
Method Article
細胞細胞性グルタチオンSトランスレセファーゼの潜在的阻害剤に対する分光測定法
* これらの著者は同等に貢献しました
要約
グルタチオンSトランスレセファーゼ(GST)は、多数の化学療法薬の代謝に関与する解毒酵素である。GSTsの過剰発現は、癌化学療法抵抗と相関している。この表現型に対抗する一つの方法は、阻害剤を使用することである。このプロトコルは、分光光測定アッセイを用いて、潜在的なGST阻害剤をスクリーニングする方法を記述する。
要約
グルタチオンSトランスレポ酵素(GST)は、グルタチオン(GSH)結合による内因性または外因性の求水化合物の除去を担う代謝酵素である。さらに、GSTは、アポトーシス経路に関与するマイトゲン活性化プロテインキナーゼ(MAPK)の調節因子です。GSTsの過剰発現は、電気的アルキル化剤を用いた化学療法を受けている患者の治療効果の低下と相関している。GST阻害剤を使用すると、この傾向を逆転させ、治療効力を増強する潜在的な解決策となり得る。この目標を達成するには、正確で迅速かつ容易な酵素アッセイを使用して、このような化合物の発見が必要です。1-クロロ-2,4-ジニトロベンゼン(CDNB)を基質として使用した分光光度法は、文献において最も採用されている方法である。しかしながら、既に説明されているGST阻害実験は、CDNBに対するミカリス・メンテン定数(Km)の測定や、使用された酵素濃度の指標など、最適な阻害アッセイの各段階を詳述するプロトコルを提供していない、試験化合物の阻害効力を評価するための重要なパラメータである。したがって、このプロトコルを用いて、最適化された分光光度GST酵素アッセイの各ステップを、潜在的な阻害剤のライブラリーをスクリーニングする方法について説明する。酵素阻害薬の効力を測定するために用いられる2つの特徴である、半最大抑制濃度(IC50)と一定の阻害(Ki)の両方の計算について説明します。記載された方法は、細胞または純粋組換えヒトGST、すなわちGSTアルファ1(GSTA1)、GST μ1(GSTM1)またはGST pi 1(GSTP1)から抽出されたGSTのプールを使用して実施することができる。ただし、このプロトコルは、このアイソフォームの基質ではないCDNBのため、GSTのsta1(GSTT1)には適用できません。この方法は、馬の肝臓からのGSTを使用してクルクミンの阻害効力をテストするために使用されました.クルクミンは、抗癌特性を示す分子であり、 シリコ ドッキング予測の後GSTアイソフォームに対する親和性を示した。我々は、クルクミンが31.6±3.6μMのIC50および23.2±3.2 μMのKi を有する強力な競争力のあるGST阻害剤であることを実証した。
概要
細胞細胞性グルタチオンS-トランストランス酵素(GSTs,EC2.5.1.18)は、グルタチオン(GSH)を化学療法剤などの種々の電子球性化合物に結合させ、体から容易に解毒して排除する。細胞細胞系GSTの7つのアイソフォームは、α、ミュー、パイ、シグマ、オメガ、シータ、およびゼータとして同定されている。GSTは主に肝臓、精巣、肺、および胃腸管2で発現する。GST α 1 (GSTA1) アイソフォームは肝細胞で高発現します。体は異種的に発現する他のサブタイプは、主に脳、心臓、肺中のGST pi1(GSTP1)、および肝臓におけるGST μ1(GSTM1)および精巣3を含む。GSTアイソフォーム間には高い配列相同性があるが、それぞれが基質特異性を示し、異なる方法で薬物代謝と癌に関与している、その差動発現44、5に従う。
電気的化合物は外因的に体内に入るか、内因的に産生される。農薬、プロスタグランジン、発がん性物質、および化学療法薬は、グルタチオン共役反応の潜在的な基質の一部である。例えば、細胞内に形成された電子欠損反応性化合物は、電子基質になる可能性が高い。クロラムブシルやメルファランなどのアルキル化剤は、GSTsによって触媒されるGSHのコンジュゲートとして排除され、これらの酵素の増加レベルはこれらの化合物66,77に対する耐性と相関している。
細胞質GSTのもう一つの重要な役割は、MAPK8(c-Jun N末端キナーゼ、またはJNK1とも呼ばれる)およびMAP3K5(アポトーシスシグナル調節キナーゼ1、またはASK1)8などのマイトゲン活性化タンパク質キナーゼ(MAPK)の活性を調節することです。それらの単量体の構造中のいくつかのアイソフォームは、これらのタンパク質に結合し、したがって、リン酸化カスケードをブロックします。通常の条件下では、GSTP1アイソフォームはMAPK8(c-Junタンパク質の活性化剤)を隔離する。c-Junとc-Fosタンパク質を組み合わせることで、アクチベータータンパク質1(AP-1)転写因子が形成され、これはプロアプトティック遺伝子の転写を担う。ストレス細胞において、GSTP1とMAPK8により形成された複合体は解化し、c-Junが活性化され、そしてアポトーシスに至る遺伝子が9に発現し始める。したがって、このGSTアイソフォームの発現が大きく、経路がブロックされ、細胞生存率の増加、細胞増殖の増加、化学療法に対する細胞感受性の低下につながる可能性があります。同様のシナリオは、GSTP1 のパラログ (たとえば、GSTM1、 MAP3K510と対話する) で発生する可能性があります。
GSTが薬物代謝およびMAPの隔離において果たした役割は、GSTの発現が化学療法治療66,1111に対する腫瘍抵抗機構の徴候であるかもしれないという仮説につながった。例えば、GSTP1は多数の癌において過剰発現しており、その存在は予後不良と再発率8の増加と相関している。これらの遺伝子の多型はまた、様々な疾患を提示する患者のための差動薬物暴露および生存率を示しており、これらの酵素は薬剤耐性のメカニズムにとって極めて重要であるという考えを強化している。例えば、GSTM1ヌル遺伝子型を有する個体は、より低い薬物クリアランスおよびよりよい生存率12、13,13と関連している。GSHアナログの使用、GSTsとの共役によって活性化されるプロドラッグ、または直接GST阻害剤14、15,15など、この過剰発現に対抗するいくつかの潜在的な手段がある。
これらの方法はすべて現在調査中であり、いくつかの化合物は患者の潜在的な使用のための臨床試験を開始しました。しかし、我々の知る限りでは、臨床現場15においてGST阻害剤として使用されている化合物はない。実際、ある種のアイソフォームに対する特異性の欠如、または正常細胞におけるGSHの枯渇は、器官系における活性酸素種(ROS)の蓄積によって引き起こされる毒性を引き起こし得るが、GST阻害剤14,15,15の可能性を低下させる欠点の一部に過ぎない。これらの化合物が体に他の薬力学的効果を発揮する可能性があるリスクも、その使用を制限しています。.例えば、エタクリン酸は、実験室環境で最も広く研究されているGST阻害剤ですが、主に強い利尿薬として使用されているため、この特性は臨床現場での他の薬剤との組み合わせでの使用を制限します。クルクミンは、GST阻害剤として正常にスクリーニングされた別の天然化合物です。この分子は、ウコンのクルクマロンガ種から抽出されたポリフェノールエーテルです。.様々な種類の腫瘍細胞株16,17のアポトーシスを誘導することにより癌に対する可能な治療選択肢として有望な結果を17示している。この化合物は、チロシンキナーゼ18またはGST経路などの多様な細胞経路を調節することができる。純粋なタンパク質を使用した研究は、GSTA1、GSTM1およびGSTP119、20,20に対する阻害力を示している。しかし、癌細胞では相反する結果が認められ、細胞がクルクミン21で治療された場合に細胞内GST活性が大きく測定された。したがって、さらなる細胞実験を計画する前に、適切な制御を用いて明確に記述されたプロトコルを使用して、半分の最大阻害濃度(IC50)および一定の阻害(Ki)を調べることが重要です。
したがって、潜在的な新しいGST阻害剤のスクリーニングおよび検査は重要な臨床的関心であり、新しい化合物は、電子性薬物と組み合わせて使用するために安全かつ効率的でなければならない。アイソフォーム特異的阻害剤に焦点を当てた研究は、GST発現の特定のパターンを示す腫瘍組織におけるGST阻害を可能にし、したがって有効な併用療法の開発を可能にする。阻害の差動モードを有する阻害剤を見つけることも関心を持つかもしれない。例えば、GSHを基質として使用する競合性阻害剤は、その枯渇を誘発する可能性がある。この細胞におけるGSH濃度の低下は、ニューロンの酸化ストレスを誘発し、アポトーシスを引き起こす。もう一つの一般的な阻害モード(非競争的阻害)は、基質が高濃度に存在する場合でも逆転することはできません。
酵素活性の速度は、ミカリス・メンテン定数(Km)および最大速度(Vmax)によって表され、これはミカリス・メンテングラフをプロットすることによって決定することができ、反応23の速度に対する基質濃度を有する。Km は、酵素活性部位の半分を占めるために必要な基質の濃度であり、高いKm がより少ない親和性を表すことを意味する。Vmax は、反応の最大速度を表し、すべての活性部位が基質によって占有される場合に到達する。Km は Vの最大値の半分に等しい。阻害には、競争、競争力のない、競争力のない3つの最も一般的なモードがあります。競合阻害の場合、阻害剤は酵素の活性部位に結合し、基質と競合する。したがって、Vmax は阻害剤の添加後に変化しないが、Km は増加し、阻害に対抗するためにより多くの基質が必要となる。非競争的阻害は、基質が酵素と複合体を形成する場合にのみ起こる。この場合、阻害のレベルが基質および酵素濃度に依存するように、阻害剤が反応に添加されるとVmax およびKm が減少する。阻害の最後のモードは非競合性であり、他の2つの阻害パターンのミックスです。阻害剤は、酵素がその基質に結合しているかどうかにかかわらず、酵素の活性部位に結合することができる。ここでVmax は阻害剤の添加後に減少するが、Km は24を変えない。
GST活性を測定した分光光測定アッセイは、1974年に1-クロロ−2,4-ジニトロベンゼン(CDNB)を反応22の基質として用いてHabigらによって初めて開発された。GSHとCDNBの間の結合は、340nmの波長で最大の光吸光度を示すGS-DNBを形成し、分光光度計で記録可能である。以下に説明する技術のほとんどは、最良の設定に関する情報および阻害アッセイのための重要な最適化点を含むHabigららから導き出される。この技術は、計算予測を用いた合理的な薬物選択によって選択されたか、文献レビューによって選択されたかにかかわらず、潜在的なGST阻害剤のスクリーニングに適用することができる。新たに合成されたGSTタンパク質または特異的なアイソフォームにプロトコルを適応させる方法も議論される。例えば、臨床的に関連する多型または一塩基多型(SNPs)を示すGSTアイソフォームの阻害力を試験することは、患者固有のGSTを標的とするこのプロトコルの潜在的な用途であり得る。
このプロトコルは、他の機能研究の前にインビトロで潜在的なGST阻害剤のスクリーニングのための迅速かつ実現可能かつ効果的な方法を提供する。酵素阻害剤の最も一般的に測定された特性を評価するために必要なステップは、酵素活性を半分に減少させるために必要な阻害剤の濃度である阻害濃度50(IC50)を説明します。阻害の定数(Ki)は、阻害剤と酵素との解離の平衡定数を表し、これら2つの分子間の親和性の特徴である。これらの2つの値は、非線形回帰と、阻害の各モードに固有の式を使用してそれぞれ測定されます。我々はまた、この阻害パターンの評価を示し、ミカレリス・メンテンプロットを用いて、阻害剤23、25、26,25,の添加後のVmaxおよびKmの変化を決定する。
プロトコル
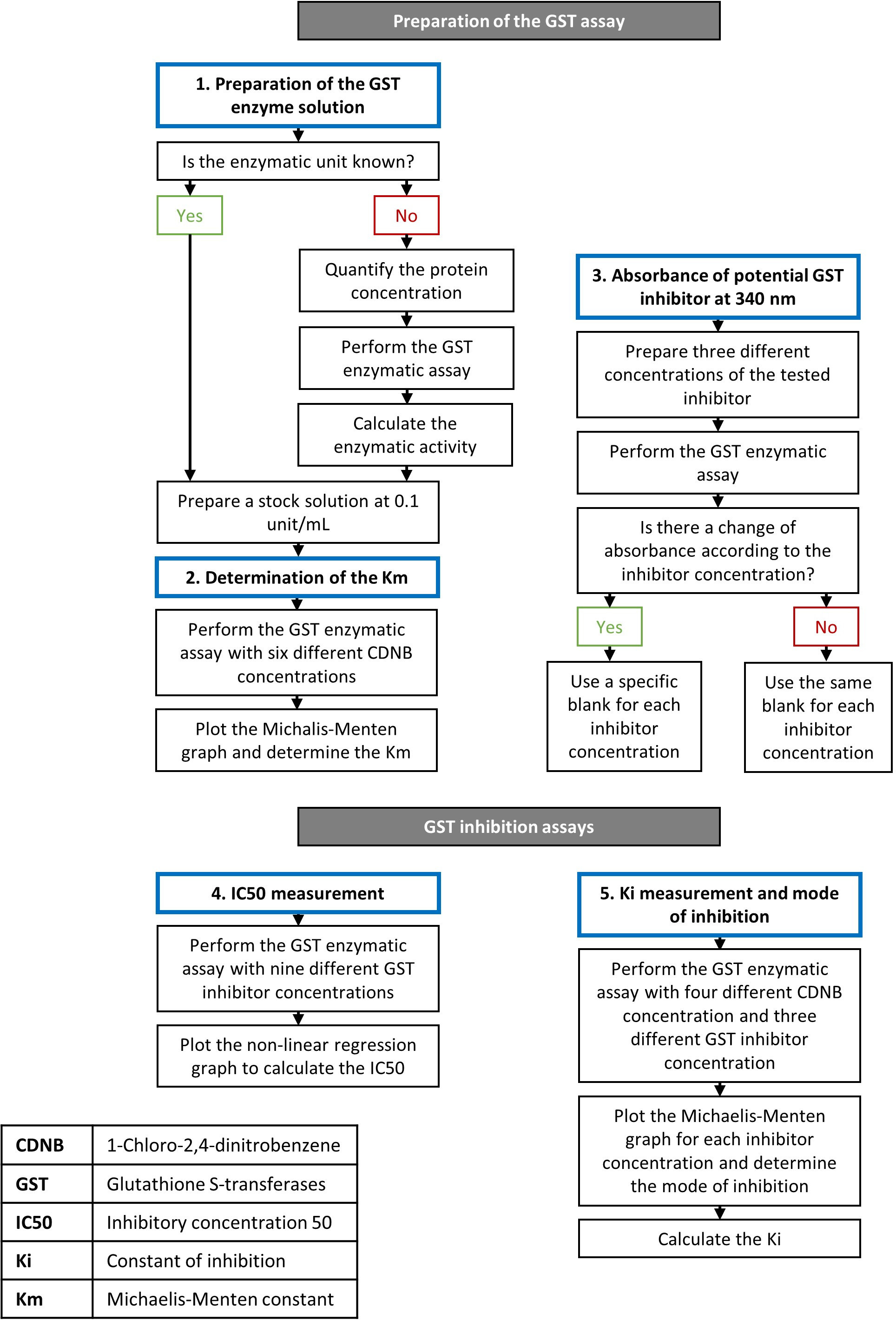
1. GST酵素溶液の調製
注:酵素溶液を調製する手順は、酵素活性の単位がアッセイ前に既知であるかどうかによって異なります。酵素単位の1つは、1分間に1μmolの生成物を合成するのに必要な酵素の量です。酵素活性は単位/mLまたはμmol/min/mLで表され、酵素溶液の希釈に依存する。特定の酵素活性は単位/mgまたはμmol/min/mgで表され、溶液の純度のみに依存する。これらの特性はともに以下に定める。単離されたGSTアイソフォームの酵素単位が不明である場合、各反応の酵素濃度を調整し、再現可能な結果を提供するために推定されなければならない。
- アッセイで使用されるGSTの酵素単位が既知の場合:
- 水に0.1 U/mLでGST酵素の新鮮なストック溶液を準備し、ステップ2に進みます。
注:このソリューションは、数ヶ月間アリコートで-20°Cで保存することも、より長い期間-80°Cで保存することもできますが、凍結/解凍サイクルは避ける必要があります。
- 水に0.1 U/mLでGST酵素の新鮮なストック溶液を準備し、ステップ2に進みます。
- アッセイで使用されるGSTの酵素単位が不明である場合:
- ビチンコニン酸(BCA)タンパク質アッセイ、または他のキットを使用して酵素溶液のタンパク質濃度を定量化します。
- タンパク質溶液を0.02mg/mLの最終的なタンパク質濃度に希釈します。
- 酵素溶液20 μL、GSH 25 mMの20 μL(分子量:307.32 g/mol)、およびDulbeccoのリン酸緩衝生理食塩水(DPBS)150 μLを96ウェルプレートに加えます。ブランクの場合は、酵素溶液の代わりに20 μLの水を加えます。
- 各ウェルにCDNB 50 mM(分子量:202.55 g/mol)の基質を10 μL加えます。
- 分光光度マイクロプレートリーダーで、ウェルを340 nmで読み取るためのパラメータを設定します。1分毎に10分間吸光度を測定することをお勧めします。
- プレートをマイクロプレートリーダーに挿入し、ステップ1.2.5の設定に従って読み取りを開始します。
- 酵素サンプルとブランクの 1 分あたりの吸光度の変化を計算します。
注: Y 軸に吸光度を、x 軸に分をプロットして、反応が線形であることを確認します。反応が線形でない場合、そして高原に達すると、すべての基質が使用され、反応が速すぎることを意味します。これにより、ストック液を2つ希釈してウェルに添加する酵素の量を減らします。 - テストサンプルの吸光度測定値をブランク補正します。
- ビール-ランバート則を表す方程式1では、毎分反応によって形成されたGS-DNBの濃度(μM)を計算します。
方程式1:
ここでCはμMの基質の濃度、A 340/min はステップ1.2.7で測定された1分間の吸光度の変化であり、εは340 nm(0.0096 μM-1*cm-1)のCDNBコンジュゲートのモル吸光係数であり、lはウェル内の光路の長さ(cm単位)である。酵素溶液200 μLで充填された96ウェルプレートの場合、パスの長さは約0.55cmです。この値はプレートモデルによって異なる場合があり、製造元に確認する必要があります。 - μmol/min の 1 ウェルに存在する製品の量を求めるには、式 1 を使用して見つかった結果に解の容積 2 x 10-4 L を掛けます。
- ステップ1.2.9の結果を4 x10-4 mgで割ることによって使用されるタンパク質の量あたりの活性を正規化する。その結果、単位/mgまたはμmol/min/mgの特定の酵素活性が得られます。
注:ステップ1.2.6で希釈が変更された場合は、非線形反応のために、それに応じてタンパク質量を調整してください。 - 酵素活性を見つけるには、ステップ1.2.10で見つかった特定の酵素活性に0.002 mg/mLを掛けることで、mg/mLのタンパク質濃度に結果を調整します。これにより、単位/mLまたはμmol/min/mLの酵素活性が与える。
注:酵素活性とは対照的に、この測定はタンパク質溶液の希釈によって変わりません。ステップ1.2.10と同じノートが、タンパク質濃度のため。 - 水で0.1単位/mLでGST酵素のストック溶液を調製し、ステップ2に進みます。
注:このソリューションは、数ヶ月間アリコートで-20°Cで保存することも、より長い期間-80°Cで保存することもできますが、凍結/解凍サイクルは避ける必要があります。タンパク質溶液の分解が起こりうる長期間にわたって実験を行う場合は、酵素活性の制御が推奨されます。
2. CDNBのGSTアイソフォームのミカエル-メンテン定数の測定
注: 手順は CDNB 基板について説明しますが、GSH などの他の基板に適用できます。CDNBの各濃度は、340 nmでの吸光度がCDNB濃度に従って増加するので、独自のブランクを必要とします。
- エタノール95%(v/v)で、10 mMから100 mMまでの6種類のCDNB濃度を調製します。
- 10 μLのCDNB、20 μLのGST酵素、20 μLのGSH 25 mM、および150 μL DPBSを使用して、アッセイ液を調製します。ブランクの場合は、CDNB溶液の代わりに、エタノールを95%だけ10 μLだけ加えます。
- CDNB濃度10μL、水20μL、GSH 25mM20μL、および150 μL DPBSを使用して、各CDNB濃度にブランクを用意します。
- マイクロプレートリーダーで1分毎に340 nmの吸光度を10分間記録します。
- ブランク補正は、正しい他の試験井戸の結果からブランクから結果を差し引くことによって吸光度を補正します。測定値に応じて、式2を用いて反応の速度を計算する。
方程式2:
ここで、A340/分は1分間に実験的に決定された吸光度の変化であり、V総量(総体積)は0.2mL、V酵素(酵素の体積)は0.02mL、GS-DNB εは340 nm(9.6 mM-1GS-DNB *cm-1)のGS-DNBコンジュゲートのモル消光係数である。96ウェルプレートの200 μLウェルでは、パスの長さは0.55cm(プレートタイプによって異なります)、絶滅係数は5.3 mM-1です。速度はμmol/mL/minまたはmM/minのいずれかによって表すことができる。 - (x軸上)の基板濃度に対する速度(y軸上)でミカエル-メンテングラフをプロットします。
- 反応の最大速度(Vmax)とミカリス-メンテン定数(Km)(すなわち、Vmaxの半分の基質濃度)を定義する。
注:GraphPad Prism などのソフトウェアを使用すると、Vmax や Kmなどのミカリス-メンテン酵素キネティクス パラメータの計算に非線形回帰を使用して酵素の動態曲線を適合させることができます。 - エタノール95%(v/v)で計算されたKm の20倍でCDNBストック溶液を調製します。
3. GST阻害剤の吸光度
注:このステップは、反応に使用される潜在的なGST阻害剤が、測定される波長で吸光度を増加させる代謝産物を生成するかどうかを調べる。その場合、使用される阻害剤の量は結果に影響を与え、濃度ごとに特定のブランクを準備する必要があります。
- 必要な濃度にインヒビターを希釈します。
注:阻害アッセイ中にテストされる最低、中、および最も高い濃度の3つの異なる希釈を準備してください。ウェル内の DMSO 濃度の最大値は、1%以下 (v/v) にする必要があります。検査に対するDMSO濃度の試験は、当研究室での1%がGST活性を大きく変化させなかった。 - 96ウェルプレートに、潜在的なGST阻害剤の2μL、GSH 25 mMの20 μL、DPBSの168 μLを加えます。阻害剤サンプルに使用する溶媒の等量を添加することにより、阻害剤を含む適切なコントロールを使用してください。
- 反応を10分間インキュベートし、酵素反応を開始する。
注:このステップは、異なるインキュベーション時間をテストすることによって、基質の完全な枯渇を避けながら反応を開始するという双子の目的で最適化することができます。 - ステップ2で見つかったKm で最終濃度を達成するために、各ウェルにCDNBの10 μLを加えます。
- プレートを数秒間振ります。
- マイクロプレートリーダーで1分毎に340 nmの吸光度を10分間記録します。
- 1分あたりの吸光度の変化を計算します。
- 阻害薬を使用しないブランク反応と陰性対照反応の両方と比較して吸光度の変化を確認します。
- 大きな変化がある場合は、結果を調整するために各阻害剤濃度にブランクを使用してください。
注:この結果は、反応の成分がGST酵素を含み、自発的に反応し、340nmで吸光度を増加させる代謝産物を生成することを示しています。この追加の吸光度を補正するには、濃度ごとに特定のブランクを測定する必要があります。 - 吸光度に大きな変化がない場合は、全ての濃度に対してGST阻害剤希釈に用いる溶媒のみを含有する一般的なブランクを使用する。
- 大きな変化がある場合は、結果を調整するために各阻害剤濃度にブランクを使用してください。
4. GSTおよびIC50評価の阻害アッセイ
- 潜在的なGST阻害剤の9つの濃度を準備する。
注:濃度は結果に応じて適応させることができます。目的は、非線形回帰曲線の底面と上面の台地を定義することです。この手順の詳細については、ディスカッションのセクションを参照してください。 - GSH溶液20 μLを25 mMのDPBS 148 μLで希釈して、アッセイ液を調製します。アッセイで使用されるウェルの数に応じてボリュームを調整します。
- 透明な96ウェルプレートで、酵素反応を最終体積190μLで準備します。これらの手順では、マルチチャンネルピペットを使用することをお勧めします。
- 試験井戸には、酵素溶液20μL、2μLの潜在的なGST阻害剤溶液、および168μLのアッセイ溶液を加えます。
- コントロールには、酵素溶液20μL、GST阻害剤に使用される希釈剤2μL、アッセイ溶液の168μLを加えます。
- ブランクウェルの場合、水20μL、潜在的なGST阻害剤2μL、アッセイ溶液168μLを加えます。
注:GST阻害剤が340nm波長を吸収する場合、テストされた濃度ごとに特定のブランクを用意する必要があります。
- CDNB溶液10μLを20xKmの各mウェル(ブランクを含む)に加えます。このステップではマルチチャンネルピペットを使用することをお勧めします。
- プレートを数秒間振ります。
- マイクロプレートリーダーを使用して10分間、毎分340 nmで吸光度を測定します。
- ブランクは、試験井戸ブランクから結果を差し引くことによって吸光度を補正します。
- GST阻害剤溶液を用いて得られた値を、阻害剤なしで1分間のコントロールの吸光度で除いて結果を正規化します。
- GST活性に対する阻害剤の対数濃度(x軸)の非線形回帰グラフをプロットし、従ってIC50を決定する。
注: GraphPad Prism は、コントロール内の酵素活性の 50% を示す濃度を予測することにより、非線形回帰プロットから IC50 を計算します。予測は底面と上の台地、ならびにシグモイドグラフによって形成された斜面の曲線に基づいている。
5.Kiの評価と阻害の種類
- 4つの異なるCDNB濃度を準備する:3より高く、1つは以前に見つかったKm に等しい。
- 3つの異なるGST阻害剤濃度を調製し、以前に見つかったIC50以下にする。
- ステップ4.2〜4.7に記載のとおりに阻害アッセイを行う。
- 方程式 2を使用して反応の速度を計算します。
- 各阻害剤濃度のミカレリス-メンテングラフをプロットし、各反応のV最大 およびKm を計算します。
- 異なる濃度を使用する場合のVmax およびKm の変化に応じて、GST阻害剤の阻害様式を評価する。
注: この手順については、結果とディスカッションのセクションで詳しく説明します。 - 阻害のモードに基づいて、Kiを計算します。
注:グラフパッドプリズムでは、阻害の性質に応じてKi を計算するために異なる方程式が使用されます。使用される方程式は、阻害剤を添加した後に観測されたKm およびVmax に基づいており、コントロールのKm およびVmaxと比較される。
結果
馬の肝臓からのGSTを伴うCDNBのミカリス-メンテン定数(Km)
クルクミンは、抗癌特性を示した高用量27で摂取した後でも安全な天然化合物である 16.この分子の阻害力はヒト組換えGSTs19,20,20に実証されている。記載されたプロトコルを用いて、馬の肝臓からのGSTアイソフォームのプールを用いて、GST活性に対するクルクミンの効果を評価した。サプライヤーによると、タンパク質のこの組み合わせの比活性は25 U / mgです。適量の水に凍結乾燥粉末を溶解することにより、0.1 U/mLのストック溶液を調製した。この化合物は、研究で最も広く使用されているGST阻害剤であるとして、エタクリン酸は、陽性対照として並行して使用されました.GST活性アッセイの設定と阻害剤の試験のステップを図1に示します。
Kmと同等の基質濃度を使用すると阻害モード28の判定に関する偏りが保証されないため、Kmは各酵素基質反応に対して定義されなければならない。m0.5 mMから5 mMまでの6つの異なる濃度のCDNBを、2.5 mM GSHの固定濃度を用いてこのパラメータを測定するために試験した。
反応の最終容積は96ウェルプレートで200 μLであった。CDNBとGSHの自発的な結合が起こり、吸光度を増加させる可能性があるため、テスト井戸と同じ濃度のCDNBを使用して、各実験に特定のブランクを使用しました。最終酵素濃度0.01単位/mLを、20 μLのストック溶液をアッセイミックスに添加することで、すべての反応に使用しました。340nmでの吸光度は10分間毎分記録した。反応の速度(mM/分)は、式2を用いて計算した。ミカリス-メンテングラフは、速度(μM/分)に対する基質濃度(mM)をプロットし、Figure 2反応のKmを計算した。実験は、少なくとも3組のデータセットが同様の結果を示すまで繰り返された。馬肝からのGSTを有するCDNBのKmは0.26±0.08mMとして測定した。このGSTのバッチを用いた各阻害アッセイに対して0.2mMのCDNB濃度を用いた。
GSHおよびCDNBと単独でインキュベートされたクルクミンを用いた実験の間に340nmで吸収された代謝産物の自発的な形成は測定されなかった。各実験において全ての阻害剤濃度に同じブランクを使用した。
GSTに対するクルクミンの阻害性
クルクミンの阻害力は、ソフトウェア とのインシリコ ドッキングシミュレーション(例えば、AutoDock Vinaバージョン1.1.2)を使用して予測された。29 異なるGSTアイソフォーム(すなわちGSTA1、GSTM1およびGSTP1)とクルクミン間の自由結合エネルギーが予測された(データは示されていない)。そして、阻害Ki の定数を 式3を用いて計算した。
方程式3:
ここで∆Gは インシリコ 分析を用いて見つかった自由結合エネルギーであり、Rはガス定数1.987 cal*K-1*mol-1であり、Tは実験中の温度であり、この場合ケルビン(298 K)では。
AutoDock Vinaによって返された自由結合エネルギーの結果に基づいて、クルクミンは、それぞれ78.1 μM、78.1 μMおよび27.4 μMのKi 値を有するヒトGSTA1、GSTM1およびGSTP1の強力なGST阻害剤である。第1の阻害アッセイは、これらの計算Ki 推定を用いて実施し、必要に応じてその結果に応じて濃度を調整した。
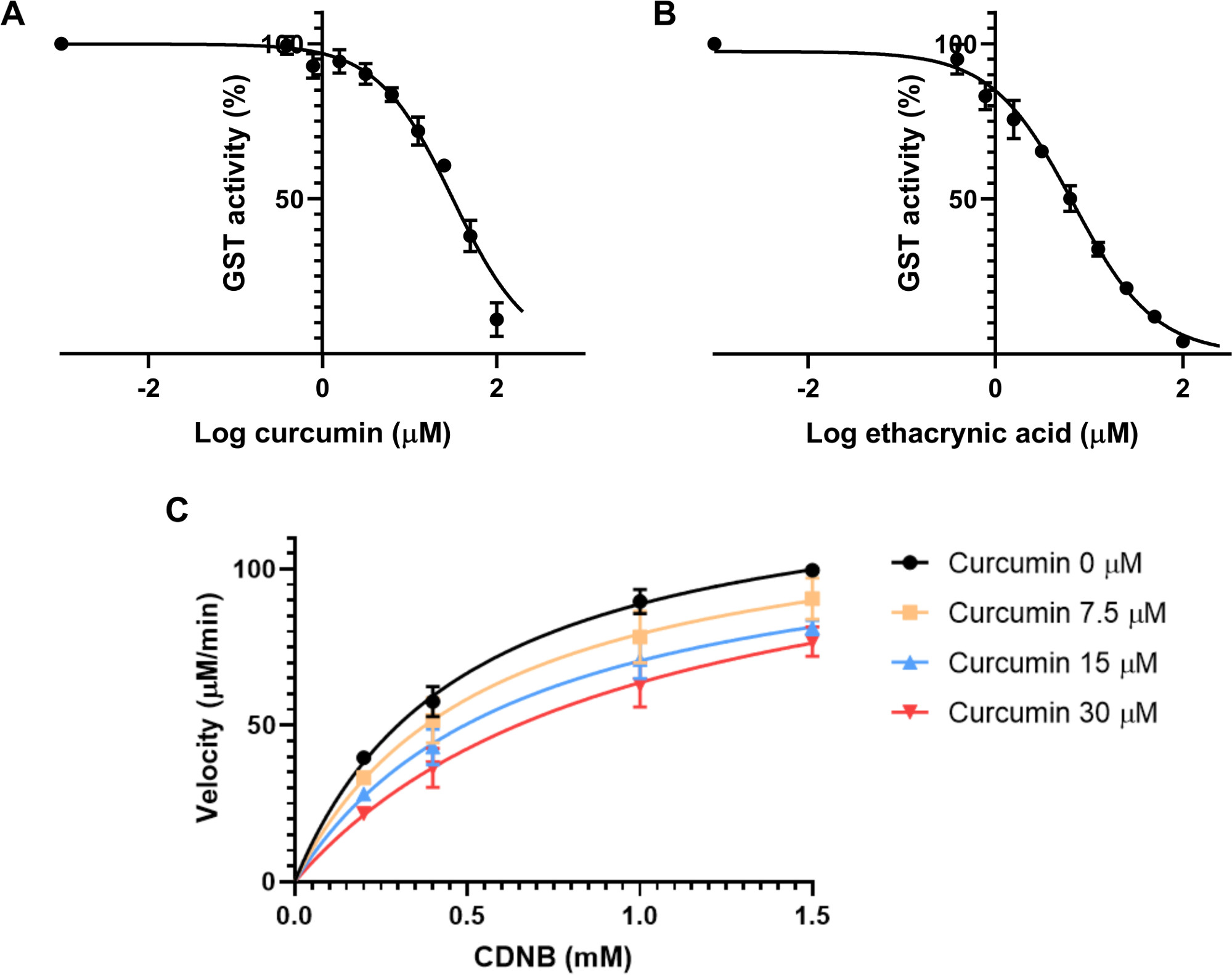
まず、IC50を推定した。この特徴は酵素の反応速度を50%低下させる阻害剤の濃度である。したがって、酵素濃度30に依存する。阻害剤の9つの最終濃度は、0.39 μMから100 μMの範囲で、各濃度は前の濃度の2倍に調製されました。アッセイ溶液は、GSH 2.5 mMの20 μL、0.01 U/mLの最終で馬の肝臓からのGSTの20 μL、クルクミン溶液の2 μL、PBSの148 μLで構成された。コントロールは、クルクミンに使用される希釈剤と同じ溶液で構成された。この混合物を酵素アッセイを開始し、アッセイ液中のクルクミンの分解を避けるために15分間インキュベートした。次に、4 mM で 10 μL の CDNB 溶液を各ウェルに添加し、最終濃度は 0.2 mM CDNB です。340nmでの吸光度を毎分10分間記録し、1分あたりの吸光度の変化を算出した。IC50を算出するために、クルクミンでインキュベートされた各サンプルをコントロールに正規化し、活性の割合を与えた。これらの結果は、阻害に対する阻害剤の対対対対対対対対対対対対の対数的な回帰を計算することによって、プロットソフトウェアを用いて分析した。馬の肝臓からGSTのクルクミンに対して検出されたIC50は31.6±3.6μMであった(図3A)。
同じ実験は、クルクミンと同様に0.39〜100μMの濃度で、エタクリン酸を陽性対照として並行して行った。IC50は6.6±1.1 μMである(図3B)。
次のステップは、これらのトランスファージの抑制のクルクミンのモードを評価することであった.4つの異なる濃度のクルクミンがテストされました:0、7.5、15、および30 μM、および4つの異なる濃度のCDNB:0.2、0.4、1、および1.5mM。プロトコルは、前の実験と同じでした。各条件の速度は、式 2を使用して計算されます。各インヒビター濃度について、基質濃度(mM)に対する速度(μM/分)を用いて曲線をプロットした(図3C)。VmaxおよびKmは、グラフパッドプリズムを使用した各プロットに基づいて決定された。潜在的阻害剤の阻害様式は、これら2つのパラメータの変化とアッセイの異なる条件に従って評価した。24 Vmaxは条件間で有意に変化しなかったが、Kmは異なる阻害剤濃度で増加したので、クルクミンのGSTの阻害は競争力がある。阻害のこのパターンは、阻害剤が酵素の活性部位の基質と競合するときに起こる。競合性阻害剤のKiを測定するために、GraphPadはコープランドら32によって記述された式4および方程式5を使用する。
方程式4:
方程式5:
Km obs は観察されたミカリス-メンテン定数、Km はコントロールのミカリス-メンテン定数、Iは抑制の一定値、Yは速度 、X は基質濃度です。
計算は阻害のモードの評価と同じ実験に基づいている。KIは 、クルクミンの馬肝からのGST阻害について推定され、23.2±3.2μMであった。

図1:GST酵素および阻害アッセイのステップを説明するフローチャート。手順の詳細については、プロトコルのセクションで説明します。 この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図2 GST酵素活性のミカエル-メンテンプロット基質CDNBの濃度の増加は、馬の肝臓からGSTのプールからGST活性を決定するためにGSH(2.5mM)の固定濃度で使用した。CDNBのKm 値は、グラフの曲線に従って0.08mM±0.26と判定した。プロット上の各データポイントは、SDを使用した4つの異なる実験実行 の平均を表します。
{kind=link}

図3: クルクミンとエタクリン酸がGST酵素活性に及ぼす影響(A-B)クルクミン(A)または正対照エタクリン酸(B)のいずれかの濃度の増加を加え、同じGST(0.01 U/mL)、GSH(2.5mM)およびCDNB(0.2mM)の最終濃度を維持しながら、非回帰性の馬肝からGSTの両方の化合物のIC50を計算するために使用した。IC50はクルクミンの31.6±3.6 μM、エタクリン酸の場合は6.6±1.1 μMと判定された。(C) ミカニス-メンテンプロットは、異なる濃度のクルクミンで、競争的な阻害様式を示すCDNB基質の様々な濃度に対して試験した。阻害剤を含み、VmaxおよびKmはそれぞれ132.7±4.6 μM、0.5±0.08 μMと測定しました。30 μMのクルクミン、Vmax、K mは130.4±13.1 μM、1.08±0.39 μMです。 A-C
{kind=link}
ディスカッション
分光光度GST酵素アッセイの各工程を記述するプロトコルを提供し、推定阻害剤(図1、表1)をスクリーニングし、その阻害力を定量化する。我々はまた、再現性のある結果を提供する正確な酵素アッセイのために考慮すべき最も重要な基準を強調した。他の着色法または質量分析に対する記述されたプロトコルの主な利点は、このプロトコルが迅速かつ容易に実行し、GST活性の定量的測定およびスクリーニングされた分子の阻害力を提供することです。
我々は、酵素阻害剤の2つの最も重要なミカリス-メンテンのパラメータを計算する方法を提示する:IC50とKi.強力な阻害剤は、可能な限り低いKiおよびIC50を発揮し、阻害剤と酵素との親和性が高いことを示す30。IC50は酵素濃度とアッセイ条件33に依存するので、異なる実験または他のアッセイ条件34を用いて得られた異なる実験からの阻害剤を比較するためにこの値を使用することが困難である場合がある。阻害の定数を使用してKiは潜在的な化合物の阻害力のより良い指標です。Kiは、阻害剤と酵素の親和性のみに依存するため、異なる阻害モードを有する2つの阻害剤を比較するために使用することができる。しかし、阻害の性質を明確に把握するには、阻害剤のパラメータ30の両方を決定しなければならない。クルクミンのIC50とKiをそれぞれ31.6±3.6μM、23.2±3.2μMとして測定し、この化合物が強力なGST阻害剤であることを示しました。これらの結果は、異なるヒトGSTアイソフォームおよびクルクミンに対して27.4〜78.1μMの間のKi値を推定したインシリコ予測で確認した。
酵素活性または反応速度と酵素量
上記のように、IC50は酵素濃度に依存しており、そして未知の酵素活性レベルで実験を行うことは誤った結論33に至る可能性がある。阻害活性を低下させる可能性のある他の因子を制御するには、GSTの新しいバッチごとに酵素活性を考慮して測定する必要があります。たとえば、フリーズ/融解サイクルが多すぎるために発生する酵素バッチの劣化は、同じ条件で実験を実行した場合でも活性を低下させ、IC50を低下させる可能性があります。つまり、0.01単位の酵素を使用しても、1単位を使用した場合と同じ結果が得られるわけではありません。酵素を多く使用すると、基質が急速に枯渇する可能性があり、反応は直線的な形状を持たない。このパラメータは、長いインキュベーション時間後に吸光度の変化が見られないため、不正確な結果につながる可能性があります。
Km 値
阻害剤によって示される阻害の種類を評価するための最良の条件を確保するために、基質濃度はミカリス-メンテン定数(Km)以下でなければなりません。Kmは、酵素28上の活性部位の半分を占めるために必要な基質の濃度で表される。例えば、より高い基質濃度は、競合性阻害剤を打ち消し、この種の阻害を評価することは、そのような設定では困難である。したがって、この方法論における重要なステップの1つは、選択された基質(ここではCDNB)に対する酵素のKmの決定である。いくつかの研究では、この値が決定されず、阻害剤によって引き起こされる阻害パターンに関する誤った結論につながり、阻害様式が誤っている場合、Kiは、式が阻害パターン26、28,28に依存するように誤って計算される。異なるGSTアイソフォームがテストされた場合、この値は酵素とリガンドのペアに固有であるため、Km値の新しい評価が必須です。0.26±0.08mMと定義したmKm(0.2mM)よりわずかに低いCDNB濃度を用い、GST上でクルクミン阻害の予測競合モードを正確に決定した。
IC50
IC50を推定する良好なシグモイド曲線を得るためには、底部と上面の両方を見つける必要があります。下台の高原は、最大の阻害活性を提供する阻害剤の濃度を表す。場合によっては、溶解性などの技術的な問題のために、化合物が高濃度であっても酵素を完全に阻害しない場合があります。しかし、GraphPadプリズムのようなツールは、非常に正確に下の高原に収めることができます。上台の高原は、酵素を阻害するのに不十分な阻害剤の濃度で構成されているため、活性は最大です。これらの高原は、両方とも、カーブの傾きを見つけるために、IC50と間の濃度を決定する上で非常に重要であり、その後、IC50はシグモイド曲線35の形状から導出することができる。クルクミンは水に難溶性であり、したがってこのアッセイで使用される最大濃度が制限され、アッセイ溶液中の沈殿を回避する。従って、GST活性を完全に阻害しない濃縮液の少ない溶液が使用された。これは、底台の高原の決定のための問題を提起しました。この問題に対抗するため、非線形回帰グラフに基づいて底値を予測し、31.6±3.6 μMのIC50をクルクミンに提供した(図3A)。エタクリン酸の場合、この化合物はアッセイ溶液に可溶性であり、IC50は6.6±1.1μMで測定したため、底面プラトーの値を予測する必要はありませんでした。
この方法は、ヒト、すなわちGSTA1、GSTM1またはGSTP1において最も発現されたGSTアイソフォームに適用することができる。しかしながら、このプロトコルは、このサブタイプの基質ではないCDNBとして、GSTT1アイソフォームの活性を定量化するのには適していない。36, 37 一方、プロトコルは、この問題に対抗するためにわずかに変更することができます。例えば、GSTT1の基材として1,2-エポキシ-3-(4'-ニトロフェオキシ)プロパン(ENPP)を用い、340nmではなく360nmでコンジュゲートの量を測定します。37
プロトコルのステップは、細胞培養実験におけるGST活性および阻害剤試験を試験するために適応および適用することができる。GST阻害剤の有無にかかわらず処理された細胞に対するGST活性の測定は、この化合物がこのような実験設定で使用できるかどうかを示す。阻害剤が親油性である場合は特に興味深い。例えば, 我々はクルクミンは、このプロトコルを使用して強力なGST阻害剤であることを提示しました.それにもかかわらず、分子は水に難溶性であり、培地中で迅速に分解するので、細胞研究への応用は限られている可能性があります。31 このプロトコルの別の改善は、基質CDNBの非アイソフォーム特異性に関して可能である。セル調査でこのプロトコルを使用すると、GSTアクティビティ全体に関する情報のみが提供されますが、正確なGSTサブタイプのアクティビティに関する情報は提供されません。アイソフォーム特異的な基質を追加したり、特定の組換えGSTアイソフォームを使用することで、アイソフォーム特異的なGST阻害剤を試験することができます。
結論として、我々は、電気化学療法と組み合わせて使用される可能性を有するGST阻害剤を試験するための完全な手順を説明する。我々は、GST酵素及び阻害アッセイの重要なステップを強調し、潜在的な興味深い分子を試験し、定量値を有する阻害剤としての効率を決定する、IC50及びKi.この方法は、任意の推定化合物に適用し、最も発現されたヒトGSTアイソフォーム(GSTA1、GSTM1およびGSTP1)に対して行うことができるか、またはわずかにGST阻害剤を用いて細胞培養研究を行うか、または選択した他の興味深いGSTアイソフォームの活性を測定するように適合する。
| 試薬 | 名前 | アッセイ溶液中の濃度 | 他 |
| 基板 | CDNB | 測定されたKm (mM) | 95%エタノールで希釈。最終的なエタノール濃度は5%≤する必要があります(v/v) |
| 結合基板 | Gsh | 2.5 mM | 水で希釈。 |
| 濃度は溶液を飽和させる必要があります。 | |||
| バッファー | Pbs | - | pH = 7.1 |
| 酵素 | GSTアイソフォームまたは純粋なアイソフォームのプール | 0.01単位/mL、実験的に決定される。 | 水で希釈。 |
| GST阻害剤 | 選択する潜在的な化合物 | IC50:最大阻害する3濃度、最小阻害を最小にする3、及び3の間に。 | DMSOで希釈し、次に水中で、DMSOの最終濃度を1%≤(v/v)にします。 |
| Ki:IC50周辺の3濃度。 | |||
| パラメーター | |||
| 室温(25°C) | |||
| pH = 7.1 | |||
| DMSO ≤ 1% (v/v) | |||
| エタノール≤5%(v/v) | |||
表1:GST阻害アッセイ中に考慮すべき試薬およびパラメータの概要。
開示事項
著者らは開示するものは何もない。
謝辞
この研究はCANSEARCH財団の支援を受けています。著者らは、ローレンス・レスネ氏とヨアン・サルミエント氏が、特に阻害剤でアッセイを標準化しながら複製実験を行う際に、技術的支援を認めたい。デニス・マリノ氏、シモナ・ムラカール博士、ヴィド・ムラカール博士による実りある議論とインプットは大いに認められています。パトリシア・ウエゾ・ディアス・カーティス博士のビデオのナレーションに協力してくれてありがとう。 私たちは、シリコ予測における 彼のインプットのためのムトゥクマール博士に感謝します。また、この原稿の英語証明読み取りで彼の助けを借りてくれたダレン・ハート氏に感謝します。
資料
| Name | Company | Catalog Number | Comments |
| 1-Chloro-2,4-dinitrobenzene (CDNB) | Sigma-Aldrich | 237329 | Substrate used for the GST enzymatic assay |
| Corning UV-Transparent Microplates | Sigma-Aldrich | CLS3635 | Transparent plate to perform the enzymatic assay. When using 200 ul, the pathlength is 0.552 cm for this plate. |
| Curcumin | Sigma-Aldrich | 8511 | Used for the results section, to test the inhibition potency of curcumin |
| Dimethyl sulfoxide (DMSO) | Sigma-Aldrich | D2650 | To prepare the stock of the putative inhibitor |
| DPBS | Sigma-Aldrich | D8537 | Buffer for the enzymatic reaction |
| Ethanol 95% | Fisher scientific | 10542382 | To dilute the CDNB |
| Glutathione S-Transferase from equine liver | Sigma-Aldrich | G6511 | Used for the results section, to test the inhibition potency of curcumin |
| L-glutathione reduced (GSH) | Sigma-Aldrich | G4251 | Co-substrate for the GST enzymatic assay |
| Pierce BCA Protein Assay Kit | ThermoFisher | 23225 | To quantify the amount of protein present in the enzymatic solution |
| Spectramax iD3 | Molecular devices | To do spectrophotometric measurments |
参考文献
- Mannervik, B., Danielson, U. H. Glutathione transferases--structure and catalytic activity. CRC Critical Reviews in Biochemistry. 23 (3), 283-337 (1988).
- Awasthi, Y. C., Sharma, R., Singhal, S. S. Human glutathione S-transferases. The International Journal of Biochemistry. 26 (3), 295-308 (1994).
- Rowe, J. D., Nieves, E., Listowsky, I. Subunit diversity and tissue distribution of human glutathione S-transferases: interpretations based on electrospray ionization-MS and peptide sequence-specific antisera. Biochemical Journal. 325, 481-486 (1997).
- Beckett, G. J., Hayes, J. D. Glutathione S-transferases: biomedical applications. Advances in Clinical Chemistry. 30, 281-380 (1993).
- Oakley, A. Glutathione transferases: a structural perspective. Drug Metabolism Reviews. 43 (2), 138-151 (2011).
- Townsend, D. M., Tew, K. D. The role of glutathione-S-transferase in anti-cancer drug resistance. Oncogene. 22 (47), 7369-7375 (2003).
- Tew, K. D. Glutathione-associated Enzymes in Anticancer Drug Resistance. Cancer Research. 54 (16), 4313-4320 (1994).
- Laborde, E. Glutathione transferases as mediators of signaling pathways involved in cell proliferation and cell death. Cell Death and Differentiation. 17 (9), 1373-1380 (2010).
- Adler, V., et al. Regulation of JNK signaling by GSTp. The EMBO Journal. 18 (5), 1321-1334 (1999).
- Cho, S. G., et al. Glutathione S-Transferase Mu Modulates the Stress-activated Signals by Suppressing Apoptosis Signal-regulating Kinase 1. Journal of Biological Chemistry. 276 (16), 12749-12755 (2001).
- Hayes, J. D., Flanagan, J. U., Jowsey, I. R. Glutathione Transferases. Annual Review of Pharmacology and Toxicology. 45 (1), 51-88 (2005).
- Lo, H. W., Ali-Osman, F. Genetic polymorphism and function of glutathione S-transferases in tumor drug resistance. Current Opinion in Pharmacology. 7 (4), 367-374 (2007).
- Ansari, M., et al. Glutathione S-transferase gene variations influence BU pharmacokinetics and outcome of hematopoietic SCT in pediatric patients. Bone Marrow Transplantation. 48 (7), 939-946 (2013).
- Mahajan, S., Atkins, W. M. The chemistry and biology of inhibitors and pro-drugs targeted to glutathione S-transferases. Cellular and molecular life sciences: CMLS. 62 (11), 1221-1233 (2005).
- Allocati, N., Masulli, M., Ilio, C. D., Federici, L. Glutathione transferases: substrates, inihibitors and pro-drugs in cancer and neurodegenerative diseases. Oncogenesis. 7 (1), 8 (2018).
- Aggarwal, B. B., Kumar, A., Bharti, A. C. Anticancer potential of curcumin: preclinical and clinical studies. Anticancer Research. 23 (1), 363-398 (2003).
- Han, S. S., Chung, S. T., Robertson, D. A., Ranjan, D., Bondada, S. Curcumin causes the growth arrest and apoptosis of B cell lymphoma by downregulation of egr-1, c-myc, bcl-XL, NF-kappa B, and p53. Clinical Immunology. 93 (2), 152-161 (1999).
- Golonko, A., Lewandowska, H., Świsłocka, R., Jasińska, U. T., Priebe, W., Lewandowski, W. Curcumin as tyrosine kinase inhibitor in cancer treatment. European Journal of Medicinal Chemistry. 181, 111512 (2019).
- Awasthi, S., et al. Curcumin-glutathione interactions and the role of human glutathione S-transferase P1-1. Chemico-Biological Interactions. 128 (1), 19-38 (2000).
- Appiah-Opong, R., Commandeur, J. N. M., Istyastono, E., Bogaards, J. J., Vermeulen, N. P. E. Inhibition of human glutathione S-transferases by curcumin and analogues. Xenobiotica; the Fate of Foreign Compounds in Biological Systems. 39 (4), 302-311 (2009).
- Dubey, V., Owusu-Apenten, R. Curcumin Restores Glutathione-S-Transferase Activity for LNCaP Prostate Cancer Cells. Pure and Applied Chemistry. 2, 61-72 (2014).
- Habig, W. H., Pabst, M. J., Jakoby, W. B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. The Journal of Biological Chemistry. 249 (22), 7130-7139 (1974).
- Johnson, K. A., Goody, R. S. The Original Michaelis Constant: Translation of the 1913 Michaelis-Menten Paper. Biochemistry. 50 (39), 8264-8269 (2011).
- Ochs, R. S. Understanding Enzyme Inhibition. Journal of Chemical Education. 77 (11), 1453 (2000).
- Bisswanger, H. Enzyme assays. Perspectives in Science. 1 (1), 41-55 (2014).
- Acker, M. G., Auld, D. S. Considerations for the design and reporting of enzyme assays in high-throughput screening applications. Perspectives in Science. 1 (1), 56-73 (2014).
- Cheng, A. L., et al. . Anticancer Research. 21, 2895-2900 (2001).
- Yang, J., Copeland, R. A., Lai, Z. Defining Balanced Conditions for Inhibitor Screening Assays That Target Bisubstrate Enzymes. Journal of Biomolecular Screening. 14 (2), 111-120 (2009).
- Uppugunduri, C. R. S., Muthukumaran, J., Santos-Silva, T., Ansari, M. Identification of putative substrates and inhibitors for Glutathione S-transferases using computational methods. Zenodo. , (2017).
- Burlingham, B. T., Widlanski, T. S. An Intuitive Look at the Relationship of Ki and IC50: A More General Use for the Dixon Plot. Journal of Chemical Education. 80 (2), 214 (2003).
- Wang, Y. J., et al. Stability of curcumin in buffer solutions and characterization of its degradation products. Journal of Pharmaceutical and Biomedical Analysis. 15 (12), 1867-1876 (1997).
- Copeland, R. A. Evaluation of enzyme inhibitors in drug discovery. A guide for medicinal chemists and pharmacologists. Methods of Biochemical Analysis. 46, 1 (2005).
- Kalliokoski, T., Kramer, C., Vulpetti, A., Gedeck, P. Comparability of Mixed IC50 Data - A Statistical Analysis. PLoS ONE. 8 (4), (2013).
- Brandt, R. B., Laux, J. E., Yates, S. W. Calculation of inhibitor Ki and inhibitor type from the concentration of inhibitor for 50% inhibition for Michaelis-Menten enzymes. Biochemical Medicine and Metabolic Biology. 37 (3), 344-349 (1987).
- Brooks, H. B., et al. Basics of Enzymatic Assays for HTS. Eli Lilly & Company and the National Center for Advancing Translational Sciences. , (2012).
- Arakawa, S., et al. Evaluation of hepatic glutathione transferase Mu 1 and Theta 1 activities in humans and mice using genotype information. Drug Metabolism and Disposition: The Biological Fate of Chemicals. 40 (3), 497-503 (2012).
- Sherratt, P. J., Pulford, D. J., Harrison, D. J., Green, T., Hayes, J. D. Evidence that human class Theta glutathione S-transferase T1-1 can catalyse the activation of dichloromethane, a liver and lung carcinogen in the mouse. Comparison of the tissue distribution of GST T1-1 with that of classes Alpha, Mu and Pi GST in human. The Biochemical Journal. 326 (3), 837-846 (1997).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved