
Method Article
Organoidi epiteliali mucociliari di cellule embrionali xenopus: generazione, coltura e imaging dal vivo ad alta risoluzione
In questo articolo
Riepilogo
Descriviamo un semplice protocollo per sviluppare organoidi epiteliali mucociliari da cellule ectodermi profonde isolate dagli embrioni xenopus laevis. I progenitori multipotenti rigenerano i precursori delle cellule del calice epiteliale e permettono il tracciamento vivo dell'iniziazione e della progressione delle transizioni cellulari sulla superficie degli organoidi.
Abstract
L'epitelio mucociliare fornisce la prima linea di difesa rimuovendo particelle estranee attraverso l'azione della produzione di muco e dello sgombero mediato dalla ciglia. Molti difetti clinicamente rilevanti nell'epitelio mucociliario sono dedotti mentre si verificano in profondità all'interno del corpo. Qui, introduciamo un modello 3D trattabile per l'epitelio mucociliario generato da progenitori multipotenti che erano microchirurgicamente isolati dagli embrioni di Xenopus laevis. Gli organoidi epiteliali mucociliari sono ricoperti da epitelio appena generato da cellule ectodermi profonde e successivamente decorati con distinte cellule multiciliate a motivi geometrici, cellule secretorie e cellule di calice che producono muco che sono indistinguibili dall'epidermide nativa entro 24 ore. Le sequenze complete di transizioni cellulari dinamiche dal mesenchimale all'epiteliale che emergono sulla superficie apicale degli organoidi possono essere tracciate da immagini dal vivo ad alta risoluzione. Questi organoidi epiteliali mucociliari in vitro coltivati e auto-organizzanti offrono notevoli vantaggi nello studio della biologia dell'epitelio mucociliario ad alta efficienza nella generazione, condizioni di coltura definite, controllo del numero e delle dimensioni e accesso diretto per l'imaging vivo durante la rigenerazione dell'epitelio differenziato.
Introduzione
Lesioni, infezioni e malattie dell'epitelio mucociliario sono associate a compromissione della produzione e della clearance del muco che si trova spesso in disturbi polmonari come malattie polmonari ostruzionistiche croniche, asma, fibrosi cistica, brochiectasi e discinesia ciliaria primaria1,2,3,4. Un recente progresso nella tecnologia organoide, ad esempio, l'organoide polmonare derivato dalla cellula basale chiamato tracheosfera che riassume la rigenerazione dell'epitelio mucociliare sorgono come un modello promettente con potenziale terapeutico1,5,6. Tuttavia, il suo uso è attualmente limitato, in parte a causa della mancanza delle condizioni di coltura definite e della bassa efficienza nelle produzioni organoidi. L'epitelio mucociliario nelle vie aeree umane e nell'epidermide della rana sono notevolmente simili nella morfologia dei tessuti, nella composizione cellulare e nella suafunzione 7,8,9,10,11,12. In entrambi gli organismi, l'epitelio mucociliare fornisce una difesa di prima linea secendo muco e sostanze antimicrobiche e cancella particelle nocive e agenti patogeni attraverso l'azione sincronizzata delle ciglia.
Qui descriviamo un semplice protocollo per generare organoidi epiteliali mucociliari usando i progenitori multipotenti degli embrioni xenopus laevis 13,14. In precedenza, abbiamoriferito 14 che in assenza di fattori di crescita esogeni e della matrice extracellulare, le cellule profonde isolate microchirurgicamente dall'ectoderma dello stadio gastrula precoce si assemblano spontaneamente in aggregati, rigenerano l'epitelio sulla sua superficie e maturano in epitelio mucociliare intercalando cellule multiciliate e altre cellule accessorie entro 24 ore. Oltre al rapido sviluppo, questo protocollo offre una distinta opportunità per accedere direttamente alle transizioni di cellule ectodermi profonde multipotenti in progenitori di cellule del calice epiteliale che riepilogano le fasi di rigenerazione di un epiteliointerrotto 14 che non sono disponibili da embrioni intatti ed ectoderma (noto anche come tappo animale)15,16,17. Il numero e le dimensioni degli organoidi prodotti sono scalabili con alta efficienza controllando i materiali di partenza dagli embrioni di Xenopus. Gli organoidi in coltura galleggiante possono essere facilmente ordinati e trasferiti nella fase desiderata per ulteriori analisi, tra cui imaging ad alta risoluzione, test meccanici, trattamento farmacologico e caratterizzazione genetica14. Questa rigenerazione spontanea, guidata dalla meccanica tissutale, dell'epitelio sulla superficie degli aggregati cellulari embrionali si traduce in organoidi epiteliali mucociliari e fornisce un nuovo modello tridimensionale (3D) per studiare la biologia dell'epitelio mucociliario.
Protocollo
L'uso degli animali e i protocolli sperimentali sono stati approvati dal comitato istituzionale per la cura e l'uso degli animali (IACUC) dell'Institute for Basic Science (IBS 18-01) e dal Korea Advanced Institute of Science and Technology (KA2017-22).
1. Embrioni
- Ottenere embrioni X. laevis utilizzando una procedura standard: raccogliere manualmente le uova dalle rane femminili stimolate ed eseguire la fecondazione in vitro18,19.
- Sgonlinare gli embrioni fecondati con delicata agitazione per circa 5 minuti in cisteina al 2% in soluzione salina di Barth modificata 1/3x (MBS; vedi la ricetta per 1X MBS sotto) a pH 819.
- Passaggio opzionale per l'imaging dal vivo: etichettare in modo fluorescente proteine specifiche e osservarne la dinamica nell'organoide, procedere alla sezione 5.
- (Facoltativo) Per monitorare la contaminazione delle cellule ectodermiche superficiali all'interno degli organoidi, etichettare la superficie apicali degli embrioni con NHS-rodiammina allo stadio 914. Incubare embrioni in 1 mg/mL NHS-Rhodamine in 1/3x MBS (pH 9.0) per 30 min con nutazione delicata. Lavare gli embrioni tre volte trasferendoli in una piastra di Petri riempita con 1/3x MBS per 15 minuti.
- Coltura dell'embrione in 1/3x MBS alla temperatura preferita (14-26 °C) fino a quando non vengono rilevati i primi segni dello stadio 10 (cioè la comparsa di cellule pigmentate scure intorno al blastopore alla vista vegetale).
2. Preparazione di strumenti, soluzioni e vasi di coltura microchirurgici
- Preparare gli strumenti necessari, tra cui un paio di forcellatrici chirurgiche e strumenti per capelli (anello per capelli e coltello per capelli) per la microchirurgia20.
- Preparare i seguenti supporti di coltura per gli embrioni: 1/3X MBS, dove 1X MBS è realizzato con NaCl (88 mM), KCl (1 mM), NaHCO3 (2,4 mM), MgSO4 (0,82 mM), Ca(NO3)2 (0,33 mM), CaCl2 (0,41 mM) e HEPES (10 mM). Regolare il pH a 7,4 con NaOH da 10 M.
NOTA: Facoltativo: aggiungere gocce di rosso fenolo per indicare il pH. - Preparare i seguenti mezzi di coltura per tessuti embrionali e organoidi: Danilchik's per Amy (DFA)21 integrato con soluzione antibiotica e antimicotica fresca all'1%. Preparare il DFA con NaCl (53 mM), Na2CO3 (5 mM), gluconato di potassio (4,5 mM), gluconato di sodio (32 mM), CaCl2 (1 mM) e MgSO4 (1 mM). Regola il pH a 8,3 con bicine granulare. Filtrare DFA (filtro bottle-top da 0,2 μm), aliquota e conservarlo a -20 °C.
- Preparare il DFA privo di calcio e magnesio per separare le cellule profonde dall'ectoderma utilizzando la ricetta sopra e omettendo CaCl2 e MgSO4. Aliquota e conservare a-20 °C.
- Preparare tubi PCR non adesivi per l'aggregazione di cellule embrionali.
- Per indurre l'aggregazione spontanea delle cellule embrionali isolate, preparare tubi PCR non adesivi rivestendo tubi PCR a fondo rotondo con 200 μL di 1% di BSA (1 g di BSA in 100 ml di acqua distillata) durante la notte a 4 °C o per 2 ore a temperatura ambiente. Ogni tubo verrà utilizzato per assemblare un organoide.
- Risciacquare tre volte i tubi PCR rivestiti con BSA con DFA per rimuovere eventuali BSA residui.
- Riempire i tubi PCR con 200 μL di DFA.
3. Isolamento delle cellule ectodermi profonde
- Seleziona e raccogli embrioni mentre raggiungono la fase iniziale 10 usando gli strumenti per capelli sotto uno stereoscopio.
- Utilizzando una pipetta di trasferimento usa e getta, trasferire gli embrioni selezionati in una piastra di Petri riempita di DFA.
- Rimuovere la membrana vitellina degli embrioni usando forcep affilate dal lato vegetale senza interrompere il lato animale dell'embrione.
NOTA: Fare attenzione a evitare di esporre embrioni all'aria. Introdurre bolle d'aria nella soluzione o portare embrioni in superficie farà scoppiare l'embrione. - Per isolare il cappuccio animale, posizionare il lato animale dell'embrione verso l'alto.
- Stima visivamente l'estensione del cappuccio animale da asportare e fai la prima incisione lungo il bordo del cappuccio dell'animale con un coltello per capelli. Tirare il coltello per capelli verso l'esterno per fare un taglio.
- Ripetere il passaggio 3.5 per creare una catena di piccoli tagli per asportare il tappo animale.
- Tagliare il bordo a strati spessi del cappuccio animale usando un coltello per capelli per prevenire l'inclusione di precursori del mesoderma.
NOTA: Per prevenire la guarigione e l'aggregazione di tappi animali isolati, procedere al passaggio successivo entro 10 minuti. Tipicamente, isoliamo 5-10 tappi per animali alla volta per assemblare più organoidi. - Per separare le cellule ectodermi profonde dal tappo animale, trasferire i tappi degli animali asportati in una piastra di Petri piena di DFA senza calcio e magnesio con una pipetta di trasferimento usa e getta. Fare attenzione a non introdurre bolle d'aria durante il trasferimento.
- Per mantenere abbastanza spazio per i passaggi successivi, utilizzando gli strumenti per capelli, posizionare i tappi degli animali per affrontare il lato animale verso l'alto e mantenere una distanza generosa da altre espianto.
- Attendere 5-10 minuti e quindi monitorare gli espianto sotto uno stereoscopio. Una volta che le cellule profonde allentate sono state trovate dal bordo dello strato superficiale pigmentato scuro, inizia a sollevare lo strato superficiale lontano dalle cellule ectodermi profonde di colore chiaro usando un coltello per capelli sotto lo stereoscopio.
- Staccare con cura (staccare) lo strato superficiale con un coltello per capelli, a partire dal bordo.
- Raccogliere le cellule ectodermi profonde con aspirazione minima (10\u201215 μL) per limitare la quantità di DFA privo di calcio e magnesio che viene trasferita al supporto di aggregazione nel passaggio successivo.
NOTA: Le cellule superficiali staccate possono essere rimosse dal supporto per evitare di contaminare le restanti cellule ectodermi profonde.
4. Generazione di organoidi epiteliali mucociliari
- Trasferire le cellule ectodermi profonde raccolte in un tubo PCR non adesivo contenente 200 μL di DFA. Pipettare delicatamente il supporto (2\u20123 volte) per disperdere le celle trasferite nel tubo PCR.
NOTA: la tempistica designata come ore dopo l'aggregazione (hpa) inizia da questo passaggio. La dimensione degli organoidi risultanti è controllata dal numero di cellule ectodermi profonde aggiunte a un tubo PCR. È possibile utilizzare cellule ectodermi profonde da uno o più tappi animali, a seconda delle dimensioni desiderate dell'organoide. - Chiudere il tubo PCR. Mantenere i tubi PCR in posizione verticale per indurre l'aggregazione spontanea nella parte inferiore.
- Monitorare il processo di aggregazione sotto uno stereoscopio. Le cellule in genere si raccolgono nella parte inferiore del tubo PCR entro un'ora e si assemblano in aggregati sferici entro 2-3 ore, a seconda delle dimensioni.
- Per condurre test di imaging vivo o farmaci durante lo sviluppo degli organoidi epiteliali mucociliari, raccogliere aggregati a 2 hpa utilizzando una pipetta da 200 μL dotata di punte ingrandite (tagliate con forbice sterilizzata) per evitare di causare danni agli aggregati durante la raccolta.
- Per consentire agli aggregati di svilupparsi nell'organoide epiteliale mucociliario in coltura, raccogliere aggregati sferici dal tubo PCR a 5 hpa e trasferirli in una piastra di Petri riempita di DFA.
- Posizionare gli aggregati lontano dagli altri per evitare che si fumino. Entro 24 ore dalla coltura a temperatura ambiente senza fattori aggiunti, si può osservare che gli organoidi epiteliali mucociliari maturi ruotano con l'azione di battere le ciglia che coprono la superficie dell'epitelio differenziato
5. Imaging vivo ad alta risoluzione ad alta risoluzione di organoidi in via di sviluppo
- Preparare l'mRNA per la microiniezione.
- Per visualizzare l'epitelializzazione che si verifica nella fase iniziale della formazione organoide, preparare l'mRNA per la proteina zonula specifica dell'epiteliale-1 (ZO-1) e per delineare le membrane cellulari amplificando i plasmidi pCS2-ZO1-RFP e pCS2-mem-GFP (un dono di Lance Davidson).
- Estrarre e linearizzare il DNA plasmide, quindi trascrivere l'mRNA chiuso utilizzando un kit di trascrizione in vitro SP6/T7.
- Aliquota l'mRNA trascritto e conservarlo a -80 °C.
- Microiniettare l'mRNA in un embrione fecondato
- Posizionare embrioni fecondati in 3% Ficoll in 1x MBS.
- Caricare 3-4 μL di mRNA utilizzando una punta del micro-caricatore in un ago di vetro tirato (una punta dell'ago affusolata lunga e fine con diametro interno di 10\u201230 μm).
- Attaccare l'ago a un microiniettore e regolare il tempo e la pressione per fornire un volume costante di mRNA per la microiniezione.
- Iniettare l'mRNA appena sotto la superficie apicale del palo animale. Una distinta macchia circolare di colore pallido causata dall'espansione della corteccia è visibile al momento della microiniezione.
- Trasferire gli embrioni iniettati a 1/3X MBS e coltarli allo stadio 9.5.
- Raccogliere gli embrioni etichettati fluorescentmente sotto uno stereoscopio a fluorescenza (impostazioni di eccitazione/emissione per GFP (488/510) e RFP (532/588)).
- Procedere con il passaggio 1.3.
- Assemblare e coltura l'organoide (sezioni 3 e 4) fino allo stadio di sviluppo desiderato.
- Esegui immagini dal vivo.
- Preparare una camera di imaging con fondo di vetro incollando un vetro di copertura a una camera acrilica fresata su misura utilizzando grasso di silicio.
NOTA: Sigillare saldamente la camera per evitare perdite dei mezzi di coltura. - Riempire la camera di imaging con DFA.
- Scegliere una griglia di microscopia elettronica a trasmissione esagonale (TEM) (75 mesh) da un contenitore utilizzando le forcep e applicare una piccola quantità di grasso sul bordo della griglia.
NOTA: La dimensione della mesh deve essere inferiore al diametro dell'aggregato in modo che l'aggregato si sieda sulla griglia. - Premere leggermente verso il basso per fissare la griglia TEM sul fondo della camera di imaging.
- Trasferire gli aggregati nella camera di imaging e posizionarli all'interno della griglia.
NOTA: Evitare di posizionare gli aggregati accanto al grasso. Durante l'esperimento, gli aggregati dovrebbero sedersi sulla griglia TEM senza contattare il fondo della camera per prevenire la compressione fisica. - Riempire la camera di imaging con DFA e sigillarla con un vetro di copertura e grasso.
NOTA: La camera deve essere ermetica senza bolle d'aria per evitare turbolenze o movimenti durante l'imaging. - Per seguire la progressione della formazione organoide epiteliale mucociliaria, raccogliere immagini z-stack time-lapse di aggregati (da 2 hpa) utilizzando un microscopio confocale.
NOTA: In genere raccogliamo z-stack spessi ~120 μm ogni 15 minuti usando un obiettivo 20X per seguire i comportamenti dinamici delle cellule, ma queste specifiche dovrebbero essere ottimizzate per l'obiettivo degli esperimenti.
- Preparare una camera di imaging con fondo di vetro incollando un vetro di copertura a una camera acrilica fresata su misura utilizzando grasso di silicio.
6. Imaging (facoltativo) Sviluppo di organoidi mediante fissazione e immunostaining
- Fissare gli organoidi nella fase di sviluppo desiderata trasferendoli in una fiala di vetro riempita con una soluzione fissante.
NOTA: aggiungere un volume di soluzione fissante >20 volte superiore a quello dei campioni per garantire la correzione completa. Eseguire i seguenti processi su un nutator, se non diversamente indicato. In generale, gli organoidi sono fissati con il 4% di paraformaldeide (PFA) in PBS. Tuttavia, possono essere necessari diversi fissivi per rilevare proteine specifiche. Ad esempio, abbiamo usato il 4% di PFA con lo 0,25% di glutaraldeide in PBS per rilevare f-actina e tubulina acetilata. Per rilevare l'intelectina (ITLN) e lo ZO-1, gli organoidi sono fissati con la soluzione di Dent ghiacciato (4:1 metanolo:dimetil solfossido) per la notte a -20 °C. Gli organoidi fissi di Dent devono essere disidratati in serie prima del lavaggio (fase 6.3). La durata per l'incubazione e il lavaggio degli anticorpi può essere ottimizzata per esigenze specifiche. - Fissare gli organoidi per 15 minuti a temperatura ambiente (RT) o durante la notte a 4 °C.
- Lavare 3 volte con PBST (PBS con 0,1% Triton X-100) per 15 minuti a RT.
- Blocca legame non specifico con siero di capra al 10% in PBST (PBSGT) per 1 h a RT.
- Incubare con anticorpo primario (1:200) in PBSGT durante la notte a 4 °C.
- Lavare 3 volte con PBST per 15 minuti a RT.
- Incubare con anticorpo secondario (1:200) in PBSGT durante la notte a 4 °C.
- Lavare 3 volte con PBST per 15 minuti a RT.
- Trasferire gli organoidi fissi e immunosostenibili in una camera di imaging e procedere con l'imaging confocale.
Risultati
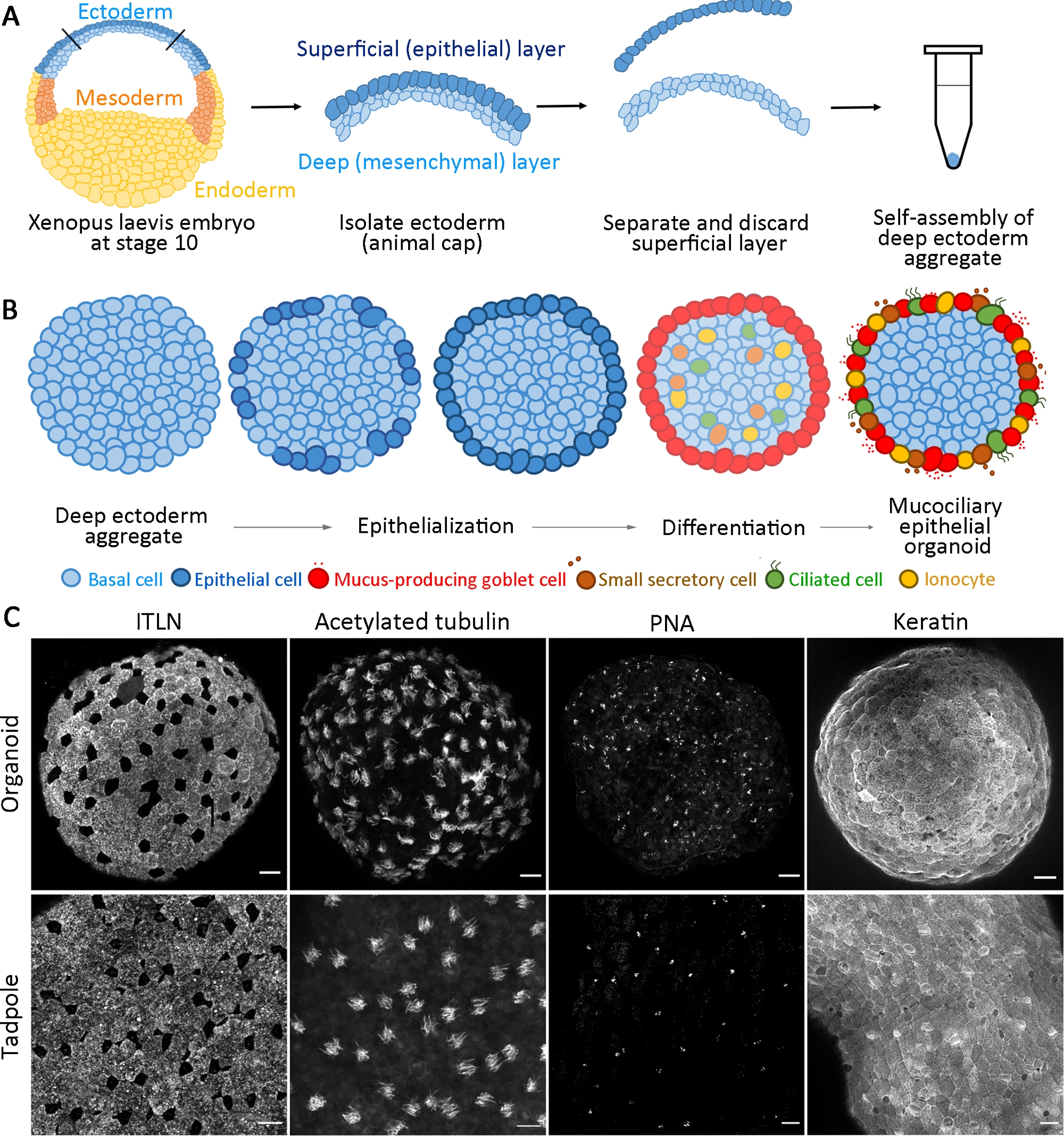
Questo protocollo standardizzato genera un organoide epiteliale mucociliare da progenitori multipotenti isolati dai primi embrioni di gastrula X. laevis entro 24 ore dalla coltivazione14. Le cellule ectodermi profonde raccolte si auto-assemblano per formare un aggregato in un tubo PCR non adesivo e subiscono l'epitelializzazione superficiale e la differenziazione delle cellule del calice. La superficie di aggregati appena epitelializzata fornisce un substrato simile all'epitelio nativo trovato in vivo per l'intercalazione delle cellule interne (ad esempio, cellule multiciliate e altre cellule accessorie) e si sviluppa per formare organoidi epiteliali mucociliari(Figura 1A,B). Entro 24 ore dall'aggregazione, gli organoidi epiteliali mucociliari auto-organizzati rigenerano un'epidermide matura indistinguibile dall'epidermide di un girino. Gli organoidi comprendono epitelio completamente differenziato (cheratina), cellule di calice che secernono muco (ITLN), cellule multiciliate (tubulina acetilata) e piccole cellule secretorie (agglutinina di arachidi, PNA)(Figura 1C).
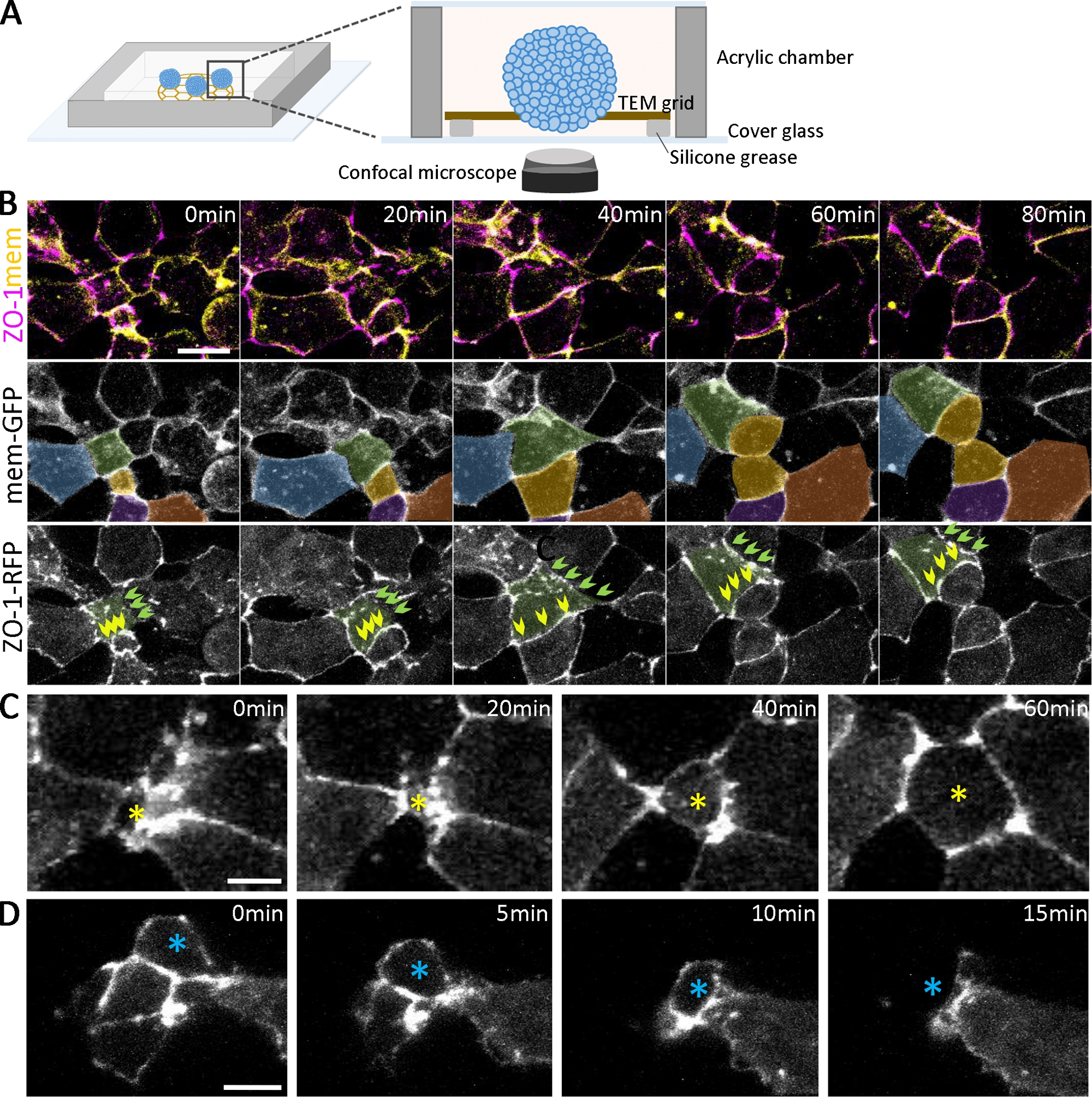
Oltre a confermare lo sviluppo di diversi tipi di cellule con immunosottenzione, la dinamica dello sviluppo organoide può essere seguita dall'imaging dal vivo (Figura 2A). Per esaminare l'epitelializzazione che emerge nella fase iniziale della formazione organoide (Figura 1B), abbiamo etichettato gli embrioni esprimendo proteine di giunzione stretta taggate fluorescentmente (ZO-1-RFP) e proteine di localizzazione della membrana (mem-GFP). Con la doppia etichettatura, i passaggi sequenziali della formazione di giunzione stretta ZO-1-positiva possono essere marcati e analizzati quantitativamente durante l'epitelializzazione (Figura 2). Ad esempio, per le cellule(Figura 2B, di colore verde) in diverse fasi di epitelializzazione (a 0 min), alcune regioni di adesione cellula-cellula hanno puncta sparsi di ZO-1 (Figura 2B, frecce verdi). Al contrario, altre aree hanno un'espressione ZO-1 completamente assemblata e contigua (Figura 2B, frecce gialle). Nel corso del tempo, la puncta si coalizza e si collega per formare giunzioni strette contigue(Figura 2B,frecce verdi) e giunzioni strette contigue mantengono la loro morfologia anche durante la divisione cellulare(Figura 2B,frecce gialle). Man mano che le giunzioni strette maturano, le cellule si muovono dinamicamente all'interno e all'uscita dalla superficie lungo i piani apicali degli organoidi (Figura 2C,D). Inoltre, tracciando le cellule in modo spaziotemporale sulla superficie degli organoidi differenzianti(Figura 2B, celle codificate a colori), è possibile un'analisi su più scale, che vanno dai singoli puncta alle giunzioni strette contigue, ai confini cellulare-cellulari e ai sottoinsiemi di popolazioni cellulari all'interno degli organoidi.

Figura 1: Generazione di organoidi epiteliali mucociliari.
(A) Schema che mostra il protocollo per assemblare aggregati di ectoderma profondi da embrioni X. laevis. (B) Schema per un modello di formazione organoide epiteliale mucociliaria originata da cellule ectodermi profonde multipotenti (vista trasversale). Le cellule posizionate in superficie transitano nelle cellule epiteliali e si differenziano in cellule di calice. Differenziare le cellule ciliate, le cellule secretorie e gli ionociti si intercalano radialmente nella superficie e rigenerano un'epidermide matura. (C) Proiezione z massima dell'epitelio mucociliare immunosostituito per ITLN (cellule di calice che producono muco), tubulina acetilata (cellule ciliate), PNA (piccole cellule secretorie) e cheratina (cellule epiteliali) in organoidi a 24 hpa (pannello superiore) ed epidermide girino (pannello inferiore). Barra di scala = 30 μm. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}

Figura 2: Imaging dal vivo degli organoidi in via di sviluppo.
(A) Schema della camera di imaging per organoidi vivi (non in scala). (B) Sequenze time-lapse di pile confocali raccolte da aggregati cellulari ectodermi profondi che esprimono ZO-1-RFP e mem-GFP da 2,5 hpa. Barra di scala = 20 μm. Le celle sono pseudocolori per il tracciamento nel tempo. Le celle di colore verde hanno diversi stati di adesione cellulare, tra cui una che sviluppa progressivamente l'adesione positiva ZO-1 (frecce verdi) e una mantenendo l'adesione positiva contigua zo-1 (frecce gialle) nel tempo. (C, D) Le immagini confocali time-lapse degli aggregati cellulari ectodermi profondi che esprimono ZO-1-RFP mostrano le cellule radialmente intercalanti che si muovono verso la superficie (C, stella gialla) e si muovono all'interno degli aggregati (D, stella blu). Barre di scala = 10 μm. Clicca qui per visualizzare una versione più grande di questa figura.
{kind=link}
Discussione
Gli organoidi epiteliali mucociliari generati da cellule ectodermi profonde dell'embrione X. laevis sono un potente strumento per studiare l'epitelializzazione e la differenziazione dei progenitori multipotenti in vitro. In contrasto con il saggio di tappi animaliampiamente adottato 16 utilizzato per l'organogenesi in vitro13 e lo sviluppo dell'epitelia mucociliale15,17,22 che utilizzano l'ectoderma intatto, gli organoidi profondi derivati dall'ectoderma presentati in questo protocollo offrono una distinta opportunità per monitorare le fasi di rigenerazione guidate dalla meccanica tissutale dell'epiteliosuperficiale 14. A circa 2 hpa, le cellule epiteliali positive ZO-1 appena generate (Figura 2) iniziano ad apparire sulla superficie apicale degli organoidi e aumentano la loro popolazione per coprire l'intero organoide man mano che il tessuto si solidifica o riduce laconformità 14. La rigenerazione dell'epitelio e le successive specifiche di lignaggio per le cellule del calice che producono muco procedono spontaneamente in un supporto di coltura definito chimicamente entro un giorno. Questi organoidi epiteliali mucociliari in rapido sviluppo forniscono una piattaforma per esaminare i comportamenti cellulari dinamici in tempo reale, ad alta risoluzione, durante i passaggi progressivi della rigenerazione epiteliale. Consentono inoltre di investigare le questioni fondamentali che sorgono durante lo sviluppo dell'epitelio mucociliario, l'omeostasi e le malattie associate2,9,23. In particolare, la sensibilità meccanica delle cellule progenitrici profonde durante il passaggio ai precursori delle cellule del calice epiteliale identificati negli organoidi14 può servire a collegare malattie respiratorie associate a una differenziazione basale anomala in cui le cellule del calice che secernono muco sono sovra- o sotto-prodotte23.
Mentre questo protocollo offre un approccio semplice per generare questi organoidi, ci sono diversi passaggi critici per il successo negli esperimenti. Per prevenire la contaminazione delle cellule epiteliali superficiali durante l'isolamento delle cellule ectodermi profonde dal cappuccio animale, è necessario monitorare il cappuccio animale posto in DFA privo di calcio e magnesio sotto uno stereoscopio e rilevare il momento giusto per iniziare la separazione dello strato superficiale pigmentato scuro del cappuccio animale. Se il tessuto viene conservato in DFA privo di calcio e magnesio per troppo tempo, l'intero tessuto si dissocia e distinguere tra cellule profonde e superficiali sarebbe quindi impossibile per gli aggregati di ectoderma profondi. Per confermare l'assenza di cellule superficiali negli aggregati ectodermi profondi, si consiglia di etichettare fluorescentmente la superficie apicali dell'embrione con NHS-rodiammina (fase1.4 14) prima della microchirurgia; ciò consentirebbe una facile identificazione delle cellule superficiali se esistono negli organoidi risultanti. Poiché la rigenerazione epiteliale è regolata dallameccanica tissutale 14, è essenziale evitare la generazione involontaria di forza per organoidi auto-organizzanti. In particolare, suggeriamo di evitare il contatto con il fondo di vetro della camera di imaging durante l'imaging dal vivo posizionando aggregati ai bordi delle griglie TEM in quanto ciò consente il libero contatto con la finestra di imaging degli aggregati vivi (fase 5.1.2.). Questo modello 3D in vitro-coltivato e auto-organizzato per l'epitelio mucociliario servirà come strumento trattabile per rispondere alle domande fondamentali che sorgono durante la rigenerazione dell'epitelio e la specifica del lignaggio delle cellule di calice.
Divulgazioni
Gli autori non hanno nulla da rivelare.
Riconoscimenti
Ringraziamo i membri del kim lab e Lance Davidson per i loro commenti e il loro supporto. Questo lavoro è stato supportato da Young Scientist Fellowship a HYK dell'Institute for Basic Science (IBS-R0250Y1).
Materiali
| Name | Company | Catalog Number | Comments |
| Equipment | |||
| Dual-stage Glass Micropipette Puller | Narishige | PC-100 | |
| Picoliter microinjector | Warner Instruments | PLI-100A | |
| Confocal Laser Microscope | |||
| Stereoscope | |||
| Tools | |||
| Forcep | Dumont | Dumont #5 | |
| Hair knife | Reference (Kay, B.K.; Peng, H.B., 1991) | ||
| Hair loop | Reference (Kay, B.K.; Peng, H.B., 1991) | ||
| hCG injection | |||
| human chorionic gonadotropin | Sigma | cg10-10vl | |
| MBS solution | |||
| 10M Sodium hydroxide | Sigma | 72068 | |
| Calcium chloride | Sigma | C3881 | |
| Calcium nitrate | Sigma | C1396 | |
| HEPES | Sigma | H4034 | |
| Magnesium sulfate | Sigma | 230391 | |
| Phenol-red | Sigma | P0290 | |
| Potassium chloride | Sigma | 7447-40-7 | |
| Sodium bicarbonate | Sigma | S6014 | |
| Sodium chloride | Sigma | S9625 | |
| Sodium hydroxide reagent grade, 97%, powder-25g | Sigma | 655104 | |
| dejellying solution | |||
| L-Cysteine hydrochloride monohydrate | Sigma | C7880 | |
| Sodium hydroxide 10M | Sigma | 72068 | |
| Ficoll solution | |||
| Ficoll | Sigma | F4375 | |
| DFA solution | |||
| Sodium chloride | Sigma | S9625 | |
| 0.22mm Filter | Millipore | S2GPT05RE | |
| Antibiotic Antimycotic Solution | Sigma | A5955 | |
| Bicine | Sigma | B3876 | |
| Calcium chloride | Sigma | C3881 | |
| Magnesium sulfate | Sigma | 230391 | |
| Potassium gluconate | Sigma | G4500 | |
| Sodium carbonate | Sigma | 222321 | |
| Sodium gluconate | Sigma | G9005 | |
| mRNA in vitro transcription | |||
| SP6/T7 in vitro transcription kit | Invitrogen | AM1340 | |
| mRNA microinjection | |||
| Borosilicate glass capillary tubes | Harvard Apparatus | GC100-10 | |
| Eppendorf microloader pipette tips | ThermoFisher | A25547 | |
| Mineral oil | Sigma | M5904 | |
| PCR tube coating | |||
| BSA | Thermofisher | 26140079 | |
| PCR tubes | SSI | SSI-3245-00 | |
| Imaging | |||
| Custom-milled acrylic chamber | |||
| Coverglass 24mmX50mm | Duran | B01_001650 | |
| SPI Hexagonal TEM Grids, Gilded Nickel (50mesh) | SPI | 275HGN-XA | |
| SPI Hexagonal TEM Grids, Gilded Nickel (75mesh) | SPI | 2775GN-XA | |
| Silicone grease | Shinetsu | HIVAC-G | |
| Fixation | |||
| 20ml screw top-cap vial | Wheaton | WH.986580 | |
| 2ml screw top-cap vial | |||
| Benzyl alcohol | Sigma | 305197 | |
| Benzyl benzoate | Sigma | B6630 | |
| Dimethyl sulfoxide (DMSO) | Sgima | D4540 | |
| Glutaraldehyde 10% EM GRADE | Electron Microscopy Sciences | 16120 | |
| Goat serum | Jackson | 005-000-121 | |
| Methanol | Sigma | 322415 | |
| Paraforlamdehyde | Sigma | P6148 | |
| Phosphate-buffered saline (PBS) | LPS Solution | CBP007B | |
| Triton X-100 | Sigma | T8787 | |
| Primary antibody (1:200) | |||
| acetylated tubulin | Sigma | clone 6-11B-1 | |
| Itln1 | Proteintech | 11770-1-AP | |
| Keratin | Developmental Studies Hybridoma Bank | 1h5 | |
| ZO1 | Invitrogen | 402200 | |
| Vectors | |||
| pCS2-mem-GFP | Gift from Dr. Lance Davidson | ||
| pCS2-ZO1-RFP | Gift from Dr. Lance Davidson |
Riferimenti
- Barkauskas, C. E., et al. Lung organoids: current uses and future promise. Development. 144 (6), 986-997 (2017).
- Puchelle, E., Zahm, J. M., Tournier, J. M., Coraux, C. Airway Epithelial Repair, Regeneration, and Remodeling after Injury in Chronic Obstructive Pulmonary Disease. Proceedings of the American Thoracic Society. 3 (8), 726-733 (2006).
- Tilley, A. E., Walters, M. S., Shaykhiev, R., Crystal, R. G. Cilia dysfunction in lung disease. Annual Review of Physiology. 77, 379-406 (2015).
- Vareille, M., Kieninger, E., Edwards, M. R., Regamey, N. The Airway Epithelium: Soldier in the Fight against Respiratory Viruses. Clinical Microbiology Reviews. 24 (1), 210-229 (2011).
- Rock, J. R., et al. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proceedings of the National Academy of Sciences of the United States of America. 106 (31), 12771-12775 (2009).
- Sachs, N., et al. Long-term expanding human airway organoids for disease modeling. The EMBO Journal. 38 (4), 100300(2019).
- Dubaissi, E., et al. A secretory cell type develops alongside multiciliated cells, ionocytes and goblet cells, and provides a protective, anti-infective function in the frog embryonic mucociliary epidermis. Development. 141 (7), 1514-1525 (2014).
- Hayes, J. M., et al. Identification of novel ciliogenesis factors using a new in vivo model for mucociliary epithelial development. Developmental Biology. 312 (1), 115-130 (2007).
- Walentek, P., Quigley, I. K. What we can learn from a tadpole about ciliopathies and airway diseases: Using systems biology in Xenopus to study cilia and mucociliary epithelia. Genesis. 55 (1-2), (2017).
- Werner, M. E., Mitchell, B. J. Understanding ciliated epithelia: The power of Xenopus. Genesis. 50 (3), 176-185 (2012).
- Dubaissi, E., Papalopulu, N. Embryonic frog epidermis: a model for the study of cell-cell interactions in the development of mucociliary disease. Disease Models & Mechanisms. 4 (2), 179-192 (2011).
- Walentek, P., et al. A novel serotonin-secreting cell type regulates ciliary motility in the mucociliary epidermis of Xenopus tadpoles. Development. 141 (7), 1526-1533 (2014).
- Asashima, M., et al. In vitro organogenesis from undifferentiated cells in Xenopus. Developmental Dynamics. 238 (6), 1309-1320 (2009).
- Kim, H. Y., Jackson, T. R., Stuckenholz, C., Davidson, L. A. Tissue mechanics drives regeneration of a mucociliated epidermis on the surface of Xenopus embryonic aggregates. Nature Communications. 11 (1), 665(2020).
- Haas, M., et al. DeltaN-Tp63 Mediates Wnt/beta-Catenin-Induced Inhibition of Differentiation in Basal Stem Cells of Mucociliary Epithelia. Cell Reports. 28 (13), 3338-3352 (2019).
- Green, J. Molecular Methods in Developmental Biology: Xenopus and Zebrafish. Guille, M. , Humana Press. 1-13 (1999).
- Stubbs, J. L., Davidson, L., Keller, R., Kintner, C. Radial intercalation of ciliated cells during Xenopus skin development. Development. 133 (13), 2507-2515 (2006).
- Nieuwkoop, P. D., Faber, J. Normal table of Xenopus laevis (Daudin) : a systematical and chronological survey of the development from the fertilized egg till the end of metamorphosis. , Garland Pub. (1994).
- Sive, H. L., Grainger, R. M., Harland, R. M. Early development of Xenopus laevis : a laboratory manual. , Cold Spring Harbor Laboratory Press. (2000).
- Joshi, S. D., Kim, H. Y., Davidson, L. A. Microscopy tools for quantifying developmental dynamics in Xenopus embryos. Methods in Molecular Biology. 917, 477-493 (2012).
- Sater, A. K., Steinhardt, R. A., Keller, R. Induction of neuronal differentiation by planar signals in Xenopus embryos. Developmental Dynamics. 197 (4), 268-280 (1993).
- Sedzinski, J., Hannezo, E., Tu, F., Biro, M., Wallingford, J. B. Emergence of an Apical Epithelial Cell Surface In Vivo. Developmental Cell. 36 (1), 24-35 (2016).
- Rock, J. R., Randell, S. H., Hogan, B. L. M. Airway basal stem cells: a perspective on their roles in epithelial homeostasis and remodeling. Disease Models & Mechanisms. 3 (9-10), 545-556 (2010).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati