
Method Article
פקיעה זמן הדמיה של העכבר מקרופאג כימוטקסיס
In This Article
Summary
כאן אנו מתארים שיטות באמצעות שעות הזמן, מיקרוסקופ משלב ניגודיות לעכבר התמונה מקרופאגים הצפק בתוך כימוטקטקטיקה המשלים C5a הדרגתי. ניתן להרחיב את הפרוטוקולים לתאים חיסוניים אחרים.
Abstract
כימוטקסיס היא קולטן בתיווך הדרכה של תאים לאורך הדרגתי כימי, בעוד chemokinesis הוא גירוי של התאים האקראיים לתנועתיות על ידי כימיקל. Chemokinesis וכימוטקוניות הן בסיסיות לגיוס ופריסה של תאים חיסוניים. לדוגמה, נוגדנים (ציטוקינים כימוטקטיק) יכול במהירות לגייס נויטרופילים ומונוציטים לextravascular אתרים של דלקת. קולטני כימוסטנט שייכים המשפחה הגדולה של G חלבון מצמידים קולטנים. כיצד כימוזסטנט (כלומר, ligand) מעברי צבע ישיר הגירה להעברת תאים באמצעות חלבון G מצמידים איתות עדיין לא מובן לגמרי. בתחום של אימונולוגיה, נויטרופילים הם תאים מודל פופולרי עבור לימוד כימוטקוניות ב מבחנה. כאן אנו מתארים בזמן אמת דו מימדי (2D) שיטת המיקרופאגים המותאמים עבור העכבר, אשר באופן מסורתי היה קשה יותר ללמוד. מקרופאגים לנוע בקצב איטי של ~ 1 μm/min על פני שטח דו-ממדי ומתאימים פחות להעברת נקודת מקור הגירה (למשל, הגירה לקראת קצה של מיקרופיפטה מלא עם כימוג) מ נויטרופילים או Dictyelium discoideum, אשר להעביר סדר גודל מהר יותר. בשימוש נרחב transwell בחני שימושי לימוד פעילות כימוטקטיק של חומרים שונים, אבל לא לספק מידע על מורפולוגיה התא, מהירות, או ניווט כימוטקטיק. כאן אנו מתארים את הזמן לשגות מיקרוסקופית מבוססי מקרופאג המערכת המבוססת על שיטה המאפשרת כימות של מהירות התא ויעילות כימוטקטיים מספק פלטפורמה כדי להתוות את הסימנים, מסלולים האות, ואת העריקים של כימווניות.
Introduction
התאים החיסוניים בדרך כלל להעביר ביחידים על משטח דו-ממדי בצורה אמבות1,2, אשר כרוך חוזרות ונשנות של בליטה של החזית, הדבקה של תא בתיווך, ונסיגה של האחורי. שלב התנאי הראשון הוא קיטוב התא, שבו תאים הטופס הקדמי והאחורי מסתיים3. כימוטקוניות מתחיל עם זיהוי של כימומי על ידי G חלבון מצמידים קולטנים ורשת איתות מורכבת מתווכת על-ידי ממברנה מעוגן חלבונים heterotrimeric g וחלבונים monomeric קטנים, כמו גם פוספוליפיד-מאוגד החליפין גואנין גורמים (gefs)4,5. הפעלה של רו gtpases של משפחות מCdc42 ומירוץ מ, לגרום בליטות בחזית6 וחברים של משפחת Rho, במיוחד rhoa, להפעיל את התכווצות של האחורי5,7. בסביבה תלת-ממדית (3d), אינטגרציה מיותרת במידה רבה להעברת לוקיציט ו-rhoa הופכת להיות חשובה יותר עבור לחיצת תאים באמצעות מעברים צרים8, בעוד Cdc42-או מירוץ המושרה Arp2/3 נשאר חשוב עבור היגוי כימוטקטיק9,10.
תאים חיסוניים עשויים להתמודד עם כימומי שונים, במיוחד בהגדרות של פציעה ברקמות, הפלישה הפתוגן, ודלקת. כימודוגני חומרים מתבטא על phagocytes משלימים C3a ו C5a, מופקים במהירות על ידי הפעלת המפל המשלים, ו מזוהים על ידי C3a משלימים וקולטני C5a. באופן דומה, התאים נמק לגייס phagocytes באמצעות קולטני פפטיד formyl, אשר לזהות המיטוסטיים, כמו גם חיידקים נגזרות formyl בקטריה11. התאים החיסוניים גם לבטא G חלבון מצמידים קולטנים עבור נוגדנים, משפחה גדולה של פפטידים כימוג מעורב בוויסות של סחר בתאים חיסוניים במהלך הומאוסטזיס ודלקת. נוגדנים מסווגים ארבע קבוצות בהתאם לריווח של שני ציסטאין הראשון (c) שאריות: c, CC, cxc, ו CX3C ציטוקינים, שם X הוא חומצת אמינו. כך, בתאי החיסון vivo צריך להגיב כראוי מורכבים מאוד אותות הרקתית הזמני, מה שהופך את המחקר של כימומוניות משימה מרתיעה. בהמשך אנו מספקים היסטוריה קצרה של כימוטקוניות, שהחלה בגישות להדמיה באמצעות הדמיה.
המחקר של כימוקוציטים מוניות שתחילתה בשנת 188812, כאשר לרופא עיניים תיאודור קארל גוסטב לייבר תיאר בבירור את הגירה מכוונת של לוקיציטים, ואת הצטברות, אתרים של דלקת במודל של פטרת (פטרייתי) דלקת. לייבר הדגיש כי המשיכה של עודף לוקיציטים על ידי גורמים הפתוגן-נגזר חשוב לחיסול של מיקרואורגניזמים מזיקים דרך phagocyציטוזה, אשר תוארה על ידי Metchnikoff (ידוע גם בשם Metchnikoff) מוקדם יותר באותו עשור13. בניסויים vivo נערכו גם בשנות ה -20 על ידי קלארק וקלארק14,15, אשר ניצל את השקיפות של טתרנים והראו כי דלקת סטרילית הנגרמת על ידי שמן קרוטון14 או אחרים משמנים15 גרימת לוקיציטים כדי לדבוק בכלי הדם, ואחריו החתלה (הגירה) והגירה מהירה דרך מרחבי רקמות לעבר הגירוי. בניסויים מחוץ למבחנה בשיטת המיקרוצילום שפותחה על ידי ז'אן קומנדון16 הראו כי הלוקוציטים הועברו לעבר מקור כימוסטנט חלקיקי כגון חיידקים17. באותה תקופה, הזהויות המולקולריות של גורמי כימוטקטיים לא היו ידועות. בשנות ה-60, סטיבן בוידן18 זיהה שטכניקות ללימוד פעילות כימוטקטיק של חומרים מסיסים היו חסרים. הוא המציא תא, הידוע לאחר מכן כחדר בוידן, ובו שני תאים המופרדים באמצעות קרום נייר מסנן. השעיית תא מתווסף לתא העליון וחומר הבדיקה מתווסף לשני התאים או רק לתא התחתון. לאחר תקופת דגירה, קרום המסנן מוסר, והתאים קבועים ומוכתמים. על-ידי השוואת מספר התאים העוברים את קרום המסנן לעבר הבאר הנמוכה יותר עם חומר המבחן בשני התאים, לא בתא, או רק בתא התחתון, ניתן לקבוע את פעילות הפעולה הכוללת כימוטקטיק. Transwell באסאומר עדיין פופולרי היום השתנו בדרכים שונות, כולל שימוש של ממברנות פוליקרבונט שונים עם גדלי נקבובית מוגדר צפיפויות19,20. החיסרון העיקרי של transwell בחני אומר הוא שזה לא מעשי להמחיש ישירות התאים הגירה הנתיב הגירה על פני הקרום בדרך כלל לא עולה על קוטר של תא החיסון.
סלי ה. Zigmond פיתחה תא כימוטווניות21 המאפשרים הדמיה של היווצרות הן מעבר הדרגתי והן מורפולוגיה התא באמצעות צבעי פלורסנט. החדר מורכב שקופית פלקסיגלס (אקריליק) עם שתי בארות לינאריות מקבילות, כל אחד עם נפח של ~ 100 μl, מופרדים על-ידי גשר רחב 1 מ"מ 3 – 10 יקרומטר מתחת למישור העליון של השקופית. כיסוי שנזרע עם תאים הוא הפוך וממוקם על השקופית כך שהיא משתרעת על שתי בארות. לאחר הוספת כימוסטנט לאחת הבארות, מעבר בין צורות מילוי משני של הגשר, בדרך כלל בטווח של 30 עד 90 דקות. הפולימולוציטים האנושיות (גרנולוציטים) בחדר זימונד מתבוננים בעצמם כלפי כימוסטנט. וריאציות של תא Zigmond דווחו, כולל דאן22 ו insall23 צ'יימברס, שניהם להשתמש שמיכות הזרע עם תאים הממוקמים על פני שתי בארות מופרדים על ידי גשר רחב 1 מ '. תא דאן מורכב מבארות קונצנטריות המופרדות על-ידי גשר עגול, בעוד שחדר האינסל קרוב יותר לאולם זימונד, אך מספק גשרים של שני רוחבי שונים, 0.5 מ"מ ו-1 מ"מ. תא כימוטווניות מספר, כינה את μ-Slide-מוניות ומיוצר על ידי הזרקת פלסטיק, תוארה על ידי זמג'ל ואח '24. תא כימוטווניות כולל מאגרים מ2 40 μL מופרדים על-ידי ערוץ רחב מאוד מ-1 עם אורך של 2 מ"מ וגובה של 70 μm. החלק התחתון של החדר נוצר על ידי חדירות גז, גיליון פלסטיק דק עם עובי זהה ותכונות אופטיות של No. 1.5 זכוכית מחליק24. כאן אנו מתארים שיטת כימוטווניות באמצעות החדר הכימוקווניות של μ-שקופיות כדי להמחיש את הגירה של מקרופאגים הצפק של העכבר למשך עד 14 שעות (המשלים C5a) הדרגתי.
Protocol
הפרוטוקולים בעקבות ההנחיות של ועדת האתיקה של המחקר המקומי, כמו גם הנחיות לטיפול בבעלי חיים.
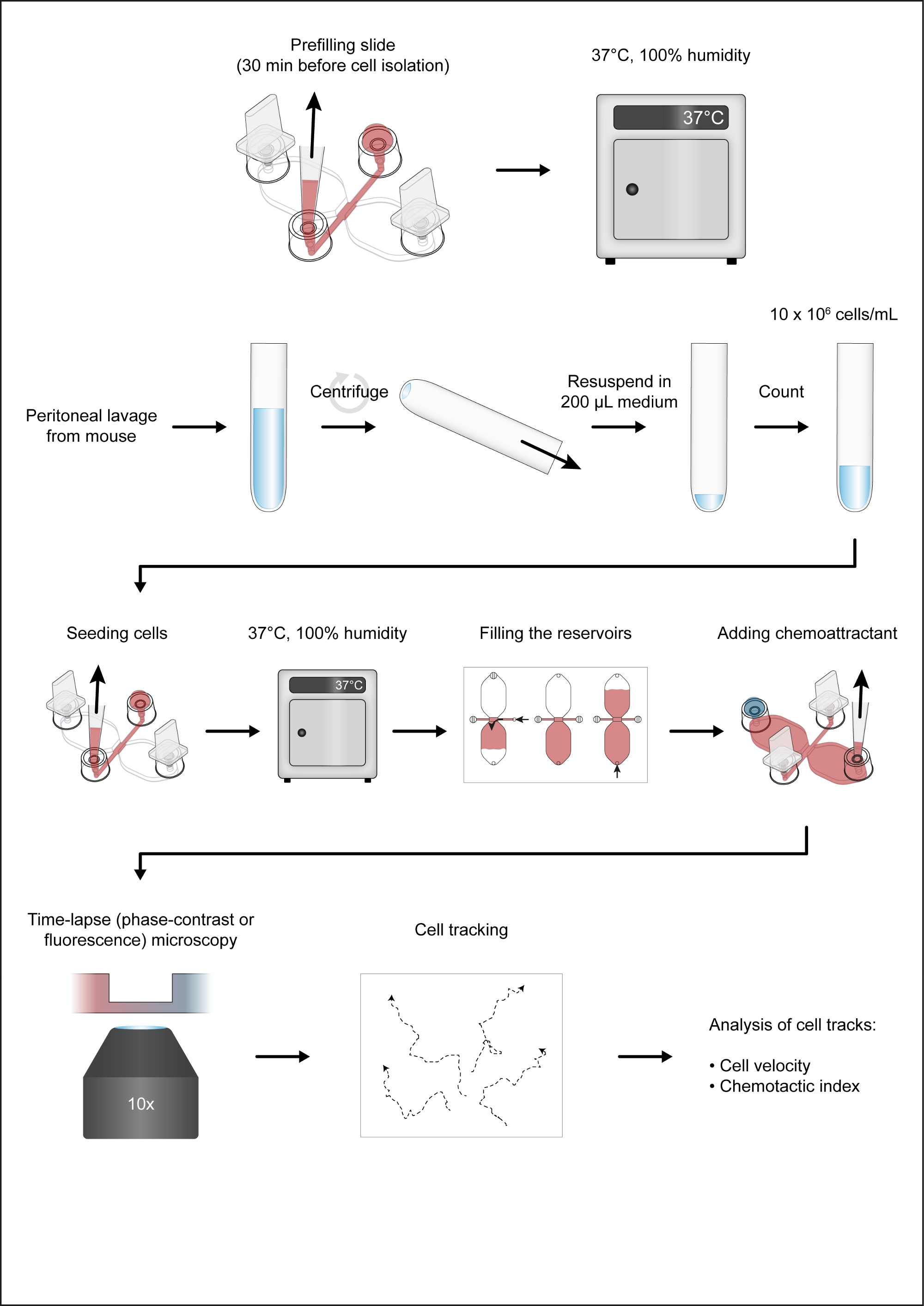
הערה: איור 1 הצגת זרימת עבודה של שיטת כימוטקסיס.
1. מילוי מראש כימוטקסיס שקופיות
-
מילוי מראש 1 מ"מ רוחב ו 2 מילימטר ערוצים החיבור של אחד או שניים שקופיות כימוטווניות באמצעות שונה RPMI 1640 hepes בינונית, המורכב ביקרבונט-free RPMI 1640 בינונית המכילה 20 מ"מ 4-(2-הידרוקסיל) -1-piperazinefonic חומצה (hepes), 10% חום בלתי מופעל העובר סרום (fbs), ואנטיביוטיקה כגון פניצילין (100 U/mL) ו סטרפטומיצין (100 μg/ml), הכין על ידי דילול 100x פניצילין/סטרפטומיצין, ו-1 μg/ml ליפופוליד (מ -E. coli), וקולטן כמו אגרה 4 ליגנד משמש כדי להפעיל את התאים.
- מניחים שקופית כימוכוניות (איור 2 א) לתוך מיכל תרבות התאים העגול (10 ס מ), שניהם מחוממים עד 37 ° c, ומניחים את המנה על גוש אלומיניום מחומם (37 ° c). הכנס תקעים ליציאות 1 ו-4 (איור 2B).
הערה: בלוק אלומיניום השמור ב-37 ° c ומוצב בתוך כיסוי הזרימה הלבינארי שימושי להכנת תאי כימוטקסיס. באופן אידיאלי, בלוק מחומם צריך לספק שטח עבודה שטוח ובארות עבור צינורות שונים, כגון 50 מ"ל צינורות ו 2 mL מיקרוצנטריפוגה צינורות. - באמצעות טיפ של 10 – 200 μL עם טיפ משופע, הפקדה 15 μL של שונה RPMI 1640 HEPES בינונית לתוך מילוי היציאה 3 (איור 2B). הבא, עם עוצמת הקול עדיין להגדיר 15 μL ולחצן הבקרה של (2 – 20 μL) פיפטה מדוכא, להכניס את העצה הפיפטה לתוך היציאה 2 ו לאכול 15 μL בקצב מהיר בינוני (איור 2B). זה ימלא מראש את הערוץ המחבר 1 מ"מ x 2 מ"מ (אזור תצפית), כמו גם את שני ערוצי האספקה האגפים (בין אזור התצפית המרכזי לבין יציאות 2 ו-3, בהתאמה). כיסוי מילוי יציאות 2 ו 3 עם כמוסות.
- לאחר המילוי מראש, הניחו את השיטווניות לארון תקשורת השמור בחדר לחות סגור בתוך אינקובטור יבשושני 2-3ב-37 ° c.
הערה: חשוב להשתמש בעצת הפיפטה הנכונה למילוי שקופית כימוטקסיס. לחלק העליון של הנמל המלא בצנרת משופעת, בעוד שבדרך כלל ניתן להוסיף עצות מחודדות לצינורות האוורור ולהגדיל את העמידות באופן משמעותי לזרימת נוזלים.
- מניחים שקופית כימוכוניות (איור 2 א) לתוך מיכל תרבות התאים העגול (10 ס מ), שניהם מחוממים עד 37 ° c, ומניחים את המנה על גוש אלומיניום מחומם (37 ° c). הכנס תקעים ליציאות 1 ו-4 (איור 2B).
2. בידוד של העכבר תושב הצפק מקרופאגים
- להקריב את 3 – 4 החודש עכבר בן באמצעות ריכוז גבוה של isof, הרדמה נדיפים (> 5% באוויר) או פחמן דו חמצני25, ואחריו פריקה צוואר הרחם. אובדן רפלקס הצתה במכרסמים התואם לאובדן התודעה בבני אדם26. נקה את הבטן של העכבר עם 80% אתנול במים ולאחר מכן לעשות 1 – 2 ס מ העור באמצע החתך באמצעות מספריים כירורגי עם טיפים קהה. לקלף את העור בחזרה כדי לחשוף את קיר הבטן המשמש כבסיס.
- הכניסו צנתר פלסטיק באורך. של 3 גרם לחלל הצפק באמצעות מזרק פלסטיק 5 מ ל, שאיפה את החלל באמצעות 2 x 4.5 mL הקרח הקרה באגירה של האנק פתרון מלח (HBSS), ללא Ca2 + ו-Mg2 +. להשאיר סביב 0.5 mL של HBSS שיורית במזרק כך רקמת למצוץ בשוגג על קצה הקטטר ניתן לגרש.
- להעביר את המדיום הלגיל, בדרך כלל 8 – 8.5 mL בסך הכל, לתוך הצינור התחתון 14 מ ל פוליפרופילן עגול. צנטריפוגה את הצינור ב 300 x g עבור 6.5 דקות בטמפרטורת החדר.
הערה: הצינור התחתון העגול מאפשר לסופרנטאנט להיות מלא ומפחית את מיזוג התאים. - להיפטר supernatant ולהשעות מחדש את התאים הצפק (בדרך כלל ~ 4 x 106 תאים לכל עכבר) ב 200 μl של שונה RPMI 1640 hepes בינונית. דלל מדגם של השעיית התא 1:20 והשתמש בהתקן ספירה, כגון תא ספירה משופר של נויבאואר, כדי לספור את התאים. לאחר מכן, לדלל את התא ההשעיה לריכוז הסופי של 10 x 106 תאים/mL ולשמור על התאים בתחתית עגול 2 מ ל פוליפרופילן מיקרוצנטריפוגה צינור ב 37 ° צ' באמצעות בלוק אלומיניום מחומם (ראה הערה בשלב 1.1.1).
3. זריעת תאים הצפק לתוך כימוטקסיס שקופיות
- לאחר ללטף את התא הבולם למעלה ולמטה 5x עם נפח הפיפטה להגדיר ב 100 μL (או במחצית הכרך ההשעיה) כדי להפחית החלקה, בעדינות הפקדה 10 μL של התא הבולם לתוך היציאה 3 של תא כימוטווניות (איור 2C). מניחים את קצה הפיפטה ליציאה 2 ומושכים לאט את המתלה לתא לערוץ המחבר (איור 2C). ברגע השעיית התא הוצג, להסיר את התקעים ביציאות 1 ו-4, אשר יסייעו לעצור את הזרימה של ההשעיה התא. הצב כובעים בכל ארבעת היציאות המלאות.
- חזור על שלב 3.1 עבור כל תאי כימוטקסיס. באמצעות מיקרוסקופ הפוך קטן העדשה 10x המטרה הניגוד האובייקטיבי, לבדוק את שקופיות כימוטקוניות עבור בועות אוויר לא רצויות.
- מניחים את מגלשות השיטווניות בתאי הצפק בחדר לחות ב-37 ° c עבור 2 – 3 h.
4. מילוי המאגרים והוספת כימוסטנט
- בדוק את אזור התצפית (הערוץ המחבר את מאגרי μL 2 40) באמצעות מיקרוסקופ הפוך.
הערה: בשלב זה, צפיפות התא תהיה גבוהה יותר מאשר לאחר מילוי המאגרים, כי תאים מחסיד חלש, בעיקר CD19+ תאים (בתאי B1), יהיה שטף מתוך אזור התצפית במהלך ההליך מילוי (איור 2C – E). - מניחים את התקעים למילוי יציאות 1 ו-2 (איור 2D). ודא כי מילוי היציאה 3 מתמלא למעלה עם בינונית וחופשית של בועות אוויר. השתמש סטרילי 27 גרם מזרק מחט להוציא בועות אוויר לא רצויות, אם נדרש.
- באמצעות 10 – 100 μL בנפח מכני הכרך, ומתה ~ 60 μL של שונה RPMI 1640 HEPES בינונית ומניחים את הקצה הפיפטה לתוך מילוי הנמל 3. השתמש בטבעת הגדרת העוצמה של הפיפטה כדי להזריק לאט ובעקביות לתוך המאגר כגון המדיום מגיע לראש מילוי הנמל 4 אחרי 1 – 2 דקות (איור 2D).
- . תמלא את המאגר השני הזז את התקע מהיציאה 1 והכנס אותו לאט ליציאה 3 (איור 2D). הבא, ומלא לאכול ~ 50 μL של שונה RPMI 1640 HEPES בינוני ולמקם את העצה הפיפטה לתוך מילוי יציאה 4. השתמש בטבעת הגדרת עוצמת הקול של 10 – 100 הצינורות הנפח μL כדי להכניס לאט ובהתמדה להזריק בינוני לתוך המאגר השני כגון בינוני מגיע לראש היציאה מילוי 1 אחרי 1 – 2 דקות (איור 2D).
- מקום 495 μL של שונה RPMI 1640 HEPES בינונית לתוך (בחלק התחתון) 2 מ"ל שפופרת מיקרוצנטריפוגה ולהוסיף 5 μL של פטנט כחול V (פתרון מניות: 10 מ"ג/mL ב פוספט מאגר מלוחים [PBS]), צבע כחול המשמש כאינדיקטור חזותי של היווצרות מעבר הצבע. מערבבים על ידי הורטקנג קצר. הוסף 5.4 μL של רקומביננטי העכבר המשלים C5a (פתרון מניות: 50 μg/mL ב-PBS עם 0.1% בסרום פרה אלבומין) ולערבב על ידי וורטקנג קצר.
- הפקדה 15 μL של כחול, המשלים C5a-המכיל בינוני לתוך מילוי יציאה 1 (איור 3A), לאחר לוודא כי הדיכאון הרדוד בחלק העליון של הנמל הוא בינוני ללא תשלום (אחרת הירידה עלולה להפוך לגולש).
- הוסף 10 – 200 μL לתוך מילוי היציאה 4 לאט ובהתמדה לסובב את הטבעת הגדרת עוצמת הקול של 10-100 μL הכונן האמצעי כדי לצייר את טיפת כחול, המשלים C5a-המכיל בינוני לתוך מאגר הפוך (איור 3B). האוויר יתחיל להיכנס לטור האנכי הקצר של מילוי היציאה 1. משוך את האוויר עד שממשק האוויר הנוזלי יהיה באמצע המעבר בעמודה האנכית ולאחר מכן הכנס באיטיות תקע לתוך היציאה.
- הרם בעדינות את הפיפטה מפורט 4, תוך שימוש בעומת זאת כדי להבטיח שהשקופית תישאר קבועה במקומה. לבסוף, תקע באיטיות יציאה 4 (איור 3B).
- בדוק את שקופית כימוטקסיס במיקרוסקופ הפוך.
הערה: התאים חסיד שנותרו באזור התצפית צריך להיות בעיקר מקרופאגים. זה יכול להיות מאושר באמצעות מתויג פלואורוסקופים anti-F4/80 נוגדנים (F4/80 הוא סמן ספציפי עבור מקרופאגים העכבר). B תאים ניתן לזהות באמצעות פלואור מתויג anti-CD19 נוגדנים F4/80-/cd19- תאים ניתן להבחין באמצעות כתם כחול גרעין חומצה פלואורסצנטי (איור 4).
5. הדמיה מקרופאג הגירה על ידי זמן-פקיעה, מיקרוסקופ הניגודיות פאזה
- הניחו שקופית כימוטקסיס על הבמה של מיקרוסקופ הפוך המצויד באינקובטור במה. שמרו על הטמפרטורה ב-37 ° c.
- תמונה באזור תצפית 1 מ"מ x 2 מ"מ באמצעות עדשת מטרה 10x-ניגודיות המטרה ולהתמקד מקרופאג lamellipodia: דק, גיליון כמו בליטות קרום. לכידת תמונות עבור 14 שעות בקצב של מסגרת 1 כל 2 דקות.
6. ניתוח הזמן-תמונות פקיעה
- מנתחים את התמונות החדות והפאזה באמצעות תוכנת ניתוח תמונה אוטומטית או תוסף המעקב הידני, המיוצר על ידי Fabrice P. קורדלירס, עבור ImageJ.
הערה: ניתן להשתמש בתוכניות מעקב אוטומטיות כדי לנתח את התאים הניתנים לדימות על-ידי שימוש בזמן הקפיצה, הפאזה, או מיקרוסקופ הקרינה. לדוגמה, התוכנה המבוססת על Java iTrack4U המיוצר על ידי Cordelières ואח '27 ניתן להשתמש עבור מעקב אוטומטי של תאים וניתוח באמצעות זמן הפקיעה, שלב-ניגודיות, או תמונות זריחה כקלט. מעקב ידני הוא זמן רב יותר, אך הרצועות שנוצרו על-ידי מעקב הידני imagej plugin יכול להיות מיובא ישירות באופן אוטומטי על ידי התוסף imagej כימוטקוניות וכלי הגירה28,29.
תוצאות
תרשים סכמטי של השקופית כימוקווניות המשמש לפקיעה זמן וידאו מיקרוסקופ של העכבר מקרופאגים להעברת הצפק בהדרגה מעבר כימוטקטקטיקה מוצג באיור 2A. השקופית מכילה שלושה תאי כימוטקא, שלכל אחד מהם ארבע יציאות מילוי. ניתן לסגור את היציאות בנפרד באמצעות התקעים המוצגים מעל השקופית. לחילופין, מכסה שאינו איטום ניתן להציב על גבי נמל מנותק כדי לשמור על עקרות. לאחר חיבור יציאות 1 ו-4, אזור התצפית (1 מ"מ רוחב x 2 מ"מ x 70 יקרומטר ערוץ גבוה המחבר את שני המאגרים) בין יציאות 2 ו-3 יכול להיות מלאה במדיום על ידי הצבת 15 μl ירידה לתוך היציאה 3 ו משתעלים עם 2 – 20 μl היקף הצינור ביציאה 2 (איור 2b) השעיה של העכבר תאים הצפק בתאי (10 x 106 תאים/mL) הופרה לתוך אזור התצפית על ידי הצבת 10 μl ירידה של ההשעיה לתוך היציאה 3 ומוחלק לאט ביציאה 2 (איור 2c). תמונה טיפוסית של תאים שנזרע באזור התצפית נלקח על ידי מיקרוסקופ הפאזה ניגודיות באמצעות עדשת היעד 10x מוצג באיור 2C. לאחר הדגירה עבור 2 – 3 h, שקופית כימוטקיית מלאה לאט עם בינונית (איור 2D). לאחר חיבור יציאות 1 ו-2, בינוני הוזרק לאט באמצעות יציאת 3 עד שהוא יצא מנמל 4. לאחר מכן, התקע הוחלף מיציאה 1 ליציאה 3, ולאחר מכן המאגר השני מילא על ידי הזרקת בינוני לאט באמצעות יציאה 4 עד שהוא יצא ביציאה 1. בשלב זה, התאים באזור התצפית נבדקו מחדש באמצעות מיקרוסקופ הפוך (איור 2E). על ידי השוואת תמונות זמן קצר לפני (איור 2C) ואחרי (איור 2c) מילוי של המאגרים, עד שני שלישים של התאים נשטפו מתוך אזור התצפית. באופן כללי, חלש חסיד CD19+ תאים (בתאי B1) נשטפו החוצה ואת התאים הנותרים היו בעיקר F4/80+ תאים (מקרופאגים). זה הוכח על ידי מיקרוסקופ הקרינה הפלואורסצנטית לאחר תיוג כל סוג תא עם התווית של נוגדנים ספציפיים (איור 4). באיור 4A, מבודד העכבר המבודד תאים הצפק מסומנים עם פלורסנט ירוק fluorophore-מעלה Anti-F4/80 נוגדנים פלורסנט פלואורואופלפור אדום-מצוקים ANTI-CD19 נוגדנים, ואת הגרעינים של תאים היו מסומנים עם כתם כחול הגרעין חומצה פלואורסצנטי. F4/80 הוא סמן ספציפי עבור מקרופאגים העכבר30, בעוד CD19 הוא סמן תא B. איור 4B מציג F4/80+ תאים התמונה על ידי ספינינג מיקרוסקופ הדיסק באזור התצפית של תא כימוטווניות. התאים סומנו לאחר שהיה שיטת כימוא לילה שנרשמה על-ידי שגיאה בזמן, מיקרוסקופ בניגוד לשלב.
המשלים C5a (הכימוסטנט) הוצג אחד משני המאגרים על ידי הצבת 15 μL ירידה של בינונית המכילה 0.54 μg/mL (רקומביננטי mouse) משלימים C5a ו 10 μg/mL פטנט כחול V לתוך מילוי יציאה 1 (איור 3A) לאחר חיבור יציאות 2 ו-3. באמצעות שאיפה איטית עם פיפטה. דרך נמל 4 איור 3B מראה את הדיפוזיה של הצבע הכחול לאחר הציור 15 μl ירידה לתוך מאגר. פטנט כחול V שימש כאינדיקציה ויזואלית עקיפה של הדיפוזיה כימוסטנט. המשלים C5a מולקולות הם גדולים במידה ניכרת מאלה של פטנט כחול V (9.0 kDa לעומת 0.57 kDa) ולפזר לאט יותר. לאחר הדיפוזיה של המשלים C5a במאגר, הריכוז שלה היה ~ 0.2 μg/mL (15 μL/40 μL [נפח אגירה] x 0.54 μg/mL = 0.2 μg/mL), שווה ערך ל ~ 22.5 nM. מעבר תלול תלולה הוקמה על פני אזור התצפית אחרי 3 h והמשיך להגדיל, להגיע למקסימום בסביבות 12 h31. איור 3C מציג את רצועות ההגירה של מקרופאגים לעבור מC5a משלים הדרגתי, בין 6 – 12 h לאחר הוספת כימוסטנט. מהירות תא ויעילות כימוטקתיות, הנכללות באינדקס כ-y-FMI (אינדקס העברה של y-forward; טווח: -1 אל + 1) ו-x-FMI, של מקרופאגים בודדים חושבו מחלקות הגירה (איור 3D). איור 3D מציג גם מגרש העברה שהופק לאחר נרמול נקודת ההתחלה של כל מסלול העברה ל-X = 0 ו-Y = 0 מתחת לחלקות התיבות. הכניסה בהתוויה של ההעברה מציגה את אופן החישוב של y-FMI עבור כל רצועת העברה.

איור 1: זרימת העבודה של שיטת כימוטקסיס. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: טיפול בשיקפי מוניות. (א) תצוגה תלת ממדית של מגלשת כימוטקיית עם ארבעה תקעים וארבעה כמוסות. השקופית מכילה שלוש תאי כימוטקסיס, כל אחד מהם מורכב מאגרי μl 2 40 מחוברים על ידי ערוץ 1 מ"מ x 2 מ"מ, אשר 70 יקרומטר גבוה, כינה את אזור התצפית. (ב) הערוץ המחבר משתרע בשני קצותיו כדי למלא את היציאות 2 ו-3. לאחר הכנסת תקעים לתוך מילוי יציאות 1 ו-4, אזור התצפית היה ממולא בינוני (אדום) על ידי החלת ירידה של בינונית ליציאה 3 ומכניס בנמל 2 עם טיפ 10-200 μL. לאחר מכן, כמוסות הוחלו על יציאות 2 ו-3 לפני הדגירה של השקופית ב 37 ° c והכנת ההשעיה התא. (ג) אזור התצפית, שם מעבר הכימוסטנט נוצר, היתה הזרע עם מקרופאגים על ידי החלת טיפה 10 μl של תאים הצפק תושב העכבר על היציאה 3 ולאחר מכן מעכפת ביציאה 2. השקופית לאחר מכן מודתה בחדר לחות ב 37 ° c עבור 2 – 3 h. התמונה חדות הפאזה המוצגת בצד ימין, מושגת באמצעות עדשת האובייקטיבי 10x, מראה תאים הצפק לאחר זריעה ודגירה ב 37 ° c עבור 2 h. סרגל קנה מידה = 500 μm. (ד) כימומוניות צ'יימברס היו מלאים בינוני על ידי חיבור יציאות 1 ו 2 ולאחר מכן לאט הזרקת בינוני דרך היציאה 3 עד מילוי איטי ויציב ניתן להשיג על-ידי הפיכת טבעת הגדרת עוצמת הקול של 20-100 μL. לאחר מילוי מאגר המים הראשון, ניתן למלא את המאגר השני באמצעות יציאות חיבור 2 ו-3 ולאחר מכן להזריק באופן איטי ביציאה 4 עד שהיא מתגלה ביציאה 1. (ה) תמונת חדות פאזה של אותו אזור תצפית המוצג לעיל (ג) לאחר מילוי שני המאגרים. סרגל בקנה מידה = 500 μm. מרכיבים גרפיים שסופקו על-ידי אליאס הורן. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
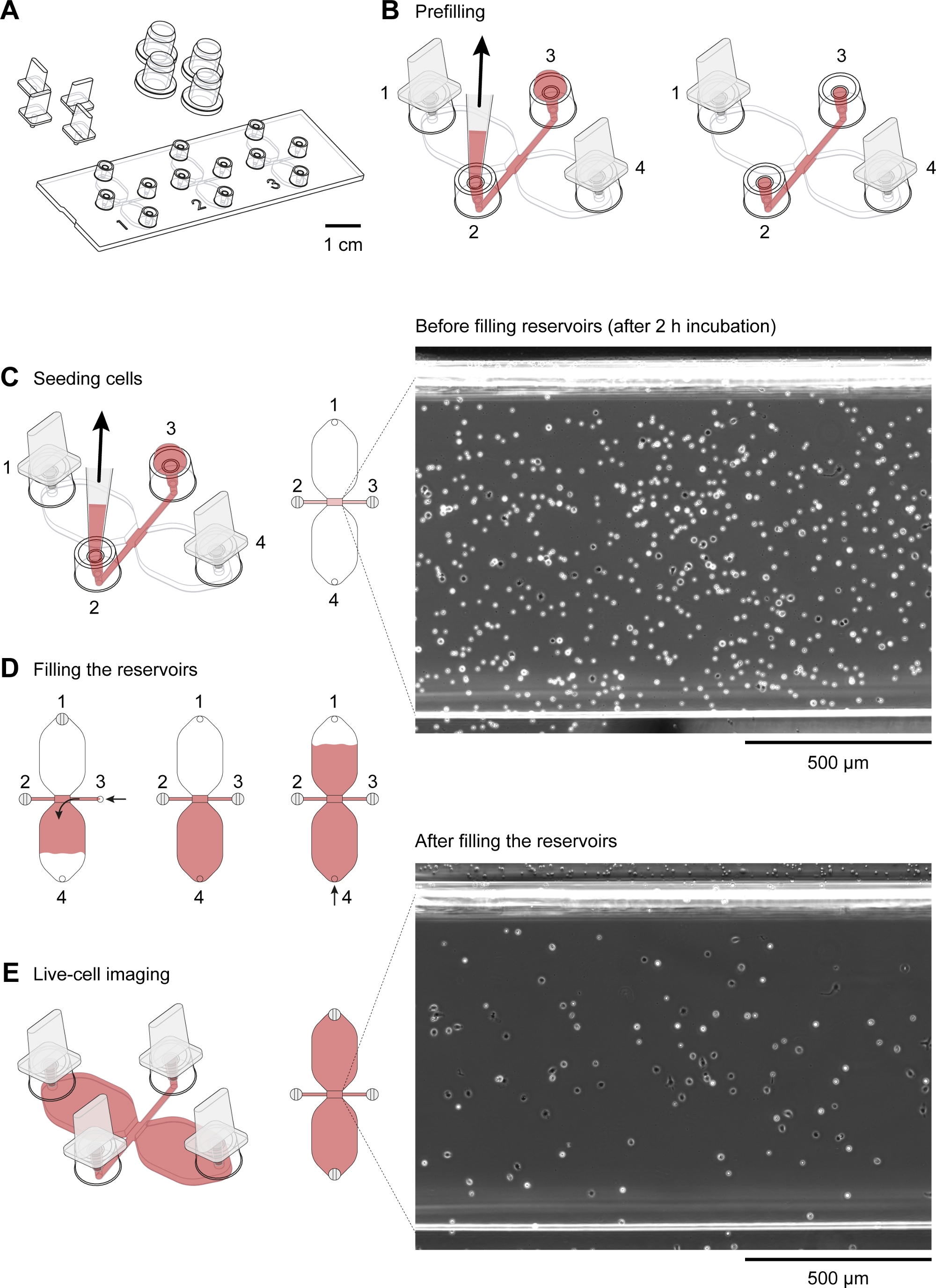{kind=link}

איור 3: שיטת כימוטקסיס. (א) כימוסטנט הוצג אחד משני מאגרים של תא כימוטקא על ידי החלת ירידה של 15 μl של בינוני המכיל 0.54 Μg/ml משלימים C5a ו 10 μg/Ml פטנטים כחול V למילוי יציאה 1, ואחריו שאיפה איטית בנמל 4. (ב) בתחילה לאחר שנשאב לתוך המאגר הכחול, המכיל כימוסטנט מכיל צורה בערך טיפה הפוכה, ולאחר מכן הפיזור לאט ברחבי המאגר. (ג) מסלולי הגירה של מקרופאגים הגירה ב כימוסטנט (המשלים C5a) הדרגתי בין 6 – 12 h לאחר החדרת כימוג לאחד המאגרים. הכיוון של מעבר הצבע מצוין מימין. הסוף של כל מסלול העברה מצוין על-ידי עיגול מלא. (ד) blox מגרשים של מהירות, x-fmi (מדד הגירה x-קדימה) ו-y-fmi (מדד הגירה y-קדימה), אינדקס של יעילות כימוטקאני נע בין -1 ל-+ 1. הנתונים הושגו על ידי ניתוח של 25 מסלולי הגירה מקרופאג. מקרופאגים במחצית התחתונה של אזור התצפית ומראה עקירה של רוחב תא אחד לפחות מעל 6 h נבחרו באופן אקראי לניתוח. להלן מגרש של רצועות העברה לאחר נרמול נקודת ההתחלה ל-X = 0 ו-Y = 0. האינדקס של כימוקסוניות (y-FMI) חושב על-ידי חלוקת ההזחה הנטו לאורך ציר Y (ד) באורך המצטבר (l) של נתיב ההעברה, כמוצג בהתאם. אלמנטים גרפיים בחלוניות A ו-B המסופקים על ידי אליאס הורן. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
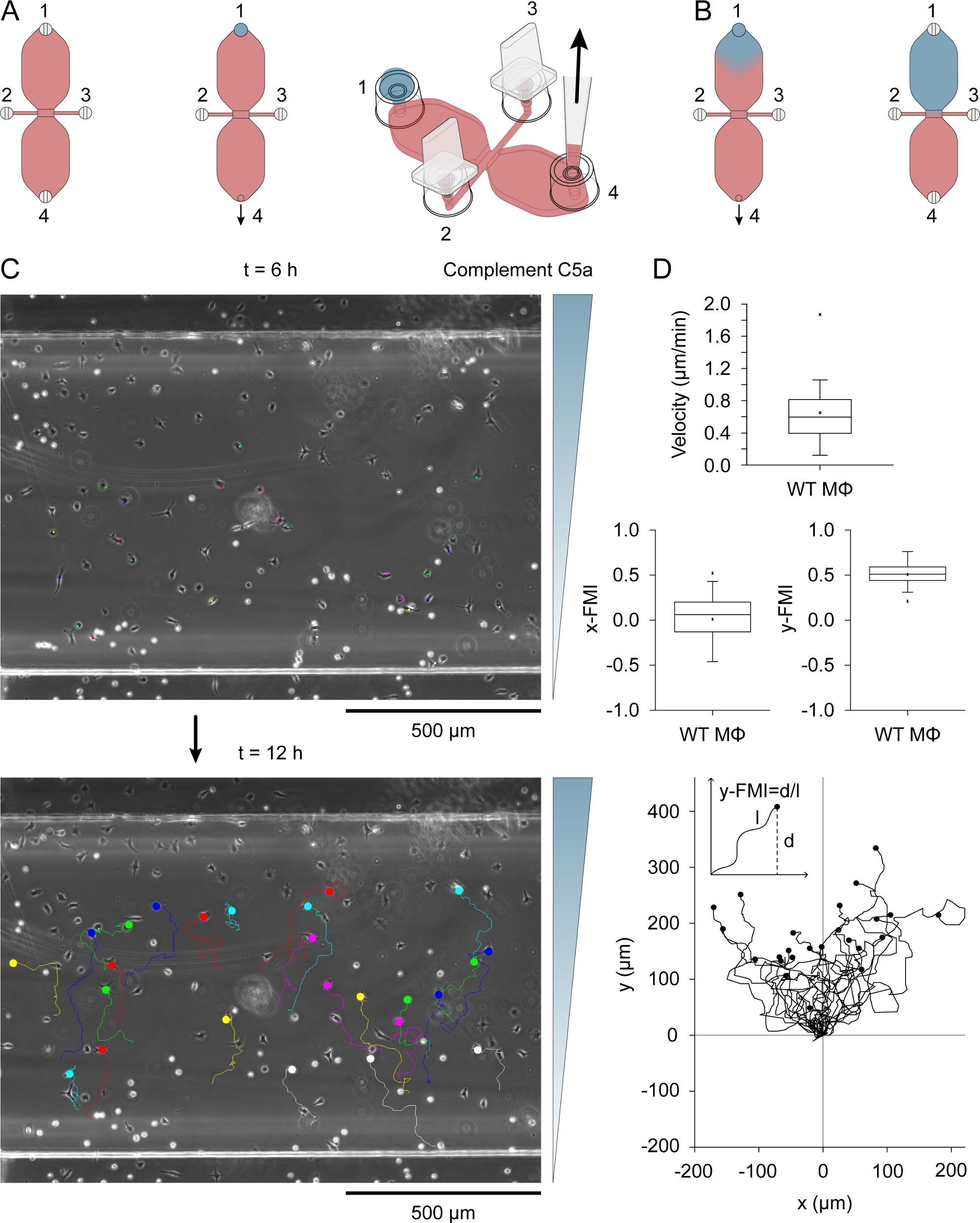{kind=link}

איור 4: תמונות פלורסנט של העכבר חי בתאי הצפק שהתקבלו על ידי ספינינג מיקרוסקופ הדיסק המסתובב. (א) תמונת מיקוד מורחבת (מיזוגהנקודה המבריק של כל Z-מטוסי) של תאים טריים הצפק מבודד העכבר מסומן עם פלורסנט ירוק אנטי F4/80 (מקרופאג marker) נוגדנים, פלורסנט אדום ANTI-CD19 (B סמן תא) נוגדנים, ו כחול הגרעין חומצה פלואורסצנטי כתם. סרגל בקנה מידה = 10 μm. (ב) תמונה (בודד Z-מישור) של F4/80+ תאים (מקרופאגים) באזור התצפית של תא כימוטווניות נלקח לאחר שיטת מוניות כימושל לילה. תאים סומנו עם פלורסנט ירוק anti-F4/80 נוגדנים ומכתם כחול הגרעין חומצה פלואורסצנטית. המשלים C5a ו פטנט כחול V הדרגתיים נשטפו על ידי הליך תיוג התא, מה שמסביר מדוע המאגר העליון בתרשים סכימטי של תא כימוטקסיס אינו כחול. סרגל קנה מידה = 10 μm. רכיב גרפי שסופק על ידי אליאס הורן. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
Discussion
הדמיה של האינוטרטל שתחילתה במאה ה -19 ומספקת אמצעים לחקר התנהגות של תאים חיסוניים חיים בסביבתם הטבעית. עם זאת, גם עם המיקרוסקופיה המתקדמת של היום וטכניקות גנטיות קשה ללמוד את התגובה של תאים כימוזרים ספציפיים vivo. כדי לעקוף בעיה זו, boyden18 פיתח transwell הוא אומר בשנות ה-60, אבל אלה הנקודה הסופית בחני לא לספק ויזואליזציה של איך התאים הועברו למעשה לקראת כימורים, מה שמקשה להבחין chemokinesis, מגורה הגירה אקראית על-ידי cue כימיים32, ו כימוטרמוניות, הגירה לעבר33ריכוזים בעיה זו נפתרה על ידי עיצוב התאים הפתוחים שונים עם גשר, בדרך כלל 1 מ"מ רחב, ממוקם בין שני מאגרים ונגישים על ידי עדשה אובייקטיבית21,22,23. החלת שובר כיסוי הפוך, הנזרע עם תאים חסיד, סוגר את התאים וכימוסטנט הוסיף לאחד המאגרים מפזרת על פני הגשר לתוך המאגר מנוגדות, יצירת מעבר ריכוז. כאן אנו מתארים שיטת כימוטווניות המשתמשת באותו עיקרון אך באמצעות חדר סגור הכולל ארבע יציאות מילוי. באמצעות מערכת זו ומעידה על זמן, מיקרוסקופ שלב-ניגודיות, פיתחנו שיטה לעכבר התמונה תושב הצפק מקרופאגים הגירה ב כימוטקטקטיקה C5a הדרגתי31,34,35,36. שיטת הפעולה הזאת, בשילוב עם מודלים של עכבר מנוקאאוט, הוכיחה כאינסטרומנטלי בחקירת התפקידים של מגוון שונים של Rho gtes וחלבונים מוטוריים ב מקרופאג מורפולוגיה, תנועתיות וכימוטקא31,34,35,36,37. השתמשנו גם גישה זו לתמונה האדם מונוציטים דם היקפי הגירה על פני משטח דו-ממדי או בסוג קולגן 3D I מטריצה38. יתר על כן, הצורך מתאים במח העצם של העכבר-מקרופאגים נגזר או מקרופאגים נגזר מותנה מותניתהתאים הקודאידמיאלואידית39,40. השתמשנו בעבר polyטטרפלואורואתילן (מצופה) תיקים עם מתאמים מצח לתאי מח עצם העצמות להשיג מקרופאגים34. היתרון של שקיות הוא כי התאים יכולים להיות מושעה מחדש ומוכן לשימוש לאחר הצבת התיק על הקרח 20 – 30 דקות. שים לב כי אנו ממלאים את האזור התצפית שקופיות כימוכוניות לפני הצגת התאים. גישה זו יש את היתרון כי בועות אוויר לא רצויות ניתן לאחר מכן החוצה (עם הצלחה משתנה) ואת אזור תצפית מראש מאפשר הקדמה איטית של תא השעיה על ידי ליטוף. מילוי מראש, למרות, מגדיל את הסבירות כי המדיום יזרום באופן חלקי לתוך אחד או שניהם של המאגרים האגפים, אשר יקדם את זריעת התאים מעבר לאזור התצפית. לחלופין, השעיית התא יכול להיות מצינורות ישירות לתוך אזור תצפית יבש, אבל בועות אוויר לא רצוי לא ניתן לסלק לאחר מכן.
חלל הצפק של העכבר מכיל שתי אוכלוסיות עיקריות של תאים: F4/80+ מקרופאגים ו (קטן) CD19+ B תאים, ביחס של כ 1:2 (איור 4a). אלה שני אוכלוסיות תאים החשבון עבור מעל 95% של תאים החלל הצפק, ואילו הנותרים F4/80/cd19- תאים בדרך כלל ניתן לזהות כמו CD11c+ תאים (תאים דנדריטים) או CD3+ תאים (T תאים). תאים בעלי מבנה חלש מתסוידים מחוץ לאזור התצפית בזמן מילוי המאגרים עם המדיום (איור 2). לאחר הוספת כימוסטנט לאחד משני מאגרי המים, צניחה בזמן, מיקרוסקופ הפאזה ניגודיות ניתן להשתמש כדי לדמות את התאים הנותרים (מקרופאגים) לעבור בהדרגה מעבר כימוסטנט מתפתחת. היווצרות C5a המשלים הדרגתי באזור התצפית, דרך דיפוזיה ממאגר אחד לשני, יכול להיות מדומה באמצעות צבע פלורסנט עם משקל מולקולרי דומה. תחליף טוב לרקומביננטי העכבר C5a (משקל מולקולרי חזוי, 9.0 kda) הוא מתויג תוספי (10 kda)31. באמצעות מיקרוסקופ קונפוקלית וקד, מעבר הקרינה הפלואורסצנטית בערוץ הצר (אזור תצפית) חיבור שני המאגרים של שקופית כימוטווניות ניתן למדוד במרווחי זמן קבועים ופרופילי ריכוז בנקודות הזמנים השונות ניתן להתוות24,31. אנו מוסיפים באופן קבוע את הצבע הכחול (פטנט כחול V) למדיום הכימוסטנט כדי לספק מחוון חזותי נוח של דיפוזיה והיווצרות מעבר צבע. בתוך 1 h של הצגת 15 μL של כחול, כימוג המכיל בינוני לתוך מאגר, המאגר מופיע כחול אחיד, על פי חוקי Fick של דיפוזיה, מעבר צבע יהיה לרוחב אזור תצפית צר חיבור המאגרים (איור 3B). מספר ימים נדרשים עבור הפתרון (צבע כחול או כימוט) כדי להיות מופץ אחיד.
מיקרוסקופ הקרינה הפלואורסצנטית יכול להיות מוחלף עבור המיקרוסקופיה הפאזה, אשר מציעה יתרונות עבור מעקב אחר תאים אוטומטיים, משום שתאים המסומנים באמצעות פלואור ניתן להבדיל בקלות מן הרקע. יתרון נוסף הוא כי אוכלוסיות ספציפיות של תאים חיסוניים יכול להיות מעקב סלקטיבי לאחר תיוג סמנים משטח עם נוגדנים פלורסנט. השתמשנו בגישה זו כדי תמונה היקפי דם CD14+ תאים (מונוולוציטים) הגירה ב כימוטקטיק fMLP (N-formylמתיונין-ליוציל-פנילאלנין) הדרגתי38. באופן דומה, הפלורסנט אנטי-F4/80 נוגדנים יכול לשמש את התמונה מקרופאגים העכבר להעברה כימוטקטקטיקה המשלים C5a הדרגתי. רעילות התמונה היא חיסרון פוטנציאלי של שימוש הדמיה פלואורסצנטית41. זה יכול להיות מופחת על ידי אמצעים שונים42, כולל שימוש fluorophores נרגש עם אורכי גל ארוכים יותר והוספת נוגדי חמצון למדיום. לחילופין, תאים המסומנים בתווית בתחילה יכול להיות מזוהה על ידי מיקרוסקופ פלואורסצנטית ולאחר מכן התמונה על ידי מעידה בזמן, מיקרוסקופ הפאזה ניגודיות. עם זאת, בפועל, תאים הנעים במהירויות נמוכות בינוני, כגון ~ 1 μm/min (מקרופאגים) או ~ 4 μm/min (מונוציטים), יכול להיות מיוצר לסירוגין על ידי המיקרוסקופיה פלואורסצנטית במרווחי זמן של דקות, אשר נסבל היטב38. בעבר השתמשנו במיקרוסקופיה פלואורסצנטית ושקופית כימוטקסיס שתוארה כאן38עבור 3d כימוטקסיסבחני 38,43. במקרה זה, שתי המאגרים היו מלאי מראש עם בינוני ו 15 μL כימותסטנט המכיל בינוני היה נשאב לתוך אחד המאגרים מיד לפני באיטיות ללטף באופן מיידי בתאים מושעה בינונית המכיל סוג קולגן אני לתוך אזור התצפית. החלק הקשה של הליך זה הוא טיפול בסוג קולגן אני, אשר מרוכז בפתרון חומצי. ה-pH של פתרון הקולגן צריך להיות מנוטרל על ידי תוספת של פתרון בסיסי לפני ערבוב את הפתרון הקרח קר קולגן עם השעיה התא. העברת תערובת הקולגן לחממה ב 37 ° c תיזום את הקולגן פולימוניזציה. במהלך הדגירה, יש לסובב באיטיות את השקופית סביב הציר הארוך שלה, כך שהתאים יישארו מפוזרים בצורה שווה בכיוונים של X-, Y ו-Z בעוד הקולגן הופך ג'ל. כימוס סגור הקשורות שקופית מתאים 3D כימוטקסיס assays, עם שישה תקעים במקום ארבעה תקעים, תיאר לאחרונה29. מערכת זו מאפשרת את התערובת תא הקולגן להיות מוכנס לאזור התצפית לפני באופן עצמאי מילוי כל אחד מאגרי המים, כי לכל מאגר יש שתי יציאות מילוי, ולא יציאה אחת.
לסיכום, אנו מתארים בזמן אמת מוניות כימוכיום המאפשר ויזואליזציה של תאים הניווט בהדרגה מעבר כימוטקטית על פני תקופה של 6 או יותר שעות. כאן אנו מתמקדים מקרופאגים, אשר לשחק תפקידים מרכזיים במחלות דלקתיות אבל כבר לא מיוצגים בזמן אמת בחני וניות כימואמר לעומת תאים נעים מהיר כמו נויטרופילים ו dictyelium אמבות.
Disclosures
. למחברים אין מה לגלות
Acknowledgements
עבודה זו הייתה נתמכת על ידי מענק (HA 3271/3-2) מ-DFG (דויטשה פורשונגסביסםהירכתיים).
Materials
| Name | Company | Catalog Number | Comments |
| µ-Slide (anodized aluminium) rack | Ibidi, Martinsried, Germany | 80003 | Autoclavable stackable rack for channel slides |
| µ-Slide Chemotaxis 2D (chemotaxis slide) | Ibidi, Martinsried, Germany | 80306 | Slide containing chemotaxis chambers (tissue culture treated) |
| 100x penicillin/streptomycin | Thermo Fisher Scientific | 15140122 | Used as supplement for RPMI 1640 media |
| 10-100 µL pipette with volume control ring | Eppendorf | 3123000047 | Eppendorf Research plus pipette |
| 10-200 µL pipette tips | Greiner Bio-One International | 739261 | Pipette tips with beveled tips (96 pieces per rack: sterile) |
| 14 mL polypropylene round bottom tubes | BD Falcon | 352059 | Used to collect peritoneal cells |
| 14-bit Hamamatsu C9100-50 Electron Multiplying-Charged Couple Device (EM-CCD) peltier-cooled camera | Hamamatsu Photonics Inc., Japan | EM-CCD camera of the spinning disk confocal microscope system | |
| 2-20 µL pipette with volume control ring | Eppendorf | 3123000039 | Eppendorf Research plus pipette |
| 24 G plastic catheter | B Braun Mesungen AG, Germany | 4254503-01 | Used for peritoneal lavage |
| 405 nm solid state laser, 50 mW | Perkin Elmer, Rodgau, Germany | Laser (405 nm) source of spinning disk confocal microscope system | |
| 488 nm solid state laser, 50 mW | Perkin Elmer, Rodgau, Germany | Laser (488 nm) source of spinning disk confocal microscope system | |
| 561 nm solid state laser, 50 mW | Perkin Elmer, Rodgau, Germany | Laser (561 nm) source of spinning disk confocal microscope system | |
| Alexa Fluor 488-conjugated rat (IgG2a) monoclonal (clone BM8) anti-mouse F4/80 antibody | Thermo Fisher Scientific | MF48020 | Mouse macrophage marker and plasma membrane label |
| Alexa Fluor 594-conjugated rat (IgG2a) monoclonal (clone 6D5) anti-mouse CD19 antibody | BioLegend | 115552 | Mouse B cell marker |
| C-Chip disposable (improved Neubauer) hemocytometer | NanoEnTek (distributed by VWR International) | 631-1098 | Used to count cells |
| CSU-X1 spinning disk scanner | Yokogawa Electric Corporation, Japan | Nipkow spinning disk unit | |
| Hank’s buffered salt solution without Ca2+ and Mg2+ | Thermo Fisher Scientific | 14170120 | Used for peritoneal lavage |
| Heat-inactivated fetal bovine serum | Thermo Fisher Scientific | 10082139 | Used as supplement for RPMI 1640 media |
| Hoechst 34580 | Thermo Fisher Scientific | H21486 | Cell permeable, blue fluorescent nucleic acid stain |
| ImageJ (image processing and analysis in Java) | National Institutes of Health (NIH) | Image analysis software | |
| Lipopolysaccharides from Escherichia coliO111:B4 | Sigma-Aldrich | L4391-1MG | Toll-like receptor 4 ligand |
| Nikon Eclipse Ti inverse microscope | Nikon, Japan | Inverted microscope | |
| Patent Blue V, sodium salt | Sigma-Aldrich | 21605-10G | Blue-colored dye used as visual indicator of gradient formation |
| Recombinant mouse complement C5a protein | R&D Systems | 2150-C5-025 | Chemoattractant for mouse macrophages |
| RPMI 1640 medium containing 20 mM Hepes | Sigma-Aldrich | R7388 | Basis medium for assays |
| UltraVIEW Vox 3D live cell imaging system + Volocity software | Perkin Elmer, Rodgau, Germany | Spinning disk confocal microscope system | |
| Zeiss LSM 510 + Axiovision software | Carl Zeiss Microscopy, Oberkochen, Germany | Confocal laser scanning microscope (LSM) adapted for phase-contrast microscopy |
References
- Lammermann, T., Germain, R. N. The multiple faces of leukocyte interstitial migration. Seminars in Immunopathology. 36, 227-251 (2014).
- Lammermann, T., Sixt, M. Mechanical modes of 'amoeboid' cell migration. Current Opinion in Cell Biology. 21, 636-644 (2009).
- Woodham, E. F., Machesky, L. M. Polarised cell migration: intrinsic and extrinsic drivers. Current Opinion in Cell Biology. 30, 25-32 (2014).
- Devreotes, P. N., et al. Excitable Signal Transduction Networks in Directed Cell Migration. Annual Review of Cell and Developmental Biology. 33, 103-125 (2017).
- Kamp, M. E., Liu, Y., Kortholt, A. Function and Regulation of Heterotrimeric G Proteins during Chemotaxis. International Journal of Molecular Sciences. 17 (1), 90 (2016).
- Miao, Y., et al. Wave patterns organize cellular protrusions and control cortical dynamics. Molecular Systems Biology. 15, 8585 (2019).
- Ridley, A. J., et al. Cell migration: integrating signals from front to back. Science. 302, 1704-1709 (2003).
- Lammermann, T., et al. Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature. 453, 51-55 (2008).
- Mullins, R. D., Heuser, J. A., Pollard, T. D. The interaction of Arp2/3 complex with actin: nucleation, high affinity pointed end capping, and formation of branching networks of filaments. Proceedings of the National Academy of Sciences of the United States of America. 95, 6181-6186 (1998).
- Leithner, A., et al. Diversified actin protrusions promote environmental exploration but are dispensable for locomotion of leukocytes. Nature Cell Biology. 18, 1253-1259 (2016).
- McDonald, B., et al. Intravascular danger signals guide neutrophils to sites of sterile inflammation. Science. 330, 362-366 (2010).
- Leber, T. Ueber die Entstehung der Entzündung und die Wirkung der entzündungserregenden Schädlichkeiten. Fortschritte der Medizin. 6, 460-464 (1888).
- Tauber, A. I. Metchnikoff and the phagocytosis theory. Nature Reviews Molecular Cell Biology. 4, 897-901 (2003).
- Clark, E. R., Linton Clark, E. Reactions of cells in the tail of amphibian larvae to injected croton oil (aseptic inflammation). American Journal of Anatomy. 27, 221-254 (1920).
- Clark, E. R., Linton Clark, E. The reaction of living cells in the tadpole's tail toward starch, agar-agar, gelatin, and gum arabic. The Anatomical Record. 24, (1922).
- Comandon, J. Phagocytose in vitro des Hématozoaires du Calfat (enregistrement cinématographique). Comptes Rendus Hebdomadaires des Séances et Mémoires de la Société de Biologie. 69, 314-316 (1917).
- McCutcheon, M. Chemotaxis in leukocytes. Physiological Reviews. 26, 319-336 (1946).
- Boyden, S. The chemotactic effect of mixtures of antibody and antigen on polymorphonuclear leucocytes. The Journal of Experimental Medicine. 115, 453-466 (1962).
- Horwitz, D. A., Garrett, M. A. Use of leukocyte chemotaxis in vitro to assay mediators generated by immune reactions. I. Quantitation of mononuclear and polymorphonuclear leukocyte chemotaxis with polycarbonate (nuclepore) filters. Journal of Immunology. 106, 649-655 (1971).
- Bignold, L. P. A novel polycarbonate (Nuclepore) membrane demonstrates chemotaxis, unaffected by chemokinesis, of polymorphonuclear leukocytes in the Boyden chamber. Journal of Immunological Methods. 105, 275-280 (1987).
- Zigmond, S. H. Ability of polymorphonuclear leukocytes to orient in gradients of chemotactic factors. The Journal of Cell Biology. 75, 606-616 (1977).
- Zicha, D., Dunn, G. A., Brown, A. F. A new direct-viewing chemotaxis chamber. Journal of Cell Science. 99, 769-775 (1991).
- Muinonen-Martin, A. J., Veltman, D. M., Kalna, G., Insall, R. H. An improved chamber for direct visualisation of chemotaxis. PLoS One. 5, 15309 (2010).
- Zengel, P., et al. mu-Slide Chemotaxis: a new chamber for long-term chemotaxis studies. BMC Cell Biology. 12, 21 (2011).
- Valentim, A. M., Guedes, S. R., Pereira, A. M., Antunes, L. M. Euthanasia using gaseous agents in laboratory rodents. Lab Animal. 50, 241-253 (2016).
- Franks, N. P. General anaesthesia: from molecular targets to neuronal pathways of sleep and arousal. Nature Reviews. Neuroscience. 9, 370-386 (2008).
- Cordelieres, F. P., et al. Automated cell tracking and analysis in phase-contrast videos (iTrack4U): development of Java software based on combined mean-shift processes. PLoS One. 8, 81266 (2013).
- Zantl, R., Horn, E. Chemotaxis of slow migrating mammalian cells analysed by video microscopy. Methods in Molecular Biology. 769, 191-203 (2011).
- Biswenger, V., et al. Characterization of EGF-guided MDA-MB-231 cell chemotaxis in vitro using a physiological and highly sensitive assay system. PLoS One. 13, 0203040 (2018).
- Austyn, J. M., Gordon, S. F4/80, a monoclonal antibody directed specifically against the mouse macrophage. European Journal of Immunology. 11, 805-815 (1981).
- Hanley, P. J., et al. Motorized RhoGAP myosin IXb (Myo9b) controls cell shape and motility. Proceedings of the National Academy of Sciences of the United States of America. 107, 12145-12150 (2010).
- Wilkinson, P. C. Cell Locomotion and Chemotaxis: Basic Concepts and Methodological Approaches. Methods. 10, 74-81 (1996).
- Pfeffer, W. Locomotorische Richtungsbewegungen durch chemische Reize. Untersuchungen aus dem Botanischen Institut zu Tübingen. 1, 363 (1884).
- Konigs, V., et al. Mouse macrophages completely lacking Rho subfamily GTPases (RhoA, RhoB, and RhoC) have severe lamellipodial retraction defects, but robust chemotactic navigation and altered motility. The Journal of Biological Chemistry. 289, 30772-30784 (2014).
- Horsthemke, M., et al. Multiple roles of filopodial dynamics in particle capture and phagocytosis and phenotypes of Cdc42 and Myo10 deletion. The Journal of Biological Chemistry. 292, 7258-7273 (2017).
- Bachg, A. C., et al. Phenotypic analysis of Myo10 knockout (Myo10(tm2/tm2)) mice lacking full-length (motorized) but not brain-specific headless myosin X. Scientific Reports. 9, 597 (2019).
- Horsthemke, M., et al. A novel isoform of myosin 18A (Myo18Agamma) is an essential sarcomeric protein in mouse heart. The Journal of Biological Chemistry. 294, 7202-7218 (2019).
- Bzymek, R., et al. Real-time two- and three-dimensional imaging of monocyte motility and navigation on planar surfaces and in collagen matrices: roles of Rho. Scientific Reports. 6, 25016 (2016).
- Wang, G. G., et al. Quantitative production of macrophages or neutrophils ex vivo using conditional Hoxb8. Nature Methods. 3, 287-293 (2006).
- Gran, S., et al. Imaging, myeloid precursor immortalization, and genome editing for defining mechanisms of leukocyte recruitment in vivo. Theranostics. 8, 2407-2423 (2018).
- Magidson, V., Khodjakov, A. Circumventing photodamage in live-cell microscopy. Methods in Cell Biology. 114, 545-560 (2013).
- Icha, J., Weber, M., Waters, J. C., Norden, C. Phototoxicity in live fluorescence microscopy, and how to avoid it. BioEssays : News and Reviews in Molecular, Cellular and Developmental Biology. 39 (8), 1700003 (2017).
- Isfort, K., et al. Real-time imaging reveals that P2Y2 and P2Y12 receptor agonists are not chemoattractants and macrophage chemotaxis to complement C5a is phosphatidylinositol 3-kinase (PI3K)- and p38 mitogen-activated protein kinase (MAPK)-independent. The Journal of Biological Chemistry. 286, 44776-44787 (2011).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved