1). עבור המצב הלא פעיל של הרודוסין, החוקה של האור רגיש ליגו 9-cis ברשתית היה מפוקח על ידי אולטרה סגול-גלוי (UV-VIS) ספקטרוסקופיית. הספקטרום חושף את המצב הגופני של הרשתית והוא מעיד על סביבתו בכיס המחייב הרשתית של הרודופסין. כרומטוגרפיה של הוצאה לפועל (SEC) הועסק על מנת להעריך מונוטוני של הרודופסין מטוהרים, כמו גם היווצרות שלמתחם המידוסין-מיני-ג'י. כמו שניה מבדיל בין מולקולות חלבונים לפי גודלם וצורתם, אוכלוסיית חלבון מצטברת יכולה להיות מזוהה כאשר הם בתוך עוצמת הריק. כדי לאשר מבנה מורכב, שברים מ-SEC העריכו על ידי נתרן dodecyl סולפט-פוליאקרילמיד ג'ל אלקטרופורזה (SDS-PAGE) כדי לאשר את הנוכחות של שני הרודוסין ו-mini-Go.
גורם נוסף שצריך להיחשב הוא שינויים פוסט-טרנסלtional (PTM) על החלבונים הממברנה. Ptm כגון N-הגליקוציה הם נצפו לעתים קרובות על חלבונים ממברנה איקריוטית המיוצרים מערכות ביטוי של תאי חרקים. זן N-גליקוזילציה מוגבלת של הכליה האדם העובריים 293 (HEK293) תאים פותחה על ידי מחיקה של קידוד הגנים N-acetylglucosaminyltransferase I (GnTI), וכתוצאה מכך הומוגנית N-הגליקוזילציה על-ידי GlcNAc2Man5 באתר הקונצנזוס Asn-X-Ser/שלושה. למרות שניתן למנוע את N-הגליסילציה על-ידי שינוי שאריות חומצות אמינו באתר הקונצנזוס, זה עשוי גם לשנות את התפקוד של החלבון או את היעילות של קיפול. בשור הרודוסין, מוטציה של השאריות N-גליסיטיות Asn15 מוביל לקיפול שגוי ולהפחית הפעלה של חלבון G9,10. הרודוסין המשמשת בדו ח זה באה לידי ביטוי בקו ה-HEK 293 GnTI לקוי. עם זאת, SDS-PAGE הראה נוכחות של שני מינים של הרודופסין. טרוגניות זה יכול למנוע היווצרות גביש ולכן deglycosylation באמצעות פפטיד-N-גליסידאז F (PNGase F) ו אנדוגליסידאז F1 (אנדו F1) נבדק. המוצר הדגליסילי התאפיין בספקטרומטר מוצרים (LC-MS) ובעזרת ספקטרומטריה לזיהוי רמת הגליקוציה וההומוגניות שלה.
Protocol
הערה: פרוטוקול זה עבור הקרנת ניקוי מפורט עבור 30 גרם של HEK293 תא הגלולה כחומר התחלתי.
1. חומרים, כימיקלים וריאגנטים
הערה: כל הפתרונות מוכנים באמצעות ריאגנטים אנליטי כיתה ומים אולטרה-טהורות, אשר מטוהר ממים מיוטים ל18.2 הגיע לMΩ ∙ ס"מ על 25 ° c.
- פתרונות מניות מאגר
- הכינו מלוחים באגירה 10x פוספט (10x PBS).
- הכן מאגר hepes: 1 M, טיטרציה כדי pH 7.5 עם naoh.
- הכינו 5 מ ל
- הכינו 2 מטרים מ-2.
הערה: כל הפתרונות מניות עוברים באמצעות מסנן 0.22 יקרומטר כדי לשמור על עקרות שלהם.
- חומרי ניקוי לתמיסה
- להכין dodecyl maltoside (DDM), 10% (w/v).
- הכינו decyl maltoside (DM), 10% (w/v).
- הכינו 6-ציקלוקסאיל-הקסאיל מלטוסייד (Cymal-6), 10% (w/v).
- הכינו 5-ציקלוקסיל-פנטאיל מלטוסייד (Cymal-5), 10% (w/v).
- הכינו את החלק של nonyl msm (C9G), 10% (w/v).
- הכינו את לאוראל מלקטוז neopentyl גליקול, 5% (w/v).
- הכינו את decyl מאלז neopentyl גליקול (DMNG), 10% (w/v).
- הכינו cymal-6 neopentyl גליקול (C6NG), 10% (w/v).
- הכינו cymal-5 neopentyl גליקול (C5NG), 10% (w/v).
- הכינו את הגלוקוז neopentyl גליקול (OGNG), 10% (w/v).
הערה: עבור 10% אבקת מלאי ניקוי, לפזר 1 גרם של אבקת כביסה במים באולטרטהורים עם נדנדה עדינה, ולאחר מכן להתאים את הנפח הסופי ל 10 mL. מניות ניקוי הפתרון יש לשמור ב-20 ° c עבור אחסון לטווח ארוך ועל קרח בזמן העבודה.
התראה: מומלץ בדרך כלל לאחסן מקפיא בבקבוקים ב-20 ° c. יש לחמם את הבקבוקים המכילים אבקת ניקוי לטמפרטורת החדר לפני הפתיחה. אבקת כביסה היא היגרוסקופי, ולכן הטמפרטורה מונעת היווצרות עיבוי כי יהיה להרטיב את הניקוי.
- כימיקלים וריאגנטים אחרים
- הכינו שרף חיסוני 1D4:10 מ ל של 50%.
הערה: בעלת הזיקה החיסונית של 1D4 הם החרוזים הקשורים בנוגדן Rho1D4 המונבטיים, אשר מאגד את 9 חומצות האמינו האחרונות של הרדוסין TETSQ, כאפיאגית. הזיקה החיסונית של 1D4 פועלת כחומר טיהור אהדה כדי ללכוד חלבונים המכילים רצף של C-terminal 1D4. חומר טיהור זה יכול להיות מוכן9,11 או לרכוש. - להכין 9-cis פתרון ברשתית: 1 מ"מ, מומס 100% אתנול.
הערה: מניעת חשיפה באור לרשתית במהלך ההכנה והאחסון. - הכינו פפטיד 1D4 (רצף TETSQ,): 800 μM, מומס במים.
- הכינו שרף חיסוני 1D4:10 מ ל של 50%.
- אגרי
הערה: כל המאגרים מעורבבים מפתרונות המניה לריכוז הרצוי. כל המאגרים מצוננים ל -4 ° c לפני השימוש.- הכנת מאגר A: PBS, 0.04% DDM.
- הכנת מאגר B: 20 מ"מ HEPES pH 7.5, 150 מ"מ הנאל, 0.04% DDM.
- הכנת מאגר C: 20 מ"מ HEPES pH 7.5, 150 מ"מ וכלי ניקוי בריכוז העבודה שלהם המפורטים בטבלה 1.
- הכנת מאגר D: 20 מ"מ HEPES pH 7.5, 150 מ"מ הנאל.
- הכנת מאגר הימנעות: 20 מ"מ HEPES pH 7.5, 150 מ"מ הייקל, 80 μM 1D4 פפטיד, ואבקת כביסה בריכוז העבודה שלהם.
- הכנת מאגר SEC: 20 מ"מ HEPES pH 7.5, 150 מ"מ הנאל, 0.025% DDM; filtrated באמצעות מסנן 0.22 יקרומטר.
- ממס עבור LC-MS
- הכנת הממס A: acetonitrile המכיל 0.1% החומצה פורמית.
- הכן ממס B: מים באולטרטהור המכיל 0.1% החומצה פורמית.
- להכין את הממס C: iso-propanol.
2. מסיסות קרום התא והפקת חלבון
- הפשרת 30 גרם של HEK293 gnti- תא גלולה המבטא את מוטציה השור N2C/M257Y/D282C3,9 לטמפרטורת החדר, להוסיף 120 mL של 1x PBS מאגר המכיל את הקוקטייל פרוטאז מעכבי המגון באמצעות מכשיר הומוגנידצר או הומוגניצר חשמלי (13,000 rpm עבור 30 s). לאסוף את הבולם התאים הומוגניים בגביע ולהתאים את עוצמת הקול ל 150 mL.
הערה: 30 גרם של גלולה תא הוא שווה ל 3 L של תרבות התא ב 2 x 106 תא/mL צפיפות. - הוסף בעדינות 10% DDM לתאים הומוגניים כדי להעניק ריכוז סופי של 1.25%. . מערבבים קרח במשך 1 שעות
- צנטריפוגה את התא ליפוסט ב 4 ° צ' ו 150,000 x g עבור 45 דקות כדי להסיר את הפסולת בלתי מסיסות.
- העבר את supernatant לבקבוק 500 mL ולהוסיף 10 מ ל של הזיקה החיסונית 1D4 שרף (50% לשתות). בעדינות לערבב את התאים מסיסות ו שרף עבור 4 h או לילה ב 4 ° c.
- טען את תערובת הליפוסט/שרף לטור פתוח כדי לאסוף את שרף.
- שטוף את השרף עם 10 אמצעי אחסון של עמודות (CV) של מאגר הכביסה A.
הערה: אמצעי האחסון של העמודה הוא הנפח של הארוז (100%) שרף agarose בשימוש. במקרה זה, 1 קורות חיים הוא 5 מ ל. - השהה מחדש את השרף עם 2 קורות חיים של מאגר A.
התראה: משלב 2.8 ואילך, שלבים שיש לבצע במצב אור אדום עמום מתויג ב-"[Dark]" בתחילת התיאור. - כהה הוסף 9-cis ברשתית אל שרף מחדש את הריכוז הסופי של 50 μM. מערבבים בעדינות ב -4 ° c עבור 4-16 h בחשיכה.
הערה: זמן דגירה קצר עלול להוביל החוקה הושלמה של רשתית. - כהה הסרת זרימה דרך העמודה. לשטוף שרף עם 20 קורות חיים מאגר A, ואחריו 15 קורות חיים מאגר B.
- כהה להשעות את שרף ב 2 קורות חיים מאגר B, ולאחר מכן לחלק את ההשעיה שרף באופן שווה 10 10-mL עמודות לסילוק.
- כהה הסר את הזרימה דרך העמודה ולאחר מכן השהה מחדש את השרף ב-1 mL מאגר C. דגירה עבור 1 h ב 4 ° c.
- כהה חזור על שלב 2.11.
- כהה הסר את הזרימה דרך העמודה ולאחר מכן השהה מחדש את השרף במאגר ההתראות של 0.8 mL עבור כל עמודה. מערבבים בעדינות עבור 2 שעות.
- כהה לאסוף הימנעות מהעמודה לתוך שפופרת 2 מ ל.
- כהה השהה מחדש את השרף ב-0.7 mL של מאגר האלוטור עבור כל עמודה. ערבב בעדינות עבור 1 h.
- כהה לאסוף הימנעות מהעמודה לתוך אותה צינורית.
3. ספקטרוסקופיית UV
- הכן את הספקטרוסקופיה לכיסוי טווח המדידה של 250-650 ננומטר. הקלט את תוכנית הבסיס באמצעות מים או מאגר הימנעות.
- כהה העמיסו את החלבון החומק לקובט הקוורץ. מדדו את הספקטרום של דגימת החלבון.
- כהה להאיר את החלבון ישירות קובט עבור 2 דקות עם אור עבר דרך מסנן 495 ננומטר ארוכת לעבור.
- למדוד את הספקטרום של המדגם המואר.
- בצע את אותה מדידה עבור כל הדגימות חלבון מטוהרים ב 9 דטרגנטים אחרים, הן מדינות כהה ומוארים.
- התווה את העקומות (ספיגת לעומת אורך הגל) בתרשים פיזור של X-Y.
4. הכללה אוטומטית בגודל כרומטוגרפיה של הרודופסיןומתחם הרודוסין-מיני-גרם
- כהה להתרכז חלבון 100 μL על ידי צנטריפוגה באמצעות מרכז ספין עם משקל מולקולרי לגזור (MWCO) של 30 kDa ב 4 ° c. ניתן לדלל דגימות יתר מרוכזת באמצעות הזרימה דרך הרכז או מאגר C. כדי לקבוע ריכוז דגימת חלבון, למדוד ספיגת ב280 ננומטר באמצעות ספקטרוסקופיה.
הערה: משלב 4.2 ואילך, הניסוי אינו דורש סביבה אפלה, ולכן ניתן להכין דגימות באור רגיל. - הכינו 100 μL הרודופסין ב 0.7 מ"ג/mL עבור כל תנאי ניקוי.
- הכינו 100 μl של הרודופסין (0.7 mg/ml) ו-mini-G4,12 (0.2 mg/ml) תערובת עבור כל מצב ניקוי. הוסף את התערובת עם 1 מ"מ MgCl2. להאיר את התערובת עם אור מ 495 ננומטר ארוך לעבור מסנן ו דגירה עבור 30 דקות.
- הר a 24 לסדר סינון ג'ל לטור עם טווח משבר של 10-600 kDa של חלבון כדורי על מטהר כרומטוגרפיה נוזלית. העמודה מעורפלת באמצעות מאגר SEC.
הערה: מטהר כרומטוגרפיה נוזלי מצויד בדוגם אוטומטי, גלאי אורך גל מרובה ואספן שבר. - העבר את הדגימות לבקבוקונים באמצעות דוגם הצלילים האוטומטיים והציבו אותם במגש המדגם. תכנת קובץ שיטה כדי להפוך את הרצפים הרציפים של שניה לאוטומטיים עבור כל דוגמה, כאשר טעינת הדגם האוטומטי מטעינה את 77 μL של המדגם לעמודה, והמיטהר משחררי 24 mL של מאגר SEC בקצב זרימה של 0.5 mL/min לכל הפעלה. להקליט את ספיגת ב 280 ננומטר ו 380 nm.
- אספו את שברי השיא של הרודופסין והרודוסין-מיני-ג'י מורכבים בנפח השמירה סביב 12.9 mL.
- לנתח את הרודופסין השמאלית דגימות משלב 4.2 ואת שברי השיא של מורכבות rhosin-מיני-G ב-4-12% SDS-denaturing מעבר ג'לים עם כתמים כחולים coomassie.
- התוויית הנתונים כרומטוגרמה (A280 או a380 לעומת אמצעי אחסון שמירה).
5. הדיגליסיציה ומחקר LC-MS
- למחקר LC-MS, השתמש רק במדגם הרודוסין מטוהר בחומר הניקוי של LMNG.
- הכינו תערובת מ200 μL של הרודופסין ב-1 מ"ג/mL ו-PNGase F13 ב 0.01 מ"ג/ml. מערבבים היטב הדגירה ב 4 ° c לילה.
- הכינו תערובת מ200 μL של הרודופסין ב-1 מ"ג/mL ואנדו F113 ב 0.01 מ"ג/ml. מערבבים היטב הדגירה ב 4 ° c לילה.
- לנתח את תוצאת העיכול על ידי SDS-PAGE ו Coomassie כחול כתמים.
- להתרכז ואנדו F1-מטופל מדגימות ובכפוף טיהור שניה במאגר D.
הערה: זה להכין את המדגם עם כמות מינימלית של חומרי ניקוי עבור LC-MS לימוד. מאגר D אינו מכיל חומרי ניקוי, אלא בגלל שיעור ההאטה של ה-LMNG מחלבון ממברנה14, הרודופסין לא יצטבר. - לאסוף את שבר השיא בנפח השמירה סביב 12.9 mL. להתרכז 1 מ"ג/mL באמצעות מרכז ספין (MWCO 30 kDa).
- הכנס 10 μg של החלבון 200 לתוך העמודה סי18-AQ של המבנה והעמודה באמצעות שיטת מעבר הצבע הליניארית עם הרכב הממס וההגדרות המפורטות בטבלה 2. הזרימה מפוצלת ל -25% ספקטרומטר מסה ו-75% עבור זיהוי UV.
תוצאות
זרימת העבודה הניסיונית להכנה וניתוח לדוגמה מסוכמת באיור 1. באמצעות עמודות פתוחות לטיהור אהדה בקנה מידה קטן אפשרה לנו להכין דגימות בתנאי ניקוי שונים רבים במקביל (איור 1א). כזה מערכת טיהור בקנה מידה קטן הניב חלבון מספיק עבור ניתוחים נוספים באמצעות ספקטרוסקופיית UV-VIS, SEC ו-SDS-עמוד (איור 1B-C).
ספקטרוסקופיית UV-VIS חשף את היציבות הרודופסין
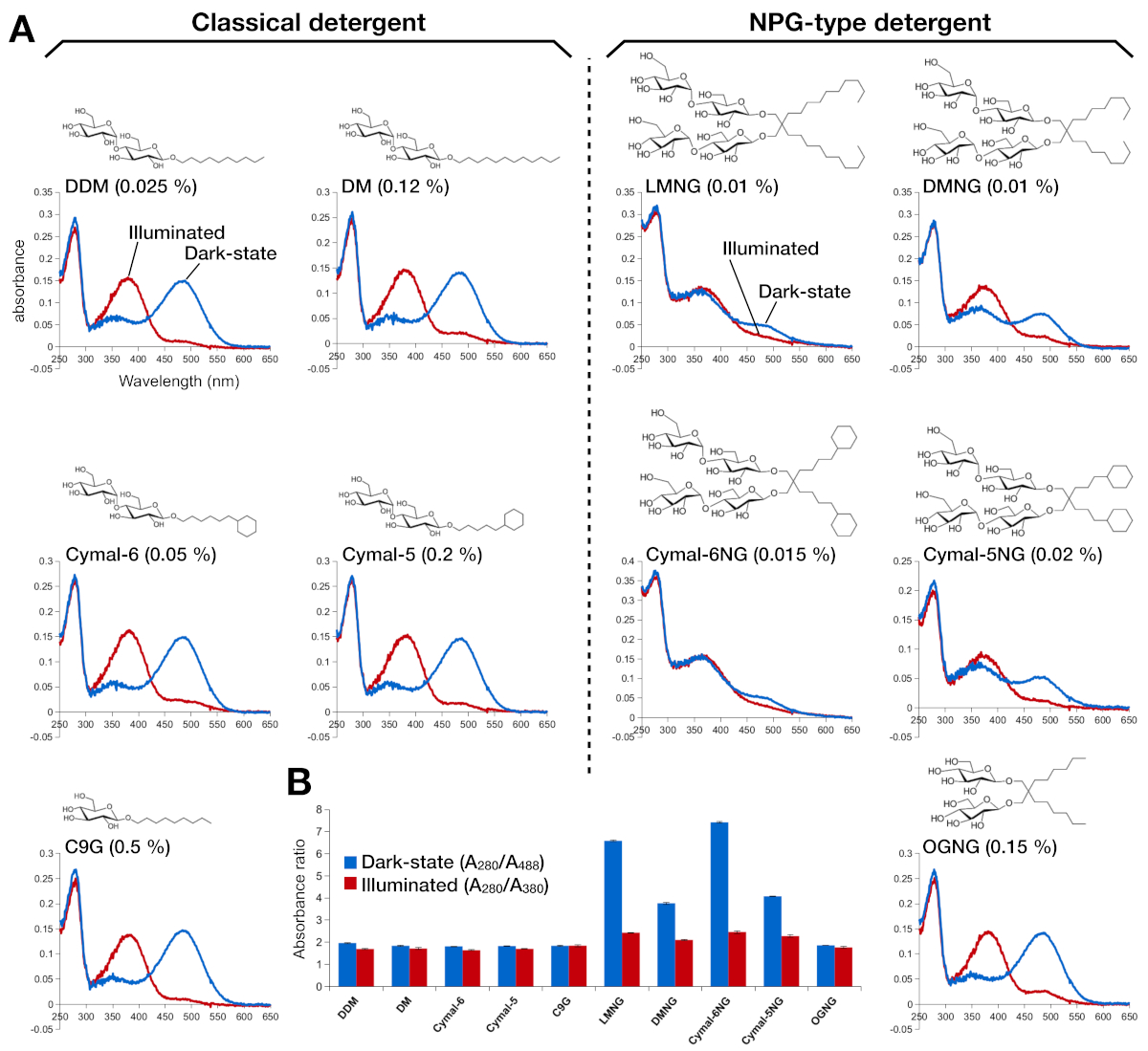
היציבות של הרידופסין הרשתית הייתה מוערך על ידי ספיגת האופטי (איור 2). במצב אפל, 9-cis ברשתית מקושרת בעקשנות Lys296 כבסיס שיף הפרו. לאחר ההארה, הרשתית של 9 העמים הוא איזוריאד אל כל טרנס isoform ואת הקישור בסיס שיף הוא deprotonated. The protonated 9-cis ברשתית נותן שיא הקליטה ב 488 nm, בעוד ברשתית deprotonated כל טרנס יש שיא ב 380 nm. ה-UV-VIS ספקטרום של הרודופסין ב DDM הראה את ספיגת אופייני של 9-cis הקשורות ברשתית מאוגד ואור המופעל, שם שינוי כחול של 108 ננומטר עם בערך אותו צפיפות אופטית נצפתה בבירור (איור 2, הלוח השמאלי העליון). כאשר הרודופסין הוא יציבות, ולאחר מכן כיס הכריכה לשינויים ברשתית, התוצאתה דה-פרוטונציה רשתית ואולי דיסוציאציה. אם זה קורה, ואז הספקטרום מראה את התרומה מדה-פרוטונציה, כמו גם את הצורה החופשית של הרשתית15. לכן, קבענו את היעילות של החוקה הרשתית לתוך הרודופסין על ידי יחס ספיגת החלבון (280 nm) ואת הרשתית (488 nm עבור protonated 9-cis ברשתית, 380 nm עבור כל טרנס הטראנס ברשתית) (איור 2B). דגימות הרודוסין מטוהרים בדטרגנטים הקלאסית (DDM, DM, Cymal-6, Cymal-5, C9G) להראות את אותו פרופיל אופטי. עם זאת, הדגימות מטוהרים בחומרי הניקוי של NPG (LMNG, DMNG, Cymal-6NG, Cymal-5NG) מציגים פרופילים אופטיים הרומז לסביבה מחייבת תת-אופטימלית עבור הרשתית, למעט דגימת ה-OGNG, שנתנה את אותו פרופיל אופטי כמו דגימת DDM.
כרומטוגרפיה בעלת גודל הוצאה לדוגמה. הראתה דגימת טוהר וחלבון מונוטוני
SEC הוא כלי אנליטי יעיל וחזק להערכת דגימות חלבון במהלך ההכנה וההקרנה. הוא מאמת דגימת טוהר משלב הטיהור הקודם, כמו גם את מונוטוני של מולקולות החלבון. עבור הרודוסיןומורכבות מיני-G שלו, איכות המדגם היתה מתורגמת מעקומות ספיגה ב 280 ננומטר ו 380 ננומטר (איור 3א). עקבות 280 ננומטר הראו נוכחות של חלבון, ו 380 מעקב ננומטר הראו נוכחות של רשתית. כל איתות המופיע בכרך הריק (כ-8 מ ל בעת שימוש בעמודה זו) יוחסו לאגרגטים חלבונים. לפיכך, התוצאות הראו כי דגימות שהוכנו בחומרי הניקוי הקלאסיים היו במצב מונוטוני למעט C9G, שם הופיעו חלק מהצבירה. לעומת זאת, דגימות שהוכנו באמצעות חומרי הניקוי NPG-type הכילו אגרגטים רבים יותר מאשר המדגם C9G; הוביל את המבנה למערך המצטבר ביותר, אך פחות אגרגטים נצפו ב-DMNG ו-Cymal-5NG. היוצא מן הכלל היה OGNG, אשר הראה פרופיל דומה DDM. אגרגטים חלבון משחררי בנפח הריק גם היה תפוסת הרשתית עניים, כפי שמוצג על ידי280/a380 היחס שגדל בהשוואה לשיא בנפח השמירה של ~ 12.9 ML מתאים 135 kda. תכונה נוספת שבחנו הייתה ששני הרודופסין והרודוסין – מיני-ג'י המריאו סביב אותו נפח שמירה (איור 3ב'). זה לא מפתיע, כי המשקל המולקולרי לכאורה של הרודופסין מאוגד הניקוי היה 120 kDa ואת זה של הרודוסין – mini-G-o 144 kda. לפיכך, לא יכולנו לוודא שמבנה מורכב בלבד מנתוני ה-SEC, כך ש-SDS-PAGE שימש לניתוח נוסף של הדוגמה שניה מטוהרים.
שירותי SDS-שאושרו על ידי מבנה מורכב
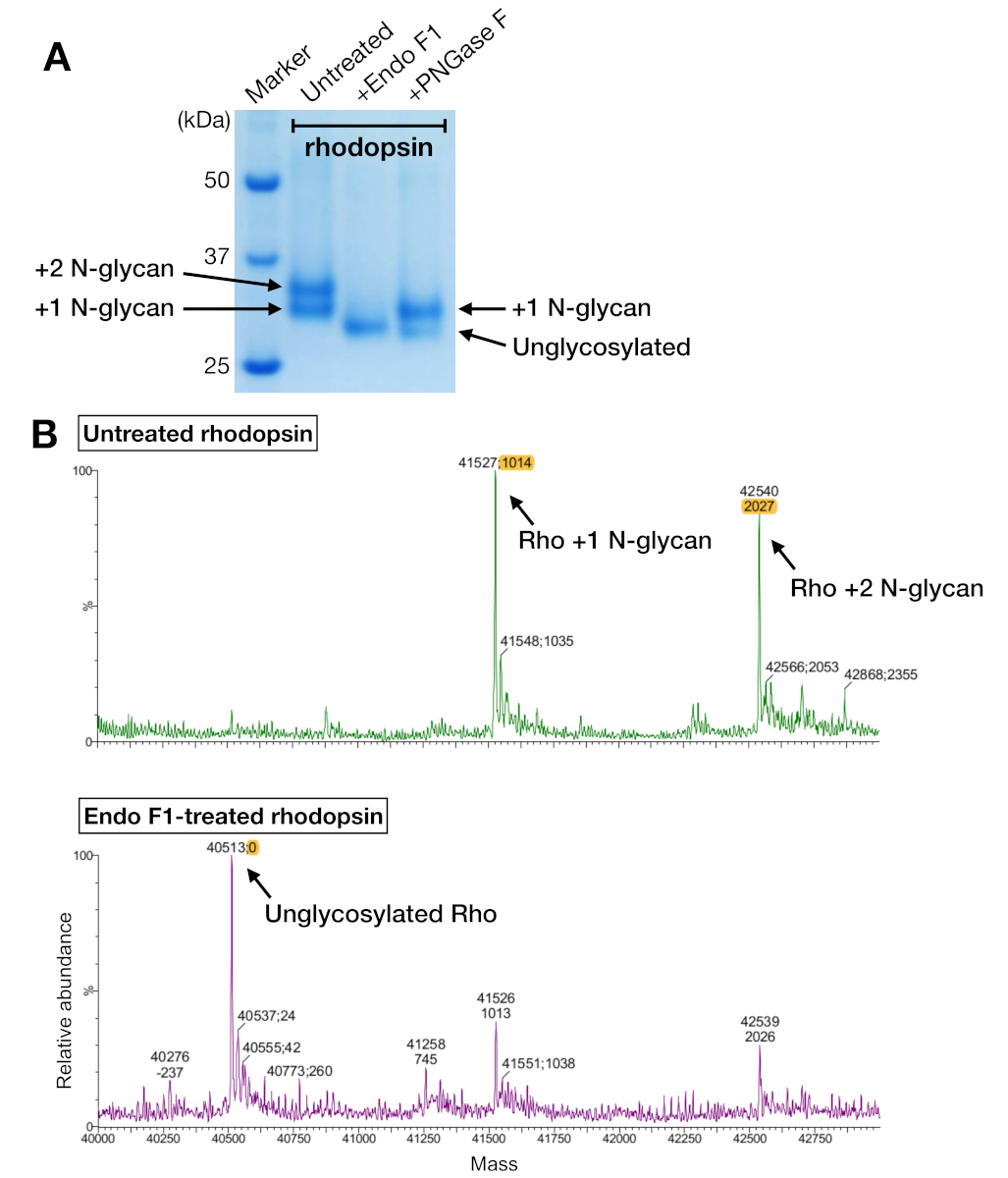
SDS-PAGE היא שיטה סטנדרטית לזיהוי רכיבי החלבון במדגם. מרוכז הרודופסין (לפני שניה טיהור) נותחו על ידי SDS-PAGE כדי לאשר את הטוהר שלה, והראה שתי להקות ליד 37 kDa ולהקה מרוח מעל 50 kDa (איור 4א). לשתי הלהקות הנמוכות יותר אישרו שיהיו מצבים שונים של N-גליסיציה. הלהקה שמעל 50 kDa היתה מתורגמת כ-oligomers מצטברים הנגרמת על ידי מאגר לדוגמה SDS-PAGE מאחר שאגרגטים אלה לא נצפו ב-SEC או כל שיטת זיהוי אחרת. כנתוני SEC לא היתה אפשרות לאשר מבנה מורכב, החלקים האלה של ה-SECשברו את שברי השימוש בדגימות של מיני-ג'י-פו באמצעות SDS-PAGE. ה-SDS-PAGE הראה להקות חלבונים של הרודופסיןומיני-ג'י בכל תנאי חומרי הניקוי, ומציע שהמתחם נוצר ללא קשר לבחירת חומרי הניקוי (איור 4ב').
ספקטרומטריה LC-MS מזהה את דפוס N-הגליסיציה באופדוסין
דגימות הרודוסין הן מטיהור האהדה והן מהניה הראו שתי להקות חלבון שהועברו עם משקל מולקולרי לכאורה של כ-37 kDa בג של SDS-PAGE, אשר לא ניתן היה להפריד ב-SEC בעת שימוש בעמודה 24-mL. דפוסים שונים של N-הגליקוזילציה על ההטרולילציה-מבוטאים מ-HEK 293 GnTI- תאים היה ההסבר הסביר ביותר. לכן, שני אנזימים, PNGase F ו אנדו F1, נבדקו על יכולתם לדמגליסיסין הרודופסין. מתוך הנתונים של SDS-PAGE, אנדו F1 הפחית את המשקל המולקולרי של שתי להקות החלבון לתוך מוצר אחד, בעוד העיכול PNGase F עדיין נתן שתי אוכלוסיות (איור 5א). הדגימות שטופלו ואנדו F1 מטופלים נותחו באמצעות ספקטרומטריה LC-MS כדי לזהות את ההמונים של מינים שונים. הנתונים הראו כי rhosin המיוצר HEK 293GnTI תאים הכילו או אחד או שניים N-גליקנים, עם הבדל במסה של 1014 ± 1 Da. אנדו F1-שטופלו הרודופסין לא הכיל כל N-גליקנים והיה הבדל המוני של 2027 ± 1 Da לעומת הרודופסין המכילים שתי N-גליקנים תוצאות אלה מתאימות עם העדר האנזים N-acetylglucosaminyltransferase I בתור התא המשמש לבטא rhodopsin, אשר תוצאות כל N-גליקנים שיש את המבנה GlcNAc2אדם5, (המוני 1014 Da).

איור 1: הכנה לדוגמא ואפיון לניסויים בסינון חומרי ניקוי. (א) הכנת דגימות הרודופסין בדטרגנטים שונים במהלך הטיהור. (ב) שיטות המשמשות בפרוטוקול: UV-VIS ספקטרוסקופיה, כרומטוגרפיה הדרה של גודל (שניות), SDS-עמוד ו-כרומטוגרפיה נוזלי-ספקטרומטר המסה (LC-MS). (ג) זרימת עבודה ניסיונית לאפיון הרודופסין, רודוסין-מיני-ג'י,והכפלת הגליקוסיציה של הרודופסין. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: ספקטרוסקופיית UV-VIS של הרודופסין. (א) אולטרה-סגולה של הרודופסין. הספקטרום של המדינה כהה, 9-cis מאוגדים ברשתית מופיעים בעקומות כחולות. לאחר תאורה, ברשתית 9-cis הוא deprotonated ו isomerizes לתוך הרשתית כל טרנס, ואת הספקטרום של הרודופסין מואר מוצגים כמו עקומות אדומות. המבנה הכימי של כל חומר ניקוי מוצג כהזחה. (ב) היחס של280/a488 (כחול בר) ו280/a380 (בר אדום) מתארת את היציבות של הרודופסין במדינה האפלה והמדינה הקלה, בהתאמה. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3: מתחם מיני-גרם המהווה כרומטוגרפיה פרופילים של הרודופסין והרודוסין-קומפלקס mini-Go מטוהר ב -10 חומרי ניקוי שונים. (א) הפאנל השמאלי מציג את הפרופילים של הזוג החדש של דגימות מטוהרים בחומרי ניקוי קלאסיים. הלוח הימני מייצג את פרופילי ה-SEC של הדגימות הנטהר בחומרי הניקוי של הסוג NPG. הפרופיל של החלבונים בסמן הרגיל מוצג כשכבת-על יחד עם דגימת DDM. הפרשנות של פרופילי שיא מוצג עבור DMNG, עם התרחיש האידיאלי (ללא אגרגטים) ראה עבור DDM, מיט, Cymal-6, Cymal-5 ו-OGNG. (ב) הפרופיל המוגדל של דגימת ה-ogng בנפח השמירה של 12-14 mL. כל הדגימות נותחו באמצעות Superdex200 להגדיל 10/300 GL עמודה. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 4: SDS-ניתוח עמודים של מתחם הרודוסין ורודוסין/מיני-גו. (א) דגימות רודוסין מטוהרים בדטרגנטים. הלהקה המרוחה מעל 50 kDa מיוחסת לאוסידונים הצבורים שנגרמה על ידי מאגר המדגם של SDS-PAGE. (ב) שניות מטוהרים של מורכבות הרודופסין/Mini-Go. הרודוסין עם 1 ו -2 N-גליקן ו-mini-Go מתוארים. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 5: זיהוי גליקוזילציה ברודופסין. (א) sds-ניתוח דפים של הרודוגליםהחטא באמצעות PNGase F ו אנדו F1. (ב) LC-MS ספקטרום של הרודוסין ללא (הפאנל העליון) עם deglycosylation על ידי אנדו F1 (הפאנל התחתון). להכנתהמבנה המיני-מיני-ג'י לצורך התגבשות, בחרנו באנדו F1 מעל PNGase F בגלל שאנדו f1 מסר מינים הומוגניות אחד של הרודופסין. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
| דטרגנט | עבודה בריכוז (%) | ריכוז מיצלה קריטי (%) |
| לפני החתונה | 0.025 | 0.0087 |
| מיט | 0.12 | 0.087 |
| Cymal-6 | 0.05 | 0.028 |
| Cymal-5 | 0.2 | 0.12 |
| C9G | 0.5 | 0.2 |
| מיכל הונג-יאנג | 0.01 | 0.001 |
| מיכל בגין | 0.01 | 0.0034 |
| Cymal-6NG | 0.015 | לא זמין; צריך להיות נמוך מ 0.056 |
| Cymal-5NG | 0.02 | 0.0056 |
| מיכל שוהם | 0.15 | 0.058 |
טבלה 1: ריכוזי חומרי ניקוי של מאגר C.
| זמן (מזערי) | ממיס A (%) | ממיס B (%) | ממיס C (%) | קצב זרימה (ml/דקה) |
| 0 | 0 | 95 | מיכל 5 | 0.5 |
| 1 | 0 | 95 | מיכל 5 | 0.5 |
| מיכל 5 | 20 | 75 | מיכל 5 | 0.6 |
| 25 | 85 | 10 | מיכל 5 | 0.6 |
| 26 | 90 | מיכל 5 | מיכל 5 | 0.6 |
| 30 | 90 | מיכל 5 | מיכל 5 | 0.6 |
טבלה 2: פרמטרי הימנעות מעמודות.
Discussion
ההצלחה התגבשות חלבונים מסתמך בחריפות על דגימת חלבון, בעיקר חלבונים ממברנה ומכלולי שלהם בשל הסיבוך הנגרם על ידי חומרי ניקוי. דו ח זה ממחיש הקרנת ניקוי והערכה של איכות דגימה עבור GPCR – מיני-G-מערכות חלבון איתות. מגוון של שיטות שימשו רבות לחקר המאפיין הביוכימי של חלבונים ממברנות, למשל, שיטת התרמויציבות באמצעות צבעי פלורסנט16,17, כריכה מחייבת לזהות היווצרות מורכב על ידי מדידת השינוי בטריפטופן שידור התדר18 או העברת אנרגיה התהודה עם ביוחיישנים19. עם זאת, הסביבות הכימיות המשמשות בשיטות אלה שונה לגמרי מאלה להכנת מדגם התגבשות, חלבונים הם בריכוז הנמוך ביותר של 1000 עבור מדידה מבוססי-פלואורסצנטית, או חלבונים מוטבעים bilayers ליפיד או במצב כביסה קבוע אחד. בפרוטוקול זה, השיטות המשמשות גם מתוקננת בהכנה לדוגמה בקנה מידה גדול לפני התגבשות. לכן, את הפרמטרים אופטימיזציה ניתן להעביר בקלות להכנה התגבשות בקנה מידה ללא הקרנה גדולה יותר ואופטימיזציה.
המטרה של פרוטוקול זה היא לייעל את הכנת GPCR יציבה והומוגנית-mini-G חלבון מורכב עבור אדים התגבשות דיפוזיה וקביעת מבנה על ידי קריסטלוגרפיה רנטגן. הפרוטוקול משלב סדרה של שיטות כדי להעריך בצורה איכותית את ההשפעה של חומרי ניקוי והדגליסיציה במהלך הכנתמתחם הרודוסין-מיני-ג'י. הרודוסין במצב לא פעיל ומדינה המופעלת באור מאוגד עם וללא התמרה בפפטיד הפכה להיות מטוהרים כאשר מטוהר בחומרי הניקוי של הדטרגנטים (C8G)20,21,22 ו C9G23,24. כאשרמתחם הרודוסין-מיני-ג'י מטוהר ב-C8G ו C9G לא התשואה קריסטלים (הנתונים אינם מוצגים), לאחר מכן בחנו מגוון רחב יותר של חומרי ניקוי אחרים באמצעות האסטרטגיה המתוארת (איור 1). על ידי לקיחת היתרון של רגישות האור של הרודוסין, אנחנו יכולים מאוד לעקוב אחר החוקה של הרשתית על אורכי גל אחר מ 280 ננומטר. ב-UV-VIS ספקטרוסקופיית ו-SEC, גילינו ברשתית ב או 380 ננומטר או 488 ננומטר. עם זאת, רוב החלבונים הממברנה אין כרומטופור כזה נוח לעקוב אחר הפונקציונליות במהלך הטיהור. אפשרויות אחרות יהיה ליצור ליגייט על-ידי הוספת כרומופור אור כרומרופאר או באמצעות רדיוligand-מחייב ומשמרת תרמית בחני25.
לרודוסין יש משקל מולקולרי של 40 kDa. בשל המסה של חומרי ניקוי זה נקשר, משקל מולקולרי לכאורה שלה על שניות הוא על 120 kDa. לכן אין מפתיע כי הכריכה של מיני-Go (24 kda) לא זוהה בקלות ב-SEC, כמו זה היה לחייב הבדלה של חלבונים עם המוני לכאורה של 120 kda ו 144 kda. לפיכך, השתמשו בניתוח של שברים של SEC על ידי SDS-PAGE כדי לאשר מדגם טוהר ומבנה מורכב. גם אם פרופילי SEC מציגים שינוי ברור בצורה מורכבת, עדיין מומלץ לבצע ניתוח של SDS-PAGE כדי לאשר את המבנה המורכב עם שותפי האיגוד הנכונים ולא מזהמים אחרים של חלבון מטוהרים.
הן הרודופסין והן מיני-ג'י היו מטוהרים בכמויות מיליגרם, שאפשרו להשתמש בזיהוי רגישות נמוך של התסביכים, כגון קליטת UV-VIS במהלך שניות וכתמים כחולים של sds-PAGE. כאשר דגימות מוגבלות, זיהוי רגיש יותר יש להשתמש, כגון מטהר LC מצויד גלאי פלואורסצנטית כדי לעקוב אחר אותות טריפטופן מן החלבון (280 ננומטר, 350 פליטת nm) ומכתים כסף עבור SDS-דף ג ' לים. גישה נוספת תהיה לנתיך חלבון פלורסנט, כגון חלבון פלואורסצנטי ירוק (GFP) על חלבון הריבית, אשר גם לאפשר איתור גם במהלך ביטוי החלבון26 אבל זה צריך להיות מוסר לפני התגבשות.
חיוני להבטיח כי חלבון מטוהר הוא גם חופשי של טרוגניות הנובעים משתנה לפטין. במקרה שמתואר כאן, שתי האוכלוסיות של הרודופסין שנצפו על SDS-דף היו מאופיינים כשיש אחד או שניים N-גליקנים. שינוי משתנה של חלבון עלול למנוע את היווצרות הקריסטלים היטב, ולכן אנו מדיגלינו את הרודופסין. האנדוגליזידאז אנדו F1 היה האפקט השכיח ביותר בדיקה וטיפול הוביל למין בודד של קולטן בלתי-גליזיאני, בעוד PNGase F הסיר באופן חלקי את הגליקנים על הרודוסין והביא לתערובת של הרודופסין באופן מלא, או עם אחד N-גליקן נשאר. הרודופסין ללא הטיפול בדגליסיקלז התגבשו בהצלחה3,27,28, ו-N-גליקן על הAsn15 ברודופסין חשוב ליצור קשר קריסטל במקרים אלה. במקרה של הרודוסין – mini-G,יש צורך להסיר את N-גליקנים על ידי אנדו F1 להשיג גבישים. אין כלל מתוקננת להסרת חלבונים מאוחרים של ריבית לפני התגבשות, אבל הסרת של הטרודוגני לפטין צריך להיחשב כאשר חלבונים להיכשל להתגבש לאחר מבחנים התגבשות נרחבת.
הנתונים והמתודולוגיה המתוארים כאן הדריכו אותנו לבחור את ה-OGNG כתכשיר הניקוי המועדף ביותר לצורך התגבשות מורכבות האודוסין-מיני-ג'י, בשל גודלו הקטן ויכולתה לייצב את הקומפלקס. השתמשנו גם אנדו F1 כדי להבטיח את הרודוסין הטהורה היה מין הומוגנית. גבישים התקבלו לאחר מכן וקבענו את מבנה הגביש כדי ~ 3.1 Å4, שהיה רק מבנה הגביש השלישי של gpcr – מורכב בחלבון איתות14,29.
עבור חלבונים ממברנה מאוגד עם חלבון שותף, הם צריכים להיחשב שני חלבונים שונים. חלבון במדינות שונות תפקודית יש קונמציות שונות והוא ברמת אנרגיה שונה. לכן, מומלץ למטב את פרוטוקול ההכנה עבור כל מצב פונקציונלי כאשר הפרמטר עבור מצב לא פעיל אינו מועבר באופן מלא למצב המופעל. כמו כן, שלא לדבר על השינוי בתכונת החלבון המורכב על ידי קשירה של חלבון שותף. הפרוטוקול משתמש בשיטות הסטנדרטיות להכנת דגימת התגבשות כדי להכין חלבון ממברנה לא פעיל בדטרגנטים שונים, ואחריו הפעלת חלבונים והיווצרות מורכבות, ולאפיון איכות החלבון. כך, פרוטוקול זה יכול בקלות להיות כללית חלבונים ממברנה אחרים מתחמי שלהם למחקרים מבניים עם שינוי קטין.
Disclosures
CGT הוא יועץ וחבר המועצה המייעצת המדעית של Sosei Heptares. לכל הסופרים האחרים. אין מה לגלות
Acknowledgements
אנו מודים לפרופסור ד ר גיהארד פ. שטלר לתמיכה ארוכת הטווח שלו בפרויקט זה, ד ר רוג'ר ג'יי. פי דוסון והופמן לה רוש לתמיכה בתרבות התאים. עבודה זו הייתה בחסות קרן המדע הלאומי השוויצרי (מענקים 210030_153145 ו310030B_173335 ל-GFXS), ומימון ל-CGT ממועצת המחקר האירופית (EMPSI, 339995) ומועצת המחקר הרפואי (MRC U105197215). FP מודה ETH ציריך באמצעות המרכז הלאומי של היכולת במחקר מולקולרי Ultrafast מדע וטכנולוגיה (NCCR חובה) ו-ETH המדע השני והטכנולוגי של הטכנולוגיה (ETH FAST). FP, JM, AB ו-CJT מכירים את התמיכה הפיננסית לטווח ארוך ממכון פאול שירר.
Materials
| Name | Company | Catalog Number | Comments |
| 1D4 peptide | Peptide2.0 | Under request | |
| 9-cis retinal | Sigma-Aldrich | R5754 | |
| Autosampler A-900 | GE Healthcare | Discontinued | |
| C9G | Anatrace | N324 | |
| cOmplete, EDTA-free protease inhibitor coctail | Roche | 5056489001 | |
| Cymal-5 | Anatrace | C325 | |
| Cymal-5NG | Anatrace | NG325 | |
| Cymal-6 | Anatrace | C326 | |
| Cymal-6NG | Anatrace | NG326 | |
| DDM | Anatrace | D310 | |
| DM | Anatrace | D322 | |
| DMNG | Anatrace | NG322 | |
| Econo column | Bio-Rad | 7372512 | |
| Ettan LC | GE Healthcare | Discontinued | |
| FRAC-950 | GE Healthcare | Discontinued | |
| HPLC Water 2795 Separation Module | Waters AG | 720000358EN | |
| InstantBlue Protein Stain | Expedeon | ISB1L | |
| LCT Premier mass spectrometer (ESI-TOF) | Waters AG | - | |
| LMNG | Anatrace | NG310 | |
| Monitor UV-900 | GE Healthcare | 18110835 | |
| Nanodrop 1000 | Witec AG/ThermoFisher | Discontinued | |
| NuPAGE 4-12% Bis-Tris gel 1.0 mm, 15 well | ThermoFisher | NP0323BOX | |
| NuPAGE MES SDS buffer (20x) | ThermoFisher | NP0002 | |
| OGNG | Anatrace | NG311 | |
| PAGEr Minigel Chamber | Lonza | 59905 | |
| Reprosil 200 C18-AQ column | Morvay Analytik GmbH | #s1503 | |
| Superdex 200 Increase GL column | GE Healthcare | 28990944 | |
| Tabletop centrifuge 5424R | Eppendorf | 5404000413 | |
| Ultracentrifuge Optima XE-100 | Beckmann Coulter | A94516 | |
| ULTRA-TURRAX T25 | IKA WERKE | 0003725003 | |
| UV-VIS spectrophotometer | Shimadzu | UV-2401PC | |
| Waters 2487 Dual λ Absorbance Detector | Waters AG | - |
References
- Tate, C. G. Practical considerations of membrane protein instability during purification and crystallisation. Methods in Molecular Biology. 601, 187-203 (2010).
- Lebon, G., Bennett, K., Jazayeri, A., Tate, C. G. Thermostabilisation of an agonist-bound conformation of the human adenosine A(2A) receptor. Journal of Molecular Biology. 409 (3), 298-310 (2011).
- Deupi, X., et al. Stabilized G protein binding site in the structure of constitutively active metarhodopsin-II. Proceedings of the National Academy of Sciences. 109 (1), 119-124 (2012).
- Tsai, C. -. J., et al. Crystal structure of rhodopsin in complex with a mini-G o sheds light on the principles of G protein selectivity. Science Advances. 4 (9), (2018).
- Carpenter, B., Tate, C. G. Engineering a minimal G protein to facilitate crystallisation of G protein-coupled receptors in their active conformation. Protein Engineering Design and Selection. 29 (12), 583-594 (2016).
- Chae, P. S., et al. Maltose-neopentyl glycol (MNG) amphiphiles for solubilization, stabilization and crystallization of membrane proteins. Nature Methods. 7 (12), 1003-1008 (2010).
- Loll, P. J. Membrane proteins, detergents and crystals: what is the state of the art. Acta Crystallographica Section F Structural Biology Communications. 70 (12), 1576-1583 (2014).
- Chae, P. S., et al. Glucose-neopentyl glycol (GNG) amphiphiles for membrane protein study. Chemical communications. 49 (23), 2287-2289 (2013).
- Standfuss, J., Xie, G., Edwards, P. C., Burghammer, M., Oprian, D. D., Schertler, G. F. X. Crystal structure of a thermally stable rhodopsin mutant. Journal of Molecular Biology. 372 (5), 1179-1188 (2007).
- Kaushal, S., Ridge, K. D., Khorana, H. G. Structure and function in rhodopsin: the role of asparagine-linked glycosylation. Proceedings of the National Academy of Sciences of the United States of America. 91 (9), 4024-4028 (1994).
- Molday, L. L., Molday, R. S. 1D4: a versatile epitope tag for the purification and characterization of expressed membrane and soluble proteins. Methods in Molecular Biology. 1177 (604), 1-15 (2014).
- Carpenter, B., Tate, C. G. Expression and Purification of Mini G Proteins from Escherichia coli. Bio-Protocol. 7 (8), (2017).
- Grueninger-Leitch, F., D'Arcy, A., D'Arcy, B., Chène, C. Deglycosylation of proteins for crystallization using recombinant fusion protein glycosidases. Protein Science. 5 (12), 2617-2622 (1996).
- Rasmussen, S. G. F., et al. Crystal structure of the β2 adrenergic receptor-Gs protein complex. Nature. 477 (7366), 549-555 (2011).
- Loginova, M. Y., Rostovtseva, Y. V., Feldman, T. B., Ostrovsky, M. A. Light damaging action of all-trans-retinal and its derivatives on rhodopsin molecules in the photoreceptor membrane. Biochemistry (Moscow). 73 (2), 130-138 (2008).
- Alexandrov, A. I., Mileni, M., Chien, E. Y. T., Hanson, M. A., Stevens, R. C. Microscale Fluorescent Thermal Stability Assay for Membrane Proteins. Structure. 16 (3), 351-359 (2008).
- Sonoda, Y., et al. Benchmarking Membrane Protein Detergent Stability for Improving Throughput of High-Resolution X-ray Structures. Structure. 19 (1), 17-25 (2011).
- Maeda, S., et al. Crystallization scale preparation of a stable GPCR signaling complex between constitutively active rhodopsin and G-protein. PloS One. 9 (6), 98714 (2014).
- Boute, N., Jockers, R., Issad, T. The use of resonance energy transfer in high-throughput screening: BRET versus FRET. Trends in Pharmacological Sciences. 23 (8), 351-354 (2002).
- Singhal, A., Guo, Y., Matkovic, M., Schertler, G., Deupi, X., Yan, E. C. Y. Structural role of the T 94 I rhodopsin mutation in congenital stationary night blindness. EMBO Report. 17 (10), 1-10 (2016).
- Choe, H. -. W., et al. Crystal structure of metarhodopsin II. Nature. 471 (7340), 651-655 (2011).
- Mattle, D., et al. Ligand channel in pharmacologically stabilized rhodopsin. Proceedings of the National Academy of Sciences of the United States of America. 115 (14), 3640-3645 (2018).
- Okada, T., Fujiyoshi, Y., Silow, M., Navarro, J., Landau, E. M., Shichida, Y. Functional role of internal water molecules in rhodopsin revealed by X-ray crystallography. Proceedings of the National Academy of Sciences of the United States of America. 99 (9), 5982-5987 (2002).
- Blankenship, E., Vahedi-Faridi, A., Lodowski, D. T. The High-Resolution Structure of Activated Opsin Reveals a Conserved Solvent Network in the Transmembrane Region Essential for Activation. Structure. 23 (12), 2358-2364 (2015).
- Magnani, F., et al. A mutagenesis and screening strategy to generate optimally thermostabilized membrane proteins for structural studies. Nature Protocols. 11 (8), 1554-1571 (2016).
- Kawate, T., Gouaux, E. Fluorescence-detection size-exclusion chromatography for precrystallization screening of integral membrane proteins. Structure. 14 (4), 673-681 (2006).
- Standfuss, J., et al. The structural basis of agonist-induced activation in constitutively active rhodopsin. Nature. 471 (7340), 656-660 (2011).
- Singhal, A., et al. Insights into congenital stationary night blindness based on the structure of G90D rhodopsin. EMBO reports. 14 (6), 520-526 (2013).
- Carpenter, B., Nehmé, R., Warne, T., Leslie, A. G. W., Tate, C. G. Structure of the adenosine A(2A) receptor bound to an engineered G protein. Nature. 536 (7614), 104-107 (2016).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved