Method Article
ניתוח בסיוע Transillumination של שלבים ספציפיים של מחזור האפיתל החצי-שנתי של העכבר לניתוחים Immunostaining במורד הזרם
In This Article
Summary
פרוטוקול זה מתאר מיקרו-דיסקציה בסיוע טרנסילומינציה של מקטעים של צינוריות עכבר בוגרות למחצה המייצגות שלבים ספציפיים של מחזור אפיתל חצי-היקפי, וסוגי תאים בו, וחיסון לאחר מכן של תכשירי סקווש ומקטעי צינור שלמים.
Abstract
Spermatogenesis הוא תהליך בידול ייחודי שבסופו של דבר מעורר את אחד מסוגי התאים המובהקים ביותר של הגוף, הזרע. התמיינות של תאי נבט מתרחשת בכיסים הציטופלזמיים של תאי סרטולי סומטיים המארחים 4 עד 5 דורות של תאי נבט בו זמנית ומתאמים ומסנכרנים את התפתחותם. לכן, ההרכב של סוגי תאי נבט בתוך חתך הוא קבוע, ואסוציאציות תאים אלה ידועים גם בשם שלבים (I-XII) של מחזור האפיתל החצי-חצי. חשוב לציין, ניתן לזהות שלבים גם מצינורות חצי-חצי שלמים המבוססים על מאפייני ספיגת האור/פיזור הדיפרנציאליים שלהם שנחשפו על ידי טרנסילומינציה, והעובדה שהשלבים עוקבים זה אחר זה לאורך הצינור בסדר מספרי. מאמר זה מתאר שיטת מיקרו-דיסקציה בסיוע טרנסילומינציה לבידוד מקטעי צינורית חצי-חצי-היקפיים המייצגים שלבים ספציפיים של מחזור אפיתל חצי-היקפי של העכבר. תבנית ספיגת האור של צינוריות חצי-היקפיות נבדקת תחילה תחת מיקרוסקופ ניתוק, ולאחר מכן מקטעי צינורית המייצגים שלבים ספציפיים נחתכים ומשמשים ליישומים במורד הזרם. כאן אנו מתארים פרוטוקולים immunostaining עבור הכנות סקווש ספציפיות לשלב עבור מקטעי tubule שלמים. שיטה זו מאפשרת לחוקר להתמקד באירועים ביולוגיים המתרחשים בשלבים ספציפיים של spermatogenesis, ובכך לספק כלי ייחודי למחקרים התפתחותיים, טוקסיקולוגיים וציטולוגיים של זרע ומנגנונים מולקולריים בסיסיים.
Introduction
בידול של תאי נבט זכר מ spermatogonia דיפלואידית כדי זרע haploid בוגרת, כלומר, spermatogenesis, הוא תהליך מורכב המתרחש באפיתל של צינוריות למחצה באשכים של אדם בוגר מינית1. צאצאים מיטוטיים של זרע A1 הראשון לחלק חמש פעמים כדי להרחיב את האוכלוסייה המחויבת לבידול, ואז להזין מיוזיס כמו זרע שבסופו של דבר להוליד זרע haploid. בידול של זרעונים עגולים לתוך זרע, כלומר, spermiogenesis, כרוך בשינויים מורכבים במורפולוגיה הסלולר, כולל דחיסה גרעינית ובניית מבנים ספציפיים לזרע כגון האקרוזום ואת הלקאה. בעכבר, כל התהליך של spermatogenesis לוקח 35 ימים כדי להשלים2,3.
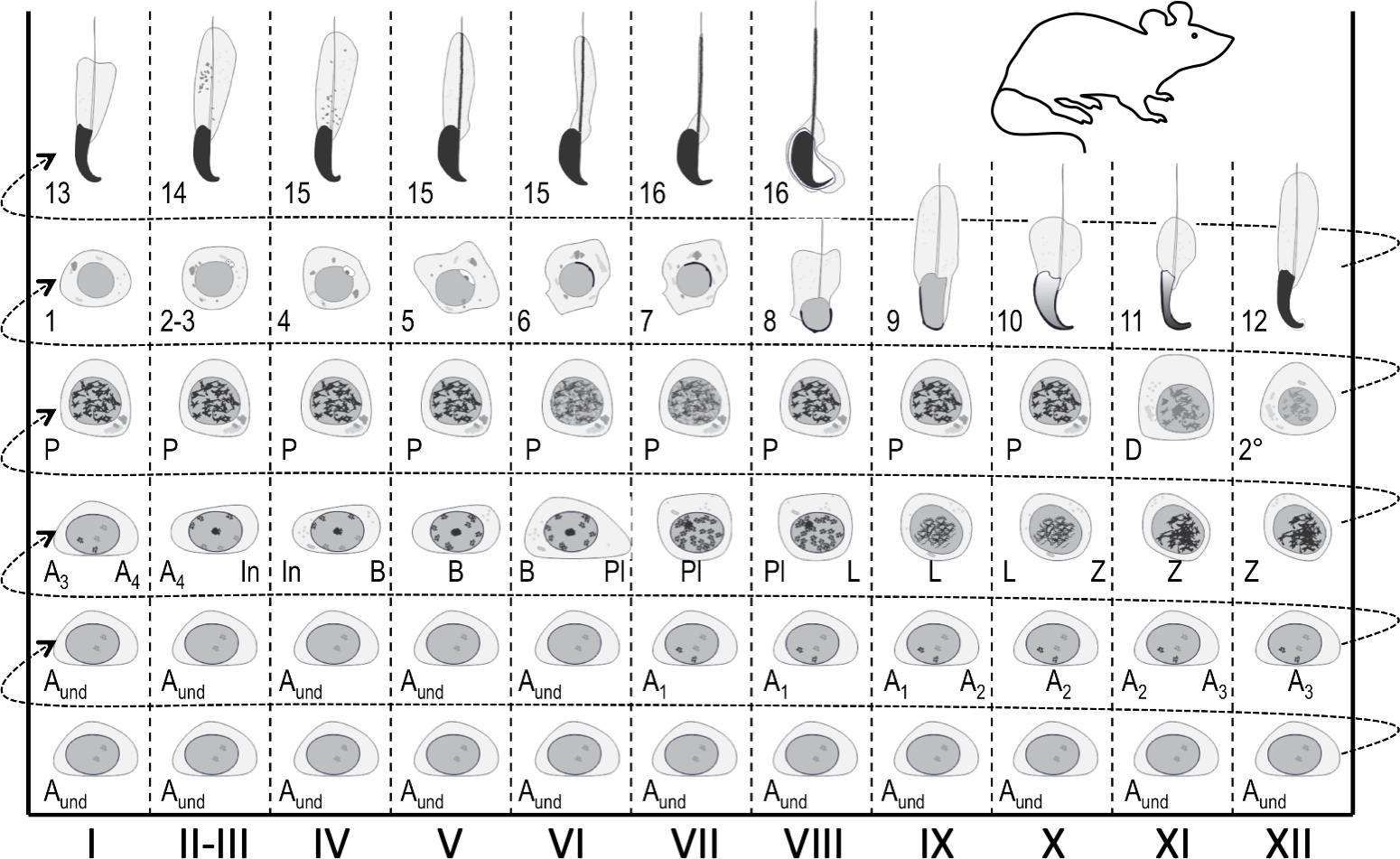
בכל אזור נתון, האפיתל החצי-שנתי מארח עד חמש קבוצות של תאי נבט מבדילים בתוספת תאי גזע/אב נבט ותאי סרטולי סומטיים1. תאי נבט מבדילים יוצרים שכבות קונצנטריות שהרכבן צפוי, ותאי הפלואיד בשלב נתון של התפתחות תמיד מקשרים לסוגים מסוימים של זרעוניםוזרעונים 4,5. לכן, כל חתך רוחב של צינור מארח קבוצות של תאי נבט של הרכב קבוע. אסוציאציות תאים ספציפיות אלה מוגדרות כשבי האפיתל החצי-שנתי. שלבים כשלעצמם אינם מציגים מצבים דמויי נקודת צ'ק-פוינט עומדים, אך מתפתחים ללא הרף כאשר הבידול של קבוצות תאי הנבט מתקדם בסנכרון1,2,6. בעכברים, ישנם 12 שלבים (I-XII)2 המסודרים באופן מגזרי לאורך הציר האורך של הצינורית החצי-שנתית, והם עוקבים זה אחר זה בסדר הגיוני ובכך יוצרים את גל האפיתל החצי-שנתי, או גל זרע7,8,9 ( איור1). השלמת spermatogenesis לוקח ארבעה מחזורים, ושכבות היררכיות או קבוצות של תאים נבט הבחנה בתוך כל חתך צינור חצי צינורי הם מחזור חצי שנתי אחד בנפרד זה מזה. אורך המחזור תלוי במינים ובלעכבר כל מחזור לוקח 8.6 ימים10.
ניתן לזהות את השלבים על בסיס ההרכב התאי והארגון של האפיתל החצי-חצי-חצי-היסטולוגי בסעיפי האשכים ההיסטולוגיים5 (איור 1 ואיור 2). עם זאת, ניתוח היסטולוגי הוא מייגע, זמן רב ודורש תיקון וכתמים, ולכן לא יכול להיות מיושם על רקמה חיה. חשוב לציין, בימוי יכול להתבצע גם על רקמה חיה תחת מיקרוסקופ דיסקציה על ידי ניצול דפוסי ספיגה/פיזור אור מובהקים המוצגים על ידי שלבים שונים של המחזור (איור 2). היכולת של כל שלב לספוג ולפזר אור היא יחסית לרמת עיבוי הכרומטין של זרעונים פוסט-מיוטיים מאוחרים שכל מארח שלב נתון ואריזת תאים אלה בחבילות7,11. בידול זרע, כלומר,spermiogenesis, מחולק עוד יותר 16 שלבים התפתחותיים, כולל 8 שלבים של זרע עגול (שלב 1-8) ו 8 שלבים של זרע מאריך (שלבים 9-16) בידול (איור 1). שלב 9-11 זרע מאריך (שלב IX-XI) להציג רק רמה נמוכה של עיבוי כרומטין וכתוצאה מכך כמות נמוכה של אור נספג. עיבוי כרומטין מתחיל בשלב 11 זרעונים (שלב XI), ובשלב 15-16 זרעונים מאריכים (שלב IV-VIII) מכילים כרומטין מרוכז במלואו, ולכן מפגינים ספיגת אור מקסימלית (איור 3). כרומטין צריך להיות מרוכז על מנת להיות ארוז בחוזקה לתוך ראש הזרע. גורמים נוספים התורמים לתבנית ספיגת האור הם מיקום של זרעונים מאריכים בתוך האפיתל (בסיסי לעומת אפיקלי) וחבילה של זרעונים מאריכים (מבוטא בשלבים II-V)11 (איור 3). חבילות נתפסות כנקודות באמצע הצינוריות ופסים בקצוות תחת מיקרוסקופ ניתוחי ככל שהכרומטין מרוכז יותר, כך הנקודה /פס11כהים יותר.
מאמר זה מתאר את השימוש בשיטת מיקרו-דיסקציה בסיוע טרנסילומינציה לבידוד מקטעי צינורית חצי-היקפיים המייצגים שלבים ספציפיים של מחזור האפיתל החצי-שנתי. לאחר בידוד, מקטעי tubule מבוימים יכולים להיות כפופים לניתוחים שונים במורד הזרם, כולל RNA ביוכימי וניתוחי חלבון12,13,14,15, cytometry זרימה16, לשעבר vivo tubule תרבות17 ו immunostaining18. כאן אנו מספקים גם פרוטוקולים מפורטים במורד הזרם כדי להכין מונוליירים מעוכים של מקטעי tubule מבוימים לניתוח מורפולוגי של תאים חיים ו immunostaining הבאים, כמו גם immunostainings הר שלם של מקטעי tubule. זרימת העבודה על קצה המזלג מתוארת באיור 4.
שיטת המיקרו-דיסקציה בסיוע transillumination מאפשרת זיהוי ובידוד מדויקים של תאי נבט בשלבים ספציפיים של בידול הודות להרכב התאי המסונכרן של השלבים. חשוב לציין, זה גם מאפשר את המחקר של אירועים תלויי במה במהלך spermatogenesis על רקמה חיה. בהתחשב בהיעדר מודלים מדרגיים במבחנה עבור spermatogenesis, שיטה זו יש גם יתרון ייחודי של מתן מחקרים התפתחותיים וטוקסיקולוגיים ממוקדים לטווח קצר על מקטעי צינורית ספציפיים לבמה ex vivo12,17. בעוד אנו מתארים את השיטה כאן עבור העכבר, אותו הליך יכול להיות מיושם על כל מין יונקים עם סידור אורך ומגזרי של שלבי אפיתל למחצה, כגון החולדה4,7,15,19,20.
Protocol
תחזוקת עכברי מעבדה וכל הניסויים בבעלי חיים נעשו בהתאם להנחיות ולתקנות הרלוונטיות לטיפול ולשימוש בחיות מעבדה באוניברסיטת טורקו.
1. הכנת צינוריות חצי-היקפיות למיקרו-דיסקציה
- להקריב עכבר זכר בוגר (≥8 שבועות, testis 80–120 מ"ג בהתאם למתח ולגיל) באמצעות CO2 חנק ואחריו נקע בצוואר הרחם.
הערה: העכבר צריך להיות בוגר מינית, ועדיף לפחות 8 שבועות. דפוס transillumination של עכברים צעירים שונה ממבוגר כי הגל של אפיתל חצי-חצי עדיין לא הוקמה במלואה, ואת העיתוי של הגל הראשון של spermatogenesis הוא ברור21,22. חוסר זרע מאריך בעכברים זכרים בני 4 שבועות מונע את השימוש בהם למיקרו-דיסקציה בסיוע טרנסילומינציה. כל זני העכבר כי יש spermatogenesis נורמלי ניתן להשתמש. - לרסס את הבטן הגחונית עם 70% אתנול. פתח את החלל abdominopelvic באמצעות מספריים סטריליים, מה שהופך פתח בצורת V.
- משיכת כרית השומן האפידילית עם מלקחיים סטריליים, לאתר את האשכים, לנתח אותם באמצעות מספריים ומניחים אותם על צלחת פטרי סטרילית 100 מ"מ המכיל PBS.
הערה: כדי לשמור על סטריליות, ודא שכל כלי המעבדה והכלים הכירורגיים סטריליים. - באמצעות מספריים עדינים לתמצת את האשכים על ידי חיתוך חריץ אלבוג'יניאה טוניקה, גיליון סיבי עבה עוטפת את האשכים. ואז לקרוע את tunica פתוח באמצעות זוג מלקחיים. כפה את הצינוריות על ידי לחיצה עם מלקחיים ולזרוק את tunica.
הערה: בעת השלכת טוניקה, זה עשוי להיות מועיל עבור כמה יישומים במורד הזרם כי האשכים arteria מוסר יחד עם טוניקה. הימנע מפגיעה בצינורות החצי-חופניים. - מעבירים את הצינוריות החצי-חצי לצלחת פטרי חדשה ויוצקים מספיק PBS סטרילי כדי לכסות את תחתית צלחת הפטרי. לאחר מכן, בעדינות למשוך את הצינוריות לגזרים אבל למנוע פגיעה tubules.
הערה: לחץ מכני רב מדי יעכב את דפוס הטרנסילומינציה וישפיע על הכדאיות של הרקמה ועל הארכיטקטורה התאית שלה. הצינוריות יכולות גם להיות מעובדות עבור immunostainings הר שלם מנקודה זו ללא היערכות (3B). לפעמים זה מספיק כדי להגדיר את השלב בדיעבד על ידי הכללת נוגדנים נגד חלבונים לידי ביטוי בידול spermatogonia, כגון SALL4, c-KIT ו DNMT3A18,23. צפיפות הזרע היא מדד שלב אמין יחסית (איור 2).
2. מיקרו-דיסקציה בסיוע טרנסילומינציה
- מניחים את צלחת פטרי תחת מיקרוסקופ ניתוח בחוזקה על ידי הקלטת אותו לבמה.
הערה: חשוב להקליט היטב את צלחת פטרי כדי למנוע את התנועה שלה אשר יכול לגרום ערבוב של קטעי tubule מבוים למחצה שנאספו. - כדי לחשוף את דפוס ספיגת האור של tubules חצי חצי בפוקוס, ודא כי המדגם מואר מלמטה ואת האור עובר דרך המדגם, כלומר, הוא transilluminated.
הערה: כמות האור שנספגת/מפוזרת היא יחסית לרמת עיבוי הכרומטין בזרעים מאריכים והצרור שלהם בתוך הצינור החצי-חצי- צינורית: ככל שהתמצית יותר, כך האור נספג יותר, כלומר, נראה כהה יותר. - הכירו את תבנית ספיגת האור של שלבים שונים כמתואר באיור 2, איור 5A ואיור S1 על ידי הזזה קפדנית של חבילות של צינוריות באמצעות מלקחיים עדינים.
הערה: השלבים תמיד עוקבים זה אחר זה בסדר הגיוני, ויוצרים את גל האפיתל החצי-שנתי. עם זאת, חשוב לדעת כי הכיוון של הגל spermatogenic מדי פעם מתהפך ולאחר מכן חוזר שוב (הידוע גם בשם אפנונים4,9 ),לפעמיםמסבך את ההליך. כמו כן, אורך כל שלב, במונחים של כמה מ"מ של tubule, משתנה במידה ניכרת. - בזהירות להרים את tubule של עניין באמצעות מלקחיים עם קצה מכור, ולאחר מכן לחתוך קטע של אורך מתאים באמצעות מספריים microdissection (ראה וידאו משלים 1). וו בקצה המלקחיים מקל על ההרמה וההחזקה של צינורית ומסייע במניעת סחיטה.
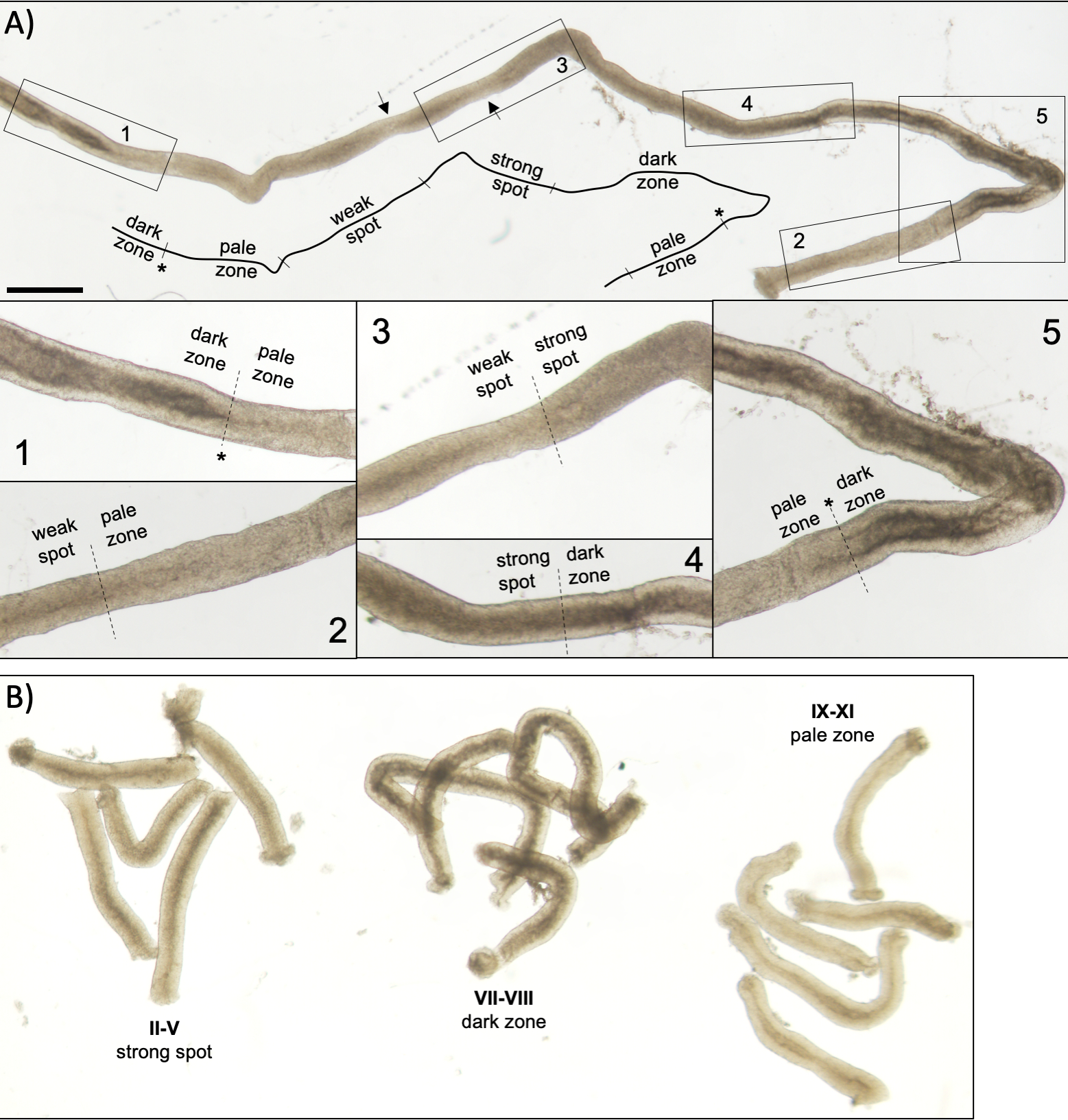
הערה: אורך המקטעים שיש לחתוך תלוי ביישומים במורד הזרם. לאיסוף חתיכות טובולה במאגר של שלב מסוים לניתוח חלבון או רנ"א12,13 (II-V, VII-VIII ו- IX-XI, איור 5B) האורך הוא בדרך כלל 2-5 מ"מ. כאשר נעשה שימוש בהפקת פנול-כלורופורם סטנדרטית, ניתן להפיק כ-200 ng של RNA מ-1 מ"מ של טובול. עבור כתמי הרכבה שלמים של מקטעי tubule מבוימים, אורך המקטעים צריך להיות >5 מ"מ. עבור ההכנות סקווש אורך המקטעים לא יעלה על 1-2 מ"מ כי התאים באמצע הקטע עלול להיכשל לצאת אם ארוך מדי. השתמש בסולם מ"מ מתחת לצלחת פטרי למדידה מדויקת של אורך הצינור.
3. חיסון של הכנות שונות
- הכנת סקווש: אימות במה וחיסון
הערה: חתיכות tubule ספציפי שלב ניתן למעוך על שקופית מיקרוסקופ עם זכוכית כיסוי לבצע ניתוח מורפולוגי של תאים חיים על ידי מיקרוסקופיה ניגוד פאזה ו immunostaining הבאים. למתחילים מומלץ להשתמש בגישה זו כדי לאמת את השלבים בעת היכרות עם שיטת microdissection בסיוע transillumination.- לאסוף את הקטע בנפח של 10 μL באמצעות פיפטה ולהעביר אותו על שקופית מיקרוסקופ.
- למעוך את הצינורית על ידי הנחת כיסוי (20 ממ x 20 ממ) בזהירות על הצינור. כתוצאה מכך, תאים יזרמו החוצה את הצינור ויוצרים מונולאי תא חי. מניחים נייר סינון על קצה זכוכית הכיסוי כדי להקל על התפשטות התאים. הימנע ממועך את התאים יותר מדי כדי לשמור אותם בחיים.
- נטר את התפשטות התאים תחת מיקרוסקופ. השתמש במיקרוסקופ עם ניגודיות פאזה במטרה 40x כדי לאמת זיהוי שלב על-ידי בדיקת סוגי התאים הקיימים (איור 2, איור S2).
- לאחר שהתאים התפשטו כדי ליצור מונולייר עגול משני קצות הצינור, טובלים את המגלשה לתוך מיכל המכיל חנקן נוזלי תוך החזקתו במלקחיים. שמור אותו שקוע במשך 10 שניות. לחלופין, הניחו את המגלשה על צלחת קרח יבשה להקפאה.
- הסר את זכוכית הכיסוי על ידי היפוך אותו באמצעות אזמל.
- ללא דיחוי, המשיכו עם הקיבעון והניחו במהירות את השקופית במיכל עם 90% אתנול למשך 2-5 דקות.
הערה: ודא הכנת סקווש לא להפשיר לפני הצבת אותו 90% אתנול. מתקנים אחרים יכולים לשמש גם, כגון אצטון, במשך 10 דקות. - יבש באוויר ומאחסן בטמפרטורת החדר (RT) (עד כמה ימים) או ב -80 מעלות צלזיוס (לטווח ארוך).
- עבור immunostaining, לאחר לתקן את הדגימות ב 4% paraformaldehyde (PFA) במשך 10 דקות ב RT.
- יש לשטוף ב-PBS ולחלחל עם 0.1% טריטון X-100 ב-PBS למשך 5 דקות.
- יש לשטוף ב-PBS ולצייר טבעת גריז סביב כל דגימת סקווש.
- הוסף 50-100 μL של 10% BSA (אלבומין סרום שור) ב 0.1% Tween ב PBS (PBST) בתוך טבעת השומן ולחסום דגימות במשך 30 דקות ב RT.
- הסר את פתרון BSA ודגר עם נוגדן ראשוני מדולל ב 10% BSA ב PBST עבור 1 שעה ב RT.
- לשטוף 3x במשך 5 דקות עם PBST.
- דגירה עם נוגדן משני מדולל ב 10% BSA ב PBST.
הערה: כדי להכתים את האקרוזומים, ניתן להדגיר את הדגימות בנוגדן אגלוטינין בוטנים (PNA, 1:1000) ב- 10% BSA ב- PBST למשך שעה אחת ב- RT (איור S3) במקום נוגדנים ראשוניים ומשניים ספציפיים. - לשטוף 3x במשך 5 דקות כל אחד עם PBST, לשטוף עם PBS ולהרכיב עם תושב המכיל DAPI.
- חיסון בהר שלם של צינוריות חצי-חופפות
הערה: הפרוטוקול שלהלן מתאר כתמי הרכבה שלמים עבור מקטעי צינורית בשלבים (משלב 2.4). אם חוקר רוצה לבצע צביעה שלמה ללא אחסון זמני (משלב 1.5), שים לב להערות ב- 3.2.1 ו- 3.2.7.- באמצעות פיפטה, להעביר את מקטעי tubule (משלב 2.4) ב PBS קר כקרח לתוך צינור חרוט 15 מ"ל ולאפשר להם משקעים על קרח.
הערה: אם משתמשים בצינורות לא מתויגים מ-1.5, מפרידים את הצינוריות בצלחת פטרי על ידי צנרת למעלה ובחזרה לצלחת מוטה מספר פעמים. השתמש פיפטה 1-mL עם קצה לחתוך. שלב זה נועד לפתוח את מבנה הרקמה. עם זאת, הימנע צינור יותר מדי כפי שהוא עלול לגרום נזק tubules. משקעים של צינוריות ייקח כמה עשרות שניות. שברי צינורית קטנים, תאים ביניים ופסולת תאים נשארים בסופרנאטנט. - הסר בזהירות את supernatant (SN) על ידי pipetting או עם שאיפה. מוסיפים 10 מ"ל של PBS קר כקרח ומערבבים על ידי היפוך.
- אפשר משקעים ולאחר מכן הסר SN כמו קודם.
- הוסף 5 מ"ל של 4% PFA ותקן במשך 5 שעות על שולחן מסתובב (20-30 סל"ד) ב +4 מעלות צלזיוס.
הערה: זמן הקיבעון תלוי בחלבונים המעניינים ובלוקליזציה התת-תאית שלהם. עבור חלבונים גרעיניים ציטופלזמיים, קיבוע 2 שעות הוא בדרך כלל מספיק, עם זאת, סמני קרום, כגון GFRα1 (אלפא קולטן משפחת GDNF 1; איור 6א',ב),תיהנה מקיבעון ארוך יותר, עד 6 שעות. - אפשר משקעים, להסיר SN (PFA) כמו קודם, ולשטוף בקצרה על ידי הוספת 10 מ"ל של PBS והפך את הצינור.
- אפשר משקעים, להסיר SN כמו קודם, ולחזור על שלב שטיפת PBS שלוש פעמים לפחות 10 דקות כל אחד על שולחן מסתובב ב +4 מעלות צלזיוס ולהמשיך עם כתמים או לאחסן ב +4 מעלות צלזיוס.
הערה: אם תנאי העבודה סטריליים ולנקות את הדגימות ניתן לאחסן ולהשתמש בהם לפחות כמה שבועות. לחלופין, הוסיפו נתרן אזיד לריכוז סופי של 0.02% (w/v) מתמיסת 2% למניה כדי לסייע בשימור הצינוריות לפני האחסון ב-4 מעלות צלזיוס. - באמצעות פיפטה של 1 מ"ל, העבר מקטעי צינורית קבועים של 10-20 לצינור עגול של 2 מ"ל. אפשר משקעים ולהסיר SN.
הערה: אם עובדים עם צינוריות ארוכות שלא בוימו, יוצקים את הצינוריות על צלחת פטרי ומשתמשים במספריים זעירים ומלקחיים חותכים מקטעים של כ-5-20 מ"מ. מקטעים ארוכים מדי יסתבכו במהלך הליך הכתם, ואילו מקטעים קצרים מדי יאבדו בקלות. - הוסף 1 מ"ל של 2% BSA + 10% FBS (סרום שור עוברי) ב 0.3% טריטון X-100 ב PBS (PBSX). חסום למשך שעה אחת לפחות בטבלה מסתובבת (20-30 סל"ד) ב- RT.
- לשטוף עם 1 מ"ל של PBSX, להסיר SN על ידי pipetting ולהוסיף 250 μL של נוגדן ראשוני מדולל 1% BSA ב PBSX (1:100-1:2000 דילול). דגירה במשך 2 שעות ב RT או לילה ב +4 מעלות צלזיוס על שולחן מסתובב (20-30 סל"ד).
- הסר את פתרון הנוגדנים על ידי pipetting ולשטוף את tubules עם 1 מ"ל של PBSX כאמור לעיל. יש לשטוף שלוש פעמים ב-PBSX על שולחן מסתובב (20-30 סל"ד) ב-RT.
הערה: לאחר שטיפה ראשונה זו ניתן להשאיר את הדגימה למשך הלילה ב-4 מעלות צלזיוס במידת הצורך. - הסר SN ולהוסיף 250 μL של נוגדן משני מדולל 1% BSA ב PBSX (בדרך כלל 1:500 דילול של נוגדן תווית פלואורסצנטית). מכסים בנייר כסף ומדגירה על שולחן מסתובב (20-30 סל"ד) ב-RT למשך שעה.
- חזור על 3.2.10.
- לבסוף להסיר SN ויוצקים את tubules למגלשת מיקרוסקופ. יש להפריד בעדינות ולסדר צינוריות ברצועות ליניאריות בעזרת טיפים להעמסת ג'ל. מסננים את המאגר העודף ומוסיפים מדיום הרכבה וכיסוי.
הערה: הימנע ייבוש של tubules בעת סידור אותם. מכתים נגד של הגרעינים עם DAPI אינו הכרחי ברוב המקרים. לאטום את קצה הכיסוי עם לק כדי למנוע ייבוש מדגם והידרדרות. ניתן לאחסן שקופיות למשך שבוע עד שבועיים ב- +4 °C (60 °F) לפני ההדמיה.
- באמצעות פיפטה, להעביר את מקטעי tubule (משלב 2.4) ב PBS קר כקרח לתוך צינור חרוט 15 מ"ל ולאפשר להם משקעים על קרח.
תוצאות
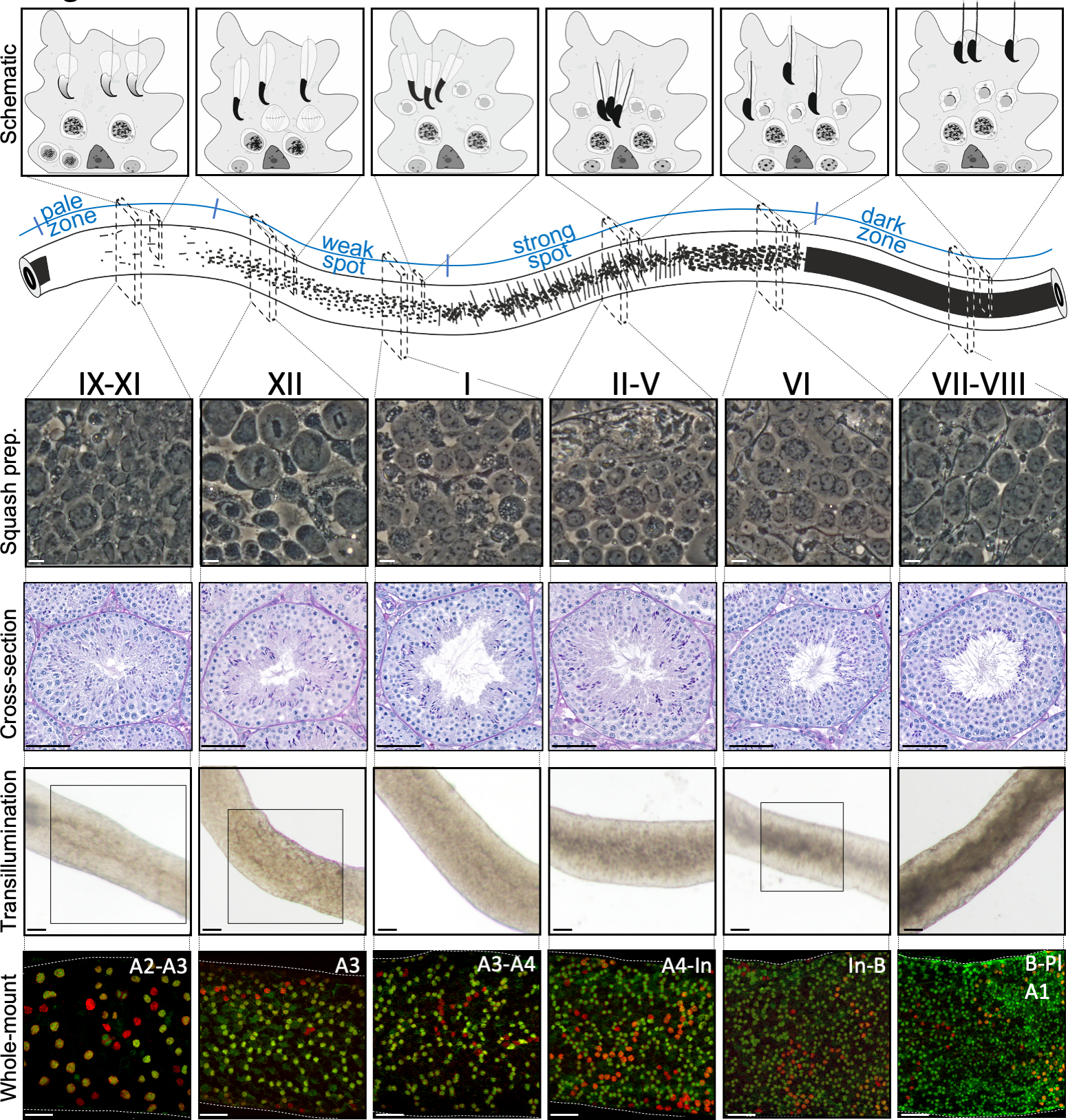
בימוי מוצלח בסיוע טרנסילומינציה ומיקרו-דיסקציה של צינוריות עכבר חצי-היקפיות תלויים בעיקר במציאת תנאי התאורה המתאימים ובמיקרוסקופ דיסקציה מתאים, וביכולת לזהות תכונות ספציפיות המאפיינות כל שלב. שלבים VII-VIII נראים כהים באופן הומוגני משום שהם מכילים מספר גבוה של זרעונים מוארכים מרוכזים במלואם המיושרים על פני השטח האפיתל של האפיתל (איור 5A ואיור S1). לאחר זרע בוגר משתחררים לתוך לומן זרע, tubule נראה חיוור מאוד בשלבים IX-XI בשל היעדר זרע מוארך מרוכז באפיתל. התכונה הקלה ביותר לזיהוי על צינוריות שעברו טרנסילומינציה היא נקודת הזרע (כוכבית באיור 5A ובאיור S1),כלומר המעבר הפתאומי מהאזור החשוך (VII-VIII) לאזור החיוור (IX-XI). האזור החיוור ואחריו אזור נקודת התורפה (XII-I). המראה המנוקד מקורו בארגון של זרע מאריך עם כרומטין מרוכז בחבילות. החבילות הופכות בולטות מאוד באזור הנקודה החזקה הבא (II-V). יתר על כן, חבילות זרע נודדות לכיוון גרעיני התאים של סרטולי הממוקמים קרוב לבזל למינה, אשר משתקף כמראה פסים של צינורית שלב II-V בעת טרנסילומינציה (איור 2, איור 5A, B ואיור S1). חבילות סוף סוף להתפזר בשלב VI ועיבוי spermatids מוארך לנוע קרוב לומן להשתחרר מן האפיתל בשלב השמיני.
ניתן לאמת במדויק את השלב המדויק של קטע הצינור באמצעות מיקרוסקופיית ניגודיות פאזה של הכנות סקווש (איור 2 ואיור S2). השלבים הספציפיים בהכנות סקווש מוכרים על בסיס ההתפתחות האקרוזומלית של זרעים עגולים בשלב 1-8, מצב עיבוי הכרומטין בזרעים מאריכים ונוכחות של חבילות זרע16 (איור 3 ואיור S2). יתר על כן, נוכחות של סוגי תאים קודמים, כגון זרע מסוג B ו leptotene או זרע zygotene, כי ניתן לזהות על בסיס התכונות המורפולוגיות שלהם, ניתן להשתמש כדי לתמוך בזיהוי הבמה. הגודל של גרעיני זרע pachytene מגדיל את גודל סביב שלב VI, אשר עשוי גם לספק עזרה נוספת בבימוי.
ההכנות סקווש מבוים ניתן להשתמש כדי ללמוד את הביטוי ואת לוקליזציה של חלבונים של עניין אפיתל חצי חצי באמצעות immunostaining. זה מאפשר ניתוח מדויק מאוד של ביטוי ספציפי לסוג התא בגלל הרכב הסלולר מוגדר היטב בכל שלב. ביטוי ספציפי לשלב יכול להיות מוגבר עוד יותר על ידי כתמים משותפים של האקרוזום (למשל, עם PNA) המאפשר הדמיה של צעדים נפרדים של בידול זרע עגול. באיור S3מוצגות תמונות מייצגות של אקרוזומים מוכתמים ב-PNA בשלבים שונים . הכתם האקרוזומלי יכול לשמש גם כדי להגדיר את הבמה בדיעבד. עם זאת, ההיערכות בסיוע טרנסילומינציה היא שיטה קלה ומהירה בהרבה למצוא את השלב הרצוי מאשר להשתמש בשברים לא מעוקלים באופן אקראי.
צביעת צינורית חצי-חצי-הררית משמשת בדרך כלל לחקר סוגי התאים הנמצאים במגע עם קרום המרתף של האפיתל החצי-צינורי, או בצד הצינורי (זרעונים, תאי זרע פרפטוטן ותאי סרטולי; איור 6A,B) או הצד הבין-כוכבי (תאי מיואידים פריטובולריים ומאקרופאגים פריטובולריים; איור 6C ואיור S4A)24. עם זאת, ניתן להשתמש בשיטה גם כדי לחקור תאים או מבנים הממוקמים עמוק יותר באפיתל, כגון מחסום בדיקת הדם (Espin, Figure S4B), או תאי נבט פוסט-מיוטיים (סמן אקרוזום PNA, איור S4C). אם משתמשים במקטעי צינורית לא מתויגים, ניתן להעריך שלב נתון בדיעבד באמצעות נוגדן נגד חלבון המתבטא בהבחנה בין זרע (A1, A2, A3, A4, In ו- B; ביחדDiff)(איור 2). הבימוי אז מסתמך על צפיפות שלdiff סינקטיאלי שעבר שש חטיבות מיטוטיות באופן תלוי שלב במהלך המחזור הראשון של בידול spermatogenic1, ולכן הכפלת מספר זרעדיף בסינטיה לאחר כלחטיבה 1,25. עם זאת, היערכות רטרוספקטיבית היא פחות מדויקת מאשר בימוי בסיוע transillumination כי אין סמנים ספציפיים עבור דורות שונים שלהבדל והערכה של צפיפותDiff עשוי להיות נוטה לטעות.

איור 1: מפת מחזור אפיתל חצי-חצי לבימוי של זרעון עכבר. עמודות אנכיות מציגות שיוכי תאים בשלבים שונים של מחזור האפיתל החצי-שנתי (מסומן בספירות רומיות I-XII). תאי הנבט הלא בוגרים ביותר נמצאים בתחתית, ואילו המובחנים ביותר נמצאים בחלק העליון. כדי לעקוב אחר ההתקדמות של בידול תא נבט, יש לעבור משמאל לימין, ומהתחתית למעלה. מחזור של האפיתל החצי-שנתי הוא סדרה שלמה של שלבים העוקבים זה אחר זה בסדר מספרי. זרעוןלא מובחן; A1-4, סוג A1–A4 זרע; ב, זרע ביניים; B, סוג B זרע; Pl, זרעון פרפטוטן; L, זרע לפטוטן; Z, זרעונים זיגוטנים; P, זרע פצ'יטן; D, זרע דיפלוטן; 2°, זרעונים משניים בתוספת חטיבות מיוטיות. ספרות ערביות 1-16 מתייחסות לשלבים של התבגרות זרע פוסט-מיוטית (זרע). לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: שיוכי תאים בשלבים של מחזור האפיתל החצי-שנתי של העכבר. שלבי מחזור האפיתל החצי-שנתי עוקבים זה אחר זה בסדר לוגי, ובכך יוצרים את הגל הזרעוני לאורך הציר האורך של צינורית שפירה למחצה. הלוח העליון מדגים את אסוציאציות תאי הנבט בשלבים שונים, ואת המיקום של סוגי תאים בתוך הכיסים הציטופלזמיים של תאי סרטולי (אפור בהיר) באפיתל החצי-חצי. האיור של הצינורית החצי-חצי-שקופית מדמיין את הגל הזרעוני ואת דפוסי הטרנסילומינציה הספציפיים של נקודת התורפה החיוורת, החלשה, הנקודה החזקה והאזורים הכהים. מכל שלב שצוין, מוצגות תמונות ייצוגיות של ניגודיות פאזה של תאים חיים של תכשירי סקווש וחומצה-שיף תקופתית (PAS) מוכתמת בחצאי אשכים. שני הלוחות התחתונים מציגים קטעים של צינורית חצי-חצי(למעלה) או כתמים עם נוגדנים (למטה) נגד SALL4 (אדום, סמן פאן-זרע) ו- DNMT3A (ירוק, סמןDiff). דפוסי transillumination לשלבים IX-XI, XII ו- VI מודגשים עם אזורים ארוזים. הן דפוס ספיגת אור (למעלה) וצפיפות של DNMT3A חיובי בידול spermatogonia (התחתון) ניתן להשתמש כדי להגדיר את השלב (משוער) של המחזור החצי-שנתי. דורות רצופים שלDiff מכונים סוג A1-A4, ביניים (In) וסוג B זרע. חלוקה של כל אחת מהן גורמת להכפלת צפיפות תאי הזרע. צפיפות התאים הגבוהה ביותר על קרום המרתף של האפיתל החצי-חצי נצפית בשלבים VI-VIII כאשר נצפו זרעונים מיוטיים פרפטוטן (Pl). סרגלי סולם: הכנת סקווש. 10 מיקרומטר, אחרים 50μm. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3: שלבים של זרעון. תכונות ייחודיות של spermatids בשלבים שונים של spermiogenesis. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 4: זרימת עבודה של מחקר זה. תבנית Transillumination הוקמה במלואה בעכבר מבוגר (>בן 8 שבועות). להקריב את העכבר ולהמשיך עם ניתוח testis ועקף ללא דיחוי. בעדינות למשוך את הצינוריות לגזרים על צלחת פטרי. תקן tubules עבור כתמים הר שלם או להמשיך עם transillumination. תחת transillumination 1) לחתוך מקטעי tubule קצר של שלבים ספציפיים(ים) עבור הכנות סקווש (בקרת איכות או immunostaining) או 2) לחתוך קטעים ארוכים יותר כדי לאסוף בריכות במה עבור ניתוחים RNA ופרוטאומיקה, תרבות רקמות או כתמים הר שלם. העבר מקטעי tubule קצרים לשקופית מיקרוסקופ בנפח 10 μL של PBS. מקם תגית כיסוי על המקטע כדי לאלץ את התאים וליצור קו monolayer תא חי. שים לב לסוגי התאים תחת מיקרוסקופ, ואז לתקן ולהכתים. WMS, כתמי הרכבה שלמים. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 5: צינורית חצי-חצי של עכבר בוגר כפי שניתן לראות תחת מיקרוסקופ טרנסילומינציה. (A)קטע ארוך של צינורית חצי-חצי-חצי כפי שניתן לראות תחת טרנסילומינציה המציגה את גל האפיתל החצי-שנתי. ניתן לזהות ארבעה אזורים נפרדים בהתבסס על דחיסת כרומטין בזרעים והלוקליזציה שלהם בתוך האפיתל: אזור חשוך (שלבים VII-VIII), אזור חיוור (שלבים IX-XI), אזור נקודת תורפה (שלבים XII-I) ואזור נקודה חזקה (שלבים II-VI). כוכבית, נקודת זרע. החצים מצביעים על שני מקטעים קצרים בתוך אזור נקודת התורפה בעלי מראה חיוור עקב לחץ מכני על הצינור. סרגל קנה מידה: 500 מיקרומטר. ערכות 1-5 הן הגדלות גבוהות יותר ממקטעי צינוריות שנבחרו. (ב) מקטעים במאגר של צינורית חצי-חצי-חצי המייצגת את שלבים II-V (אזור נקודה חזקה), VII-VIII (אזור כהה) ו- IX-XI (אזור חיוור). לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 6: מערכת חיסונית שלמה באמצעות תאי זרע, תאי סרטולי וסמני מקרופאגים פריטובולריים. (A)מכתים הרכבה שלמה עבור קטע המייצג את שלב XI-II עם נוגדנים נגד GFRα1 (אדום), SALL4 (ירוק) ו DNMT3B (כחול). הכתם חושף שלוש אוכלוסיות שונות של זרע: מבודדים(A)או מזווגים(יח"צ)זרע גזע מובחן (GFRα1+/ SALL4+/ DNMT3B- ;אזורים מנוקדים לבנים), סינכרוניאלי קצר (כאן 4 תאים מיושרים; A al4) זרע אב (GFRα1-/ SALL4+ /DNMT3B-; אזורים מנוקדים צהובים) ומבדיל זרעונים (GFRα1-/ SALL4+ /DNMT3B+). SALL4+/ / DNMT3B+ תאים הם סוג A3–A4 זרע. (B)כתמי צינור חצי-חצי הרריים עם נוגדנים נגד GFRα1 (אדום), USF1 (ירוק) ו- SOX9 (כחול). GFRα1 מכתים את תת-קבוצת הגזע של זרעונים. USF1 מתבטא הן תאי זרע והן SOX9+ סרטולי. (C)מקרופאגים פריטובולריים בעכברים בוגרים הם חיוביים הן עבור F4/80 (אדום) והן עבור MHCII (כחול). DAPI מכתים DNA (ירוק). גרעינים גדולים בהירים הם גרעיני תאים מיואידיים פריטובולריים. סרגלי קנה מידה: 50 מיקרומטר. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}
איור S1: קטע ארוך של צינורית חצי-חצי-חצי כפי שניתן לראות תחת טרנסילומינציה המציגה את גל האפיתל החצי-שנתי. ניתן לזהות ארבעה אזורים נפרדים בהתבסס על דחיסת כרומטין בזרעים והלוקליזציה שלהם בתוך האפיתל: אזור נקודת תורפה (שלבים XII-I), אזור נקודה חזק (שלבים II-VI), אזור חשוך (שלבים VII-VIII) ואזור חיוור (שלבים IX-XI). ניתן לזהות את נקודת הזרע (כוכבית) כאזור החשוך העובר בפתאומיות לאזור החיוור. סרגל קנה מידה: 500 מיקרומטר. ערכות כניסה 1-4 הן הגדלות גבוהות יותר ממקטעי tubule שנבחרו. אנא לחץ כאן כדי להוריד נתון זה.
{kind=link}
איור S2: מיקרוסקופיית ניגודיות פאזה של מונוליירים של תאים חיים בשלבים שונים של מחזור האפיתל החצי-שנתי. (A)שלב I. שלב 1 זרע עגול עדיין חסר את המבנה האקרוזומלי. הם מאופיינים בגרעין עגול קטן (עיגולים אדומים) עם כרומוצנטרי יחיד ייחודי (חצים לבנים). הגוף הכרומטואידי נראה בציטופלסמה כגרגר כהה במגע הדוק עם הקרום הגרעיני (חצים כחולים). גרעין התא סרטולי (עיגול לבן) מכיל שלושה מוקדים כהים: גרעין גדול עם שני כרומוזנטים לווין. תאי סרטולי אינם נראים בהכנות סקווש אבל הם זורמים מדי פעם מן הצינור יחד עם תאי נבט. (B)שלב II-IV. בשלב 4 זרעונים עגולים (עיגולים אדומים), המבנה האקרוזומלי (חצים אדומים) מופיע כסקל לבן שטוח מעט המחובר למעטפה הגרעינית. זרעים מוארכים יוצרים חבילות וכבר יש להם הלקאה עבה (חצים לבנים) המציינת את נוכחותו של נדן מיטוכונדריאלי בחתיכה האמצעית של זנב הזרע. הגרעינים של זרעון pachytene (PSpc) הם בערך כפול גדולים כמו אלה של זרע עגול מאופיינים באזורי כרומטין כהים מופץ ברחבי הגרעין. (C)שלב V-VI. בשלב 5-6 זרעונים עגולים (עיגול אדום), האקרוזום שטוח עוד יותר ומתפשט על הגרעין (חצים אדומים). אזור המעטפה הגרעינית הפונה לאקרוזום נראה חשוך בשל נוכחות האקרופלקסום, צלחת עשירה בחלבון בין הקרום האקרוזומלי לקרום הגרעיני. זרעים מאריכים משתחררים מחבילות (חצים לבנים). זרעון Pachytene (PSpc) נצפו לעתים קרובות באפיתל. (ד)שלב VII-השמיני. האקרוזום (חצים אדומים), בטנה כהה על המעטפה הגרעינית עם צללים לבנים, מורחב במלואו ומכסה כמעט את כל הצד האפור של שלב 7-8 זרעונים עגולים (עיגולים אדומים). שלב זה מאופיין בזרעים בוגרים (חצים לבנים) שעשויים להיות בשפע בחלקים רבים של הכנת הדלעת. הגרעין של זרעון פצ'יטן (PSpc) גדל בגודל במהלך הפיתוח ומופיע גדול יותר בשלב השביעי השמיני מאשר בשלבים קודמים. אזורי הכרומטין הכהים בתוך הגרעין של זרעון פצ'יטן (PSpc) נראים מטושטשים עקב פעילות שעתוק גבוהה ואירועים מיוטיים. (E)שלב X. שלב 10 גרעיני זרע (חצים לבנים) יזמו התארכות אבל הכרומטין עדיין לא התרכז. האקרוזומים מתחילים ליצור וו בקצה הגרעיני (חץ אדום). הגרעינים של זרע pachytene (PSpc) מופיעים גדולים מאוד כפי שהם מתכוננים חטיבות מיוטי. (ו)שלב 12. שלב XII מאופיין בנוכחות לוחות מטפאזה מיוטיים (עיגולים מקווקווים לבנים). תאים עגולים קטנים עם דפוס כרומטין מרוכז טיפוסי הם זרעונים זיגוטנים (ZSpc), שבו כרומוזומי האחות מתיישרים כדי ליזום היווצרות מורכבת סינפטון. סרגלי קנה מידה: 10 מיקרומטר. אנא לחץ כאן כדי להוריד נתון זה.
{kind=link}
איור S3: זיהוי השלב של מחזור אפיתל חצי-שנתי על בסיס כתמים אקרוסומליים וגרעיניים. ההכנות סקווש קבוע אצטון היו מוכתמים עם PNA רודמין מצומד ו DAPI. שלב I: אמנם אין אקרוזום קיים בשלב 1 זרע עגול acrosome מפותח יכול להיות מזוהה זרע מאריך. שלבים II-IV: התפתחות אקרוסומלית מתחילה עם הופעתם של גרגירים פרואקרוסומליים/אקרוזומליים בזרעים עגולים. Vesicle אקרוזומלי על פני השטח הגרעיניים מופיע עגול עד שלב 3 spermatids, ולאחר מכן משטח בשלב 4 spermatids. שלב V: הזווית הממוזגת על ידי האקרוזום משתרעת מ-40 מעלות למקסימום של 95°. שלבים VI-VII: הזווית של האקרוזום משתרעת מ-95 מעלות ל-120 מעלות. שלבים VIII-IX: האקרוזום הוארך במלואו בשלב 8 spermatids (שלב VIII), וגרעינים מקוטבים בצד apical של התא יצירת קשר עם קרום הפלזמה (לא מוצג). בשלב התשיעי גרעין הזרע הופך מעוות; משטחים הגביים והיפנים נראים לראשונה. שלבים X-XI: זרע להראות את הזווית הגבית. שלב 12: שלב זה מאופיין בצורה הטובה ביותר על ידי הופעת חטיבות מיוטיות; לוחות metaphase מסומנים עם חצים לבנים. זרעים מאריכים עם האקרוזומים שלהם נראים גם. סרגלי קנה מידה: 10μm. אנא לחץ כאן כדי להוריד נתון זה.
{kind=link}
איור S4: כתמי הרכבה שלמים לסמנים לא קונבנציונליים. (A) אקטין שרירים חלק אלפא (aSMA) מתבטא על ידי תאים מיואידיים peritubular. (ב) אספין תמקם בצמתים הדוקים של תאי סרטולי ותורם למחסום בדיקת הדם. (ג) PNA תמקם לאקרוזומים של זרעונים. סרגלי קנה מידה: 50μm. אנא לחץ כאן כדי להוריד נתון זה.
{kind=link}
וידאו משלים 1: חיתוך קטע קצר של tubule חצי-צינורי המייצג את שלב VII-VIII. אנא לחץ כאן כדי להוריד וידאו זה.
Discussion
שיטת microdissection בסיוע transillumination שתיארנו לעיל מאפשרת גישה מוכוונת שלב לחקר זרע. Spermatogenesis הוא תהליך מסונכרן מאוד, וכל הצעדים העיקריים במהלך הבידול spermatogenic מוסדרים ומבוצעים באופן תלוי שלב, כגון מחויבות בידול (בשלבים VII-VIII), הופעת מיוזיס (VII-VIII), חטיבות מיוטיות (XII), הופעת התארכות זרע (VIII) ו זרע (VIII)1,26,27. הניתוח מונחה הבמה מספק כלי רב עוצמה ללמוד אירועים מסוימים אלה המוגבלים לשלבים ספציפיים של spermatogenesis ולכן נמצא רק בשלבים מוגדרים של מחזור האפיתל חצי. מאסטרינג השיטה לוקח קצת תרגול ושימוש במיקרוסקופ ניתוח באיכות טובה ותנאי תאורה נאותים הם המפתח להצלחה. יישום שיטה זו כחלק ערכת הכלים היומיומית יש יכולת לשפר באופן משמעותי את ההשפעה ואת הרלוונטיות הביולוגית של מחקר על תפקודי הרבייה הגברית על ידי מתן ניתוח מדויק יותר של אירועים מולקולריים במהלך spermatogenesis.
כל זני העכבר WT שלמדנו מציגים תבנית טרנסילומינציה דומה ומציגים אסוציאציות תאים שמורות בשלבים של מחזור האפיתל החצי-שנתי. בתנאי כי בידול spermiogenic של תאי נבט אינו שונה באופן גס מעכברי WT, אותו הדבר חל גם על כל המודלים בעכבר נוקאאוט כי למדנו. יתר על כן, זה יכול להיות מיושם על מינים אחרים המציגים סידור סגמנטי אורך של שלבים של מחזור אפיתל למחצה7. עם זאת, לא ניתן להשתמש במינים עם שלבים לא סגמנטיים (כגון בני אדם). בהתחשב בתפקיד המהותי של עיבוי כרומטין בהארכת זרע בהגדרת דפוס הטרנסילומינציה, ברור כי כל רגולציה שגויה של תהליך זה בהכרח תעכב את יישום שיטה זו. בעכברים צעירים ומבוגרים צעירים (5-6 שבועות) דפוס transillumination עדיין לא הוקמה במלואה, ולכן, רק עכברים מעל 8 שבועות יש להשתמש. חשוב גם לזכור כי סחיטה ומשיכת הצינוריות בהכרח לפגוע בדפוס transillumination כי זה מעוות את הארכיטקטורה התאית בתוך האפיתל חצי.
מקטעי השחפת המבודדים למחצה יכולים גם להיות תרבותיים המאפשרים התבוננות אקס ויוו ומניפולציה של תהליכים מצמד זרע, כולל מיוזיס. כדי להבטיח את הכדאיות של הרקמה ולמנוע השפלה RNA וחלבון, יש לאסוף את הדגימות ולעבד אותן לא יותר משעתיים לאחר הקרבת העכבר. עבור תרבות ex vivo של tubules חצי, הזמן מהקרבה לתחילת התרבות לא יעלה על 1 שעה. השלמות של שברי tubule בדרך כלל ניתן לשמור עד 72 שעות במבחנה אם נקצר כראוי.
השלב של מחזור האפיתל החצי-חצי יכול להיות מאומת ואף מוגדר בצורה מדויקת יותר באמצעות מיקרוסקופיית פאזה-ניגודיות של הכנות סקווש16. מיקרוסקופיה מבוצעת על תאים חיים, אשר מספק מימד נוסף לתוך הניתוח ומאפשר תצפית של organelle או תנועות תא בשלבים ספציפיים של spermatogenesis28,29,30. מיקרוסקופיית ניגודיות פאזה מספקת היערכות מדויקת לכשל חיסוני עוקב, המאפשר ניתוח מפורט מאוד של ביטוי החלבון ודינמיקת לוקליזציה במהלך spermatogenesis, כולל שינויים ספציפיים לשלב.
בעוד תאים משתחררים מההקשר האפיתל בהכנות סקווש, immunostainings הר שלם של מקטעי tubule לאפשר את המחקר של תאים spermatogenic בסביבה הפיזיולוגית שלהם. לכן, ההכנות להרכבה שלמה עשויות לספק הדמיה טובה יותר של ארכיטקטורת הצינור החצי-צינורית והמגעים הבין-תאיים שלה מאשר חיסון על חתכים. חשוב לציין, היערכות בסיוע transillumination של מקטעי tubule לפני immunostaining עושה את הגישה אפילו יותר חזק על ידי הכללת מידע על השלב הספציפי של קטע נתון. הכתמת הרכבה שלמה היא כלי שימושי במיוחד לחקר תאים בפריפריה של צינוריות חצי-צינוריות, כגון תאי מיואידים פריטובולריים, מקרופאגים פריטובולריים וזרעונים, אך עשויה גם לפתוח תובנות חדשניות על מחקר על תאי נבט מיוטיים ופוסט-מיוטיים.
Disclosures
למחברים אין מה לחשוף.
Acknowledgements
עבודה זו נתמכה על ידי מענקים מהאקדמיה של פינלנד [315948, 314387 ל- N.K.]; קרן סיגריד ג'וסליוס [ל-נ.ק., ג'יי.טי]; קרן אמיל אלטונן [לג'יי-איי.M,ט.ל.]; תוכנית הדוקטורט של טורקו לרפואה מולקולרית [S.C.-M., O.O.].
Materials
| Name | Company | Catalog Number | Comments |
| bovine serum albumin (BSA) | Sigma | A9647 | |
| cover glass 20x20 mm | Menzel Gläser | 11961988 | |
| Falcon conical tube 15-ml | Sarstedt | 62.554.502 | |
| fetal bovine serum (FBS) | Biowest | S1810 | |
| grease pen (ImmEdge) | Vector Laboratories | H-4000 | |
| microscope slide Superfrost Plus | Thermo Scientific | 22-037-246 | |
| Parafolmaldehyde (PFA) | Electron Microscopy Sciences | 15714 | |
| Petri dish (100-mm) | Greiner | 664160 | |
| phosphate-buffered saline (PBS) | Gibco | 11503387 | |
| ProLong Diamond Antifade Mountant | Thermo Fisher | P36962 | |
| rhodamine-labelled Peanut agglutinin (PNA) | Vector Laboratories | RL-1072 | |
| Triton X-100 | Sigma | 93443 | |
| Tween-20 | Sigma | P2287 |
References
- Mäkelä, J. A., Toppari, J. Testis Physiology: Seminiferous Cycle. Encyclopedia of Reproduction. , (2018).
- Oakberg, E. F. Duration of spermatogenesis in the mouse and timing of stages of the cycle of the seminiferous epithelium. The American Journal of Anatomy. 99 (3), 507-516 (1956).
- Clermont, Y. Kinetics of spermatogenesis in mammals: seminiferous epithelium cycle and spermatogonial renewal. Physiological Reviews. 52 (1), 198-236 (1972).
- Perey, B., Clermont, Y., Leblond, C. The wave of the seminiferous epithelium in the rat. American Journal of Anatomy. 108 (1), 47-77 (1961).
- de Lima e Martins Lara, N., Costa, G., Avelar, G., Lacerda, S., Hess, R., França, L. Testis Physiology-Overview and Histology. Encyclopedia of Reproduction. , (2018).
- Leblond, C. P., Clermont, Y. Definition of the stages of the cycle of the seminiferous epithelium in the rat. Annals of the New York Academy of Sciences. 55 (4), 548-573 (1952).
- Parvinen, M. Regulation of the seminiferous epithelium. Endocrine Reviews. 3 (4), 404-417 (1982).
- Kotaja, N., et al. Preparation, isolation and characterization of stage-specific spermatogenic cells for cellular and molecular analysis. Nature Methods. 1 (3), 249-254 (2004).
- Nakata, H., Sonomura, T., Iseki, S. Three-dimensional analysis of seminiferous tubules and spermatogenic waves in mice. Reproduction. 154 (5), 569-579 (2017).
- Russell, L. D., Ettlin, R. A., SinhaHikim, A. P., Clegg, E. D. . Histological and histopathological evaluation of the testis. , (1990).
- Parvinen, M., Hecht, N. B. Identification of living spermatogenic cells of the mouse by transillumination-phase contrast microscopic technique for "in situ" analyses of DNA polymerase activities. Histochemistry. 71 (4), 567-579 (1981).
- Ventelä, S., Mäkelä, J. A., Kulmala, J., Westermarck, J., Toppari, J. Identification and regulation of a stage-specific stem cell niche enriched by Nanog-positive spermatogonial stem cells in the mouse testis. Stem Cells. 30 (5), 1008-1020 (2012).
- Faisal, I., et al. Transcription factor USF1 is required for maintenance of germline stem cells in male mice. Endocrinology. 160 (5), 1119-1136 (2019).
- Wright, W. W., et al. Identification of stage-specific proteins synthesized by rat seminiferous tubules. Biology of Reproduction. 29 (1), 257-270 (1983).
- Johnston, D. S., et al. Stage-specific gene expression is a fundamental characteristic of rat spermatogenic cells and Sertoli cells. Proceedings of the National Academy of Sciences of the United States of America. 105 (24), 8315-8320 (2008).
- Toppari, J., Bishop, P. C., Parker, J. W., diZerega, G. S. DNA flow cytometric analysis of mouse seminiferous epithelium. Cytometry. 9 (5), 456-462 (1988).
- Mäkelä, J. A., et al. Hedgehog signalling promotes germ cell survival in the rat testis. Reproduction. 142 (5), 711-721 (2011).
- La, H. M., et al. Identification of dynamic undifferentiated cell states within the male germline. Nature Communications. 9 (1), 04827 (2018).
- Toppari, J., Parvinen, M. In vitro differentiation of rat seminiferous tubular segments from defined stages of the epithelial cycle morphologic and immunolocalization analysis. Journal of Andrology. 6 (6), 334-343 (1985).
- Parvinen, M., Vanha-Perttula, T. Identification and enzyme quantitation of the stages of the seminiferous epithelial wave in the rat. The Anatomical Record. 174 (4), 435-449 (1972).
- Kluin, P. M., Kramer, M. F., de Rooij, D. G. Spermatogenesis in the immature mouse proceeds faster than in the adult. International Journal of Andrology. 5 (3), 282-294 (1982).
- Yoshida, S., et al. The first round of mouse spermatogenesis is a distinctive program that lacks the self-renewing spermatogonia stage. Development. 133 (8), 1495-1505 (2006).
- Chan, A. L., et al. Germline stem cell activity is sustained by SALL4-dependent silencing of distinct tumor suppressor genes. Stem Cell Reports. 9 (3), 956-971 (2017).
- Lokka, E., et al. Generation, localization and functions of macrophages during the development of testis. Nat Commun. 11 (1), 4375 (2020).
- Mäkelä, J. A., Toppari, J. Testis physiology: Spermatogenic cell syncytium. Encyclopedia of Reproduction. , (2018).
- Griswold, M. D. Spermatogenesis: The commitment to meiosis. Physiological Reviews. 96 (1), 1-17 (2016).
- Mäkelä, J. A., Toppari, J. Spermatogenesis. Endocrinology of the Testis and Male Reproduction. , 1-39 (2017).
- Ventela, S., Toppari, J., Parvinen, M. Intercellular organelle traffic through cytoplasmic bridges in early spermatids of the rat: mechanisms of haploid gene product sharing. Molecular Biology of the Cell. 14 (7), 2768-2780 (2003).
- Parvinen, M., Parvinen, L. M. Active movements of the chromatoid body. A possible transport mechanism for haploid gene products. The Journal of Cell Biology. 80 (3), 621-628 (1979).
- Parvinen, M., Söderström, K. O. Chromosome rotation and formation of synapsis. Nature. 260 (5551), 534-535 (1976).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved