
Method Article
Triagem espectrofotométrica para potenciais inibidores de Glutathione Citosolítica S-Transferases
* Estes autores contribuíram igualmente
Neste Artigo
Resumo
Glutationa S-transferases (GSTs) são enzimas desintoxicação envolvidas no metabolismo de numerosas drogas quimioterápicas. A superexpressão dos GSTs está correlacionada com a resistência à quimioterapia do câncer. Uma maneira de combater este fenótipo é usar inibidores. Este protocolo descreve um método usando um ensaio espectrofotométrico para rastrear potenciais inibidores GST.
Resumo
Glutationas S-transferases (GSTs) são enzimas metabólicas responsáveis pela eliminação de compostos eletrofílicos endógenos ou exógenos por conjugação de glutationa (GSH). Além disso, os GSTs são reguladores de quinases de proteína ativadas por mitogênio (MAPKs) envolvidas em vias apoptóticas. A superexpressão dos GSTs está correlacionada com a diminuição da eficácia terapêutica entre os pacientes submetidos à quimioterapia com agentes eletrofílicos. O uso de inibidores GST pode ser uma solução potencial para reverter essa tendência e aumentar a potência do tratamento. Alcançar esse objetivo requer a descoberta de tais compostos, com um ensaio preciso, rápido e fácil de enzimas. Um protocolo espectrofotométrico utilizando 1-cloro-2,4-dinitrobenzeno (CDNB) como substrato é o método mais empregado na literatura. No entanto, os experimentos de inibição gst já descritos não fornecem um protocolo detalhando cada estágio de um ensaio de inibição ideal, como a medição da constante Michaelis-Menten (Km) para CDNB ou indicação da concentração de enzimas empregadas, parâmetros cruciais para avaliar a potência de inibição de um composto testado. Assim, com este protocolo, descrevemos cada passo de um ensaio otimizado de enzimas GST espectrofotométricas, para selecionar bibliotecas de potenciais inibidores. Explicamos o cálculo tanto da concentração inibitória semi-máxima (IC50) quanto da constante de inibição (Ki)— duas características usadas para medir a potência de um inibidor de enzimas. O método descrito pode ser implementado usando um pool de GSTs extraídos de células ou GSTs humanos recombinantes puros, ou seja, GST alfa 1 (GSTA1), GST mu 1 (GSTM1) ou GST pi 1 (GSTP1). No entanto, este protocolo não pode ser aplicado ao GST theta 1 (GSTT1), uma vez que o CDNB não é um substrato para esta isoforme. Este método foi utilizado para testar a potência de inibição da curcumina usando GSTs do fígado equino. Curcumina é uma molécula que exibe propriedades anticânces e mostrou afinidade com isoforms GST depois em previsões de acoplamento de silico. Demonstramos que a curcumina é um potente inibidor gst competitivo, com ic50 de 31,6 ± 3,6 μM e um Ki de 23,2 ± 3,2 μM. A curcumina tem potencial para ser combinada com medicação quimiofica de quimioterapia para melhorar sua eficácia.
Introdução
Enzimas citosolíticas glutationa S-transferase (GSTs, EC 2.5.1.18) catalisam a conjugação da glutationa (GSH) em vários compostos eletrofílicos, como agentes quimioterápicos, para desintoxicar e eliminá-los facilmente do corpo1. Sete isoformes de GST citosósico foram identificados como alfa, mu, pi, sigma, ômega, theta e zeta. Os GSTs são expressos principalmente no fígado, testículos, pulmões e trato gastrointestinal2. O isoforme GST alfa 1 (GSTA1) é altamente expresso em hepatócitos. O corpo expressa heterogêneamente outros subtipos, incluindo GST pi 1 (GSTP1) predominantemente no cérebro, coração e pulmões, e GST mu 1 (GSTM1) no fígado e testículos3. Embora exista uma disselogia de alta sequência entre isoformes GST, cada uma exibe especificidade de substrato e está implicada no metabolismo de drogas e câncer de diferentes formas, de acordo com sua expressão diferencial4,5.
Compostos eletrofílicos entram no corpo exogenously ou são produzidos endógenamente. Pesticidas, prostaglandinas, cancerígenos e quimioterapia são alguns dos substratos potenciais para reações de conjugação de glutationa6. Por exemplo, qualquer composto reativo deficiente de elétrons formado dentro de uma célula é provável que se torne um substrato eletrofílico. Agentes alquilantes como clororambucil ou melfalana são eliminados como conjugados de GSH catalisados por GSTs, e o aumento dos níveis dessas enzimas foram correlacionados com a resistência a esses compostos6,,7.
Outro papel importante dos GSTs citolíticos é regular a atividade de quinases proteicas ativadas por mitogênio (MAPK) como MAPK8 (também conhecido como c-Jun N-terminal kinase, ou JNK1) e MAP3K5 (também conhecido como quinase que regula o sinal de apoptose 1, ou ASK1)8. Alguns isóformes em sua conformação monomérica se ligarão a essas proteínas e, assim, bloquearão a cascata de fosforilação. Em condições normais, o isoforme GSTP1 sequestrará MAPK8 (o ativador da proteína c-Jun). A combinação do c-Jun com a proteína c-Fos forma o fator de transcrição da proteína ativadora 1 (AP-1), responsável pela transcrição dos genes pró-apoptóticos. Em células estressadas, o complexo formado por GSTP1 e MAPK8 se dissocia, c-Jun é ativado, e os genes que levam à apoptose começam a ser expressos9. Uma maior expressão dessa isoforme GST pode, portanto, bloquear o caminho, levando ao aumento da viabilidade celular, mais proliferação celular e menor sensibilidade celular à quimioterapia. Cenários semelhantes podem ocorrer com paralogs de GSTP1, por exemplo, GSTM1, que interage com MAP3K510.
Os papéis desempenhados pelos GSTs no metabolismo de drogas e no sequestro de MAPKs levaram à hipótese de que uma maior expressão dos GSTs poderia ser um sinal de um mecanismo de resistência tumoral ao tratamento quimioterápico6,,11. Por exemplo, o GSTP1 é superexpresso em inúmeros cânceres e sua presença tem sido correlacionada com um prognóstico ruim e uma incidência aumentada de recaída8. O polimorfismo nesses genes também tem mostrado taxas diferenciadas de exposição e sobrevivência de medicamentos para pacientes que apresentam várias doenças, reforçando a ideia de que essas enzimas são cruciais para mecanismos de resistência a medicamentos. Por exemplo, indivíduos com o genótipo nulo GSTM1 estão associados com menor desembaraço de drogas e melhor sobrevida12,13. Existem vários meios potenciais de combater essa superexpressão, como o uso de análogos GSH, prodrugs que são ativados por conjugação com GSTs, ou inibidores GST diretos14,15.
Todos esses métodos estão atualmente sob investigação, e alguns compostos iniciaram testes clínicos para seu potencial uso entre os pacientes. No entanto, até onde sabemos, não há compostos em uso como inibidores gst em ambientes clínicos15. De fato, a falta de especificidade para certos isoformes ou o esgotamento do GSH em células normais, que podem levar a toxicidades causadas pelo acúmulo de espécies reativas de oxigênio (ROS) nos sistemas de órgãos, são apenas algumas das desvantagens que reduzem o potencial dos inibidores gst14,15. O risco de que esses compostos possam exercer outros efeitos farmacodinâmicos no corpo também está limitando seu uso. O ácido ethacúnico, por exemplo, é o inibidor GST mais estudado em ambientes laboratoriais, mas por ser usado principalmente como um diurético forte, esta propriedade limita seu uso em combinação com outras drogas em ambientes clínicos. Curcumina é outro composto natural com sucesso rastreado como um inibidor GST. Esta molécula é um éter polifenól extraído da espécie Curcuma longa de cúrcuma. Tem mostrado resultados promissores como uma possível opção de tratamento contra o câncer, induzindo a apoptose de vários tipos de linhas de células tumorais16,17. O composto pode regular diversas vias celulares, como a quinase de tirasina18 ou a via GST. Estudos com proteínas puras têm mostrado sua potência de inibição em GSTA1, GSTM1 e GSTP119,20. Entretanto, foram observados resultados conflitantes nas células cancerosas, onde a maior atividade do GST intracelular foi medida quando as células foram tratadas com curcumina21. Assim, é importante investigar a concentração inibitória semi-máxima (IC50) e a constante de inibição (Ki) de qualquer inibidor de GST putativo usando um protocolo claramente descrito com controles adequados antes de planejar quaisquer outros experimentos celulares.
A triagem e o teste de potenciais novos inibidores GST é, portanto, de interesse clínico significativo, e qualquer novo composto encontrado deve ser seguro e eficiente para uso em combinação com drogas eletrofílicas. O foco de pesquisas em inibidores específicos de isoforme pode permitir a inibição do GST em tecidos tumorais que exibem padrões específicos de expressão GST, permitindo assim o desenvolvimento de uma terapia de combinação eficaz. Encontrar inibidores com modos diferenciais de inibição também pode ser de interesse. Por exemplo, um inibidor competitivo que usa GSH como substrato pode induzir seu esgotamento. Essa redução da concentração de GSH nas células mostrou-se induzir o estresse oxidativo nos neurônios, levando à apoptose. Outro modo comum de inibição — inibição não competitiva — não pode ser revertido mesmo que o substrato esteja presente em altas concentrações.
A taxa de atividade enzimática é representada pela constante Michaelis-Menten (Km) e a velocidade máxima (Vmax),que pode ser determinada plotando um gráfico Michaelis-Menten, com a concentração do substrato contra a velocidade da reação23. Km é a concentração de substrato necessário para ocupar metade dos locais ativos enzimáticos, o que significa que um alto Km representa menos afinidade. Vmax representa a velocidade máxima da reação, alcançada quando todos os locais ativos são ocupados pelo substrato. Km é igual a metade Vno máximo. Existem três modos mais comuns de inibição: competitivo, não competitivo e não competitivo. Em caso de inibição competitiva, o inibidor se liga ao local ativo da enzima e compete com o substrato. Assim, Vmax não muda após a adição do inibidor, mas Km aumenta, pois mais substrato é necessário para combater a inibição. A inibição não competitiva ocorre apenas quando o substrato forma um complexo com a enzima. Neste caso, como o nível de inibição depende da concentração de substrato e enzima, Vmax e Km diminuem quando o inibidor é adicionado à reação. O último modo de inibição não é competitivo e é uma mistura dos dois outros padrões de inibição. O inibidor pode ligar-se ao local ativo da enzima, quer a enzima esteja ligada ao seu substrato ou não. Aqui Vmax diminui após a adição do inibidor, mas Km não muda24.
Um ensaio espectrofotométrico que mediu a atividade GST foi desenvolvido pela primeira vez por Habig et al. em 1974 usando 1-cloro-2,4-dinitrobenzeno (CDNB) como substrato para a reação22. A conjugação entre GSH e CDNB forma GS-DNB, que exibe uma absorção máxima de luz no comprimento de onda de 340 nm, gravável com um espectrômetro. A maior parte da técnica explicada abaixo é derivada de Habig et al., incluindo informações sobre as melhores configurações e pontos de otimização importantes para um ensaio inibidor. A técnica pode ser aplicada à triagem de potenciais inibidores gst, seja escolhido pela seleção racional de medicamentos usando previsões computacionais ou por revisão da literatura. Como adaptar o protocolo a proteínas GST recém-sintetizadas ou isoforms específicos também é discutido. Por exemplo, testar a potência inibitória dos isoformas GST exibindo um polimorfismo clinicamente relevante ou de polimorfismos de nucleotídeos únicos (SNPs) pode ser uso potencial para este protocolo, visando GSTs específicos do paciente.
Este protocolo fornece um método rápido, viável e eficaz para a triagem de potenciais inibidores GST in vitro antes de qualquer outro estudo funcional. As etapas necessárias para avaliar as características mais comumente medidas de um inibidor enzimático são descritas: a concentração inibitória 50 (IC50) que é a concentração do inibidor necessário para diminuir a atividade enzimática pela metade; e a constante de inibição (Ki) que representa a constante de equilíbrio da dissociação entre o inibidor e a enzima, característica da afinidade entre essas duas moléculas. Esses dois valores são medidos por regressão não linear e utilizando equações específicas para cada modo de inibição, respectivamente. Também demonstramos a avaliação desse padrão de inibição, utilizando tramas de Michaelis-Menten para determinar a alteração de Vmax e Km após a adição do inibidor23,,25,26.
Protocolo
1. Preparação da solução enzimápica GST
NOTA: O procedimento de preparação da solução enzimática depende se as unidades de atividade enzimática são conhecidas ou não antes do ensaio. Uma unidade enzimática é a quantidade de enzima necessária para sintetizar 1 μmol do produto por minuto. A atividade enzimática é representada em unit/mL ou μmol/min/mL e depende da diluição da solução enzimática. A atividade enzimática específica é representada em unidade/mg ou μmol/min/mg e depende unicamente da pureza da solução. Ambas as características são determinadas abaixo. Se a unidade enzimática de uma isóforma GST isolada for desconhecida, deve-se estimar ajustar as concentrações enzimáticas para cada reação e fornecer resultados reprodutíveis.
- Se a unidade enzimática do GST utilizada no ensaio for conhecida:
- Prepare uma solução de estoque fresco da enzima GST a 0,1 U/mL na água e, em seguida, prossiga com o passo 2.
NOTA: Esta solução pode ser armazenada a -20 °C em alíquotas por vários meses ou a -80 °C por períodos mais longos, mas o ciclo de congelamento/degelo deve ser evitado.
- Prepare uma solução de estoque fresco da enzima GST a 0,1 U/mL na água e, em seguida, prossiga com o passo 2.
- Se a unidade enzimática do GST utilizada no ensaio for desconhecida:
- Quantifique a concentração proteica da solução enzimática usando um ensaio proteico ácido bicinchonínico (BCA) ou qualquer outro kit.
- Diluir a solução proteica para uma concentração proteica final de 0,02 mg/mL.
- Adicione 20 μL da solução enzimápica, 20 μL de GSH 25 mM (peso molecular: 307,32 g/mol) e 150 μL da solução salina tamponada de fosfato de Dulbecco (DPBS) a uma placa de 96 poços. Para o espaço em branco, adicione 20 μL de água em vez da solução enzimápica.
- Adicione 10 μL de CDNB 50 mM (peso molecular: 202,55 g/mol) substrato a cada poço.
- Em um leitor de microplacas espectrofotométricas, defina os parâmetros para a leitura dos poços em 340 nm. Recomenda-se medir a absorvância a cada minuto durante 10 minutos.
- Insira a placa no leitor de microplacos e inicie a leitura de acordo com as configurações da etapa 1.2.5.
- Calcule a mudança de absorvância por minuto para as amostras enzimáticas e o em branco.
NOTA: Verifique se a reação é linear plotando a absorvância no eixo y e minutos no eixo x. Se a reação não é linear e atinge um platô, significa que todo o substrato é usado e a reação é muito rápida. Assim, reduza a quantidade de enzima adicionada ao poço diluindo a solução de estoque por dois. - Corrija em branco as leituras de absorvência das amostras de teste.
- Com a equação 1, representando a lei Beer-Lambert, calcule a concentração de GS-DNB formada (em μM) pela reação a cada minuto.
Equação 1:
onde C é a concentração do substrato em μM, Um340/min é a mudança de absorvância por minuto, medida na etapa 1.2.7, ε é o coeficiente de extinção molar para o conjugado CDNB a 340 nm (0,0096 μM-1*cm-1), e l é o comprimento do caminho de luz no poço (em cm). Para uma placa de 96 poços preenchida com 200 μL de solução enzimática, o comprimento do caminho é de cerca de 0,55 cm. Este valor pode variar de acordo com o modelo da placa e deve ser verificado com o fabricante. - Para determinar a quantidade de produto presente em um poço em μmol/min, multiplique os resultados encontrados utilizando a equação 1 pelo volume da solução, 2 x 10-4 L.
- Normalize a atividade por quantidade de proteína utilizada dividindo os resultados da etapa 1.2.9 por 4 x 10-4 mgs. O resultado é a atividade enzimática específica na unidade/mg ou μmol/min/mg.
NOTA: Se a diluição foi alterada na etapa 1.2.6, por causa de uma reação não linear, ajuste a quantidade de proteína em conformidade. - Para encontrar a atividade enzimática, ajuste os resultados à concentração proteica em mg/mL multiplicando a atividade enzimática específica encontrada na etapa 1.2.10 por 0,002 mg/mL. Isso dará a atividade enzimática na unidade/mL ou μmol/min/mL.
NOTA: Em contraste com a atividade enzimática, esta medida não mudará dependendo da diluição da solução proteica. Mesma nota do passo 1.2.10, mas para a concentração de proteínas. - Prepare uma solução de estoque da enzima GST a 0,1 unidade/mL na água e, em seguida, prossiga com a etapa 2.
NOTA: Esta solução pode ser armazenada a -20 °C em alíquotas por vários meses ou a -80 °C por períodos mais longos, mas o ciclo de congelamento/degelo deve ser evitado. Recomenda-se o controle da atividade enzimática se os experimentos forem realizados durante um longo período de tempo, pois a degradação da solução proteica pode ocorrer.
2. Medição da constante Michaelis-Menten do isoform GST para CDNB
NOTA: O procedimento é explicado para um substrato CDNB, mas pode ser aplicado a qualquer outro substrato, como o GSH. Cada concentração de CDNB precisa de seu próprio espaço em branco, pois a absorvância a 340 nm aumentará de acordo com a concentração do CDNB.
- Prepare seis concentrações diferentes de CDNB, variando de 10 mM a 100 mM, em etanol 95% (v/v).
- Prepare a solução de ensaio, com 10 μL de CDNB, 20 μL de enzima GST, 20 μL de GSH 25 mM e 150 μL DPBS. Para o branco, em vez da solução CDNB, adicione apenas 10 μL de etanol 95%.
- Prepare um branco para cada concentração de CDNB, com 10 μL de CDNB, 20 μL de água, 20 μL de GSH 25 mM e 150 μL DPBS.
- Regisso de absorvância a 340 nm a cada minuto durante 10 minutos com um leitor de microplacas.
- Corrija a absorvância subtraindo os resultados do vazio do dos outros poços de teste corretos. De acordo com os valores medidos, calcule a velocidade da reação usando a Equação 2.
Equação 2:
onde A340/min é a mudança experimentalmente determinada na absorvência por minuto, Vtotal (volume total) é igual a 0,2 mL,enzima V (volume de enzima) é de 0,02 mL, e εGS-DNB é o coeficiente de extinção molar do conjugado GS-DNB a 340 nm (9,6 mM-1*cm-1). Em um poço de 200 μL de uma placa de 96 poços, o comprimento do caminho é de 0,55 cm (dependendo do tipo de placa) e o coeficiente de extinção equivale a 5,3 mM-1. A velocidade pode ser representada por μmol/mL/min ou mM/min. - Plote o gráfico Michaelis-Menten com a velocidade (no eixo y) contra a concentração do substrato (no eixo x).
- Defina a velocidade máxima (Vmax) da reação e a constante Michaelis-Menten (Km) (ou seja, a concentração do substrato na metade de Vmax).
NOTA: Utilizando softwares como GraphPad Prism, a curva cinética enzimática pode ser montada usando uma regressão não linear para o cálculo dos parâmetros de cinética enzimática enzimática Michaelis-Menten, como Vmax e Km. - Prepare uma solução de estoque cdnb em 20 vezes o Km calculado em etanol 95% (v/v).
3. Absorvância do potencial inibidor GST
NOTA: Esta etapa é realizada para investigar se o potencial inibidor GST usado na reação produz um metabólito que aumenta a absorção no comprimento de onda que está sendo medido. Se isso acontecer, a quantidade de inibidor utilizado terá um impacto nos resultados e um branco específico deve ser preparado para cada concentração.
- Diluir o inibidor às concentrações necessárias.
NOTA: Prepare três diluições diferentes, de preferência as concentrações mais baixas, médias e altas testadas durante o ensaio de inibição. A concentração máxima de DMSO no poço deve ser igual ou inferior a 1% (v/v). Testamos o efeito da concentração de DMSO no ensaio em nosso laboratório, e 1% não alterou significativamente a atividade do GST. - Em uma placa de 96 poços, adicione 2 μL do potencial inibidor GST, 20 μL de GSH 25 mM e 168 μL de DPBS. Use um controle adequado, sem inibidor, adicionando volumes iguais do solvente utilizado para as amostras inibidoras.
- Incubar a reação por 10 minutos, para começar a reação enzimática.
NOTA: Esta etapa pode ser otimizada, testando diferentes tempos de incubação, com o objetivo gêmeo de iniciar a reação, evitando o esgotamento total do substrato. - Adicione 10 μL de CDNB a cada poço para alcançar uma concentração final no Km encontrado na etapa 2.
- Agite a placa por alguns segundos.
- Regisso de absorvância a 340 nm a cada minuto durante 10 minutos com um leitor de microplacas.
- Calcule a mudança de absorvência por minuto.
- Verifique a mudança na absorvância em comparação com a reação em branco e a reação de controle negativo sem inibidor.
- Se houver uma mudança significativa, use em branco para cada concentração inibidora para ajustar os resultados.
NOTA: Este resultado indica que os constituintes da reação, sem enzima GST, reagem espontaneamente e produzem um metabólito que aumenta a absorção a 340 nm. Para corrigir essa absorvência adicional, deve-se medir um branco específico para cada concentração. - Se não houver alteração significativa na absorção, utilize um branco geral, contendo apenas o solvente utilizado para a diluição inibidora gst, para todas as concentrações.
- Se houver uma mudança significativa, use em branco para cada concentração inibidora para ajustar os resultados.
4. Ensaio de inibição da avaliação gst e IC50
- Prepare nove concentrações do potencial inibidor GST.
NOTA: As concentrações podem ser adaptadas de acordo com os resultados. O objetivo é definir os platôs inferiores e superiores da curva de regressão não linear. Consulte a seção de discussão para obter mais detalhes sobre esta etapa. - Prepare a solução de ensaio diluindo 20 μL da solução GSH a 25 mM com 148 μL de DPBS. Adapte o volume de acordo com o número de poços utilizados no ensaio.
- Em uma placa clara de 96 poços, prepare a reação enzimática em um volume final de 190 μL. Recomenda-se o uso de uma pipeta multicanal para estas etapas.
- Para os poços de teste, adicione 20 μL da solução enzimápica, 2 μL da solução potencial inibidora GST e 168 μL da solução de ensaio.
- Para o controle, adicione 20 μL da solução enzimápica, 2 μL do diluente utilizado para o inibidor GST e 168 μL da solução de ensaio.
- Para os poços em branco, adicione 20 μL de água, 2 μL do potencial inibidor GST e 168 μL da solução de ensaio.
NOTA: Se o inibidor GST absorver o comprimento de onda de 340 nm, então um branco específico deve ser preparado para cada concentração testada.
- Adicione 10 μL da solução CDNB a 20x Km a cada poço, incluindo o em branco. Recomenda-se o uso de uma pipeta multicanal para esta etapa.
- Agite a placa por alguns segundos.
- Meça a absorvância a 340 nm a cada minuto durante 10 minutos usando um leitor de microplacas.
- Corrija a absorvância subtraindo os resultados dos espaços em branco do teste.
- Normalizar os resultados dividindo os valores obtidos utilizando a solução inibidora GST pela absorvância do controle por minuto sem inibidor.
- Plote os gráficos de regressão não linear da concentração logarítmica (x-eixo) do inibidor contra a atividade GST (eixo y), e assim determinar o IC50.
NOTA: O Prisma do GraphPad calcula o IC50 a partir das parcelas de regressão não linear, prevendo a concentração que mostrará 50% da atividade enzimática no controle. A previsão é baseada nos platôs inferiores e superiores, bem como na curva da inclinação formada pelo gráfico sigmoide.
5. Avaliação do Kie do tipo de inibição
- Prepare quatro concentrações diferentes de CDNB: três maiores e uma igual ao Km encontrado anteriormente.
- Prepare três concentrações inibidoras GST diferentes, iguais ou inferiores ao IC50 encontrado anteriormente.
- Realize o ensaio de inibição conforme descrito nas etapas 4.2 a 4.7.
- Calcule a velocidade das reações usando a Equação 2.
- Plote os gráficos Michaelis-Menten para cada concentração inibidora e calcule os Vmax e Km de cada reação.
- De acordo com as mudanças em Vmax e Km ao usar diferentes concentrações, avalie o modo de inibição do inibidor GST.
NOTA: Esta etapa é explicada com mais detalhes nas seções de resultados e discussão. - Com base no modo de inibição, calcule o Ki.
NOTA: No Prisma do GraphPad, diferentes equações serão usadas para calcular o Ki de acordo com a natureza da inibição. As equações utilizadas baseiam-se no Km e Vmax observados após a adição do inibidor e comparados com os Km e Vmaxdo controle .
Resultados
Michaelis-Menten constante (Km) de CDNB com GST de fígado equino
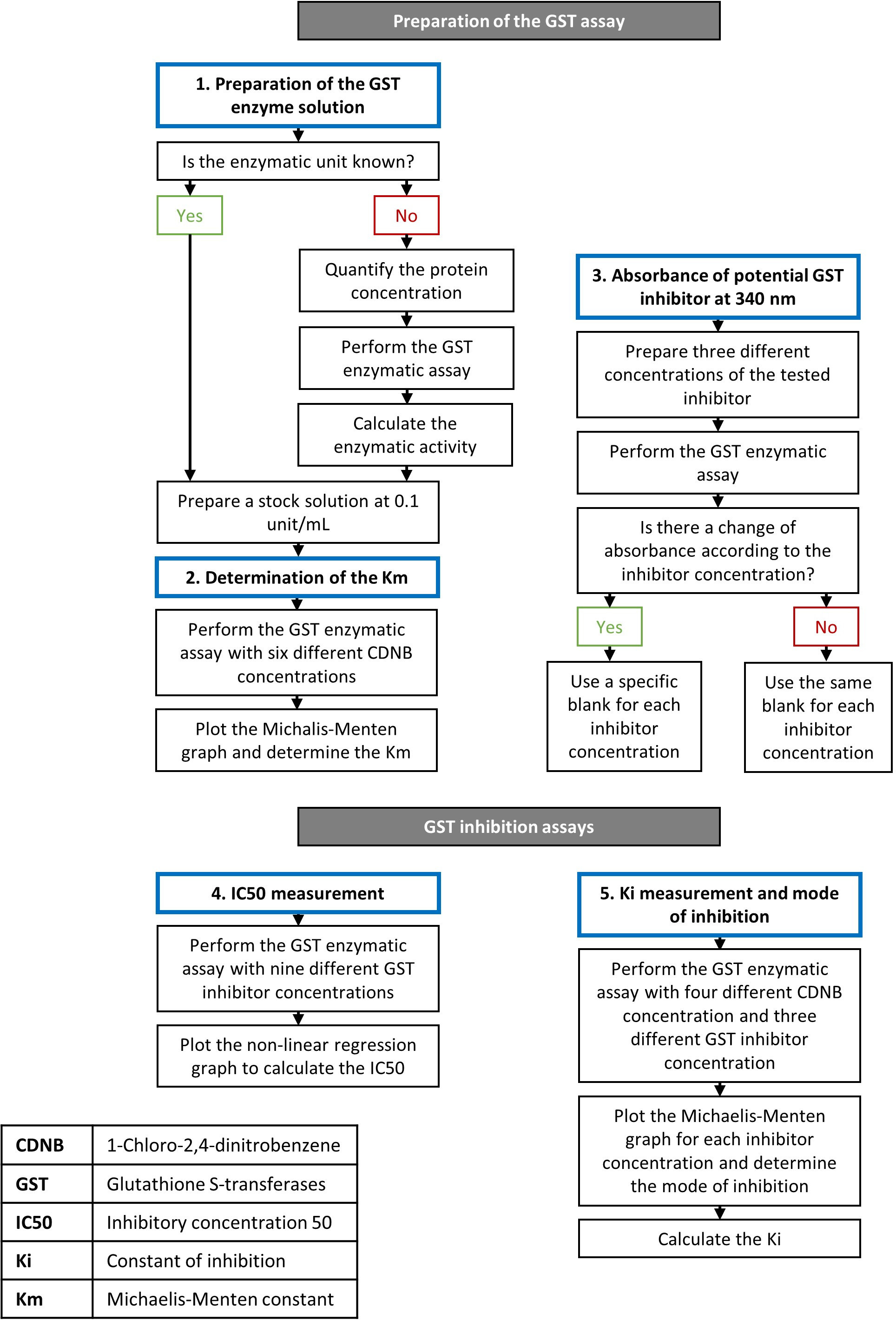
A curcumina é um composto natural seguro mesmo após a ingestão em doses mais altas27 que apresentaram propriedades anticancerígenas16. A potência inibitória desta molécula foi demonstrada em GSTs recombinantes humanas19,20. Usando o protocolo descrito, avaliamos o efeito da curcumina na atividade GST usando um pool de isoformas GST do fígado equino. Segundo o fornecedor, a atividade específica desse mix de proteínas é de 25 U/mg. Uma solução de estoque de 0,1 U/mL foi preparada dissolvendo o pó liofilizado na quantidade adequada de água. O ácido ethacúnico foi usado em paralelo como um controle positivo, pois este composto é o inibidor GST mais utilizado em estudos. As etapas na configuração de um ensaio de atividade GST e testes de inibidores são descritas na Figura 1.
O Km deve ser definido para cada reação substrato enzimática, pois o uso da concentração de substrato equivalente a Km garantiria nenhum viés quanto à determinação do modo de inibição28. Seis concentrações diferentes de CDNB, variando de 0,5 mM a 5 mM, foram testadas para medir este parâmetro, utilizando uma concentração fixa de 2,5 mM GSH.
O volume final da reação foi de 200 μL, em uma placa de 96 poços. Um branco específico foi utilizado para cada experimento, utilizando a mesma concentração de CDNB que o teste bem porque a conjugação espontânea de CDNB e GSH poderia ocorrer e poderia aumentar a absorvância. Foi utilizada uma concentração enzimática final de 0,01 unidade/mL para todas as reações, adicionando 20 μL da solução de estoque ao mix de ensaios. A absorção a 340 nm foi registrada a cada minuto durante 10 minutos. A velocidade (em mM/min) da reação foi calculada utilizando-se a Equação 2. Um gráfico de Michaelis-Menten foi plotado da concentração do substrato (mM) contra a velocidade (μM/min)(Figura 2), e o Km da reação foi calculado. O experimento foi repetido até que pelo menos três conjuntos de dados mostraram resultados semelhantes. O Km de CDNB com GST de fígado equino foi medido como 0,26 ± 0,08 mM. Foi utilizada uma concentração de 0,2 mM de CDNB para cada ensaio inibitório utilizando este lote de GSTs.
Nenhuma formação espontânea de metabólito que absorvia a 340 nm foi medida durante experimentos usando curcumina incubada sozinha com GSH e CDNB. O mesmo espaço em branco foi usado para cada concentração inibidora em cada experimento.
Potência inibitória da curcumina em GSTs
A potência inibitória da curcumina foi prevista usando uma simulação de acoplamento em silico com software (por exemplo, AutoDock Vina versão 1.1.2). 29 A energia de ligação livre entre diferentes isoformas GST (ou seja, GSTA1, GSTM1 e GSTP1) e curcumina foi prevista (dados não mostrados). Em seguida, a constante de inibição Ki foi calculada usando a Equação 3.
Equação 3:
onde ∆G é a energia de ligação livre encontrada usando a análise em silico, R é a constante de gás de 1,987 cal*K-1*mol-1, e T é a temperatura durante o experimento, neste caso em Kelvin (298 K).
Com base nos resultados de energia de ligação livre retornados pelo AutoDock Vina, a curcumina é um potente inibidor GST do humano GSTA1, GSTM1 e GSTP1, com valores Ki de 78,1 μM, 78,1 μM e 27,4 μM, respectivamente. Os primeiros ensaios inibitórios foram realizados utilizando-se essas estimativas computacionais Ki, e as concentrações foram ajustadas de acordo com os resultados, se necessário.
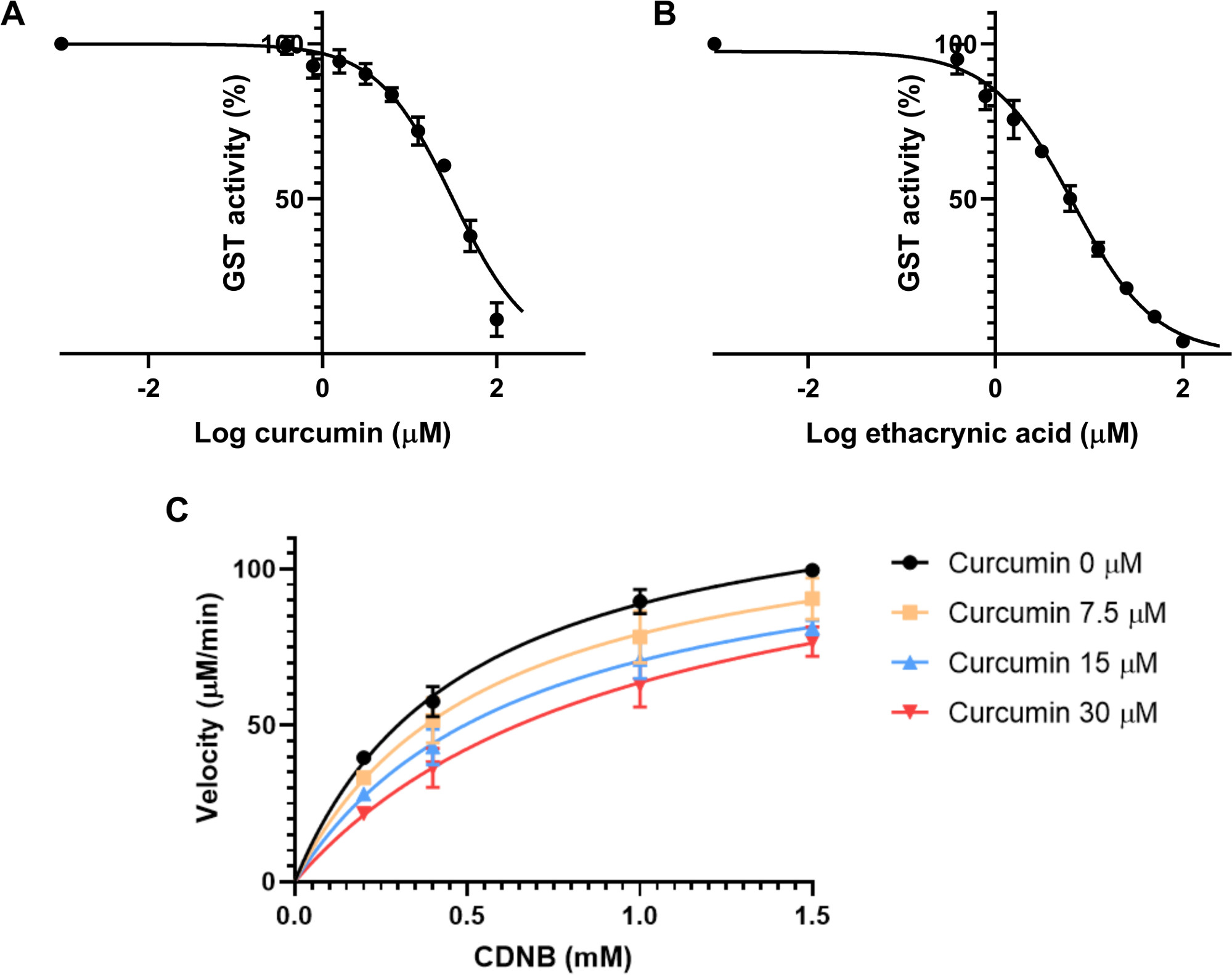
Primeiro, o IC50 foi estimado. Essa característica é a concentração do inibidor que reduz a taxa de reação de uma enzima em 50%. Depende, portanto, da concentração enzimática30. Foram preparadas nove concentrações finais do inibidor, variando de 0,39 μM a 100 μM com cada concentração o dobro da anterior. A solução de ensaio foi composta por 20 μL de GSH 2,5 mM, 20 μL de GST de fígado equino em 0,01 U/mL final, 2 μL da solução de curcumina e 148 μL de PBS. O controle foi composto pela mesma solução com o diluente utilizado para curcumina. Esta mistura foi incubada por 15 minutos para iniciar o ensaio enzimático e evitar a degradação da curcumina na solução de ensaio, uma vez que este composto é instável em soluções tampão31. Em seguida, 10 μL de solução CDNB a 4 mM foi adicionado a cada poço, para uma concentração final de 0,2 mM CDNB. A absorvância a 340 nm foi registrada a cada minuto durante 10 minutos, para calcular mudanças na absorvância por minuto. Para calcular o IC50, cada amostra incubada com curcumina foi normalizada ao controle, para dar a porcentagem de atividade. Esses resultados foram analisados utilizando-se o software de plotagem através do cálculo da regressão não linear da concentração logarítmica do inibidor versus a inibição. O IC50 encontrado para curcumina em GST do fígado equino foi de 31,6 ± 3,6 μM(Figura 3A).
O mesmo experimento foi realizado em paralelo utilizando ácido ethacúnico como controle positivo, com concentrações variando de 0,39 a 100 μM, como com curcumina. O IC50 foi encontrado em 6,6 ± 1,1 μM(Figura 3B).
O passo seguinte foi avaliar o modo de inibição da curcumina nessas transferências. Foram testadas quatro concentrações diferentes de curcumina: 0, 7,5, 15 e 30 μM, além de quatro concentrações diferentes de CDNB: 0,2, 0,4, 1 e 1,5 mM. O protocolo era o mesmo do experimento anterior. A velocidade de cada condição foi calculada usando a Equação 2. Uma curva foi traçada para cada concentração inibidora, com a velocidade (em μM/min) contra a concentração de substrato (mM)(Figura 3C). Os Vmax e Km foram determinados com base em cada parcela com GraphPad Prism. O modo de inibição do potencial inibidor foi avaliado de acordo com as mudanças nesses dois parâmetros e a condição diferente dos ensaios. 24 Como Vmax não mudou significativamente entre as condições, mas Km aumentou com as diferentes concentrações inibidoras, a inibição da curcumina de GSTs é competitiva. Este padrão de inibição ocorre quando o inibidor compete com o substrato para o local ativo da enzima. Para medir o Ki de um inibidor competitivo, o GraphPad usa a Equação 4 e a Equação 5, conforme descrito por Copeland et al.32.
Equação 4:
Equação 5:
onde Km obs é a constante de Michaelis-Menten observada, Km é a constante Michaelis-Menten do controle, [I] é a concentração inibidora, Ki é a constante da inibição, Y é a velocidade, e X é a concentração de substrato.
Os cálculos baseiam-se nos mesmos experimentos que a avaliação do modo de inibição. O Ki estimado para a inibição de curcumina de GST do fígado equino foi de 23,2 ± 3,2 μM.

Figura 1: Fluxograma descrevendo os passos dos ensaios enzimáticos e inibidores do GST. Detalhes sobre o procedimento são apresentados na seção de protocolo. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2: Enredo michaelis-menten da atividade enzimada GST. O aumento da concentração do substrato CDNB foi utilizado com uma concentração fixa de GSH (2,5 mM) para determinar a atividade GST a partir de uma piscina de GST do fígado equino. O valor de Km para CDNB foi determinado como 0,26 ± 0,08 mM de acordo com a curva do gráfico. Cada ponto de dados no plot representa a média de quatro corridas experimentais diferentes com OD. Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3: Efeito da curcumina e ácido ethacúnico na atividade enzimátil GST. (A-B) Além de concentrações crescentes de curcumina (A) ou do controle positivo ácido ethacúnico(B),mantendo as mesmas concentrações finais GST (0,01 U/mL), GSH (2,5 mM) e CDNB (0,2 mM) foram utilizadas para calcular o IC50 de ambos os compostos para GST do fígado equino com regressão não linear. IC50 foi determinado como 31,6 ± 3,6 μM para curcumina e 6,6 ± 1,1 μM para ácido ethacúnico. (C) Enredo michaelis-menten de diferentes concentrações de curcumina, testado contra concentrações variadas de substrato CDNB indicando um modo competitivo de inibição. Sem inibidor, Vmax e Km foram medidos como 132,7 ± 4,6 μM e 0,5 ± 0,08 μM, respectivamente. Com 30 μM de curcumina, Vmax e Km foram 130,4 ± 13,1 μM e 1,08 ± 0,39 μM. Os pontos de dados em todos os gráficos(A-C) representam os meios de três diferentes corridas experimentais com SD. Por favor, clique aqui para ver uma versão maior deste número.
{kind=link}
Discussão
Fornecemos um protocolo descrevendo cada passo de um ensaio enzimático GST espectrofotométrico, para tela de inibidores putativos(Figura 1, Tabela 1) e quantificar sua potência inibitória. Também enfatizamos os critérios mais cruciais a serem considerados para ensaios precisos de enzimas que proporcionam resultados reprodutíveis. As principais vantagens do protocolo descrito sobre outros métodos colorimétricos ou espectrometria de massa é que este protocolo é rápido e fácil de executar e fornecer medições quantitativas da atividade GST, bem como a potência de inibição das moléculas rastreadas.
Apresentamos a maneira de calcular os dois parâmetros michaelis-menten mais importantes de um inibidor de enzimas: o IC50 e o Ki. Um inibidor potente exercerá o menor Ki e IC50 possíveis, indicando que a afinidade entre o inibidor e a enzima é alta30. Como o IC50 depende da concentração esticada e das condições de ensaio33,pode ser difícil usar esse valor para comparar inibidores de diferentes experimentos ou obtidos utilizando outras condições de ensaio34. Usando a constante de inibição Ki é um melhor indicador da potência inibitória dos compostos potenciais. Ki pode ser usado para comparar dois inibidores com diferentes modos de inibição, pois depende apenas da afinidade entre o inibidor e a enzima. No entanto, para ter uma ideia clara da natureza da inibição é preciso determinar ambos os parâmetros do inibidor30. Medimos o IC50 e O Ki das curcuminas como 31,6 ± 3,6 μM e 23,2 ± 3,2 μM, respectivamente, indicando que este composto é um potente inibidor GST. Esses resultados confirmaram as previsões in silico, que estimaram os valores Ki entre 27,4 e 78,1 μM para diferentes isoformas humanas gst e curcumina.
Atividade enzimática ou taxa de reação e quantidade de enzima
Como dito acima, o IC50 depende da concentração enzimática, e conduzir um experimento com um nível desconhecido de atividade enzimática pode levar a falsas conclusões33. Para controlar outros fatores, que podem reduzir a atividade inibitória, deve-se considerar e medir a atividade enzimática para cada novo lote de GSTs. Por exemplo, a degradação de um lote enzimático, causada por muitos ciclos de congelamento/degelo, pode diminuir a atividade e, assim, levar a um IC50 menor, mesmo que o experimento tenha sido executado sob as mesmas condições. Em outras palavras, o uso de 0,01 unidades de enzima não dará os mesmos resultados que o uso de 1 unidade. O uso de muitas enzimas pode levar a um rápido esgotamento do substrato e a reação não terá uma forma linear. Este parâmetro poderia, assim, levar a um resultado inexato, pois nenhuma mudança na absorção seria vista após um longo período de incubação.
Valor Km
Para garantir as melhores condições para avaliar o tipo de inibição exibida por um inibidor, a concentração de substrato deve ser igual ou inferior à constante Michaelis-Menten (Km). Km é representado pela concentração de substrato necessário para ocupar metade dos locais ativos na enzima28. Por exemplo, uma maior concentração de substratos pode neutralizar um inibidor competitivo e avaliar esse tipo de inibição será difícil nesse cenário. Assim, um dos passos cruciais dessa metodologia é a determinação do Km da enzima para o substrato selecionado (aqui CDNB). Em alguns estudos, esse valor não foi determinado e isso poderia levar a falsas conclusões sobre o padrão de inibição causado pelo inibidor, e, se o modo de inibição estiver incorreto, o Ki será calculado incorretamente, pois a equação se baseia no padrão de inibição26,28. Se uma isoforma GST diferente for testada, uma nova avaliação do valor Km é obrigatória, pois este valor é único para um par de enzimas e ligantes. Como utilizamos uma concentração de CDNB ligeiramente inferior ao Km (0,2 mM), que definimos como 0,26 ± 0,08 mM, o modo competitivo previsto de inibição da curcumina no GST foi precisamente determinado.
IC50
Obter uma boa curva sigmoidal com a qual estimar o IC50 requer encontrar tanto os platôs inferiores quanto os superiores. O planalto inferior representa as concentrações de um inibidor que proporciona a atividade inibitória máxima. Em alguns casos, um composto pode não inibir totalmente a enzima, mesmo em altas concentrações, devido a questões técnicas como solubilidade. No entanto, ferramentas como GraphPad Prism podem caber no planalto inferior com bastante precisão. O planalto superior é composto por concentrações do inibidor, que são insuficientes para inibir a enzima, e, portanto, a atividade é máxima. Ambos os platôs são cruciais para determinar o IC50, bem como concentrações no meio, a fim de encontrar a inclinação da curva — então o IC50 pode ser derivado da forma da curva sigmoide35. A curcumina é mal solúvel em água, assim a concentração máxima utilizada neste ensaio é limitada, para evitar precipitação na solução de ensaio. Assim, foram utilizadas soluções menos concentradas, que não inibem totalmente a atividade GST. Isso levantou questões para a determinação do planalto inferior. Para combater esse problema, previmos valores inferiores baseados no gráfico de regressão não linear, que forneceu ic50 de 31,6 ± 3,6 μM para curcumina(Figura 3A). Para o ácido ethacúnico, não havia necessidade de prever os valores para o planalto inferior, uma vez que este composto é solúvel na solução de ensaio e o IC50 foi medido em 6,6 ± 1,1 μM.
Este método pode ser aplicado aos isóformes GST mais expressos em humanos, ou seja, GSTA1, GSTM1 ou GSTP1. No entanto, este protocolo não é adequado para quantificar a atividade da isoforme GSTT1, uma vez que o CDNB não é um substrato para este subtipo. 36, 37 Enquanto isso, o protocolo pode ser ligeiramente modificado para combater esta questão. Por exemplo, usando 1,2-epóxi-3-(4'-nitrofenoxy)propano (ENPP) como substrato para GSTT1 e medir a quantidade de conjugado a 360nm em vez de 340nm. 37
As etapas do protocolo podem ser adaptadas e aplicadas para testar a atividade GST e testes inibidores em experimentos de cultura celular. A medição da atividade GST em células tratadas com e sem um inibidor GST indicará se este composto pode ser usado em tal configuração experimental. É especialmente interessante quando o inibidor é lipofílico. Por exemplo, apresentamos que a curcumina é um potente inibidor GST usando este protocolo. No entanto, sua aplicação a estudos celulares pode ser limitada, uma vez que a molécula é mal solúvel em água e se degrada rapidamente em médio. 31 Outra amenização deste protocolo é possível quanto à especificidade não isoforme do cdnb substrato. O uso deste protocolo em estudos celulares só dará informações sobre a atividade global do GST, mas não sobre a atividade do subtipo GST preciso. A adição de substrato específico de isoforme e/ou o uso de isoformes GST recombinantes específicos permitirá testar inibidores GST específicos de isoforme.
Para concluir, descrevemos um procedimento completo para testar inibidores GST que têm potencial para serem usados em combinação com quimioterapia eletrofílica. Enfatizamos os passos cruciais de um ensaio de enzima GST e inibição, para testar moléculas interessantes potenciais e determinar sua eficiência como inibidor com valores quantitativos, o IC50 e o Ki. Este método pode ser aplicado a qualquer composto putativo e realizado nos isoforms GST humanos mais expressos (GSTA1, GSTM1 e GSTP1), ou ligeiramente adaptado para realizar estudos de cultura celular com inibidores gst ou medir a atividade de outras interessantes isoformas GST de escolha.
| Reagentes | Nome | Concentração na solução de ensaio | Outros |
| Substrato | CDNB | Mede Km (mM) | Diluído em 95% de etanol. A concentração final de etanol deve ser ≤ 5% (v/v) |
| Substrato conjugal | Gsh | 2,5 mM | Diluído na água. |
| A concentração deve saturar a solução. | |||
| Buffer | Pbs | - | pH = 7,1 |
| Enzima | Pool de isoforms GST ou isoforme puro | 0,01 unidade/mL, determinada experimentalmente. | Diluído na água. |
| Inibidor GST | Potencial composto de escolha | IC50: 3 concentrações que inibem, 3 que inibem minimamente, e 3 no meio. | Diluído em DMSO e depois na água, para ter uma concentração final de DMSO ≤ 1% (v/v) |
| Ki: 3 concentrações ao redor do IC50. | |||
| Parâmetros | |||
| Temperatura ambiente (25°C) | |||
| pH = 7,1 | |||
| DMSO ≤ 1% (v/v) | |||
| Etanol ≤ 5% (v/v) | |||
Tabela 1: Resumo dos reagentes e parâmetros a serem considerados durante um ensaio de inibição GST.
Divulgações
Os autores não têm nada a revelar.
Agradecimentos
Este trabalho de pesquisa foi apoiado pela Fundação CANSEARCH. Os autores gostariam de reconhecer a Sra. Laurence Lesne e o Sr. Yoann Sarmiento para assistência técnica, especialmente na condução dos experimentos de replicação enquanto padronizam os ensaios com inibidores. Discussões frutíferas e contribuições do Sr. Denis Marino, Dr. Simona Mlakar e Dr. Vid Mlakar são muito reconhecidas. Agradecemos à Dra. Agradecemos ao Dr. Muthukumar por suas informações sobre as previsões de Silico. Também agradecemos ao Sr. Darren Hart por sua ajuda na leitura da prova em inglês deste manuscrito.
Materiais
| Name | Company | Catalog Number | Comments |
| 1-Chloro-2,4-dinitrobenzene (CDNB) | Sigma-Aldrich | 237329 | Substrate used for the GST enzymatic assay |
| Corning UV-Transparent Microplates | Sigma-Aldrich | CLS3635 | Transparent plate to perform the enzymatic assay. When using 200 ul, the pathlength is 0.552 cm for this plate. |
| Curcumin | Sigma-Aldrich | 8511 | Used for the results section, to test the inhibition potency of curcumin |
| Dimethyl sulfoxide (DMSO) | Sigma-Aldrich | D2650 | To prepare the stock of the putative inhibitor |
| DPBS | Sigma-Aldrich | D8537 | Buffer for the enzymatic reaction |
| Ethanol 95% | Fisher scientific | 10542382 | To dilute the CDNB |
| Glutathione S-Transferase from equine liver | Sigma-Aldrich | G6511 | Used for the results section, to test the inhibition potency of curcumin |
| L-glutathione reduced (GSH) | Sigma-Aldrich | G4251 | Co-substrate for the GST enzymatic assay |
| Pierce BCA Protein Assay Kit | ThermoFisher | 23225 | To quantify the amount of protein present in the enzymatic solution |
| Spectramax iD3 | Molecular devices | To do spectrophotometric measurments |
Referências
- Mannervik, B., Danielson, U. H. Glutathione transferases--structure and catalytic activity. CRC Critical Reviews in Biochemistry. 23 (3), 283-337 (1988).
- Awasthi, Y. C., Sharma, R., Singhal, S. S. Human glutathione S-transferases. The International Journal of Biochemistry. 26 (3), 295-308 (1994).
- Rowe, J. D., Nieves, E., Listowsky, I. Subunit diversity and tissue distribution of human glutathione S-transferases: interpretations based on electrospray ionization-MS and peptide sequence-specific antisera. Biochemical Journal. 325, 481-486 (1997).
- Beckett, G. J., Hayes, J. D. Glutathione S-transferases: biomedical applications. Advances in Clinical Chemistry. 30, 281-380 (1993).
- Oakley, A. Glutathione transferases: a structural perspective. Drug Metabolism Reviews. 43 (2), 138-151 (2011).
- Townsend, D. M., Tew, K. D. The role of glutathione-S-transferase in anti-cancer drug resistance. Oncogene. 22 (47), 7369-7375 (2003).
- Tew, K. D. Glutathione-associated Enzymes in Anticancer Drug Resistance. Cancer Research. 54 (16), 4313-4320 (1994).
- Laborde, E. Glutathione transferases as mediators of signaling pathways involved in cell proliferation and cell death. Cell Death and Differentiation. 17 (9), 1373-1380 (2010).
- Adler, V., et al. Regulation of JNK signaling by GSTp. The EMBO Journal. 18 (5), 1321-1334 (1999).
- Cho, S. G., et al. Glutathione S-Transferase Mu Modulates the Stress-activated Signals by Suppressing Apoptosis Signal-regulating Kinase 1. Journal of Biological Chemistry. 276 (16), 12749-12755 (2001).
- Hayes, J. D., Flanagan, J. U., Jowsey, I. R. Glutathione Transferases. Annual Review of Pharmacology and Toxicology. 45 (1), 51-88 (2005).
- Lo, H. W., Ali-Osman, F. Genetic polymorphism and function of glutathione S-transferases in tumor drug resistance. Current Opinion in Pharmacology. 7 (4), 367-374 (2007).
- Ansari, M., et al. Glutathione S-transferase gene variations influence BU pharmacokinetics and outcome of hematopoietic SCT in pediatric patients. Bone Marrow Transplantation. 48 (7), 939-946 (2013).
- Mahajan, S., Atkins, W. M. The chemistry and biology of inhibitors and pro-drugs targeted to glutathione S-transferases. Cellular and molecular life sciences: CMLS. 62 (11), 1221-1233 (2005).
- Allocati, N., Masulli, M., Ilio, C. D., Federici, L. Glutathione transferases: substrates, inihibitors and pro-drugs in cancer and neurodegenerative diseases. Oncogenesis. 7 (1), 8 (2018).
- Aggarwal, B. B., Kumar, A., Bharti, A. C. Anticancer potential of curcumin: preclinical and clinical studies. Anticancer Research. 23 (1), 363-398 (2003).
- Han, S. S., Chung, S. T., Robertson, D. A., Ranjan, D., Bondada, S. Curcumin causes the growth arrest and apoptosis of B cell lymphoma by downregulation of egr-1, c-myc, bcl-XL, NF-kappa B, and p53. Clinical Immunology. 93 (2), 152-161 (1999).
- Golonko, A., Lewandowska, H., Świsłocka, R., Jasińska, U. T., Priebe, W., Lewandowski, W. Curcumin as tyrosine kinase inhibitor in cancer treatment. European Journal of Medicinal Chemistry. 181, 111512 (2019).
- Awasthi, S., et al. Curcumin-glutathione interactions and the role of human glutathione S-transferase P1-1. Chemico-Biological Interactions. 128 (1), 19-38 (2000).
- Appiah-Opong, R., Commandeur, J. N. M., Istyastono, E., Bogaards, J. J., Vermeulen, N. P. E. Inhibition of human glutathione S-transferases by curcumin and analogues. Xenobiotica; the Fate of Foreign Compounds in Biological Systems. 39 (4), 302-311 (2009).
- Dubey, V., Owusu-Apenten, R. Curcumin Restores Glutathione-S-Transferase Activity for LNCaP Prostate Cancer Cells. Pure and Applied Chemistry. 2, 61-72 (2014).
- Habig, W. H., Pabst, M. J., Jakoby, W. B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. The Journal of Biological Chemistry. 249 (22), 7130-7139 (1974).
- Johnson, K. A., Goody, R. S. The Original Michaelis Constant: Translation of the 1913 Michaelis-Menten Paper. Biochemistry. 50 (39), 8264-8269 (2011).
- Ochs, R. S. Understanding Enzyme Inhibition. Journal of Chemical Education. 77 (11), 1453 (2000).
- Bisswanger, H. Enzyme assays. Perspectives in Science. 1 (1), 41-55 (2014).
- Acker, M. G., Auld, D. S. Considerations for the design and reporting of enzyme assays in high-throughput screening applications. Perspectives in Science. 1 (1), 56-73 (2014).
- Cheng, A. L., et al. . Anticancer Research. 21, 2895-2900 (2001).
- Yang, J., Copeland, R. A., Lai, Z. Defining Balanced Conditions for Inhibitor Screening Assays That Target Bisubstrate Enzymes. Journal of Biomolecular Screening. 14 (2), 111-120 (2009).
- Uppugunduri, C. R. S., Muthukumaran, J., Santos-Silva, T., Ansari, M. Identification of putative substrates and inhibitors for Glutathione S-transferases using computational methods. Zenodo. , (2017).
- Burlingham, B. T., Widlanski, T. S. An Intuitive Look at the Relationship of Ki and IC50: A More General Use for the Dixon Plot. Journal of Chemical Education. 80 (2), 214 (2003).
- Wang, Y. J., et al. Stability of curcumin in buffer solutions and characterization of its degradation products. Journal of Pharmaceutical and Biomedical Analysis. 15 (12), 1867-1876 (1997).
- Copeland, R. A. Evaluation of enzyme inhibitors in drug discovery. A guide for medicinal chemists and pharmacologists. Methods of Biochemical Analysis. 46, 1 (2005).
- Kalliokoski, T., Kramer, C., Vulpetti, A., Gedeck, P. Comparability of Mixed IC50 Data - A Statistical Analysis. PLoS ONE. 8 (4), (2013).
- Brandt, R. B., Laux, J. E., Yates, S. W. Calculation of inhibitor Ki and inhibitor type from the concentration of inhibitor for 50% inhibition for Michaelis-Menten enzymes. Biochemical Medicine and Metabolic Biology. 37 (3), 344-349 (1987).
- Brooks, H. B., et al. Basics of Enzymatic Assays for HTS. Eli Lilly & Company and the National Center for Advancing Translational Sciences. , (2012).
- Arakawa, S., et al. Evaluation of hepatic glutathione transferase Mu 1 and Theta 1 activities in humans and mice using genotype information. Drug Metabolism and Disposition: The Biological Fate of Chemicals. 40 (3), 497-503 (2012).
- Sherratt, P. J., Pulford, D. J., Harrison, D. J., Green, T., Hayes, J. D. Evidence that human class Theta glutathione S-transferase T1-1 can catalyse the activation of dichloromethane, a liver and lung carcinogen in the mouse. Comparison of the tissue distribution of GST T1-1 with that of classes Alpha, Mu and Pi GST in human. The Biochemical Journal. 326 (3), 837-846 (1997).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados