
Method Article
Başarılı Kristalizasyon Için Görsel GPCR-mini-G Protein Sinyal Kompleksi Stratejik Tarama ve Karakterizasyonu
Bu Makalede
Özet
Bu rapor, görsel GPCR, rodopsin ve mini-Goile karmaşık hazırlamak için farklı deterjanların tarama açıklar. Arınma sırasında kompleksin farklı aşamalarında kalitesini karakterize eden biyokimyasal yöntemler gösterilmiştir. Bu protokol gelecekteki yapısal çalışmaları için diğer membran protein komplekslerine genelleştirilebilir.
Özet
Membran protein komplekslerinin kristal yapılarını belirlemenin anahtarı kristalleşmeden önce numunenin kalitesidir. Özellikle deterjan seçimi önemlidir, çünkü kompleksin hem stabilitesini hem de monodispersitliğini etkiler. Yakın zamanda sığır rodopsinaktif bir devlet kristal yapısını tespit mühendislik G proteini ile birleştiğinde, mini-Go, 3.1 şçözünürlükte. Burada rodopsin-mini-Go kompleksinin hazırlanmasını optimize etme prosedürünü ayrıntılarıyla anlatacağız. Koyu hal rodopsin klasik ve neopentil glikol (NPG) deterjanlarda hazırlandı ve ardından ışık maruziyeti altında mini-Go ile karmaşık oluşum lar yapıldı. Rodopsin stabilitesi ultraviyole görünür (UV-VIS) spektroskopisi ile değerlendirildi, Hangi ışığa duyarlı ligand rodopsin içine reconstitution izler, 9-cis retinal. Rodopsin ve rodopsin-mini-Go kompleksinin monodispersitlüğünü karakterize etmek için otomatik boyut dışlama kromatografisi (SEC) kullanıldı. SDS-poliakrilamid elektroforezi (SDS-PAGE), jeli Coomassie mavisi ile boyandan sonra rodopsin ve mini-Go arasında 1:1 molar oranı belirleyerek kompleksin oluşumunu doğruladı. Tüm bu analitik verileri çapraz doğruladıktan sonra, uygun olmayan deterjanları ortadan kaldırdık ve büyük ölçekli hazırlık ve kristalizasyon için en iyi aday deterjanla devam ettik. N-glikozilasyonun heterojenliği nden kaynaklanan ek bir sorun daha ortaya çıkmıştır. Heterolog olarak ifade edilen rodopsin, SDS-PAGE'de iki farklı N-glikozilasyon popülasyonuna sahip olduğu gözlendi, bu da muhtemelen kristallogenezi engelledi. Bu nedenle, farklı deglikozilasyon enzimleri test edildi ve endoglycosidase F1 (EndoF1) N-glikozilasyon tek bir tür ile rodopsin üretti. Protein kalitesini niteleme amaçlı bu stratejik boru hattı ile rodopsin-mini-Go kompleksinin hazırlanması kristal yapıyı sağlamak için optimize edildi. Bu, GPCR-G protein sinyal kompleksinin sadece üçüncü kristal yapısıydı. Bu yaklaşım, numune hazırlama ve yapı tayinini kolaylaştırmak için diğer membran proteinleri ve kompleksleri için de genellenebilir.
Giriş
Membran proteinlerinin ve komplekslerinin kristal yapılarının belirlenmesi, iyi yayılan kristallerin elde edilmesindeki güçlükler nedeniyle her zaman zor olmuştur. Çözünür proteinlerin aksine, integral membran proteinleri hücre zarını kapsayan hidrofobik bir çekirdek oluşturur. Hücre zarından sulu tampona membran proteinlerini çıkarmak için deterjanlar deterjan-protein mikelleri oluşturmak için kullanılmalı ve böylece membran proteinlerinin hidrofobik çekirdeğietrafındaki lipitlerin değiştirilmesi gerekir. Membran proteinlerinin stabilitesi, aktivitesi ve bütünlüğü doğrudan deterjan1'inkimyasal ve yapısal özelliklerine bağlıdır ve deterjan ın özellikleri de mikenin boyutunu belirler. Büyük bir deterjan micelle küçük bir membran proteininhidrofilik yüzeyleri tıkamak, böylece buhar difüzyon yöntemi kullanırken kristal temas eksikliği nedeniyle kristalizasyon önleyebilir. Küçük bir deterjan micelle kristalografi için avantajlıdır, ancak kısa zincirli deterjanlar genellikle daha serttir ve bu nedenle membran proteininin dengesinibozan ve toplanmasına yol açar. Bu nedenle, kristalizasyondan önce, ek bir deterjan tarama prosedürü vazgeçilmezdir, genellikle protein stabilitesini hala koruyan daha kısa deterjanları hedeflemek.
G protein-coupled reseptörleri (GpR) yedi transmembran a-helikler içeren integral membran proteinleridir. KPCR iki ana durumda var, ya ters agonistler veya antagonistler tarafından stabilize bir inaktif devlet, ya da aktif bir devlet bir agonist bağlı ve bir G protein tarafından stabilize, bu iki uç arasında çok sayıda alt devletler var olması muhtemeldir rağmen. GPCR yapı tayini başlangıçta aktif devletlere göre daha yüksek istikrar nedeniyle ters agonistler ve antagonistler bağlı inaktif devletler üzerinde duruldu2. KPCR agonist bağlama üzerine aktive edildiğinde, reseptörleri son derece dinamik, ve yarık g protein kaplin reseptörünün sitoplazmik yüzünde geçici olarak formları. Bu dinamizmin agonist bağlı GPCR'lerin genellikle aktif olmayan durumdan daha kararsız olmasının nedeni olduğu düşünülmektedir. Bu nedenle, çalışma altındaki reseptörün konformasyonel durumuna uygun deterjanların taranması gerekli hale gelir, çünkü daha hafif deterjanların aktif bir durumu incelemek için inaktif bir duruma göre gerekli olması muhtemeldir.
Bu raporda, görsel GPCR, sığır rodopsin3ve mini-Go protein4,ile kompleks,5 deterjan tarama deneyleri için, inaktif durumu ve aktif durumu temsil, sırasıyla kullanın. Deterjan taraması klasik alkil maltoside ve glukoside deterjanlar ve neopentil glikol (NPG) deterjanları üzerinde duruldu. Bu bağlamda, klasik bir deterjan bir şeker kafa grubu ve alkil zinciri inşa edilir, NPG tipi deterjanlar şekerler ve alkil zincirleri arasındaki arayüz de kuaternary karbon tarafından erimiş iki özdeş klasik deterjan içerir iken6,7,8.
Farklı deterjanlarda rodopsinin arıtılmasından başlayarak deneysel bir iş akışı tasarlanmıştır, ardından rodopsin-mini-Go kompleksinin oluşumu ve kompleksin çeşitli yöntemlerle karakterizasyonu ile sona erdirilmiştir(Şekil 1). Rodopsinin inaktif durumu için ışığa duyarlı ligand 9-cis retinal reconstitution ultraviyole görünür (UV-VIS) spektroskopi ile izlendi. Spektrum retinanın fizikokimyasal durumunu ortaya çıkarır ve rodopsin retina bağlayıcı cebinde çevresinin göstergesidir. Arınmış rodopsinin monodispersitlüğünü ve rodopsin-mini-Go kompleksinin oluşumunu değerlendirmek için boyut dışlama kromatografisi (SEC) kullanılmıştır. SEC protein moleküllerini büyüklüklerine ve şekillerine göre farklılaştırırken, birleştirilmiş protein popülasyonu boşluk hacminde elat olarak tanımlanabilir. Karmaşık oluşumu doğrulamak için, SEC kesirler sodyum dodecyl sülfat-poliakrilamid jel elektroforez (SDS-PAGE) hem rodopsin ve mini-Govarlığını doğrulamak için değerlendirildi .
Dikkate alınması gereken bir diğer faktör membran proteinleri üzerinde post-translational değişiklikler (PTM) olduğunu. N-glikozilasyon gibi PTM genellikle memeli ve böcek hücre ekspresyon sistemlerinde üretilen ökaryotik membran proteinlerinde gözlenmektedir. İnsan embriyonik böbrek 293 (HEK293) hücrelerinin sınırlı bir N-glikozilasyon suş n-asetilglucosaminyltransferaz I (GnTI) kodlayan genin silinmesi ile geliştirilmiştir, glcNAc2Man5 tarafından homojen N-glikozilasyon sonuçlanan asn-X-Ser/Thr. N-glikozilasyon konsensüs yerinde bir amino asit kalıntısı mutasyona uğratılması ile önlenebilir olsa da, bu da protein in işlevini veya katlama verimliliğini değiştirebilir. Büyükbaş rodopsinde, N-glikozile kalıntı Asn15 mutasyonu yanlış katlama ve azaltılmış G protein aktivasyonu yol açar9,10. Bu raporda kullanılan rodopsin HEK 293 GnTI eksikliği hücre hattında ifade edilebildi. Ancak, SDS-PAGE rodopsin iki tür varlığını gösterdi. Bu heterojenlik kristal oluşumunu engelleyebilir ve bu nedenle peptid-N-glikosidaz F (PNGase F) ve endoglycosidase F1 (Endo F1) kullanılarak deglikozilasyon test edildi. Deglikozilile ürün glikozilasyon düzeyini ve homojenliğini belirlemek için SDS-PAGE ve sıvı kromatografi-kütle spektrometresi (LC-MS) ile karakterize edildi.
Protokol
NOT: Deterjan taraması için bu protokol başlangıç malzemesi olarak 30 g HEK293 hücreli pelet için ayrıntılı olarak açıklanır.
1. Malzemeler, kimyasallar ve reaktifler
NOT: Tüm çözümler analitik sınıf reaktifler ve 25 °C'de 18,2 MΩ°cm direnç sağlamak için deiyonize sudan arıtılmış ultra saf su kullanılarak hazırlanır.
- Tampon stok çözümleri
- 10x fosfat tamponlu salin (10x PBS) hazırlayın.
- HEPES tampon hazırlayın: 1 M, NaOH ile pH 7.5 titrated.
- 5 M Nacl hazırlayın.
- Hazırlayın 2 M MgCl2.
NOT: Tüm stok çözümleri, sterilitelerini korumak için 0,22 μm'lik bir filtreden geçirilir.
- Deterjan stok çözümleri
- Dodecyl maltoside (DDM), 10% (w / v) hazırlayın.
- Decil maltoside (DM), %10 (w/v) hazırlayın.
- 6-sikloheksil-heksil maltoside (Cymal-6), %10 (w/v) hazırlayın.
- Hazırlamak 5-sikloheksil-pentyl maltoside (Cymal-5), 10% (w / v).
- Nonyl glukoside (C9G), %10 (w/v) hazırlayın.
- Lauryl maltoz neopentil glikol (LMNG), %5 (w/v) hazırlayın.
- Decyl maltoz neopentil glikol (DMNG), %10 (w/v) hazırlayın.
- Simal-6 neopentil glikol (C6NG), %10 (w/v) hazırlayın.
- Simal-5 neopentil glikol (C5NG), %10 (w/v) hazırlayın.
- Otol glukoz neopentil glikol (OGNG), %10 (w/v) hazırlayın.
NOT: %10 deterjan stok çözeltisi için, 1 g deterjan tozunu hafif sallanan ultra saf suda çözünve son hacmi 10 mL'ye ayarlayın. Deterjan stok çözeltisi uzun süreli depolama için -20 °C'de ve çalışırken buz üzerinde tutulmalıdır.
DİkKAT: Şişelenmiş deterjanların genellikle -20 °C'lik dondurucuda sakkullanılması tavsiye edilir. Deterjan tozu içeren şişeler açılmadan önce oda sıcaklığına kadar ısıtılmalıdır. Deterjan tozu higroskopiktir, bu nedenle sıcaklık dengesi deterjanı ıslayacak yoğuşma oluşumunu önler.
- Diğer kimyasallar ve reaktifler
- Hazırlamak 1D4 immünafinite agarose resin: 10 mL 50% bulamaç.
NOT: 1D4 immünafinite agarose monoklonal Rho1D4 antikor ile bağlantılı agarose boncuklar, epitop olarak büyükbaş rodopsin TETSQVAPA son 9 amino asitler bağlar. 1D4 immünafinite agarose c-terminal 1D4 dizisi içeren proteinleri yakalamak için afinite saflaştırma malzemesi olarak çalışır. Bu arıtma malzemesi9,11 veyasatın alınabilir. - 9-cis retinal çözelti hazırlayın: 1 mM, % 100 etanol içinde çözünmüş.
NOT: Hazırlık ve depolama sırasında retinaya ışık maruziyetini önleyin. - Hazırlamak 1D4 peptid (sıraTETSQVAPA): 800 μM, suda çözünmüş.
- Hazırlamak 1D4 immünafinite agarose resin: 10 mL 50% bulamaç.
- Arabellek
NOT: Tüm tamponlar stok çözeltilerinden istenilen konsantrasyona karıştırılır. Tüm tamponlar kullanılmadan önce 4 °C'ye kadar soğutulur.- Arabellek A hazırlayın: PBS, 0.04% DDM.
- Hazırara B: 20 mM HEPES pH 7.5, 150 mM NaCl, 0.04% DDM.
- Hazırlayın Tampon C: 20 mM HEPES pH 7.5, 150 mM NaCl ve deterjan çalışma konsantrasyonlarında Tablo 1'delistelenmiştir.
- Hazırara D: 20 mM HEPES pH 7.5, 150 mM NaCl.
- Elüsyon tamponunu hazırlayın: 20 mM HEPES pH 7.5, 150 mM NaCl, 80 μM 1D4 peptid ve deterjan çalışma konsantrasyonlarında.
- SEC tamponhazırlayın: 20 mM HEPES pH 7.5, 150 mM NaCl, 0.025% DDM; 0,22 μm'lik bir filtre ile filtratedilir.
- LC-MS için çözücü
- Çözücü A hazırlayın: %0.1 formik asit içeren asetonitril.
- Çözücü B hazırlayın: %0,1 formik asit içeren ultra saf su.
- Solvent C: iso-propanol hazırlayın.
2. Hücre zarı çözünürasyonu ve protein ekstraksiyonu
- HeK293 GnTI 30 g- - hücre pelet büyükbaş rodopsin mutant N2C/M257Y/D282C3ifade,oda sıcaklığına9, 120 mL 1x PBS tampon protetratör kokteyl içeren ekleyin ve bir Dounce homogenizer veya bir homozergeni (13.000 rpm 30 s) kullanarak homojenize. Homojenleştirilmiş hücre süspansiyonuna bir kabın içinde toplayın ve hacmi 150 mL'ye ayarlayın.
NOT: 30 g hücre peleti 2 x 106 hücre/mL yoğunluğunda hücre kültürünün 3 L'sine eşittir. - %1.25'lik son konsantrasyonu vermek için homojenize hücrelere %10 DDM ekleyin. 1 saat buz üzerinde karıştırın.
- 4 °C ve 150.000 x g'de hücre lisatını 45 dk'da santrifüj edin ve çözünmemiş enkazı temizleyin.
- Supernatant'ı 500 mL'lik bir şişeye aktarın ve 1D4 immünafinite sinin 10 mL'sini ekleyin agarose reçine (%50 bulamaç). Çözünür hücreli ve reçin'i 4 saat veya gece boyunca 4 °C'de hafifçe karıştırın.
- Reçine toplamak için lysate/reçine karışımını açık bir kolona yükleyin.
- Reçineyi 10 sütun hacmi (CV) ile yıkayın Arabellek A.
NOT: Sütun hacmi paketlenmiş hacmi (%100) agarose reşin kullanılır. Bu durumda 1 CV 5 mL'dir. - Buffer A'nın 2 CV'si ile resenin isini yeniden askıya alın.
DİkKAT: 2.8. adımdan itibaren, loş kırmızı ışık koşulualtında yapılması gereken adımlar açıklamanın başında "[Karanlık]" olarak etiketlenir. - [Karanlık] Resuspended resin'e 9 cis retinal ekleyin ve 50 μM'lik son konsantrasyona ekleyin. Karanlıkta 4°C'de 4-16 saat hafifçe karıştırın.
NOT: Daha kısa kuluçka süresi retinanın eksik yeniden yapılandırılmasına yol açabilir. - [Karanlık] Sütundaki akışı kaldırın. 20 CV Tampon A ile reçine yıkayın, ardından 15 CV Tampon B.
- [Karanlık] Rezorini 2 CV Arabellek B'de yeniden askıya alın ve rezorin süspansiyonuna eşit olarak 10 mL'lik imha sütununa bölün.
- [Karanlık] Sütundan akışı çıkarın ve 1 mL Tampon C. 4 °C'de 1 saat kuluçka için reçineyi yeniden askıya alın.
- [Karanlık] Adımı tekrarlayın 2.11.
- [Karanlık] Sütundaki akışı kaldırın ve her sütun için 0,8 mL Elüsyon Arabelleği'nde reçineyi yeniden askıya alın. Yavaşça 2 saat karıştırın.
- [Karanlık] 2 mL tüp içine sütundan elütion toplayın.
- [Karanlık] Her sütun için 0,7 mL Elütion Arabelleği reçinesini yeniden askıya alın. Hafifçe 1 saat karıştırın.
- [Karanlık] Aynı tüp içine sütundan elütion toplamak.
3. UV-VIS spektroskopisi
- Spektrofotometreyi 250-650 nm ölçüm aralığını kapsayacak şekilde hazırlayın. Su veya Elüsyon Arabelleği kullanarak taban çizgisini kaydedin.
- [Karanlık] Eluted proteini kuvars cuvette yükleyin. Protein örneğinin spektrumu ölçün.
- [Karanlık] 495 nm uzun geçişli bir filtreden geçen ışıkla 2 dakika boyunca proteini doğrudan cuvette'de aydınlatın.
- Işıklı numunenin tayfını ölçün.
- Diğer 9 deterjanda saflaştırılmış tüm protein örnekleri için aynı ölçümü yapın, hem karanlık hem de aydınlatılmış hallerde.
- X-Y dağılım grafiğindeki eğrileri (absorbance versus dalga boyu) çizin.
4. Rodopsin ve rodopsin-mini-Go kompleksinin otomatik boyut dışlama kromatografisi
- [Karanlık] 4 °C'de 30 kDa moleküler ağırlık kesme (MWCO) ile bir spin konsantratörü kullanarak 100 μL'ye konsantre protein. Aşırı konsantre numuneler, konsantratör veya Tampon C'den gelen akış kullanılarak seyreltilebilir. Protein numunesi konsantrasyonu belirlemek için, bir spektrofotometre kullanarak 280 nm'de absorbansı ölçün.
NOT: Adım 4.2'den itibaren, deney karanlık bir ortam gerektirmez ve bu nedenle numuneler normal ışık altında hazırlanabilir. - Her deterjan durumu için 0,7 mg/mL'de 100 μL rodopsin hazırlayın.
- Her deterjan durumu için 100 μL rodopsin (0.7 mg/mL) ve mini-Go4,12 (0.2 mg/mL) karışımı hazırlayın. Karışımı 1 mM MgCl2ile tamamlayın. Karışımı 495 nm uzun geçişli filtreden gelen ışıkla aydınlatın ve 30 dakika kuluçkaya yatırın.
- Sıvı kromatografi temizleyiciüzerine 10-600 kDa küresel protein fraksiyonu olan 24 mL jel filtrasyon kolonunu monte edin. Sütunu SEC arabelleğiyle dengeler.
NOT: Sıvı kromatografi temizleyici bir autosampler, çoklu dalga boyu dedektörü ve bir fraksiyon toplayıcı ile donatılmıştır. - Örnekleri otonumune şişelerine aktarın ve numune tepsisine yerleştirin. Her örnek için sıralı SEC çalışanlarını otomatikleştirmek için bir yöntem dosyası programlayın, otomatik örnekleyici numunenin 77 μL'sini kolona yükleyerek ve 24 mL'lik SEC arabelleğini çalıştıran 0,5 mL/dk akış hızında temizleyici yedirin. 280 nm ve 380 nm de absorbans kaydedin.
- 12.9 mL civarında tutma hacminde rodopsin ve rodopsin-mini-Go kompleksinin tepe fraksiyonlarını toplayın.
- 4.2. adımdan sol rodopsin örneklerini ve coomassie mavisi boyama ile %4-12 SDS-denatüre gradyan jeller üzerinde rodopsin-mini-Go kompleksinin pik fraksiyonlarını analiz edin.
- Elütion kromatogramını (A280 veya A380 ve bekletme hacmi) çizin.
5. Deglikozilasyon ve LC-MS çalışması
- LC-MS çalışması için sadece LMNG deterjanı ile saflaştırılmış rodopsin örneğini kullanın.
- 1 mg/mL'de 200 μL rodopsin ve 0.01 mg/mL'de PNGase F13 karışımı hazırlayın. İyice karıştırın ve gece boyunca 4 °C'de kuluçkaya yatırın.
- 1 mg/mL'de 200 μL rodopsin ve 0.01 mg/mL'de Endo F113 karışımıhazırlayın. İyice karıştırın ve gece boyunca 4 °C'de kuluçkaya yatırın.
- SDS-PAGE ve Coomassie mavi boyama ile sindirim sonucu analiz edin.
- İşlenmemiş ve Endo F1 ile işlenmiş rodopsin örneklerine konsantre olun ve Tampon D'de SEC saflığına tabi olun.
NOT: Bu LC-MS çalışması için deterjan en az miktarda örnek hazırlamaktır. Tampon D herhangi bir deterjan içermez, ancak bir membran protein14LMNG yavaş off-rate nedeniyle , rodopsin agrega olmaz. - 12,9 mL civarında tutma hacminde tepe fraksiyonu toplayın. Spin konsantratörü (MWCO 30 kDa) kullanarak 1 mg/mL'ye konsantre olun.
- Bir Reprosil 200 C18-AQ sütununa proteinin 10 μg'sini enjekte edin ve tablo 2'delistelenen çözücü bileşimi ve ayarları ile doğrusal degrade yöntemini kullanarak kolonu kullanın. Akış kütle spektrometresi için %25'e, UV tespiti için %75'e bölünür.
Sonuçlar
Numune hazırlama ve analiz için deneysel iş akışı Şekil 1'deözetlenmiştir. Küçük ölçekli afinite arınma için açık sütunlar kullanmak, birçok farklı deterjan koşullarında numune hazırlamamızı sağlar(Şekil 1A). Böyle küçük ölçekli bir arınma kurulumu UV-VIS spektroskopisi, SEC ve SDS-PAGE(Şekil 1B-C)kullanılarak daha fazla analiz için yeterli protein vermiştir.
UV-VIS spektroskopisinde rodopsin stabilitesi saptadı
Retinal rekonkorpsinin stabilitesi optik emiciliği ile değerlendirildi (Şekil 2). Karanlık durumda, 9 cis retinal kovalent lys296 bir protonlanmış Schiff baz olarak bağlantılıdır. Aydınlatmadan sonra, 9-cis retinal all-trans izoform izomerize edilir ve Schiff baz bağlantısı deprotonated. Protonlanmış 9-cis retinal 488 nm emilim tepe verir, deprotonated all-trans retinal 380 nm bir tepe ye sahiptir. DDM'de rodopsinIN UV-VIS spektrumları 9 cis retinal bağlı ve ışıkla aktive edilmiş rodopsinin tipik emilimini gösterdi, kabaca aynı optik yoğunlukta 108 nm mavi bir kayma açıkça gözlendi(Şekil 2A, sol üst panel). Rodopsin dengesizolduğunda, ve sonra retina değişiklikleri için bağlayıcı cep, hangi retina deprotonation ve muhtemelen dissosilasyon ile sonuçlanır. Bu durumda, ve sonra spektrum deprotonasyon katkısı yanı sıra retinal serbest formu gösterir15. Bu nedenle, protein (280 nm) ve retinal (protonlanmış 9 cis retinal için 488 nm, deprotonated all-trans retinal için 380 nm) arasındaki absorbans oranı ile rodopin içine retina reconstitutionin etkinliğini belirledik (Şekil 2B). Klasik deterjanlarda (DDM, DM, Cymal-6, Cymal-5, C9G) arıtılmış rodopsin örnekleri aynı optik profili gösterir. Ancak, NPG deterjanlarında (LMNG, DMNG, Cymal-6NG, Cymal-5NG) saflaştırılan numunelerde, DDM örneği ile aynı optik profili veren OGNG örneği dışında retinaiçin en uygun olmayan bağlama ortamını öneren optik profiller gösterilmiştir.
Boyut dışlama kromatografisinde örnek saflık ve protein monodispersitliği saptadı.
SEC, hazırlama ve tarama sırasında protein örneklerini değerlendirmek için etkili ve sağlam bir analitik araçtır. Bir önceki saflaştırma adımından alınan numune saflığını ve protein moleküllerinin monodispersitliğini doğrular. Rodopsin ve mini-Go kompleksi için numune kalitesi 280 nm ve 380 nm 'deki emilim eğrilerinden yorumlandı(Şekil 3A). 280 nm iz protein in varlığını gösterdi, ve 380 nm iz retinal varlığını gösterdi. Boşluk hacminde görünen tüm sinyaller (bu sütunu kullanırken yaklaşık 8 mL) protein agregalarına atfedilen. Bu nedenle, sonuçlar klasik deterjanlarda hazırlanan örneklerin C9G dışında monodispers durumda olduğunu göstermiştir ve burada agreganın bir kısmı ortaya çıkmıştır. Buna karşılık, NPG tipi deterjanlar kullanılarak hazırlanan numuneler C9G örneğinden çok daha fazla agrega içeriyordu; LMNG ve Cymal-6NG en agrega oluşumuna yol açtı, ancak DMNG ve Cymal-5NG'de daha az agrega gözlendi. Bunun istisnası, DDM'ye benzer bir profil gösteren OGNG idi. ~12.9 mL'lik tutma hacmindeki zirveye kıyasla 135 kDa'ya karşılık gelen A280/A380 oranında gösterildiği gibi, boşluk hacminde bulunan protein agregaları da daha zayıf retinal doluluk oranına sahipti. Gözlemlediğimiz bir diğer özellik de hem rodopsin hem de rodopsin-mini-Go'nun aynı tutma hacmietrafında eluted olmasıdır (Şekil 3B). Deterjana bağlı rodopsinin moleküler ağırlığı 120 kDa ve rodopsin-mini-Go 144 kDa olduğu için bu şaşırtıcı değildir. Bu nedenle karmaşık oluşumu yalnızca SEC verilerinden tespit edemedik, bu yüzden SDS-PAGE SEC tarafından saflaştırılmış örneği daha fazla analiz etmek için kullanıldı.
SDS-PAGE doğrulanmış karmaşık oluşum
SDS-PAGE, bir numunedeki protein bileşenlerini tanımlamak için kullanılan standart bir yöntemdir. Konsantre rodopsin (SEC arınma önce) saflığını doğrulamak için SDS-PAGE tarafından analiz edildi ve 37 kDa yakın iki bant ve 50 kDa üzerinde bir lekeli bant gösterdi(Şekil 4A). Alt iki bant daha sonra farklı N-glikozilasyon durumları olduğu doğrulandı. 50 kDa'nın üzerindeki bant, SDS-PAGE örnek tamponu tarafından indüklenen toplanmış rodopsin oligomerleri olarak yorumlandı, çünkü bu agregalar SEC veya başka bir algılama yönteminde gözlenmemiştir. SEC verileri karmaşık oluşumu doğrulayamadığından, SEC rodopsin-mini-Go örneklerinden kesirler sds-PAGE kullanılarak analiz edildi. SDS-PAGE tüm deterjan koşullarında hem rodopsin hem de mini-Go protein bantlarını göstererek, deterjan seçimine bakılmaksızın kompleksin oluştuğunu düşündürmektedir(Şekil 4B).
LC-MS spektrometresi rodopsinde N-glikozilasyon paterni saptandı
Hem yakınlık arınması hem de SEC'den alınan rodopsin örnekleri, 24 mL'lik bir kolon kullanırken SEC ile ayrılamayan bir SDS-PAGE jelinde yaklaşık 37 kDa molekül ağırlığıyla göç eden iki protein bandı gösterdi. HEK 293 GnTI heterolog ifade rodopsin n-glikozilasyon farklıdesenleri - hücreler en olası açıklama oldu. Bu nedenle, iki enzim, PNGase F ve Endo F1, deglikozilat rodopsin yetenekleri için test edildi. SDS-PAGE verilerinden, Endo F1 her iki protein bandının moleküler ağırlığını tek bir ürüne dönüştürürken, PNGase F sindirimi hala iki popülasyon almıştır(Şekil 5A). Sindirilmemiş ve Endo F1 ile tedavi edilen örnekler farklı türlerin kütlelerini belirlemek için LC-MS spektrometresi kullanılarak analiz edildi. Veriler, HEK 293 GnTI'da üretilen- rodopsinin 1014±1 Da. Endo F1-tedavi rhodopsin kütle farkı ile bir veya iki N-glikan içerdiğini ve iki N-glikan içeren rodopsinle karşılaştırıldığında 2027±1 Da kütle farkı olduğunu gösterdi. Bu sonuçlar rodopsin ifade etmek için kullanılan hücre hattında enzim N-asetilglucosaminyltransferaz I yokluğu ile tutarlıdır, hangi yapısı GlcNAc sahip tüm N-glikanlar sonuçlanır2Man5, (kütle 1014 Da).

Şekil 1: Deterjan tarama deneyi için numune hazırlama ve karakterizasyon. (A) Arınma sırasında farklı deterjanlarda rodopsin örneklerinin hazırlanması. (B) Protokolde kullanılan yöntemler: UV-VIS spektroskopisi, boyut-dışlama kromatografisi (SEC), SDS-PAGE ve sıvı kromatografi-kütle spektrometresi (LC-MS). (C) Rodopsin, rodopsin-mini-Gove rodopsin deglikozilasyon ürünü karakterizasyonu için deneysel iş akışı. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
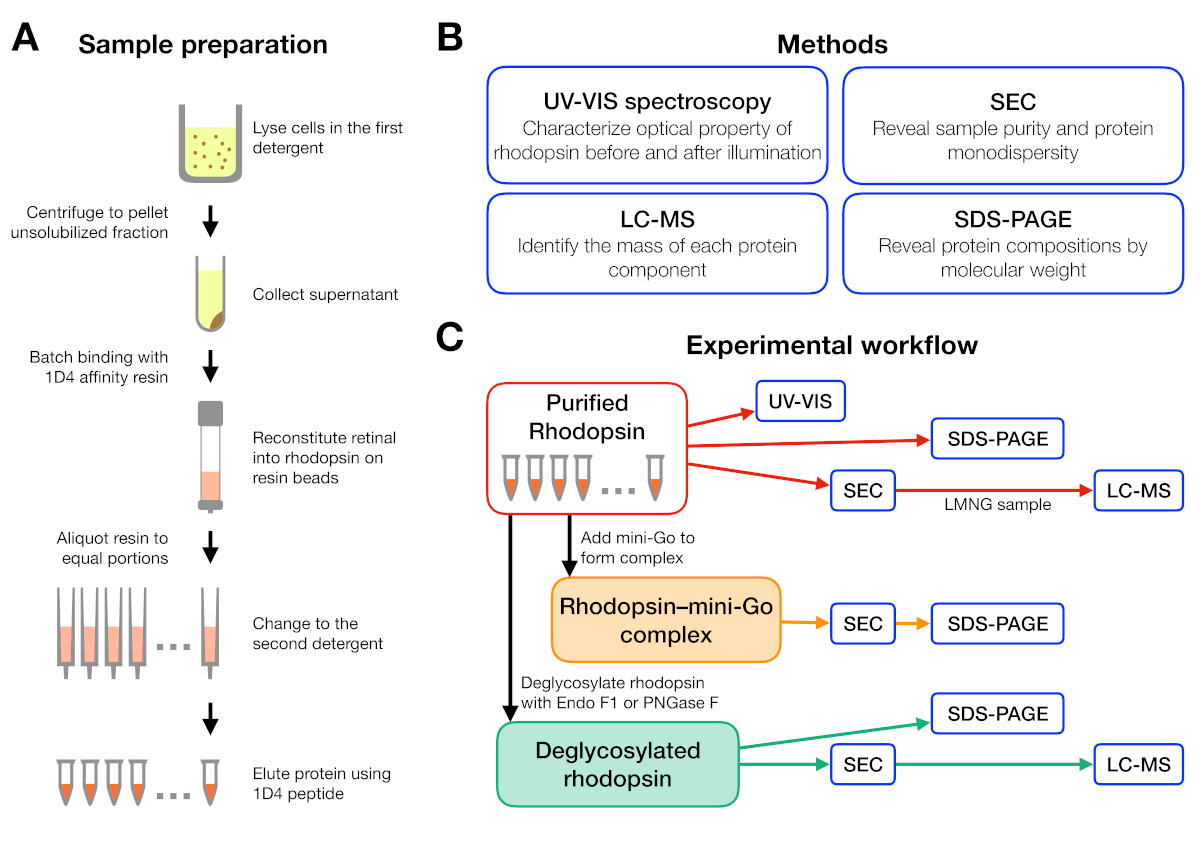{kind=link}

Şekil 2: Rodopsin UV-VIS spektroskopisi. (A) rodopsin UV-VIS spektrumları. Koyu hal, 9 cis retinal bağlı rodopsin spektrumları mavi eğriler gösterilir. Aydınlatmadan sonra, 9-cis retinal deprotonated ve tüm trans retina içine isomerizes, ve ışıklı rodopsin spektrumları kırmızı eğriler olarak gösterilir. Her deterjan kimyasal yapısı bir inset olarak gösterilir. (B) A280/A488 (mavi çubuk) ve A280/A380 (kırmızı çubuk) oranları sırasıyla karanlık durumda ve açık durumda rodopsin kararlılığını belirtir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
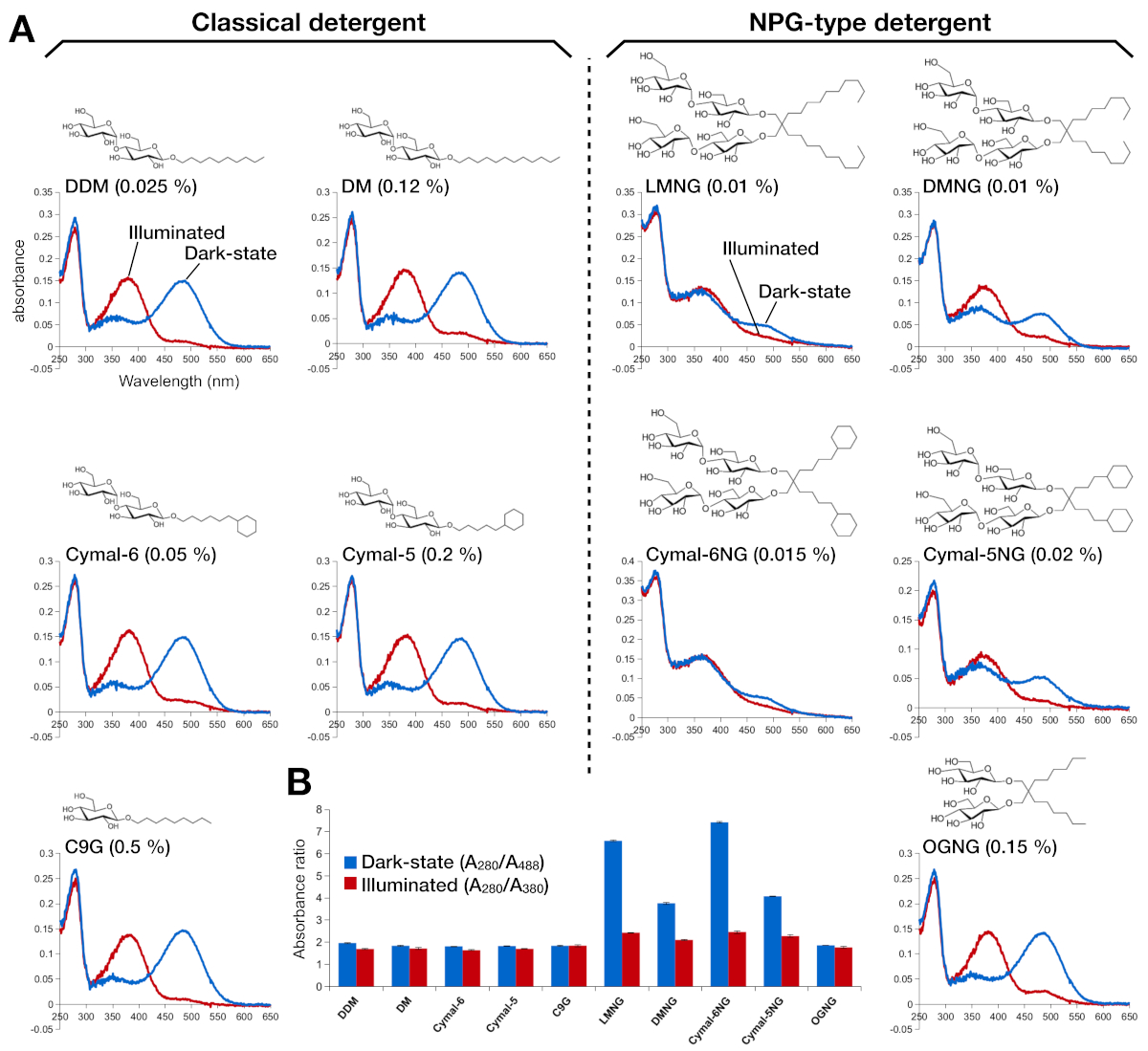{kind=link}

Şekil 3: Rodopsin ve rodopsinin boyut dışlama kromatografisi profilleri–10 farklı deterjanda saflaştırılmış mini-Go kompleksi. (A) Sol panel klasik deterjanlarda saflaştırılı örneklerin SEC profillerini gösterir. Sağ panel, NPG tipi deterjanlarda saflaştırılı numunelerin SEC profillerini temsil eder. Standart marker proteinlerin profili DDM örneği ile birlikte bindirme olarak gösterilir. DDM, DM, Cymal-6, Cymal-5 ve OGNG için ideal senaryo (agrega yok) ile en yüksek profillerin yorumlanması DMNG için gösterilir. (B) OGNG örneğinin 12-14 mL saklama hacmindeki büyütülmüş profili. Tüm örnekler Superdex200 Artış 10/300 GL sütunu kullanılarak analiz edildi. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: Rodopsin ve rodopsin/mini-Go kompleksinin SDS-PAGE analizi. (A) Deterjanlarda arıtılmış rodopsin örnekleri. 50 kDa'nın üzerindeki bulaşmış bant, SDS-PAGE örnek tamponu tarafından indüklenen birleştirilmiş rodopsin oligomerlerine atfedilir. (B) RODOPSİn/mini-Go kompleksi SEC saflaştırılmış örnekleri. 1 ve 2 N-glikan ve mini-Go ile rodopsin gösterilmiştir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 5: Rodopsinde glikozilasyonun belirlenmesi. (A) PNGase F ve Endo F1 kullanılarak deglikozilile rodopsin SDS-PAGE analizi. (B) LC-MS spektrumları rodopsin olmadan (üst panel) ve Endo F1 (alt panel) ile deglikozilasyon ile. Rodopsin-mini-Go kompleksini kristalizasyon için hazırlamak için PNGase F yerine Endo F1'i seçtik çünkü Endo F1 tek bir homojen rodopsin türü sundu. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
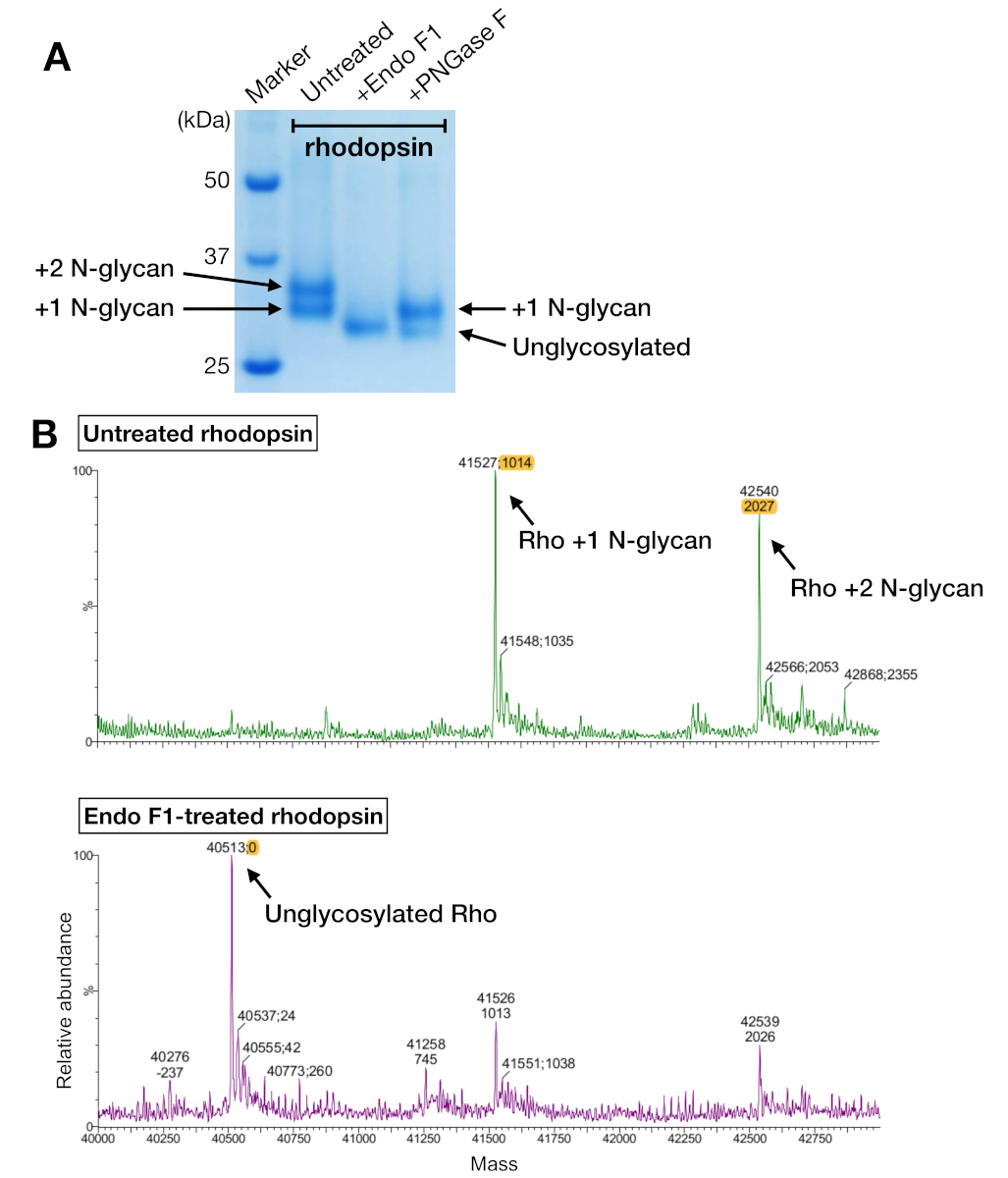{kind=link}
| Deterjan | Çalışma konsantrasyonu (%) | Kritik mikel konsantrasyonu (%) |
| DDM | 0.025 | 0.0087 |
| Dm | 0.12 | 0.087 |
| Cymal-6 | 0.05 | 0.028 |
| Cymal-5 | 0.2 | 0.12 |
| C9G | 0.5 | 0.2 |
| LMNG | 0.01 | 0.001 |
| DMNG | 0.01 | 0.0034 |
| Cymal-6NG | 0.015 | Mevcut değil; 0,056'dan düşük olmalıdır |
| Cymal-5NG | 0.02 | 0.0056 |
| OGNG | 0.15 | 0.058 |
Tablo 1: Tampon C deterjan konsantrasyonları.
| Süre (dk) | Çözücü A (%) | Çözücü B (%) | Çözücü C (%) | Akış hızı (ml/dk) |
| 0 | 0 | 95 | 5 | 0.5 |
| 1 | 0 | 95 | 5 | 0.5 |
| 5 | 20 | 75 | 5 | 0.6 |
| 25 | 85 | 10 | 5 | 0.6 |
| 26 | 90 | 5 | 5 | 0.6 |
| 30 | 90 | 5 | 5 | 0.6 |
Tablo 2: Sütun elüsyon parametreleri.
Tartışmalar
Protein kristalizasyonundaki başarı, deterjanların neden olduğu komplikasyon nedeniyle protein örneğine, özellikle membran proteinlerine ve komplekslerine bağlıdır. Bu rapor, GPCR-mini-G protein sinyal kompleksleri için deterjan taraması ve numune kalitesinin değerlendirilmesini göstermektedir. Çeşitli yöntemler yaygın membran proteinlerinin biyokimyasal özelliği ni incelemek için kullanılmıştır, örneğin, floresan boyalar kullanarak termostabilite tsay16,17, bağlayıcı teşbik triptofan floresan sinyali18 veya biyosensörler ile rezonans enerji transferi değişikliği ölçerek karmaşık oluşumunu tespit etmek için19. Ancak, bu yöntemlerde kullanılan kimyasal ortamlar bir kristalizasyon örneği hazırlamak için oldukça farklıdır, ya proteinler floresan bazlı ölçüm için bin kat daha düşük konsantrasyonda, ya da proteinler lipid iki katmanlı veya bir sabit deterjan durumda gömülüdür. Bu protokolde kullanılan yöntemler kristalizasyon dan önce büyük ölçekli numune hazırlamada da standartlaştırılmışolarak kullanılmaktadır. Bu nedenle, optimize edilmiş parametreler daha fazla ana tarama ve optimizasyon olmadan kristalizasyon ölçeğinde hazırlık için kolayca aktarılabilir.
Bu protokolün amacı, x-ışını kristalografisi ile buhar difüzyon kristalizasyonu ve yapı tayini için istikrarlı ve homojen bir GPCR-mini-G protein kompleksinin hazırlanmasını optimize etmektir. Protokol, rodopsin-mini-Go kompleksinin hazırlanması sırasında deterjan ve deglikozilasyonun etkisini nitel olarak değerlendirmek için bir dizi yöntemi entegre eder. Rodopin inaktif durumda ve transdüsin peptid olmadan bağlı ve ışık aktive devlet deterjanlar octyl glukoside saflaştırılmış zaman kristalize edilmiştir (C8G)20,21,22 ve C9G23,24. C8G ve C9G'de saflaştırılan rodopsin-mini-Go kompleksi kristal leri verim vermedikçe (veriler gösterilmedi), daha sonra açıklanan stratejiyi kullanarak daha geniş bir diğer deterjan yelpazesini araştırdık (Şekil 1). Rodopsinin ışık hassasiyetinden yararlanarak retinanın 280 nm'den başka dalga boylarında yeniden yapılandırılabileceğini söyleyebiliriz. Hem UV-VIS spektroskopisinde hem de SEC'de retinal 380 nm veya 488 nm olarak saptın. Ancak, çoğu membran proteinlerinin arınma sırasında işlevselliği takip etmek için böyle uygun bir kromofor yok. Diğer seçenekler bir ışık tespit kromofor ekleyerek veya radyoligand-bağlayıcı ve termal vardiya tahlilleri25kullanarak bir ligand tespit edilebilir hale getirmek olacaktır.
Rodopsin in molekül ağırlığı 40 kDa'dır. Bağlaçdaki deterjan kütlesi nedeniyle SEC'deki molekül ağırlığı yaklaşık 120 kDa'dır. Bu nedenle mini-Go (24 kDa) bağlanmasının SEC'de kolayca tespit edilememesi şaşırtıcı değildir, çünkü bu proteinlerin belirgin kütleleri 120 kDa ve 144 kDa olan farklılaşmasını gerektirecektir. Bu nedenle SDS-PAGE ile SEC fraksiyonlarının analizi örnek saflığı ve karmaşık oluşumu doğrulamak için kullanılmıştır. SEC profilleri karmaşık oluşum üzerinde net bir kayma gösterse bile, karmaşık oluşumu diğer saflaştırılmış protein kirleticileri yerine doğru bağlama ortakları ile doğrulamak için SDS-PAGE analizinin gerçekleştirilmesi önerilir.
Hem rodopsin hem de mini-Go miligram miktarlarda saflaştırılmıştır, bu da sds-PAGE jellerinin SEC ve Commassie Blue boyama sı sırasında UV-VIS emilimi gibi komplekslerin düşük duyarlılık algılamasının kullanılmasını sağlamıştır. Numunelerin sınırlı olduğu durumlarda, proteinden gelen triptofan sinyallerini (280 nm uyarma, 350 nm emisyon) izlemek için floresan dedektörü ile donatılmış lc temizleyici ve SDS-PAGE jelleri için gümüş boyama gibi daha hassas algılama kullanılmalıdır. Başka bir yaklaşım bir floresan protein sigorta olacaktır, yeşil floresan protein gibi (GFP) ilgi protein, aynı zamanda protein ekspresyonu sırasında bile algılama sağlayacak26 ama kristalizasyon önce kaldırılmalıdır.
Saflaştırılmış proteinin değişken TBM'lerden kaynaklanan heterojenlikten de arındırılmış olmasını sağlamak esastır. Burada açıklanan durumda, SDS-PAGE jellerinde gözlenen iki rodopsin popülasyonu bir veya iki N-glikana sahip olarak karakterize edildi. Bir proteinin değişken modifikasyonu potansiyel olarak iyi dağılan kristallerin oluşumunu önler, bu nedenle rodopsinde deglikozilite. Endoglycosidase Endo F1 en etkili endoglycosidase test edildi ve tedavi unglycosylated reseptör tek bir tür yol açtı, PNGase F sadece kısmen rodopsin üzerinde glikan kaldırıldı ve rodopsin karışımı ile sonuçlanan tamamen unglycosileted veya bir N-glikan kaldı. Deglikozilaz tedavisi olmadan rodopin başarıyla kristalize olmuştur3,27,28, rodopsin Asn15 üzerinde N-glikan bu durumlarda kristal temas oluşturmak için önemlidir. Rodopsin-mini-Godurumunda, kristalleri elde etmek için Endo F1 ile N-glikanların çıkarılması gereklidir. Kristalizasyon dan önce ilgi deglikozilat proteinleri için standart bir kural yoktur, ancak proteinler kapsamlı kristalizasyon denemeleri sonra kristalize başarısız olduğunda heterojen PTM'lerin kaldırılması düşünülmelidir.
Burada açıklanan veri ve metodoloji, küçük micelle boyutu ve kompleksi stabilize etme yeteneği nedeniyle rodopsin-mini-Go kompleksinin kristalizasyonu için en çok tercih edilen deterjan olarak OGNG'yi seçmemize rehberlik etti. Ayrıca arıtılmış rodopsinin homojen bir tür olduğundan emin olmak için Endo F1'i kullandık. Kristaller daha sonra elde edildi ve biz ~ 3.1 ş4kristal yapısını belirledi , hangi bir GPCR-G protein sinyal kompleksi sadece üçüncü kristal yapısı oldu14,29.
Ortak proteinle bağlanan ve olmayan membran proteinleri için iki farklı protein olarak düşünülmelidir. Farklı fonksiyonel hallerde bir protein farklı konformasyonlara sahiptir ve farklı enerji düzeyindedir. Bu nedenle, etkin olmayan durum için parametre etkinleştirilen duruma tam olarak aktarılamaz gibi her işlevsel durum için hazırlama protokolü optimize etmek önerilir. Ayrıca, bir ortak protein bağlanarak karmaşık protein özelliği değişikliği söz değil. Protokol, farklı deterjanlarda inaktif membran proteini hazırlamak, ardından protein aktivasyonu ve kompleks oluşumunu sağlamak ve protein kalitesini karakterize etmek için kristalizasyon numunesi hazırlamak için standartlaştırılmış yöntemler kullanır. Böylece, bu protokol kolayca diğer membran proteinleri ve küçük değişiklikler ile yapısal çalışmalar için kompleksleri için genelleştirilmiş olabilir.
Açıklamalar
CGT, Sosei Heptares'in Bilimsel Danışma Kurulu'nun danışmanı ve üyesidir. Diğer tüm yazarların açıklayacak bir şeyi yok.
Teşekkürler
Prof. Dr. Gebhard F. X. Schertler'a bu projedeki uzun süreli desteği için teşekkür ederiz, Dr. Roger J.P. Dawson ve Hoffmann La Roche hücre kültürüne destek için. Bu çalışma İsviçre Ulusal Bilim Vakfı (210030_153145 ve 310030B_173335 GFXS'e hibe ler) ve Avrupa Araştırma Konseyi (EMPSI, 339995) ve Tıbbi Araştırma Konseyi'nden (MRC U105197215) CGT'ye fon sağlamak la desteklenmiştir. FP, ETH Zürich'i Ulusal Araştırma Yetkinlik Merkezi Moleküler Ultrafast Bilim ve Teknoloji (NCCR MUST) ve ETH Femtosecond ve Attosecond Science and Technology (ETH FAST) programları aracılığıyla kabul eder. FP, JM, AB ve CJT Paul Scherrer Enstitüsü'nden uzun vadeli mali destek kabul.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 1D4 peptide | Peptide2.0 | Under request | |
| 9-cis retinal | Sigma-Aldrich | R5754 | |
| Autosampler A-900 | GE Healthcare | Discontinued | |
| C9G | Anatrace | N324 | |
| cOmplete, EDTA-free protease inhibitor coctail | Roche | 5056489001 | |
| Cymal-5 | Anatrace | C325 | |
| Cymal-5NG | Anatrace | NG325 | |
| Cymal-6 | Anatrace | C326 | |
| Cymal-6NG | Anatrace | NG326 | |
| DDM | Anatrace | D310 | |
| DM | Anatrace | D322 | |
| DMNG | Anatrace | NG322 | |
| Econo column | Bio-Rad | 7372512 | |
| Ettan LC | GE Healthcare | Discontinued | |
| FRAC-950 | GE Healthcare | Discontinued | |
| HPLC Water 2795 Separation Module | Waters AG | 720000358EN | |
| InstantBlue Protein Stain | Expedeon | ISB1L | |
| LCT Premier mass spectrometer (ESI-TOF) | Waters AG | - | |
| LMNG | Anatrace | NG310 | |
| Monitor UV-900 | GE Healthcare | 18110835 | |
| Nanodrop 1000 | Witec AG/ThermoFisher | Discontinued | |
| NuPAGE 4-12% Bis-Tris gel 1.0 mm, 15 well | ThermoFisher | NP0323BOX | |
| NuPAGE MES SDS buffer (20x) | ThermoFisher | NP0002 | |
| OGNG | Anatrace | NG311 | |
| PAGEr Minigel Chamber | Lonza | 59905 | |
| Reprosil 200 C18-AQ column | Morvay Analytik GmbH | #s1503 | |
| Superdex 200 Increase GL column | GE Healthcare | 28990944 | |
| Tabletop centrifuge 5424R | Eppendorf | 5404000413 | |
| Ultracentrifuge Optima XE-100 | Beckmann Coulter | A94516 | |
| ULTRA-TURRAX T25 | IKA WERKE | 0003725003 | |
| UV-VIS spectrophotometer | Shimadzu | UV-2401PC | |
| Waters 2487 Dual λ Absorbance Detector | Waters AG | - |
Referanslar
- Tate, C. G. Practical considerations of membrane protein instability during purification and crystallisation. Methods in Molecular Biology. 601, 187-203 (2010).
- Lebon, G., Bennett, K., Jazayeri, A., Tate, C. G. Thermostabilisation of an agonist-bound conformation of the human adenosine A(2A) receptor. Journal of Molecular Biology. 409 (3), 298-310 (2011).
- Deupi, X., et al. Stabilized G protein binding site in the structure of constitutively active metarhodopsin-II. Proceedings of the National Academy of Sciences. 109 (1), 119-124 (2012).
- Tsai, C. -. J., et al. Crystal structure of rhodopsin in complex with a mini-G o sheds light on the principles of G protein selectivity. Science Advances. 4 (9), (2018).
- Carpenter, B., Tate, C. G. Engineering a minimal G protein to facilitate crystallisation of G protein-coupled receptors in their active conformation. Protein Engineering Design and Selection. 29 (12), 583-594 (2016).
- Chae, P. S., et al. Maltose-neopentyl glycol (MNG) amphiphiles for solubilization, stabilization and crystallization of membrane proteins. Nature Methods. 7 (12), 1003-1008 (2010).
- Loll, P. J. Membrane proteins, detergents and crystals: what is the state of the art. Acta Crystallographica Section F Structural Biology Communications. 70 (12), 1576-1583 (2014).
- Chae, P. S., et al. Glucose-neopentyl glycol (GNG) amphiphiles for membrane protein study. Chemical communications. 49 (23), 2287-2289 (2013).
- Standfuss, J., Xie, G., Edwards, P. C., Burghammer, M., Oprian, D. D., Schertler, G. F. X. Crystal structure of a thermally stable rhodopsin mutant. Journal of Molecular Biology. 372 (5), 1179-1188 (2007).
- Kaushal, S., Ridge, K. D., Khorana, H. G. Structure and function in rhodopsin: the role of asparagine-linked glycosylation. Proceedings of the National Academy of Sciences of the United States of America. 91 (9), 4024-4028 (1994).
- Molday, L. L., Molday, R. S. 1D4: a versatile epitope tag for the purification and characterization of expressed membrane and soluble proteins. Methods in Molecular Biology. 1177 (604), 1-15 (2014).
- Carpenter, B., Tate, C. G. Expression and Purification of Mini G Proteins from Escherichia coli. Bio-Protocol. 7 (8), (2017).
- Grueninger-Leitch, F., D'Arcy, A., D'Arcy, B., Chène, C. Deglycosylation of proteins for crystallization using recombinant fusion protein glycosidases. Protein Science. 5 (12), 2617-2622 (1996).
- Rasmussen, S. G. F., et al. Crystal structure of the β2 adrenergic receptor-Gs protein complex. Nature. 477 (7366), 549-555 (2011).
- Loginova, M. Y., Rostovtseva, Y. V., Feldman, T. B., Ostrovsky, M. A. Light damaging action of all-trans-retinal and its derivatives on rhodopsin molecules in the photoreceptor membrane. Biochemistry (Moscow). 73 (2), 130-138 (2008).
- Alexandrov, A. I., Mileni, M., Chien, E. Y. T., Hanson, M. A., Stevens, R. C. Microscale Fluorescent Thermal Stability Assay for Membrane Proteins. Structure. 16 (3), 351-359 (2008).
- Sonoda, Y., et al. Benchmarking Membrane Protein Detergent Stability for Improving Throughput of High-Resolution X-ray Structures. Structure. 19 (1), 17-25 (2011).
- Maeda, S., et al. Crystallization scale preparation of a stable GPCR signaling complex between constitutively active rhodopsin and G-protein. PloS One. 9 (6), 98714 (2014).
- Boute, N., Jockers, R., Issad, T. The use of resonance energy transfer in high-throughput screening: BRET versus FRET. Trends in Pharmacological Sciences. 23 (8), 351-354 (2002).
- Singhal, A., Guo, Y., Matkovic, M., Schertler, G., Deupi, X., Yan, E. C. Y. Structural role of the T 94 I rhodopsin mutation in congenital stationary night blindness. EMBO Report. 17 (10), 1-10 (2016).
- Choe, H. -. W., et al. Crystal structure of metarhodopsin II. Nature. 471 (7340), 651-655 (2011).
- Mattle, D., et al. Ligand channel in pharmacologically stabilized rhodopsin. Proceedings of the National Academy of Sciences of the United States of America. 115 (14), 3640-3645 (2018).
- Okada, T., Fujiyoshi, Y., Silow, M., Navarro, J., Landau, E. M., Shichida, Y. Functional role of internal water molecules in rhodopsin revealed by X-ray crystallography. Proceedings of the National Academy of Sciences of the United States of America. 99 (9), 5982-5987 (2002).
- Blankenship, E., Vahedi-Faridi, A., Lodowski, D. T. The High-Resolution Structure of Activated Opsin Reveals a Conserved Solvent Network in the Transmembrane Region Essential for Activation. Structure. 23 (12), 2358-2364 (2015).
- Magnani, F., et al. A mutagenesis and screening strategy to generate optimally thermostabilized membrane proteins for structural studies. Nature Protocols. 11 (8), 1554-1571 (2016).
- Kawate, T., Gouaux, E. Fluorescence-detection size-exclusion chromatography for precrystallization screening of integral membrane proteins. Structure. 14 (4), 673-681 (2006).
- Standfuss, J., et al. The structural basis of agonist-induced activation in constitutively active rhodopsin. Nature. 471 (7340), 656-660 (2011).
- Singhal, A., et al. Insights into congenital stationary night blindness based on the structure of G90D rhodopsin. EMBO reports. 14 (6), 520-526 (2013).
- Carpenter, B., Nehmé, R., Warne, T., Leslie, A. G. W., Tate, C. G. Structure of the adenosine A(2A) receptor bound to an engineered G protein. Nature. 536 (7614), 104-107 (2016).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır