
Method Article
Zeitraffer-Bildgebung von Mausmakrophage Chemotaxis
In diesem Artikel
Zusammenfassung
Hier beschreiben wir Methoden mit Zeitraffer-Phasen-Kontrast-Mikroskopie, um peritoneale Makrophagen in einem chemotaktischen Komplement C5a-Gradienten zu beleben. Die Protokolle können auf andere Immunzellen ausgedehnt werden.
Zusammenfassung
Chemotaxis ist rezeptorvermittelte Führung von Zellen entlang eines chemischen Gradienten, während Chemokinese die Stimulation der zufälligen Zellmotilität durch eine Chemikalie ist. Chemokinese und Chemotaxis sind für die Mobilisierung und den Einsatz von Immunzellen von grundlegender Bedeutung. Zum Beispiel können Chemokine (chemotaktische Zytokine) schnell zirkulierende Neutrophile und Monozyten an extravaskuläre Entzündungsstellen rekrutieren. Chemoattractant-Rezeptoren gehören zur großen Familie der G-Protein-gekoppelten Rezeptoren. Wie Chemoattractant (d.h. Liganden) die direkte Zellmigration über G-Protein-gekoppelte Rezeptor-Signalisierung verläufet, ist noch nicht vollständig geklärt. Im Bereich der Immunologie sind Neutrophile beliebte Modellzellen zur Untersuchung von Chemotaxis in vitro. Hier beschreiben wir einen in Echtzeit zweidimensionalen (2D) Chemotaxis-Assay, der auf Maus-Resident-Makrophagen zugeschnitten ist, die traditionell schwieriger zu untersuchen sind. Makrophagen bewegen sich mit einem langsamen Tempo von 1 m/min auf einer 2D-Oberfläche und eignen sich weniger gut für Punkt-Quell-Migrationstests (z. B. Migration zur Spitze einer mit Chemoattractant gefüllten Mikropipette) als Neutrophile oder Dictyostelium-Discoideum, die eine Größenordnung schneller bewegen. Weit verbreitete Transwell-Assays sind nützlich für die Untersuchung der chemotaktischen Aktivität verschiedener Substanzen, liefern aber keine Informationen über Zellmorphologie, Geschwindigkeit oder chemotaktische Navigation. Hier beschreiben wir einen zeitraffiumbasierten Makrophagen-Chemotaxis-Assay, der die Quantifizierung der Zellgeschwindigkeit und der chemotaktischen Effizienz ermöglicht und eine Plattform bietet, um die Wandler, Signalwege und Effektoren von Chemotaxis abzuleiten.
Einleitung
Immunzellen wandern in der Regel singend auf einer 2D-Oberfläche in einer amoebariden Art und Weise1,2, die wiederholte Zyklen der Vorkonstruktion der Vorderseite, Integrin-vermittelte Zellhaftung und Rückzug der Rückseite beinhaltet. Ein Voraussetzung ist die Zellpolarisation, bei der Zellen vordere und hintere Ends3bilden. Chemotaxis beginnt mit dem Nachweis von Chemoattractanten durch G-Protein-gekoppelte Rezeptoren und einem komplexen Signalnetzwerk, das durch membranverankerte heterotrimere G-Proteine und kleine monomere G-Proteine sowie phospholipidgebundene Guanin-Nukleotid-Austauschfaktoren (GEFs)4,5vermittelt wird. Aktivierung von Rho GTPases der Cdc42 und Rac Subfamilien induzieren Vorsprünge an der Vorderen6 und Mitglieder der Rho Unterfamilie, vor allem RhoA, aktivieren Kontraktion der hinteren5,7. In einer dreidimensionalen (3D) Umgebung sind Integrine für die Leukozytenmigration weitgehend redundant und RhoA wird wichtiger für das Quetschen von Zellen durch enge Passagen8, während die Cdc42- oder Rac-induzierte Arp2/3-Aktivierung für die chemotaktische Lenkung9,10wichtig bleibt.
Immunzellen können verschiedenen Chemo-Peugern ausgesetzt sein, vor allem in den Umgebungen von Gewebeverletzungen, Pathogen-Invasion, und Entzündungen. Die endogenen Chemoattractants, die auf Phagozyten exprimiert werden, ergänzen C3a und C5a, werden durch Aktivierung der Komplementkaskade schnell erzeugt und werden durch komplementierte C3a- und C5a-Rezeptoren erkannt. In ähnlicher Weise rekrutieren nekrotische Zellen Phagozyten über Formylpeptid-Rezeptoren, die mitochondrie-abgeleitete sowie bakterielle Formylpeptide erkennen11. Immunzellen exprimieren auch G-Protein-gekoppelte Rezeptoren für Chemokine, eine große Familie von Chemoattractanten peptiden, die an der Regulierung des Immunzellhandels während der Homöostase und Entzündung beteiligt sind. Chemokine werden in vier Gruppen eingeteilt, abhängig vom Abstand der ersten beiden Cystein (C)-Rückstände: C, CC, CXC und CX3C Zytokine, wobei X eine Aminosäure ist. Daher müssen in vivo Immunzellen angemessen auf hochkomplexe räumliche und zeitliche Signale reagieren, was die Untersuchung von Chemotaxis zu einer entmutigenden Aufgabe macht. Im Folgenden stellen wir eine kurze Geschichte der Chemotaxis, die mit intravitalen bildgebenden Ansätzen begann.
Die Untersuchung der Leukozyten-Chemotaxis geht auf das Jahr 188812zurück, als der Augenarzt Theodor Karl Gustav Leber die gezielte Migration von Leukozyten zu und die Anhäufung von Entzündungsstellen in einem Modell mykotischer (pilzförmiger) Keratitis klar beschrieb. Leber betonte, dass die Anziehungskraft überschüssiger Leukozyten durch pathogene Substanzen wichtig für die Eliminierung schädlicher Mikroorganismen mittels Phagozytose sei, die von Metchnikoff (auch bekannt als Metschnikoff) Anfang des Jahrzehnts13beschrieben worden sei. In vivo-Experimente wurden auch in den 1920er Jahren von Clark und Clark14,15, durchgeführt, die die Transparenz der Kaulquappen nutzten und zeigten, dass sterile Entzündungen, die durch Crotonöl 14 oder andere Reizstoffe15 induzierten, Dazu führten, dass Leukozyten an den Blutgefäßen haften, gefolgt von Diapedese (transendotheliale Migration) und einer schnellen Migration durch die Geweberäume zum Reizstoff.15 In-vitro-Experimente mit der von Jean Comandon16 entwickelten Mikrokinzographie-Methode zeigten, dass Leukozyten zu einer partikelartigen Chemoattractantenquelle wie Bakterien17wanderten. Zu dieser Zeit waren die molekularen Identitäten chemotaktischer Faktoren unbekannt. In den 1960er Jahren erkannte Stephen Boyden18, dass Techniken zur Untersuchung der chemotaktischen Aktivität löslicher Substanzen fehlten. Er entwarf eine Kammer, die später als Boydenkammer bekannt war, mit zwei Fächern, die durch eine Filterpapiermembran getrennt waren. Dem oberen Fach wird eine Zellaufhängung zugesetzt und die Prüfsubstanz entweder beiden Fächern oder nur dem unteren Fach zugesetzt. Nach einer Inkubationszeit wird die Filtermembran entfernt und die Zellen fixiert und gebeizt. Durch den Vergleich, wie viele Zellen über die Filtermembran zum unteren Brunnen mit der Prüfsubstanz in beiden Fächern, weder in den Fächern noch nur im unteren Fach, wandern, kann die chemotaktische Aktivität bestimmt werden. Transwell-Assays sind auch heute noch beliebt und wurden auf verschiedene Weise modifiziert, einschließlich der Verwendung von verschiedenen Polycarbonat-Membranen mit definierten Porengrößen und Dichten19,20. Ein großer Nachteil von Transwell-Assays ist, dass es unpraktisch ist, Zellen, die migrieren, direkt zu visualisieren und der Migrationspfad über die Membran in der Regel den Durchmesser einer Immunzelle nicht überschreitet.
Sally H. Zigmond entwickelte eine Chemotaxiskammer21, die die Visualisierung sowohl der Gradientenbildung als auch der Zellmorphologie mit Fluoreszenzfarbstoffen ermöglichte. Die Kammer besteht aus einem Plexiglas (Acryl) Schlitten mit zwei parallelen linearen Bohrungen, jeweils mit einem Volumen von 100 l, getrennt durch eine 1 mm breite Brücke 3–10 m unter der oberen Ebene des Dias. Ein mit Zellen gesäter Deckslip wird invertiert und so auf die Folie gelegt, dass er die beiden Brunnen überspannt. Nach Zugabe eines Chemoattractants zu einem der Brunnen bildet sich über die Brücke ein steiler chemoattractantgradienter Gradient, typischerweise innerhalb von 30–90 min. Menschliche polymorphonukleare Leukozyten (Granulozyten) in der Zigmond-Kammer werden beobachtet, die sich am Chemoanziehen orientieren. Es wurden Variationen der Zigmond-Kammer berichtet, darunter die Kammern Dunn22 und Insall23, die beide einen Deckschein verwenden, der mit Zellen bestückt ist, die über zwei Brunnen gelegt werden, die durch eine 1 mm breite Brücke getrennt sind. Die Dunn-Kammer besteht aus konzentrischen Brunnen, die durch eine kreisförmige Brücke getrennt sind, während die Insall-Kammer enger mit der Zigmond-Kammer verwandt ist, aber Brücken von zwei verschiedenen Breiten, 0,5 mm und 1 mm, bietet. Zemgel et al.24beschrieben eine neuartige Chemotaxis-Kammer, die als "Slide Chemotaxis" bezeichnet und aus Kunststoffspritzguss hergestellt wird. Die Chemotaxiskammer besteht aus zwei 40-L-Reservoirs, die durch einen 1 mm breiten Kanal mit einer Länge von 2 mm und einer Höhe von 70 mm voneinander getrennt sind. Der Boden der Kammer besteht aus einer gasdurchlässigen, dünnen Kunststoffplatte mit der gleichen Dicke und optischen Eigenschaften eines Glasdeckels Nr. 1,524. Hier beschreiben wir einen Chemotaxis-Assay, der die Kammer der Slide Chemotaxis verwendet, um die Migration von mausansässigen Peritonealmakrophagen für bis zu 14 h in einem chemotaktischen (Komplement C5a) Gradienten zu visualisieren.
Protokoll
Die Protokolle folgen den Richtlinien unserer lokalen Forschungsethikkommission sowie den Tierpflegerichtlinien.
HINWEIS: Abbildung 1 zeigt einen Workflow des Chemotaxis-Assays.
1. Prefilling Chemotaxis Rutschen
-
Vorfüllen Sie die 1 mm breiten und 2 mm langen Verbindungskanäle eines oder zweier Chemotaxis-Dias mit modifiziertem RPMI 1640 HEPES-Medium, bestehend aus bikarbonatfreiem RPMI 1640-Medium mit 20 mM 4-(2-Hydroxyethyl)-1-Piperazinthanulfonsäure (HE PES), 10 % hitzeinaktiviertes fetales Rinderserum (FBS) und Antibiotika wie Penicillin (100 U/ml) und Streptomycin (100 g/ml), hergestellt durch Verdünnung von 100x Penicillin/Streptomycin, und 1 g/ml Lipopolysaccharid (aus E. coli) und ein toller Rezeptor 4 Liganden , um die Zellen zu aktivieren.
- Legen Sie eine Chemotaxis -Folie (Abbildung 2A) in eine runde (10 cm Durchmesser) Zellkulturschale, die beide auf 37 °C vorgewärmt sind, und stellen Sie die Schale auf einen beheizten (37 °C) Aluminiumblock. Stecken Sie Stecker in die Ports 1 und 4 (Abbildung 2B).
HINWEIS: Ein Aluminiumblock, der bei 37 °C gehalten und in der laminaren Durchflusshaube platziert wird, ist nützlich für die Vorbereitung von Chemotaxiskammern. Idealerweise sollte der beheizte Block einen flachen Arbeitsbereich und Brunnen für verschiedene Rohre, wie 50 ml-Rohre und 2 ml Mikrozentrifugenrohre, bieten. - Mit einer 10–200 L PipetteSpitze mit abgeschrägter Spitze 15 l modifiziertes RPMI 1640 HEPES Medium in Füllanschluss 3 ablagern (Abbildung 2B). Als nächstes, wenn das Volumen noch auf 15 l eingestellt ist und die Steuertaste der Pipette (2–20 l Volumen) gedrückt ist, legen Sie die Pipettenspitze in Anschluss 2 ein und aspirieren Sie 15 l mit einer mäßig schnellen Rate(Abbildung 2B). Dadurch werden der 1 mm x 2 mm Verbindungskanal (Beobachtungsbereich) sowie die beiden flankierenden Versorgungskanäle (zwischen dem zentralen Beobachtungsgebiet bzw. den Ports 2 bzw. 3) vorgefüllt. Abdeckung der Füllöffnungen 2 und 3 mit Kappen.
- Nach dem Vorfüllen die Chemotaxis auf ein Regal, das in einer geschlossenen Feuchtigkeitskammer in einem ansonsten trockenen undCO2-freienInkubator bei 37 °C aufbewahrt wird, legen.
HINWEIS: Es ist wichtig, die richtige Pipettenspitze zum Füllen der Chemotaxis-Rutsche zu verwenden. Eine abgeschrägte Pipettenspitze verkeilt sich in die Oberseite des Füllanschlusses, während häufig verwendete spitze Pipettenspitzen tiefer in den Füllanschluss eingeführt werden können und den Widerstand gegen Flüssigkeitsfluss erheblich erhöhen können.
- Legen Sie eine Chemotaxis -Folie (Abbildung 2A) in eine runde (10 cm Durchmesser) Zellkulturschale, die beide auf 37 °C vorgewärmt sind, und stellen Sie die Schale auf einen beheizten (37 °C) Aluminiumblock. Stecken Sie Stecker in die Ports 1 und 4 (Abbildung 2B).
2. Isolierung von Maus Resident Peritoneal Macropages
- Opfern Sie eine 3-4 Monate alte Maus mit einer hohen Konzentration des flüchtigen Anästhetikums Isofluran (>5% in der Luft) oder Kohlendioxid25, gefolgt von zervikalen Dislokation. Der Verlust des Rechtenreflexes bei Nagetieren korreliert mit dem Verlust des Bewusstseins beim Menschen26. Reinigen Sie den Bauch der Maus mit 80% Ethanol in Wasser und machen Sie dann einen 1–2 cm Mittellinienhautschnitt mit chirurgischer Schere mit stumpfen Spitzen. Schälen Sie die Haut zurück, um die darunter liegende Bauchwand freizulegen.
- Setzen Sie einen 24 G Kunststoffkatheter in die Peritonealhöhle ein. Mit einer 5 ml Kunststoffspritze den Hohlraum mit 2 x 4,5 ml eiskalter Hanks gepufferter Salzlösung (HBSS) ohne Ca2+ und Mg2+spülen. Lassen Sie ca. 0,5 ml Rest-HBSS in der Spritze, so dass versehentlich auf die Katheterspitze gesaugtes Gewebe ausgestoßen werden kann.
- Übertragen Sie das lavaged Medium, in der Regel 8-8,5 ml insgesamt, in ein 14 ml Polypropylen rundes Unterrohr. Zentrifugieren Sie das Rohr bei 300 x g für 6,5 min bei Raumtemperatur.
HINWEIS: Das runde Unterrohr ermöglicht eine vollständige Dekantierung des Überstandes und reduziert zellverstopfte. - Entsorgen Sie den Überstand und setzen Sie die Peritonealzellen (in der Regel 4 x 106 Zellen pro Maus) in 200 L modifiziertem RPMI 1640 HEPES-Medium wieder aus. Verdünnen Sie eine Probe der Zellsuspension 1:20 und verwenden Sie eine Zählvorrichtung, wie z. B. eine Neubauer verbesserte Zählkammer, um die Zellen zu zählen. Anschließend verdünnen Sie die Zellsuspension auf eine Endkonzentration von 10 x 106 Zellen/ml und halten Sie die Zellen in einem runden unteren 2 ml Polypropylen-Mikrozentrifugenrohr bei 37 °C mit einem beheizten Aluminiumblock (siehe ANMERKUNG in Schritt 1.1.1).
3. Saat peritoneale Zellen in Chemotaxis-Dias
- Nach dem Pipetieren der Zellsuspension nach oben und unten 5x mit dem Pipettenvolumen auf 100 l (oder bei der Hälfte des Suspensionsvolumens) eingestellt, um Verklumpungen zu reduzieren, legen Sie vorsichtig 10 'L der Zellsuspension in Port 3 einer Chemotaxiskammer ab (Abbildung 2C). Legen Sie die Pipettenspitze in Port 2 und ziehen Sie die Zellaufhängung langsam in den Verbindungskanal (Abbildung 2C). Sobald die Zellsuspension eingeführt wurde, entfernen Sie die Stecker an den Ports 1 und 4, was dazu beitragen wird, den Fluss der Zellsuspension zu verhaften. Platzieren Sie Kappen an allen vier Füllanschlüssen.
- Wiederholen Sie Schritt 3.1 für alle Chemotaxiskammern. Mit einem kleinen invertierten Mikroskop und einer 10x Phasenkontrastobjektivlinse inspizieren Sie die Chemotaxis-Dias auf unerwünschte Luftblasen.
- Die Chemotaxis-Dias mit Peritonealzellen in eine Feuchtigkeitskammer bei 37 °C für 2-3 h setzen.
4. Füllen der Reservoirs und Hinzufügen von Chemoattractant
- Prüfen Sie den Beobachtungsbereich (Kanal, der die beiden 40-L-Reservoirs verbindet) mit einem invertierten Mikroskop.
HINWEIS: In diesem Stadium wird die Zelldichte höher sein als nach dem Füllen der Reservoirs, da schwach anhaftende Zellen, überwiegend CD19+ Zellen (B1-Zellen), während des Füllvorgangs aus dem Beobachtungsbereich gewaschen werden (Abbildung 2C-E). - Stecken Sie Stecker in die Füllanschlüsse 1 und 2 (Abbildung 2D). Stellen Sie sicher, dass der Füllanschluss 3 nach oben mit medium und frei von Luftblasen gefüllt ist. Verwenden Sie eine sterile 27 G Spritzennadel, um bei Bedarf unerwünschte Luftblasen zu lösen.
- Mit einer mechanischen Pipette mit 10–100 l Volumen, aspirieren Sie 60 L modifiziertes RPMI 1640 HEPES-Medium und legen Sie die Pipettenspitze in Denfüllanschluss 3. Verwenden Sie den Volumeneinstellungsring der Pipette, um langsam und stetig Medium in das Reservoir zu injizieren, so dass Das Medium nach 1–2 min die Oberseite des Füllanschlusses 4 erreicht (Abbildung 2D).
- Füllen Sie das zweite Reservoir. Verschieben Sie den Stecker von Port 1 und setzen Sie ihn langsam in Port 3 ein (Abbildung 2D). Als nächstes aspirieren Sie 50 L modifiziertes RPMI 1640 HEPES Medium und legen Sie die Pipettenspitze in Füllanschluss 4. Verwenden Sie den Volumeneinstellungsring der 10–100 L Volumenpipette, um langsam und stetig Medium in das zweite Reservoir zu injizieren, so dass das Medium nach 1–2 min die Oberseite des Füllanschlusses 1 erreicht(Abbildung 2D).
- Legen Sie 495 l modifiziertes RPMI 1640 HEPES-Medium in ein (rundes Untermal) 2 ml Mikrozentrifugenrohr und fügen Sie 5 l Patent Blue V (Lagerlösung: 10 mg/ml in phosphatgepufferter Saline [PBS]) hinzu), einem blauen Farbstoff, der als visueller Indikator für die Konzentrationsgradientenbildung verwendet wird. Mix durch kurze Wirbel. Fügen Sie 5,4 l rekombinante Maus komplementieren C5a (Lagerlösung: 50 g/ml in PBS mit 0,1% Rinderserumalbumin) und mischen Durch kurze Wirbel.
- Legen Sie 15 l blau ab, ergänzen Sie C5a-haltiges Medium in Füllanschluss 1 (Abbildung 3A), nachdem Sie sichergestellt haben, dass die flache Vertiefung an der Spitze des Ports mittelfrei ist (sonst kann der Tropfen überschwappen).
- Legen Sie eine 10–200 L Pipettenspitze in Füllanschluss 4 ein und drehen Sie langsam und stetig den Volumeneinstellungsring der 10–100 L Volumenpipette, um den Tropfen des blauen, C5a-haltigen Mediums in das gegenüberliegende Reservoir zu ziehen (Abbildung 3B). Luft beginnt, die kurze vertikale Spalte des Füllanschlusses 1 zu betreten. Zeichnen Sie Luft ein, bis sich die Flüssigkeits-Luft-Schnittstelle in der Mitte der vertikalen Spalte befindet, und legen Sie dann langsam einen Stecker in den Anschluss ein.
- Heben Sie die Pipette vorsichtig von Port 4 an, indem Sie die andere Hand verwenden, um sicherzustellen, dass die Rutsche an Ort und Stelle fixiert bleibt. Schließlich, langsam Stecker-Port 4 (Abbildung 3B).
- Prüfen Sie das Chemotaxis-Dia auf einem invertierten Mikroskop.
HINWEIS: Die verbleibenden haftenden Zellen im Beobachtungsgebiet sollten überwiegend Makrophagen sein. Dies kann mit fluoreszierend markierten Anti-F4/80-Antikörpern bestätigt werden (F4/80 ist ein spezifischer Marker für Mausmakrophagen). B-Zellen können mit fluoreszierend markierten Anti-CD19-Antikörpern und F4/80-/CD19- Zellen mit einem blauen fluoreszierenden Nukleinsäurefleck nachgewiesen werden (Abbildung 4).
5. Bildgebende Makrophagenmigration durch Zeitraffer, Phasenkontrastmikroskopie
- Legen Sie eine Chemotaxis-Folie auf die Bühne eines invertierten Mikroskops, das mit einem Bühneninkubator ausgestattet ist. Halten Sie die Temperatur bei 37 °C.
- Stellen Sie den 1 mm x 2 mm Beobachtungsbereich mit einer 10x Phasenkontrastobjektivlinse ab und konzentrieren Sie sich auf die Makrophagenlamellipodie: dünne, blechartige Membranvorsprünge. Erfassen Sie Bilder für 14 h mit einer Rate von 1 Frame alle 2 min.
6. Analyse von Zeitrafferbildern
- Analysieren Sie die Zeitraffer-Phasen-Kontrast-Bilder mit der automatisierten Bildanalyse-Software oder dem Manual Tracking Plugin, das von Fabrice P. Cordeliéres für ImageJ erstellt wurde.
HINWEIS: Automatisierte Tracking-Programme können verwendet werden, um Zellen zu analysieren, die entweder durch Zeitraffer-, Phasenkontrast- oder Fluoreszenzmikroskopie abgebildet sind. Beispielsweise kann die Java-basierte Software iTrack4U, die von Cordeliéres et al.27 produziert wurde, zur automatisierten Zellverfolgung und -analyse mit Zeitraffer-, Phasenkontrast- oder Fluoreszenzbildern als Eingabe verwendet werden. Manuelle Seperge ist zeitaufwändiger, aber die Tracks, die durch das ImageJ Plugin Manual Tracking generiert werden, können direkt importiert und automatisch durch das ImageJ Plugin Chemotaxis und Migration Tool28,29analysiert werden.
Ergebnisse
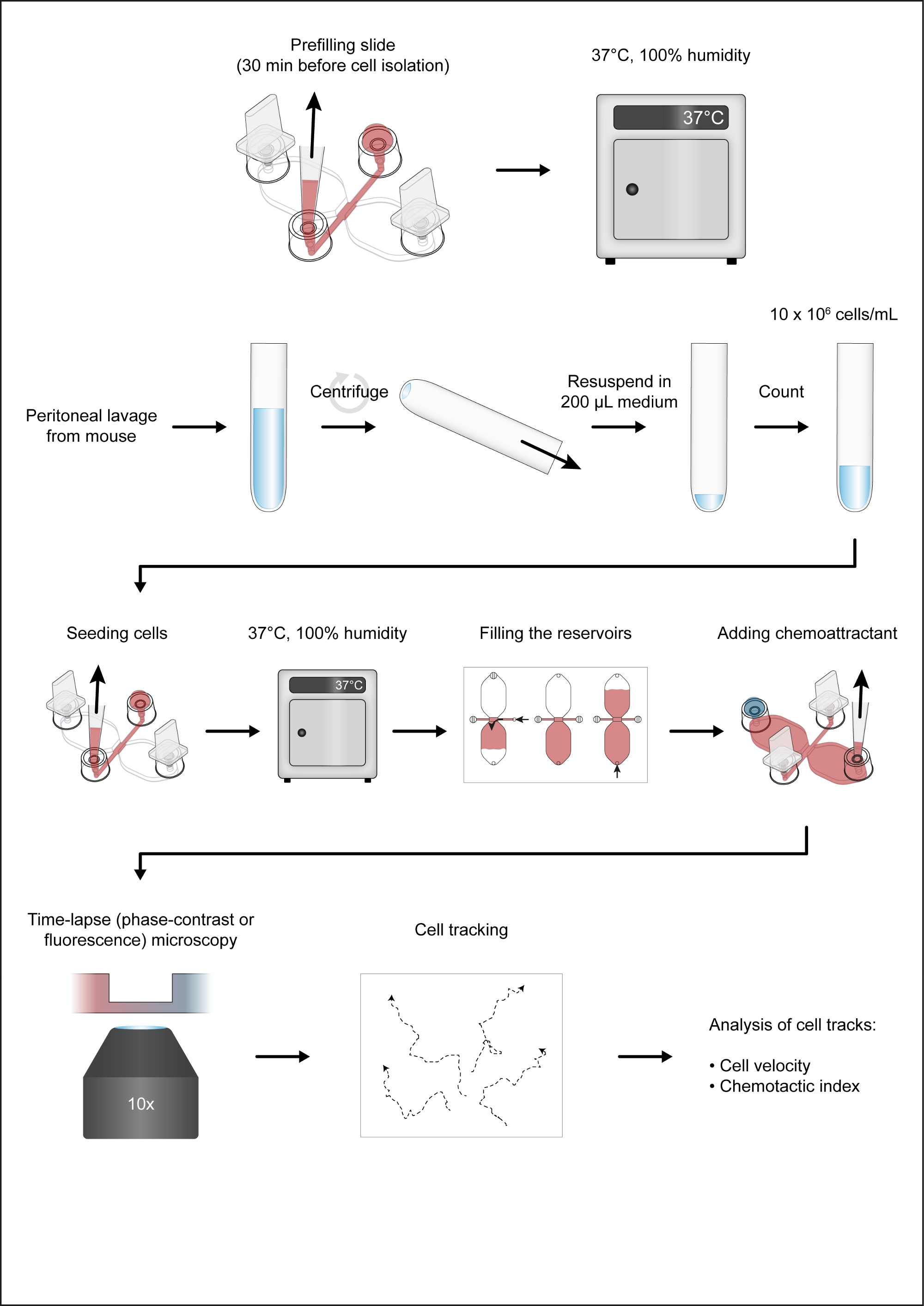
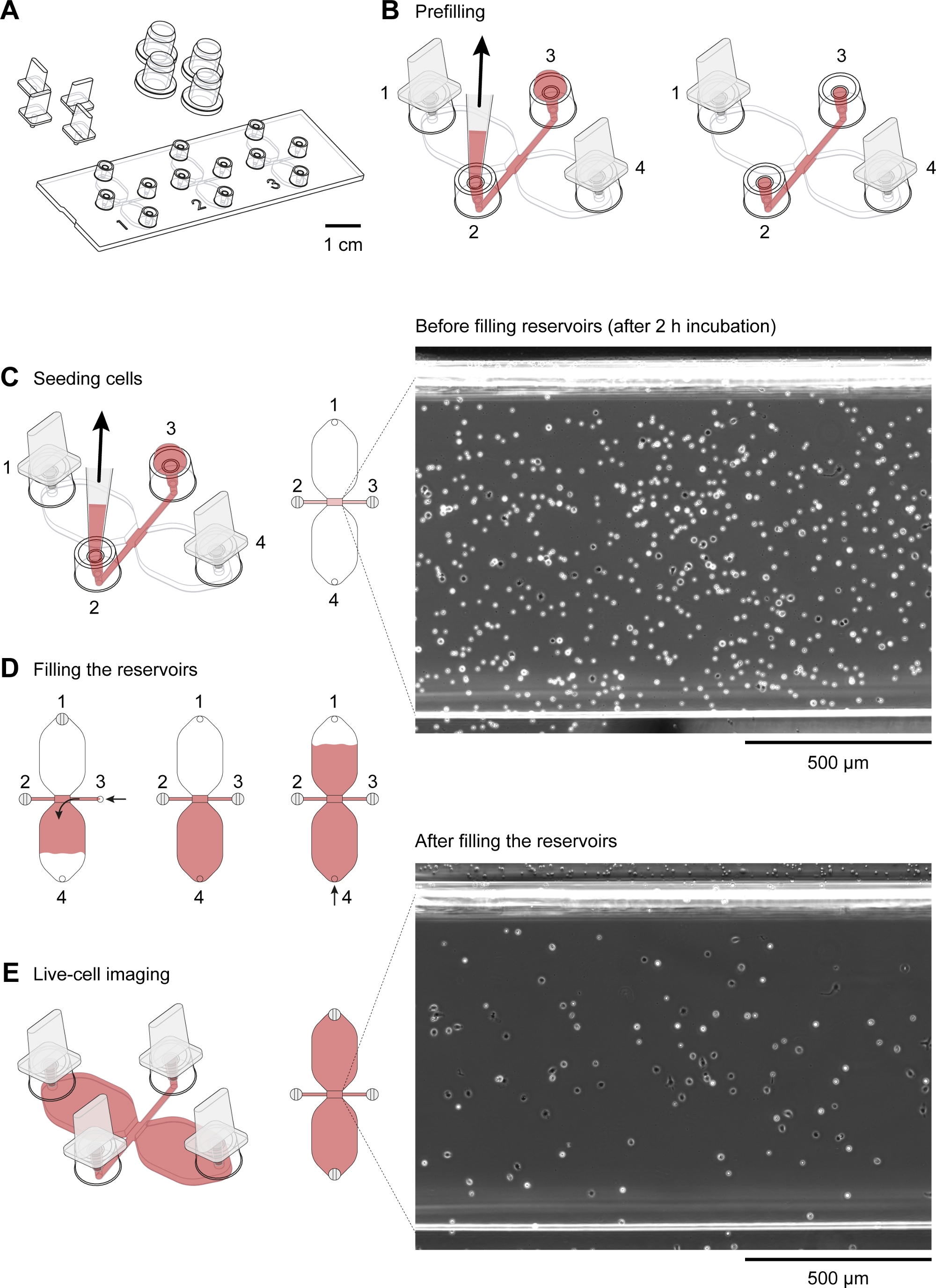
Ein schematisches Diagramm des Chemotaxis-Dias, das für die Zeitraffer-Videomikroskopie von peritonealen Makrophagen verwendet wird, die in einem chemotaktischen Gradienten wandern, ist in Abbildung 2Adargestellt. Die Rutsche enthält drei Chemotaxis-Kammern, von denen jede vier Füllöffnungen hat. Ports können mit den über der Folie gezeigten Steckern einzeln geschlossen werden. Alternativ kann eine nicht versiegelnde Kappe über einen unplugged Port gelegt werden, um die Sterilität aufrechtzuerhalten. Nach dem Einstecken der Ports 1 und 4 kann der Beobachtungsbereich (1 mm breit x 2 mm lang x 70 m hochkanalig, der die beiden Reservoirs verbindet) zwischen den Ports 2 und 3 mit Medium vorgefüllt werden, indem ein 15-L-Abfall in Port 3 platziert und mit einer 2–20-L-Volumenpipette an Port 2(Abbildung 2B)gesaugt wird. Eine Suspension von mausresidenten Peritonealzellen (10 x 106 Zellen/ml) wurde in den Beobachtungsbereich eingesät, indem ein 10 L-Tropfen der Suspension in Port 3 platziert und langsam an Port 2 angesaugt wurde (Abbildung 2C). Ein typisches Bild von Zellen, die im Beobachtungsbereich ausgesät wurden, die durch Phasenkontrastmikroskopie mit einer 10-fachen Objektivlinse aufgenommen wurden, ist in Abbildung 2Cdargestellt. Nach der Inkubation für 2-3 h wurde die Chemotaxis-Rutsche langsam mit Medium gefüllt (Abbildung 2D). Nach dem Schließen der Ports 1 und 2 wurde das Medium langsam über Port 3 injiziert, bis es aus Port 4 auftauchte. Als nächstes wurde der Stecker von Port 1 auf Port 3 umgeschaltet, und dann wurde das zweite Reservoir durch langsames Einspritzen von Medium über Port 4 gefüllt, bis er an Port 1 auftauchte. In diesem Stadium wurden die Zellen im Beobachtungsbereich mit einem invertierten Mikroskop erneut untersucht (Abbildung 2E). Durch den Vergleich der Bilder kurz vor (Abbildung 2C) und nach der Füllung der Reservoirs (Abbildung 2E) waren bis zu zwei Drittel der Zellen aus dem Beobachtungsgebiet ausgewaschen worden. Im Allgemeinen wurden schwach anhängliche CD19+ Zellen (B1-Zellen) ausgewaschen und die übrigen Zellen waren überwiegend F4/80+ Zellen (Makrophagen). Dies wurde durch fluoreszenzmikroskopische Mikroskopie nach der Kennzeichnung jedes Zelltyps mit fluoreszierend gekennzeichneten spezifischen Antikörpern nachgewiesen (Abbildung 4). In Abbildung 4Awurden frisch isolierte, mausresidente Peritonealzellen mit grünen fluoreszierenden Fluorophor-konjugierten Anti-F4/80-Antikörpern und roten fluoreszierenden Fluorophor-konjugierten Anti-CD19-Antikörpern gekennzeichnet, und die Zellkerne wurden mit einem blauen fluoreszierenden Nukleinsäurefleck gekennzeichnet. F4/80 ist ein spezifischer Marker für Mausmakrophagen30, während CD19 ein B-Zellmarker ist. Abbildung 4B zeigt F4/80+ Zellen, die durch die konfokale Mikroskopie der Spinnscheibe im Beobachtungsbereich einer Chemotaxiskammer abgebildet sind. Die Zellen wurden nach einem über Nacht chemotaxis-Assay durch Zeitraffer-Phasen-Kontrastmikroskopie aufgezeichnet.
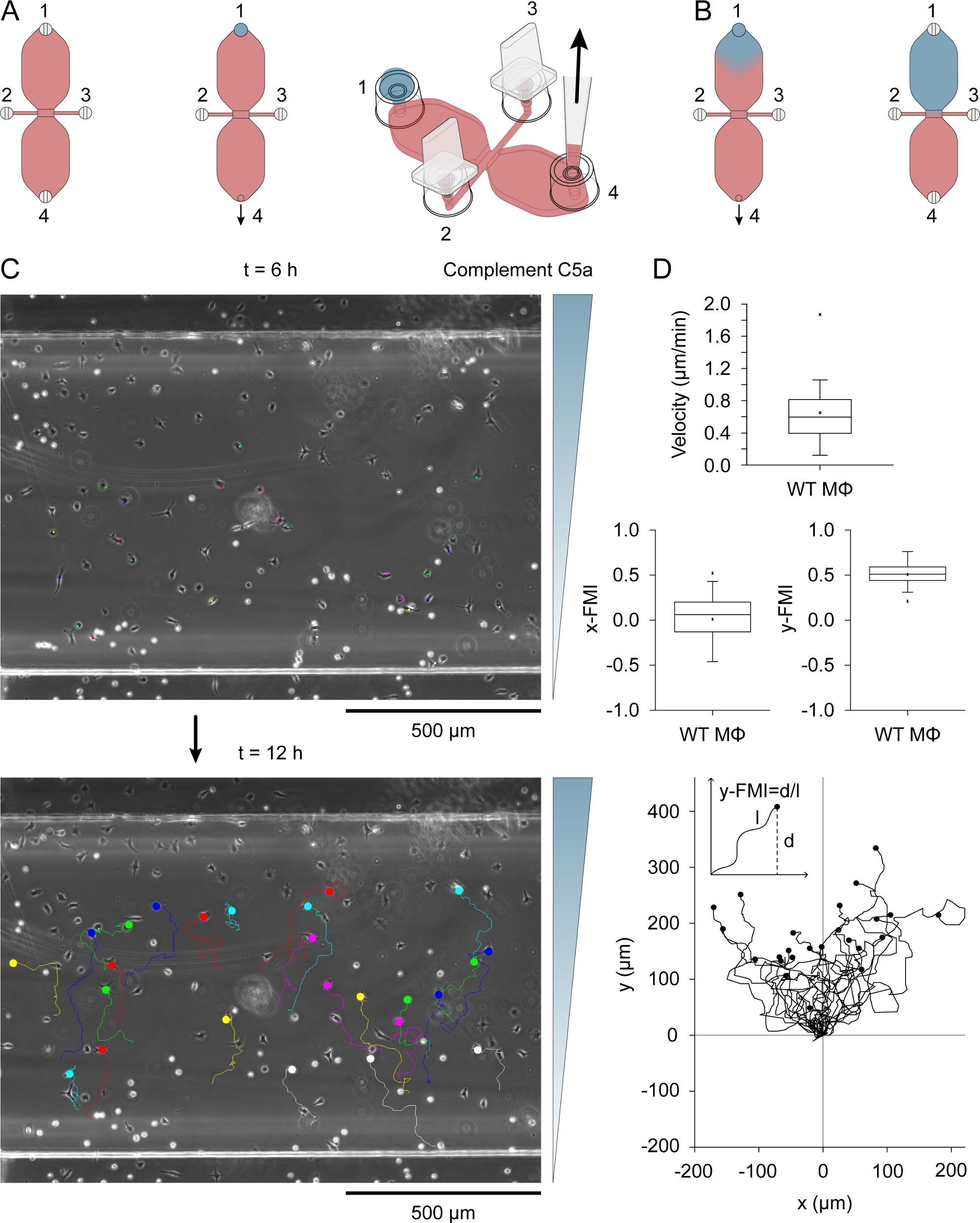
Komplement C5a (Chemoattractant) wurde in eines der beiden Reservoirs eingeführt, indem ein 15-L-Tropfen des Mediums, das 0,54 g/ml (rekombinante Maus) enthält, C5a und 10 g/ml Patent Blue V nach dem Einstecken der Ports 2 und 3 in den Füllanschluss 1 (Abbildung 3A)einführte. Das chemoattraktionant Medium wurde langsam durch langsames Aspirationen mit einer Pipette über Port 4 in das Reservoir gezogen. Abbildung 3B zeigt die Diffusion des blauen Farbstoffs nach dem Zeichnen des 15-L-Tropfens in ein Reservoir. Patent Blue V wurde als indirekter visueller Indikator für chemolockante Diffusion verwendet. Komplement-C5a-Moleküle sind deutlich größer als die von Patent Blue V (9,0 kDa versus 0,57 kDa) und diffundieren langsamer. Nach der Diffusion des Komplements C5a im Reservoir betrug seine Konzentration 0,2 g/ml (15 l/40 l [Reservoirvolumen] x 0,54 g/ml = 0,2 g/ml), was 22,5 nM entspricht. Ein bescheiden steiler Gefälle bildete sich nach 3 h über das Beobachtungsgebiet und nahm weiter zu, wo ein Maximum um etwa 12 h31erreicht wurde. Abbildung 3C zeigt die Migrationsspuren von Makrophagen, die in einem Komplement-C5a-Gradienten zwischen 6 und 12 h nach DemHinzufügen des Chemoattractants wandern. Zellgeschwindigkeit und chemotaktische Effizienz, indiziert als y-FMI (y-Forward-Migrationsindex; Bereich: -1 bis +1) und x-FMI, der einzelnen Makrophagen wurde aus den Migrationsdiagrammen berechnet (Abbildung 3D). Abbildung 3D zeigt auch ein Migrationsdiagramm, das nach der Normalisierung des Startpunkts jeder Migrationsspur auf X = 0 und Y = 0 unter den Boxplots erstellt wurde. Der Einset im Migrationsdiagramm zeigt, wie der y-FMI für jede Migrationsspur berechnet wurde.

Abbildung 1: Der Arbeitsablauf des Chemotaxis-Assays. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 2: Handhabung von Chemotaxis-Dias. (A) Eine 3D-Ansicht einer Chemotaxis-Folie mit vier Steckern und vier Kappen. Die Rutsche enthält drei Chemotaxis-Kammern, von denen jede aus zwei 40-L-Reservoirs besteht, die durch einen 1 mm x 2 mm Kanal verbunden sind, der 70 m hoch ist und als Beobachtungsgebiet bezeichnet wird. (B) Der Verbindungskanal erstreckt sich an beiden Enden bis zu den Füllöffnungen 2 und 3. Nach dem Einsetzen von Steckern in die Füllöffnungen 1 und 4 wurde der Beobachtungsbereich mit Medium (rot) vorgefüllt, indem ein Tropfen Medium auf Port 3 aufgebracht und an Port 2 mit einer Pipettespitze von 10–200 l angesaugt wurde. Anschließend wurden die Kappen auf die Ports 2 und 3 aufgebracht, bevor der Schlitten bei 37 °C inkubiert und die Zellsuspension vorbereitet wurde. (C) Das Beobachtungsgebiet, in dem sich der chemolockige Gradient bildete, wurde mit Makrophagen gesät, indem ein 10 L Tropfen der mausansässigen Peritonealzellen an Port 3 aufgebracht und langsam an Port 2 angesaugt wurde. Die Rutsche wurde dann in einer Feuchtigkeitskammer bei 37 °C für 2-3 h inkubiert. Das auf der rechten Seite gezeigte Phasenkontrastbild, das über eine 10-fache Objektivlinse erhalten wurde, zeigt Peritonealzellen nach Aussaat und Inkubation bei 37 °C für 2 h. Scale bar = 500 m. (D) Chemotaxis-Kammern wurden durch Stecköffnungen 1 und 2 mit Medium gefüllt und dann langsam über Port 3 injiziert, bis es an Port 4 auftauchte. Eine langsame und gleichmäßige Befüllung kann durch Drehen des Volumeneinstellrings einer 20–100 L Volumenpipette erreicht werden. Nach dem Befüllen des ersten Reservoirs kann das zweite Reservoir durch Einstecken der Ports 2 und 3 gefüllt werden und dann langsam Medium an Port 4 injizieren, bis es an Port 1 auftaucht. (E) Phasenkontrastbild des oben gezeigten Beobachtungsbereichs (C) nach dem Befüllen der beiden Reservoirs. Maßstabsleiste = 500 m. Grafische Elemente von Elias Horn. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 3: Chemotaxis-Assay. (A) Chemoattractant wurde in eines der beiden Reservoirs einer Chemotaxiskammer eingeführt, indem ein 15 l Tropfen Medium mit 0,54 g/ml Komplement C5a und 10 g/ml Patent Blue V auf den Füllanschluss 1 aufgebracht wurde, gefolgt von einer langsamen Aspiration an Port 4. (B) Zunächst, nachdem das blaue, chemolockhaltige Medium in das Reservoir gezogen wurde, hatte es eine grob umgekehrte Tropfenform und diffundierte dann langsam durch das Reservoir. (C) Migrationsspuren von Makrophagen, die in einem Chemoattraktions (Komplement C5a) Gradienten zwischen 6–12 h nach Dereinführung von Chemoattractant in einem der Reservoirs wandern. Die Richtung des Farbverlaufs ist auf der rechten Seite angezeigt. Das Ende jeder Migrationsspur wird durch einen gefüllten Kreis angezeigt. (D) Blox-Plots mit Geschwindigkeit, x-FMI (x-Forward-Migrationsindex) und y-FMI (y-forward migration index), ein Index der chemotaktischen Effizienz, der von -1 bis +1 reicht. Die Daten wurden durch die Analyse von 25 Makrophagen-Migrationsspuren ermittelt. Makrophagen in der unteren Hälfte des Beobachtungsbereichs und die Darstellung einer Verschiebung von mindestens einer Zellbreite über 6 h wurden zufällig für die Analyse ausgewählt. Unten ist ein Diagramm von Migrationsspuren nach der Normalisierung des Startpunkts auf X = 0 und Y = 0. Der Chemotaxis-Index (y-FMI) wurde berechnet, indem die Nettoverschiebung entlang der Y-Achse (d) durch die akkumulierte Länge (l) des Migrationspfads dividiert wurde, wie schematisch dargestellt. Grafische Elemente in den Paneelen A und B von Elias Horn. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 4: Fluoreszierende Bilder von lebenden Maus-Resident-Peritonealzellen, die durch drehende Plattenkonfokalmikroskopie gewonnen werden. (A) Erweitertes Fokusbild (hellste Punktverschmelzung aller Z-Ebenen) von frisch isolierten Mausperitonealzellen, die mit grünen fluoreszierenden Anti-F4/80-Antikörpern (Makrophagenmarker), roten fluoreszierenden Anti-CD19-Antikörpern (B-Zellmarker) und einem blauen fluoreszierenden Nukleinsäurefleck beschriftet sind. Skala bar = 10 m . (B) Schnappschuss (einzelne Z-Ebene) von F4/80+ Zellen (Makrophagen) im Beobachtungsbereich einer Chemotaxis-Kammer, die nach einem über Nacht erfolgten Chemotaxis-Assay aufgenommen wurde. Die Zellen wurden mit grünen fluoreszierenden Anti-F4/80-Antikörpern und einem blauen fluoreszierenden Nukleinsäurefleck gekennzeichnet. Die Komplement-Gradienten C5a und Patent Blue V wurden durch das Zellbeschriftungsverfahren ausgewaschen, was erklärt, warum das obere Reservoir im Schaltplan der Chemotaxiskammer nicht blau ist. Maßstabsleiste = 10 m. Grafikelement zur Verfügung gestellt von Elias Horn. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
Diskussion
Intravitale Bildgebung stammt aus dem 19. Jahrhundert und bietet eine Möglichkeit, das Verhalten lebender Immunzellen in ihrer natürlichen Umgebung zu untersuchen. Doch selbst mit den heutigen fortgeschrittenen Mikroskopie- und Gentechniken ist es schwierig, die Reaktion von Zellen auf bestimmte Chemoattractants in vivo zu untersuchen. Um dieses Problem zu umgehen, entwickelte Boyden18 Transwell-Assays in den 1960er Jahren, aber diese Endpunkt-Assays lieferten keine Visualisierung, wie Zellen tatsächlich zu Chemolockanten migrierten, was es schwierig machte, Chemokinese zu unterscheiden, die zufällige Migration durch einen chemischen Cue32und Chemotaxis, Migration in Richtung höherer Konzentrationen chemischer Reize voneinander33. Dieses Problem wurde gelöst, indem verschiedene offene Kammern mit einer Brücke, typischerweise 1 mm breit, zwischen zwei Reservoirs und zugänglich durch eine Objektivlinse21,22,23. Durch das Anwenden eines invertierten Deckelschlupfs, der mit anhaftenden Zellen gesät ist, schließt er die Kammern und chemoattractant, das einem der Reservoirs zugesetzt wird, diffundiert über die Brücke zum gegenüberliegenden Reservoir, wodurch ein Konzentrationsgradient entsteht. Hier beschreiben wir einen Chemotaxis-Assay nach dem gleichen Prinzip, aber mit einer geschlossenen Kammer mit vier Füllöffnungen. Mit diesem System und der Zeitraffer-Phasen-Kontrastmikroskopie haben wir einen Assay entwickelt, um bildmausresidente Peritonealmakrophagen zu bilden, die in einer chemotaktischen Komplementierung C5a Gradient31,34,35,36wandern. Dieser Assay, kombiniert mit Knockout-Maus-Modellen, erwies sich als entscheidend für die Untersuchung der Rollen verschiedener Rho GTPases und motorischer Proteine in Makrophagenmorphologie, Beweglichkeit und Chemotaxis31,34,35,36,37. Wir haben diesen Ansatz auch verwendet, um menschliche periphere Blutmonozyten abzubilden, die auf einer 2D-Oberfläche oder in einer 3D-Kollagen-Matrix Typ I38wandern. Darüber hinaus eignet sich der Assay für Mausknochenmark-abgeleitete Makrophagen oder Makrophagen, die aus bedingt verewigten myeloischen Vorläuferzellen39,40abgeleitet werden. Wir haben zuvor Polytetrafluorethylen (PTFE) Taschen mit Luer-Adaptern verwendet, um Knochenmarkzellen zu kultizieren und Makrophagen zu erhalten34. Der Vorteil von PTFE-Beuteln ist, dass die Zellen nach 20–30 min leicht resuspendiert und einsatzbereit sein können. Beachten Sie, dass wir den Chemotaxis-Dia-Beobachtungsbereich vorfüllen, bevor wir die Zellen einführen. Dieser Ansatz hat den Vorteil, dass unerwünschte Luftblasen anschließend (mit variablem Erfolg) ausgespült werden können und der vorgetränkte Beobachtungsbereich die langsame Einführung einer Zellsuspension durch Pipettieren ermöglicht. Die Vorfüllung erhöht jedoch die Wahrscheinlichkeit, dass das Medium teilweise in eines oder beide der flankierenden Reservoirs fließt, was die Aussaat von Zellen außerhalb des Beobachtungsgebiets fördert. Alternativ kann die Zellsuspension direkt in ein trockenes Beobachtungsgebiet geleitet werden, unerwünschte Luftblasen können jedoch nicht später ausgestoßen werden.
Die Peritonealhöhle der Maus enthält zwei Hauptpopulationen von Zellen: F4/80+ Makrophagen und (kleinere) CD19+ B-Zellen, im Verhältnis von etwa 1:2 (Abbildung 4A). Diese beiden Zellpopulationen machen über 95% der Peritonealhöhlenzellen aus, während die verbleibenden F4/80-/CD19- Zellen in der Regel als CD11c+ Zellen (dendritische Zellen) oder CD3+ Zellen (T-Zellen) identifiziert werden können. Schwach anhaftende B-Zellen werden während der Befüllung der Reservoirs mit dem Medium(Abbildung 2) aus dem Beobachtungsbereich gewaschen. Nach Zugabe von Chemoattractant zu einem der beiden Reservoirs kann die Zeitraffer-Phasen-Kontrastmikroskopie verwendet werden, um die verbleibenden Zellen (Makrophagen) abzubilden, die in einem sich entwickelnden Chemoattractantgradienten wandern. Die Bildung des Komplement-C5a-Gradienten im Beobachtungsgebiet, durch Diffusion von einem Reservoir zum anderen, kann mit einem Fluoreszenzfarbstoff mit ähnlichem Molekulargewicht simuliert werden. Ein guter Ersatz für rekombinante Mauskomplement C5a (vorhergesagtes Molekulargewicht, 9,0 kDa) ist fluoreszierend dextran (10 kDa)31gekennzeichnet. Mittels konfokaler Mikroskopie kann der Fluoreszenzgradient im schmalen Kanal (Beobachtungsbereich), der die beiden Reservoirs des Chemotaxis-Dias verbindet, in festen Intervallen gemessen werden und Konzentrationsprofile an den verschiedenen Zeitpunkten24,31dargestellt werden. Wir fügen dem Chemolock-Medium routinemäßig einen nicht fluoreszierenden, blauen Farbstoff (Patent Blue V) hinzu, um einen praktischen visuellen Indikator für Diffusion und Gradientenbildung zu liefern. Innerhalb von 1 h nach der Einführung von 15 l blauem, chemolockhaltigem Medium in ein Reservoir erscheint das Reservoir einheitlich blau und nach Diffusionsgesetzen bildet sich ein Gradient über das enge Beobachtungsgebiet, das die Reservoirs verbindet (Abbildung 3B). Mehrere Tage sind erforderlich, damit der Gelöste (blauer Farbstoff oder Chemoattractant) gleichmäßig verteilt wird.
Die Fluoreszenzmikroskopie kann durch die Phasenkontrastmikroskopie ersetzt werden, die Vorteile für die automatisierte Zellverfolgung bietet, da fluoreszierend markierte Zellen leicht vom Hintergrund unterschieden werden können. Ein weiterer Vorteil ist, dass bestimmte Populationen von Immunzellen selektiv nach der Kennzeichnung von Oberflächenmarkern mit fluoreszierenden Antikörpern verfolgt werden können. Wir verwendeten diesen Ansatz, um menschliches peripheres Blut CD14+ Zellen (Monozyten) zu bilden, die in einem chemotaktischen fMLP (N-Formylmethionin-Leucyl-Phenylalanin)Gradienten 38wandern. In ähnlicher Weise könnten fluoreszierende Anti-F4/80-Antikörper verwendet werden, um Mausmakrophagen abzubilden, die in einem chemotaktischen Komplement C5a-Gradienten migrieren. Phototoxizität ist ein potenzieller Nachteil der Verwendung von Fluoreszenz-Bildgebung41. Dies kann mit verschiedenen Mitteln reduziert werden42, einschließlich der Verwendung von Fluorophoren mit längeren Wellenlängen angeregt und Zugabe von Antioxidantien auf das Medium. Alternativ konnten markierte Zellen zunächst durch Fluoreszenzmikroskopie identifiziert und anschließend durch Zeitraffer-Phasen-Kontrastmikroskopie abgebildet werden. In der Praxis können jedoch Zellen, die sich bei mäßig niedrigen Geschwindigkeiten bewegen, wie z. B. 1 m/min (Makrophagen) oder 4 m/min (Monozyten), in regelmäßigen Abständen durch Fluoreszenzmikroskopie in Minutenintervallen dargestellt werden, was gut verträglichist 38. Wir haben zuvor Fluoreszenzmikroskopie und die hier beschriebene Chemotaxis-Folie für 3D-Chemotaxis-Assays38,43verwendet. In diesem Fall wurden beide Reservoirs mit einem mittel- und 15-L-Chemo-haltigen Medium vorgefüllt, das unmittelbar vor der langsamen Pipettierung fluoreszierend markierter Zellen, die in einem mittelhaltigen Kollagentyp I in den Beobachtungsbereich schweben, in eines der Reservoirs gezogen wurde. Der schwierige Teil dieses Verfahrens ist der Umgang mit Kollagen Typ I, der in saurer Lösung konzentriert ist. Der pH-Wert der Kollagenlösung muss durch Zugabe einer alkalischen Lösung neutralisiert werden, bevor die eiskalte Kollagenlösung mit der Zellsuspension vermischt wird. Die Übertragung der Kollagen-Zell-Mischung auf einen Inkubator bei 37 °C wird eine Kollagenpolymerisation initiieren. Während der Inkubation sollte der Schlitten langsam um seine lange Achse gedreht werden, so dass die Zellen gleichmäßig in x-, Y- und Z-Achse verteilt bleiben, während das Kollagen zu einem Gel polymerisiert. Eine verwandte geschlossene Chemotaxis-Folie, die für 3D-Chemotaxis-Assays geeignet ist, mit sechs Steckern statt vier Steckern, wurde kürzlich29beschrieben. Dieses System ermöglicht es, das Kollagen-Zell-Gemisch in den Beobachtungsbereich einzuführen, bevor jedes der flankierenden Reservoirs unabhängig gefüllt wird, da jedes Reservoir über zwei Füllöffnungen und nicht über einen einzigen Port verfügt.
Zusammenfassend beschreiben wir einen Echtzeit-Chemotaxis-Assay, der die Visualisierung von Zellen ermöglicht, die in einem chemotaktischen Gradienten über einen Zeitraum von 6 oder mehr Stunden navigieren. Dabei konzentrieren wir uns auf Makrophagen, die bei entzündlichen Erkrankungen eine wichtige Rolle spielen, aber in Echtzeit-Chemotaxis-Assays unterrepräsentiert sind, verglichen mit schneller endenden Zellen wie Neutrophilen und Dictyostelium-Amöben.
Offenlegungen
Die Autoren haben nichts zu verraten.
Danksagungen
Diese Arbeit wurde durch ein Stipendium (HA 3271/3-2) der DFG (Deutsche Forschungsgemeinschaft) unterstützt.
Materialien
| Name | Company | Catalog Number | Comments |
| µ-Slide (anodized aluminium) rack | Ibidi, Martinsried, Germany | 80003 | Autoclavable stackable rack for channel slides |
| µ-Slide Chemotaxis 2D (chemotaxis slide) | Ibidi, Martinsried, Germany | 80306 | Slide containing chemotaxis chambers (tissue culture treated) |
| 100x penicillin/streptomycin | Thermo Fisher Scientific | 15140122 | Used as supplement for RPMI 1640 media |
| 10-100 µL pipette with volume control ring | Eppendorf | 3123000047 | Eppendorf Research plus pipette |
| 10-200 µL pipette tips | Greiner Bio-One International | 739261 | Pipette tips with beveled tips (96 pieces per rack: sterile) |
| 14 mL polypropylene round bottom tubes | BD Falcon | 352059 | Used to collect peritoneal cells |
| 14-bit Hamamatsu C9100-50 Electron Multiplying-Charged Couple Device (EM-CCD) peltier-cooled camera | Hamamatsu Photonics Inc., Japan | EM-CCD camera of the spinning disk confocal microscope system | |
| 2-20 µL pipette with volume control ring | Eppendorf | 3123000039 | Eppendorf Research plus pipette |
| 24 G plastic catheter | B Braun Mesungen AG, Germany | 4254503-01 | Used for peritoneal lavage |
| 405 nm solid state laser, 50 mW | Perkin Elmer, Rodgau, Germany | Laser (405 nm) source of spinning disk confocal microscope system | |
| 488 nm solid state laser, 50 mW | Perkin Elmer, Rodgau, Germany | Laser (488 nm) source of spinning disk confocal microscope system | |
| 561 nm solid state laser, 50 mW | Perkin Elmer, Rodgau, Germany | Laser (561 nm) source of spinning disk confocal microscope system | |
| Alexa Fluor 488-conjugated rat (IgG2a) monoclonal (clone BM8) anti-mouse F4/80 antibody | Thermo Fisher Scientific | MF48020 | Mouse macrophage marker and plasma membrane label |
| Alexa Fluor 594-conjugated rat (IgG2a) monoclonal (clone 6D5) anti-mouse CD19 antibody | BioLegend | 115552 | Mouse B cell marker |
| C-Chip disposable (improved Neubauer) hemocytometer | NanoEnTek (distributed by VWR International) | 631-1098 | Used to count cells |
| CSU-X1 spinning disk scanner | Yokogawa Electric Corporation, Japan | Nipkow spinning disk unit | |
| Hank’s buffered salt solution without Ca2+ and Mg2+ | Thermo Fisher Scientific | 14170120 | Used for peritoneal lavage |
| Heat-inactivated fetal bovine serum | Thermo Fisher Scientific | 10082139 | Used as supplement for RPMI 1640 media |
| Hoechst 34580 | Thermo Fisher Scientific | H21486 | Cell permeable, blue fluorescent nucleic acid stain |
| ImageJ (image processing and analysis in Java) | National Institutes of Health (NIH) | Image analysis software | |
| Lipopolysaccharides from Escherichia coliO111:B4 | Sigma-Aldrich | L4391-1MG | Toll-like receptor 4 ligand |
| Nikon Eclipse Ti inverse microscope | Nikon, Japan | Inverted microscope | |
| Patent Blue V, sodium salt | Sigma-Aldrich | 21605-10G | Blue-colored dye used as visual indicator of gradient formation |
| Recombinant mouse complement C5a protein | R&D Systems | 2150-C5-025 | Chemoattractant for mouse macrophages |
| RPMI 1640 medium containing 20 mM Hepes | Sigma-Aldrich | R7388 | Basis medium for assays |
| UltraVIEW Vox 3D live cell imaging system + Volocity software | Perkin Elmer, Rodgau, Germany | Spinning disk confocal microscope system | |
| Zeiss LSM 510 + Axiovision software | Carl Zeiss Microscopy, Oberkochen, Germany | Confocal laser scanning microscope (LSM) adapted for phase-contrast microscopy |
Referenzen
- Lammermann, T., Germain, R. N. The multiple faces of leukocyte interstitial migration. Seminars in Immunopathology. 36, 227-251 (2014).
- Lammermann, T., Sixt, M. Mechanical modes of 'amoeboid' cell migration. Current Opinion in Cell Biology. 21, 636-644 (2009).
- Woodham, E. F., Machesky, L. M. Polarised cell migration: intrinsic and extrinsic drivers. Current Opinion in Cell Biology. 30, 25-32 (2014).
- Devreotes, P. N., et al. Excitable Signal Transduction Networks in Directed Cell Migration. Annual Review of Cell and Developmental Biology. 33, 103-125 (2017).
- Kamp, M. E., Liu, Y., Kortholt, A. Function and Regulation of Heterotrimeric G Proteins during Chemotaxis. International Journal of Molecular Sciences. 17 (1), 90(2016).
- Miao, Y., et al. Wave patterns organize cellular protrusions and control cortical dynamics. Molecular Systems Biology. 15, 8585(2019).
- Ridley, A. J., et al. Cell migration: integrating signals from front to back. Science. 302, 1704-1709 (2003).
- Lammermann, T., et al. Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature. 453, 51-55 (2008).
- Mullins, R. D., Heuser, J. A., Pollard, T. D. The interaction of Arp2/3 complex with actin: nucleation, high affinity pointed end capping, and formation of branching networks of filaments. Proceedings of the National Academy of Sciences of the United States of America. 95, 6181-6186 (1998).
- Leithner, A., et al. Diversified actin protrusions promote environmental exploration but are dispensable for locomotion of leukocytes. Nature Cell Biology. 18, 1253-1259 (2016).
- McDonald, B., et al. Intravascular danger signals guide neutrophils to sites of sterile inflammation. Science. 330, 362-366 (2010).
- Leber, T. Ueber die Entstehung der Entzündung und die Wirkung der entzündungserregenden Schädlichkeiten. Fortschritte der Medizin. 6, 460-464 (1888).
- Tauber, A. I. Metchnikoff and the phagocytosis theory. Nature Reviews Molecular Cell Biology. 4, 897-901 (2003).
- Clark, E. R., Linton Clark, E. Reactions of cells in the tail of amphibian larvae to injected croton oil (aseptic inflammation). American Journal of Anatomy. 27, 221-254 (1920).
- Clark, E. R., Linton Clark, E. The reaction of living cells in the tadpole's tail toward starch, agar-agar, gelatin, and gum arabic. The Anatomical Record. 24, (1922).
- Comandon, J. Phagocytose in vitro des Hématozoaires du Calfat (enregistrement cinématographique). Comptes Rendus Hebdomadaires des Séances et Mémoires de la Société de Biologie. 69, 314-316 (1917).
- McCutcheon, M. Chemotaxis in leukocytes. Physiological Reviews. 26, 319-336 (1946).
- Boyden, S. The chemotactic effect of mixtures of antibody and antigen on polymorphonuclear leucocytes. The Journal of Experimental Medicine. 115, 453-466 (1962).
- Horwitz, D. A., Garrett, M. A. Use of leukocyte chemotaxis in vitro to assay mediators generated by immune reactions. I. Quantitation of mononuclear and polymorphonuclear leukocyte chemotaxis with polycarbonate (nuclepore) filters. Journal of Immunology. 106, 649-655 (1971).
- Bignold, L. P. A novel polycarbonate (Nuclepore) membrane demonstrates chemotaxis, unaffected by chemokinesis, of polymorphonuclear leukocytes in the Boyden chamber. Journal of Immunological Methods. 105, 275-280 (1987).
- Zigmond, S. H. Ability of polymorphonuclear leukocytes to orient in gradients of chemotactic factors. The Journal of Cell Biology. 75, 606-616 (1977).
- Zicha, D., Dunn, G. A., Brown, A. F. A new direct-viewing chemotaxis chamber. Journal of Cell Science. 99, Pt 4 769-775 (1991).
- Muinonen-Martin, A. J., Veltman, D. M., Kalna, G., Insall, R. H. An improved chamber for direct visualisation of chemotaxis. PLoS One. 5, 15309(2010).
- Zengel, P., et al. mu-Slide Chemotaxis: a new chamber for long-term chemotaxis studies. BMC Cell Biology. 12, 21(2011).
- Valentim, A. M., Guedes, S. R., Pereira, A. M., Antunes, L. M. Euthanasia using gaseous agents in laboratory rodents. Lab Animal. 50, 241-253 (2016).
- Franks, N. P. General anaesthesia: from molecular targets to neuronal pathways of sleep and arousal. Nature Reviews. Neuroscience. 9, 370-386 (2008).
- Cordelieres, F. P., et al. Automated cell tracking and analysis in phase-contrast videos (iTrack4U): development of Java software based on combined mean-shift processes. PLoS One. 8, 81266(2013).
- Zantl, R., Horn, E. Chemotaxis of slow migrating mammalian cells analysed by video microscopy. Methods in Molecular Biology. 769, 191-203 (2011).
- Biswenger, V., et al. Characterization of EGF-guided MDA-MB-231 cell chemotaxis in vitro using a physiological and highly sensitive assay system. PLoS One. 13, 0203040(2018).
- Austyn, J. M., Gordon, S. F4/80, a monoclonal antibody directed specifically against the mouse macrophage. European Journal of Immunology. 11, 805-815 (1981).
- Hanley, P. J., et al. Motorized RhoGAP myosin IXb (Myo9b) controls cell shape and motility. Proceedings of the National Academy of Sciences of the United States of America. 107, 12145-12150 (2010).
- Wilkinson, P. C. Cell Locomotion and Chemotaxis: Basic Concepts and Methodological Approaches. Methods. 10, 74-81 (1996).
- Pfeffer, W. Locomotorische Richtungsbewegungen durch chemische Reize. Untersuchungen aus dem Botanischen Institut zu Tübingen. 1, 363(1884).
- Konigs, V., et al. Mouse macrophages completely lacking Rho subfamily GTPases (RhoA, RhoB, and RhoC) have severe lamellipodial retraction defects, but robust chemotactic navigation and altered motility. The Journal of Biological Chemistry. 289, 30772-30784 (2014).
- Horsthemke, M., et al. Multiple roles of filopodial dynamics in particle capture and phagocytosis and phenotypes of Cdc42 and Myo10 deletion. The Journal of Biological Chemistry. 292, 7258-7273 (2017).
- Bachg, A. C., et al. Phenotypic analysis of Myo10 knockout (Myo10(tm2/tm2)) mice lacking full-length (motorized) but not brain-specific headless myosin X. Scientific Reports. 9, 597(2019).
- Horsthemke, M., et al. A novel isoform of myosin 18A (Myo18Agamma) is an essential sarcomeric protein in mouse heart. The Journal of Biological Chemistry. 294, 7202-7218 (2019).
- Bzymek, R., et al. Real-time two- and three-dimensional imaging of monocyte motility and navigation on planar surfaces and in collagen matrices: roles of Rho. Scientific Reports. 6, 25016(2016).
- Wang, G. G., et al. Quantitative production of macrophages or neutrophils ex vivo using conditional Hoxb8. Nature Methods. 3, 287-293 (2006).
- Gran, S., et al. Imaging, myeloid precursor immortalization, and genome editing for defining mechanisms of leukocyte recruitment in vivo. Theranostics. 8, 2407-2423 (2018).
- Magidson, V., Khodjakov, A. Circumventing photodamage in live-cell microscopy. Methods in Cell Biology. 114, 545-560 (2013).
- Icha, J., Weber, M., Waters, J. C., Norden, C. Phototoxicity in live fluorescence microscopy, and how to avoid it. BioEssays : News and Reviews in Molecular, Cellular and Developmental Biology. 39 (8), 1700003(2017).
- Isfort, K., et al. Real-time imaging reveals that P2Y2 and P2Y12 receptor agonists are not chemoattractants and macrophage chemotaxis to complement C5a is phosphatidylinositol 3-kinase (PI3K)- and p38 mitogen-activated protein kinase (MAPK)-independent. The Journal of Biological Chemistry. 286, 44776-44787 (2011).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten