
Method Article
Optimizado coloração negativa: um protocolo de alta capacidade para Examinando pequeno e assimétrico Estrutura de proteínas por microscopia eletrônica
Neste Artigo
Resumo
Mais da metade das proteínas são pequenas proteínas (massa molecular <200 kDa) que são um desafio tanto para elétron imagem ao microscópio e reconstruções tridimensionais. Coloração negativa otimizado é um protocolo robusto e de alto rendimento para a obtenção de alto contraste e imagens de pequenas proteínas ou complexos assimétricas em diferentes condições fisiológicas relativamente alta resolução (~ 1 nm).
Resumo
Determinação estrutural de proteínas é bastante desafiador para proteínas com massas moleculares entre 40 - 200 kDa. Considerando-se que mais de metade das proteínas naturais têm uma massa molecular entre 40 - 200 kDa, 1,2, é necessário um método robusto e de alto rendimento, com uma capacidade de resolução nanométrica. Microscopia coloração negativa (NS) elétron (EM) é uma abordagem fácil, rápida e qualitativa que tem sido frequentemente usado em laboratórios de pesquisa para examinar a estrutura de proteína e proteína-proteína interações. Infelizmente, os protocolos de NS convencionais muitas vezes geram artefatos estruturais em proteínas, especialmente com as lipoproteínas que normalmente formam apresentando artefatos Rouleaux. Usando imagens de lipoproteínas de microscopia crio-elétrons (crio-EM) como um padrão, os principais parâmetros em NS condições de preparação da amostra foram recentemente selecionados e relatou como o protocolo NS otimizado (OPNS), um protocolo NS convencional modificado 3. Artefatos como rouleaux pode ser muito limitado por OPNS, adicionalmente, proporcionando alto contraste junto com razoavelmente alta resolução (perto de 1 nm) imagens de pequenas e assimétricas proteínas. Estas imagens de alta resolução e alto contraste são ainda favoráveis para uma proteína individual (um único objeto, não média) reconstrução 3D, como um anticorpo de 160 kDa, através do método de tomografia eletrônica de 4,5. Além disso, OPNS pode ser uma ferramenta de alto rendimento para analisar amostras de centenas de pequenas proteínas. Por exemplo, o mecanismo anteriormente publicada de 53 kDa da proteína de transferência de éster de colesterol (CETP) envolveu a triagem e imagiologia de centenas de amostras 6. Considerando crio-EM raramente com sucesso imagens proteínas inferior a 200 kDa ainda tem de publicar qualquer estudo envolvendo triagem mais de cem condições da amostra, é justo chamar OPNS um método de alto rendimento para o estudo de pequenas proteínas. Esperemos que o protocolo OPNS aqui apresentada pode ser uma ferramenta útil para empurrar os limites de EM e acelerar estudos de EM na estrutura da proteína pequena, dinâmica e mecanismos.
Introdução
Compreender a função da proteína exige conhecimento da estrutura da proteína. A determinação estrutural é um desafio para as proteínas cujas massas molecular estão dentro de 40 - 200 kDa. A cristalografia de raios-X está limitado por cristalização de proteínas; ressonância magnética nuclear (RMN) é limitado a massas moleculares inferiores a 40 kDa, ao passo que a microscopia crio-electrão (crio-EM) tem dificuldade em ambos aquisição de imagem e reconstruções tridimensionais (3D) de pequenas proteínas, que as massas moleculares são menos a 200 kDa. Notavelmente,% de proteínas mais de 50 têm uma massa molecular na faixa de 40 - 200 kDa 1,2, como os métodos atuais são um desafio em estudar proteínas desse porte, é necessário um novo método.
Embora a maioria dos microscópios eletrônicos de transmissão (ETM) são capazes de resolução atômica, ou seja, mais de 3 Å de resolução, conseguindo mesmo uma estrutura perto resolução nanométrica a partir de um material biológico é bastante challenGing 7. Os danos da radiação, baixo contraste, os desvios estruturais, bem como artefatos como desidratação todos os impedem de alta resolução de imagem TEM 3,8.
Entre as várias abordagens TEM, crio-EM é um método avançado e de ponta para alcançar estruturas de resolução atômicas de macromoléculas altamente simétricas sob condições fisiológicas perto de 9-12. A amostra de crio-EM é preparada flash de congelação da solução de amostra, a incorporação das macromoléculas em gelo vítreo, que é posteriormente trabalhada a temperaturas criogénicas tais como azoto ou hélio temperaturas do líquido 13. Cryo-EM é vantajoso em que as amostras não apresentam artefatos e são quase nativa em estrutura 8-12. Cryo-EM tem suas desvantagens: i) dispositivos adicionais são necessários para ser instalado ou comprado para atualizar um instrumento TEM padrão para uma capacidade de crio-EM. Os dispositivos incluem: anti-contaminador, porta-crio, software modo de baixa dose e sensi baixa dosecâmera CCD tiva, embora os preços destes dispositivos são muito mais baixos do que o preço do próprio instrumento TEM; ii) operação de crio-EM tem mais tempo do que a operação NS. Examinando uma amostra de crio-EM, muitas vezes requer mais tempo para preparar as amostras e operar o instrumento TEM do que a de NS porque crio-EM, é necessário resolver as dificuldades adicionais, incluindo: líquido operação temperatura do nitrogênio, amostra de carga, tração de imagem, gradientes de temperatura, baixa dose modelo de operação, as sensibilidades de radiação das amostras e os limites de dosagem. Estas etapas extras vão diminuir a velocidade de aquisição de dados úteis em relação a aquisição de dados NS, embora algumas imagens crio-EM podem certamente ser obtido em 1 hora ou menos por especialistas crio-los com o instrumento preparado com um gradiente de temperatura equilibrada; iii) os usuários precisam de treinamento adicional, como a manipulação de nitrogênio líquido, congelamento grades crio-EM, a operação em baixa dose, medida de dose, a manipulação da carga, à deriva e conhecimento em proc imagemessing; iv) falta de imagiologia repetível para os mesmos crio exemplar durante as diferentes sessões de TEM. Espécimes Cryo-EM são facilmente danificados pela contaminação de gelo durante a carga e descarga de amostra de / para o instrumento TEM. Este dano é especialmente uma preocupação quando as amostras são difíceis de ser isolados / purificados 14; v) proteínas pequenas (<200 kDa de massa molecular) são um desafio a ser trabalhada por causa do baixo contraste; vi) a partir de contraste e de ruído elevado de imagens crio-EM reduz o valor da correlação cruzada entre as imagens, portanto, diminuindo a precisão total na determinação da orientação da proteína, conformações e classificações, especialmente para as proteínas que são estruturalmente flexível e, naturalmente, variar em solução 4,5.
Coloração negativa (NS) é relativamente "antigo" e histórica método que qualquer laboratório, com qualquer tipo EM, pode utilizar para analisar a estrutura da proteína. Brenner e Horne primeiro desenvolveu o conceito of coloração negativa de meio século atrás para examinar vírus 15. NS é realizado por meio de revestimento da amostra com sais de metais pesados carregados. Este conceito originalmente vindo de microscopia de luz e a prática de incorporar as bactérias em uma solução mancha fornecendo escuridão ao redor dos espécimes, permitindo maior contraste de imagem para ver a imagem negativa 16. Uma vez que os íons de metais pesados têm uma maior capacidade para dispersar os elétrons em comparação com átomos de menos densas nas proteínas 17-20, e revestimento de metais pesados mancha permite uma limitação dosagem superior com melhor contraste. Espécime NS pode fornecer imagens de alto contraste 8 para facilitar a determinação orientação das partículas e reconstrução 3D de imagens de crio-EM.
NS tradicionais, infelizmente, pode produzir artefatos induzidos por interações mancha em proteínas, como a agregação geral, a dissociação molecular, achatamento e empilhamento 8,21,22. Para lipídico relacionado proteins, como lipoproteínas 16,23-30, um artefato comum resulta em partículas que são empilhados e embalados juntos em um rouleaux (Figura 1) 31-36. Muitos estudos de lipoproteína, tais como a electroforese em gel não desnaturante de poliacrilamida de gradiente, crio-EM estuda 13,29,37-40, espectrometria de massa de 39,41, e os dados de difracção de raios-X de ângulo pequeno 42 mostra que todas as partículas de lipoproteínas são partículas isoladas naturalmente, em vez de empilhada em conjunto, formando uma rouleaux 21,29,30,35,42-45. A observação da formação rouleaux por NS convencionais é possivelmente causada por interações dinâmicas entre lipoproteínas compostas de apolipoproteínas (APO) e fosfolipídios que são estruturalmente flexível em solução 13,29,30,46-49 e sensibilidade para o protocolo NS padrão. Para identificar esse artefacto, apolipoproteína E4 (apoE4) palmitoil-oleoylphosphatidylcholine (POPC) lipoproteína de alta densidade (HDL) da amostra foram utilizados como uma amostra de teste e crioterapiaImagens EM para um padrão artefato livre 29, a triagem das amostras NS elaborados sob uma série de condições. Ao comparar os tamanhos de partículas e formas obtidas a partir de NS e crio-EM, o tipo específico de reagente de coloração e concentração de sal foram considerados dois parâmetros principais fazendo com que os fenómenos ROULEAUX bem conhecidos. Assim, um protocolo de coloração negativa optimizado (OPNS) foi relatada.
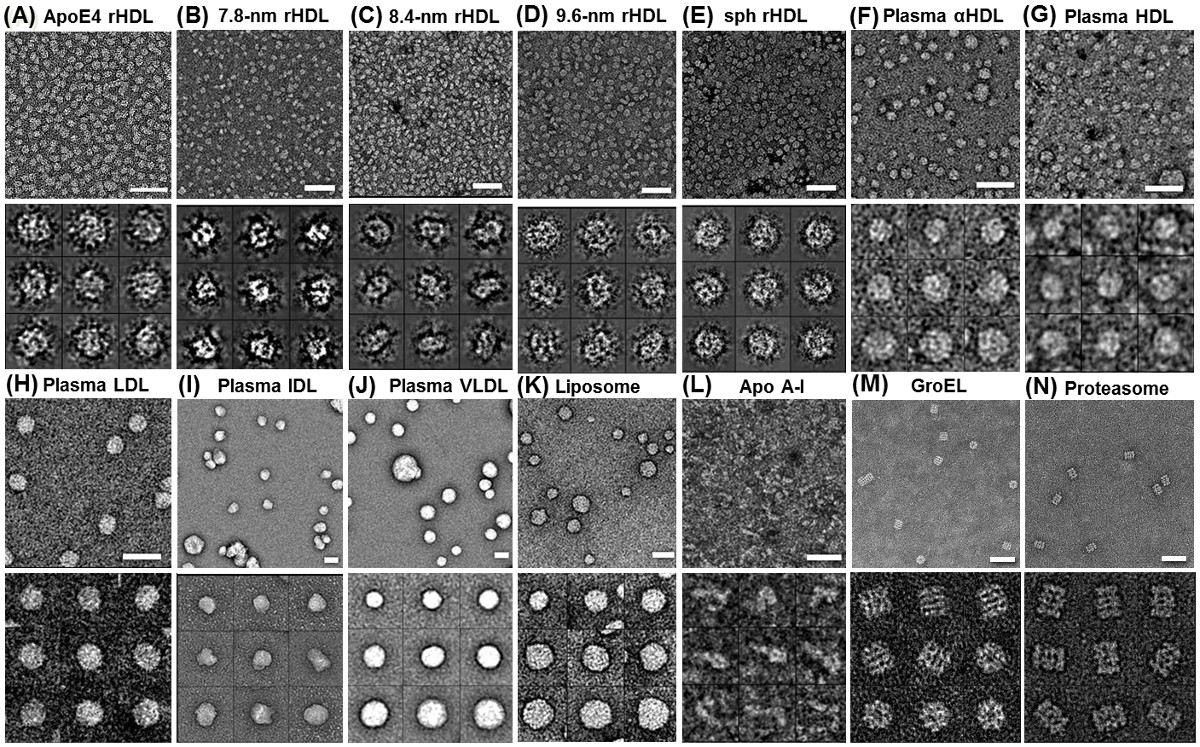
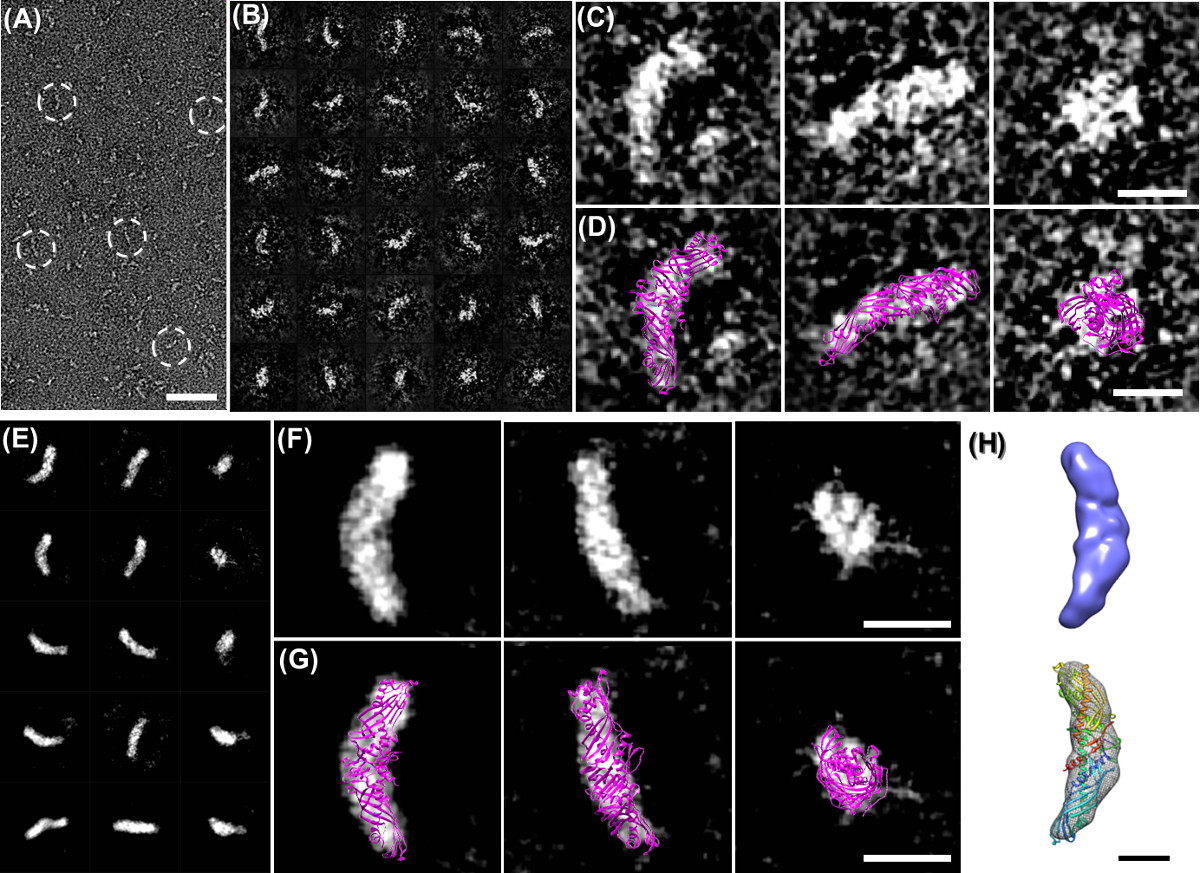
Por OPNS, o fenómeno bem conhecido de rouleaux apoE4 HDL foi eliminado por OPNS (Figura 2A). A análise estatística demonstrou OPNS rendimentos imagens muito semelhantes (menos de 5% do desvio) em tamanho e forma, em comparação com os de crio-EM, no entanto, o contraste foi eliminado. As validações de OPNS foram realizadas por análise de eliminação do artefacto rouleaux de quase todas as classes e subclasses de amostras de lipoproteína 6,29,30,50,51, incluindo a apo AI 7,8 nm (Figura 2B), 8,4 nm (Figura 2C) , 9,6 nm discoidal HDL reconstituídas (rHDL) (Figura 2D), 9,3 nm esférica rHDL (Figura 2E), o HDL no plasma humano (Figura 2F), livre de lípidos de apo AI (Figura 2G), o HDL no plasma (Figura 2H), lipoproteína de baixa densidade (LDL ) (Figura 2I), lipoproteínas de densidade intermédia (IDL) (Figura 2J), lipoproteínas de muito baixa densidade (VLDL) (Figura 2K), e lipossomas de POPC (Figura 2D) 30. Validações adicionais foram realizados por imagiologia pequenas e assimétricas proteínas, incluindo a proteína de 53 kDa de transferência de éster de colesterol (CETP) (Figura 3A - C) de 6,29, e altamente flexível 160 kDa anticorpo IgG (Figura 4A e B) 4,5,29 , 52, e duas proteínas estruturalmente bem conhecidos, e GroEL de proteassoma (Figura 2 M e N). Para requerer qualquer validação adicional de colleagues, estamos abertos a quaisquer testes cegos sobre este método OPNS.
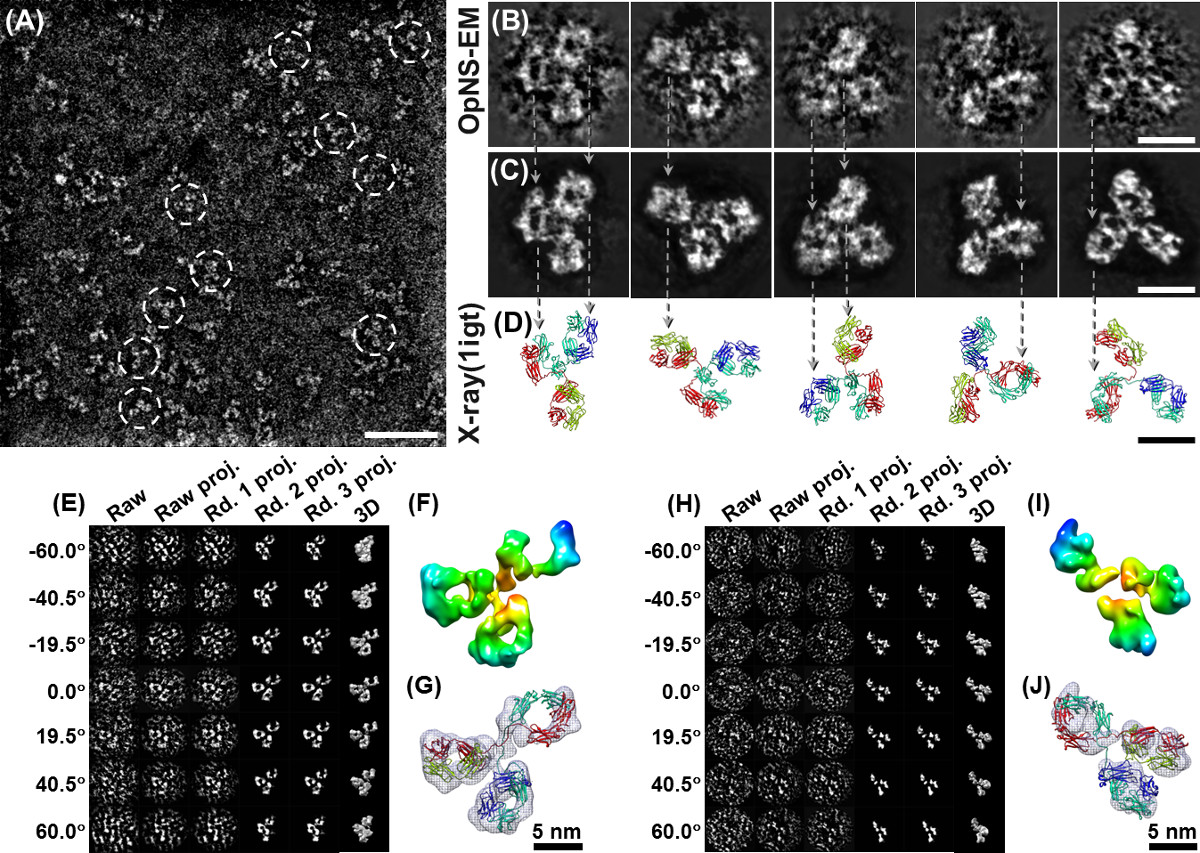
OPNS como um protocolo de alto rendimento também tem sido utilizada para estudar mecanismo de proteína através de análise de centenas de amostras de pequenas proteínas, tais como foi CETP que se ligam a várias proteínas sob uma série de condições, incluindo (CETP interagindo 4 classes de lipoproteínas, recombinados HDL plasma HDL, LDL e VLDL, com / sem dois anticorpos, H300 e N13, sob vezes 9 de incubação, incluindo 3 min, 20 min, 1 hora, 2 horas, 4 horas, 8 horas, 24 horas, 48 horas e 72 horas, com menos de 4 proporções molares, por exemplo, 1: 0,5, 1: 1, 1: 2, 1: 4, e 3 diluições, ou seja, 0,1 mg / ml, 0,01 mg / ml e 0,001 mg / ml, mais amostras de controlo adicionais, incluindo CETP sozinho, LDL sozinho, e VLDL sozinho, com testes de triplos de experiências anteriores por parte de pessoas diferentes) 6,29. OPNS imagens de CETP fornecido imagens de alto contraste com detalhes estruturais razoavelmente finas; o que nos permite reconstruir com sucesso um mapa de densidade 3D da 53 kDa smaproteína CETP ll (Figura 3D - F) pela reconstrução única partícula. Além disso, as imagens OPNS alto contraste nos fornecer um sinal suficiente de uma proteína individual (Figura 4A - C), o que nos permitiu alcançar a resolução intermediária (~ 1,5 nm) de um único (um objeto, não média) estrutura de anticorpos IgG 3D através do método (Figura 4E - J) tomografia elétron-partícula individual (Ipet) 5. A descrição detalhada da estratégia ipet reconstrução, a metodologia, os processos passo-a-passo e análise de variações estruturais foram previamente relatados 4. Um filme sobre procedimentos de reconstrução de anticorpos ipet, incluindo as imagens cruas e resultados intermediários, mapa de densidade 3D e acoplagem estrutural foi também disponível ao público por upload para o YouTube 5. Comparação das reconstruções 3D a partir de diferentes partículas de anticorpos individuais poderiam revelar a dinâmica das proteínas e cmudanças onformational durante reações químicas 4,5.
Considerando-se que mais de 50% de proteínas têm massa molecular variando 40-200 kDa 1,2, o sucesso em imagem destas pequenas proteínas evidenciado que o método OPNS é uma ferramenta útil para empurrar a fronteira EM convencional para determinações estruturais pequenas e assimétricas e um mecanismo de descobertas . Assim, o protocolo detalhada é fornecida abaixo.
Protocolo
1 Preparação da solução fresca de coloração negativa em 1% (w / v)
- Coloque 1 mg uranilo Formate (UF) do pó em frasco de vidro contendo 100 ml de água deionizada em uma sala ou caixa escura.
- Agita-se a solução de O / N à temperatura ambiente numa sala escura / caixa. Embrulhe o frasco da solução com papel alumínio para evitar que a luz incida a solução.
- Filtra-se a solução suavemente, através de 0,2 um (tamanho de poro) de filtro montado em uma seringa de 5 ml. A solução filtrada foi recolhido em várias folhas de alumínio cobertos tubos Falcon. A seringa de filtro também deve ser envolvida com papel de alumínio para evitar a exposição à luz.
- Filtra-se a solução novamente através de 0,02 um (tamanho de poro) de filtro montado em seringa de 1 ml. A solução foi filtrada e foi recolhido em alíquotas de 2 ml. As seringas e tubos de ensaio a ser coberto com uma folha de alumínio, antes da utilização.
- Congelar os frascos alíquotas usando uma pinça de cabo longo submergindo os frascos dentro de um recipiente cheio de nitrogênio líquido imediatamentedepois a solução foi filtrada.
- Transfira os frascos congelados em um freezer -80 ° C para armazenamento e uso futuro.
2 Preparação de Workstation coloração negativa e Incubação Workstation
- Descongelar um frasco de solução UF 1% em um banho de água regulado para RT. Certifique-se de que a folha de alumínio permanece enrolada em torno do frasco para evitar a exposição à luz.
- Filtra-se a solução depois de ser completamente descongelado, usando uma folha de alumínio enrolado seringa de 1 ml montado com um filtro de 0,02 ^ m. Coletar a solução filtrada para uma nova folha de alumínio envolto frasco, e colocou-a sobre o gelo dentro de uma caixa de gelo cap coberto espera para o uso.
- Faça placas de solução pelo dedo empurrando uma folha longa Parafilm ~ 8 polegadas para a superfície de uma placa de cristalização de proteínas vazio. Ele irá gerar linhas de placas circulares com um diâmetro de ~ 5 milímetros. Fazer 6 placas em cada fila lugar as placas Parafilm sobre uma superfície de gelo achatado dentro de gelo contêmer tampa, como uma estação de trabalho de coloração.
- Pipeta ~ 35 mL de água deionizada em cada um dos restantes três placas em cada linha, e pipetas ~ 35 mL filtrada solução UF para direita três placas em cada linha. Cobrir a estação de trabalho de coloração com uma tampa para evitar a exposição à luz antes de corar as proteínas.
- Prepara-se uma estação de incubação grade EM preenchendo uma caixa a meio caminho vazio com gelo, e adere ao lado de um gancho que pode ser montado sobre uma base de apoio. Coloque as pinças de exemplo no cabide, nos quais dicas das pinças estão próximas da superfície do gelo. Metade cobrir dicas das pinças por a altura da caixa lábio. Uma caixa de gelo ideal para fazer uma estação de incubação está usando um recipiente pontas de pipeta vazio.
3. OPNS Operação
- Glow-descarregar a película de carbono fina revestida 300 grids EM cobre malha por 10 seg.
- Pegue uma grade com uma pinça e ligar a pinça para dentro do hangar, mantendo a rede em um 45 ° de inclinação e de 1 polegada acima do gelosuperfície interior da estação de incubação.
- Dilui-se a amostra de proteína com fosfato de Dulbecco tamponado salino (DBPS) a concentração final de proteína de 0,01 a ~ ~ 0,005 mg / ml. Depósito ~ 4 ul diluída amostra imediatamente após a diluição do lado da grade de carbono de EM. (DPBS pode ser mudado para um outro tampão, se a proteína for sensível a DPBS)
- Incubar a amostra nas grades EM para ~ 1 min dentro da estação de incubação.
- Remover o excesso de solução através rapidamente tocar a borda da rede de papel de filtro.
- Toque a grelha rapidamente para a primeira gota (~ 35 ul) de superfície de água pura em cima da folha de Parafilm direito depois o excesso de solução foi removido da grelha EM. E em seguida, retire o excesso de água rapidamente com papel de filtro.
- Repita o passo 3.6 para mais duas vezes, lavando a grade EM nas restantes duas gotas de água na Parafilm (Passo 3.6 e 3.7 pode ser ignorada se a amostra é muito sensível à água).
- Flutuar o gri EMd imediatamente sobre a superfície da primeira gota de UF solução logo após o excesso de água sobre a rede EM foi removido. E então incubar durante 10 seg. Certifique-se para terminar os procedimentos de lavagem de água dentro de 3 segundos antes da grelha flutuante na superfície da solução UF. Quanto mais curto for o tempo de lavagem, o melhor será a qualidade.
- Limpe as pinças penetrando as dicas em um papel de filtro para ~ 2-3 vezes.
- Remover o excesso de solução da grelha por meio de contacto da borda de grade com o papel de filtro, e em seguida a grelha flutuar sobre a segunda queda de UF.
- Continuar acima passo 3.10 do segundo gota.
- Float a grade no terceiro gota de solução UF e cobrir a estação de coloração para ~ 1 min.
- Passo opcional: lavar a grelha rapidamente no topo de água, conforme descrito em 3.6. Este passo adicional irá beneficiar amostras contendo mais de partículas de ouro livre carregado ou fundo mancha.
- Retire o excesso de solução tocando no papel de filtro para potodo e backside grade (lado oposto ao de carbono). Seca-se a grade sob uma ligeira corrente de gás de azoto à temperatura ambiente, depois de observar bem a solução de absorção para o papel de filtro.
- Coloque a grelha sobre uma folha de papel de filtro no interior de uma caixa de Petri, e cobrir parcialmente a placa com uma tampa para secar pelo menos durante 30 minutos sob temperatura ambiente. Para algumas amostras, coloque a grelha em um 40 ° C incubadora assar por uma hora.
- Guarde a grade em uma caixa de grade EM para o futuro EM examinando.
4 EM Examinando
- Alinhe o TEM EM corretamente antes de exame das pequenas proteínas no grid.
- Verifique a resolução do mais alto visível Thon-ring que é o espectro de um filme de carbono amorfo poder de determinar se o TEM foi alinhada corretamente. Desde o TEM é compartilhada por muitos usuários diferentes, o TEM foi muitas vezes não devidamente alinhados.
- Verifique cuidadosamente o espectro de energia na área de filme de carbono da amostra para ajustar o alinhamento condition da máquina. As maiores visíveis Thon-rings são melhores do que a resolução de destino.
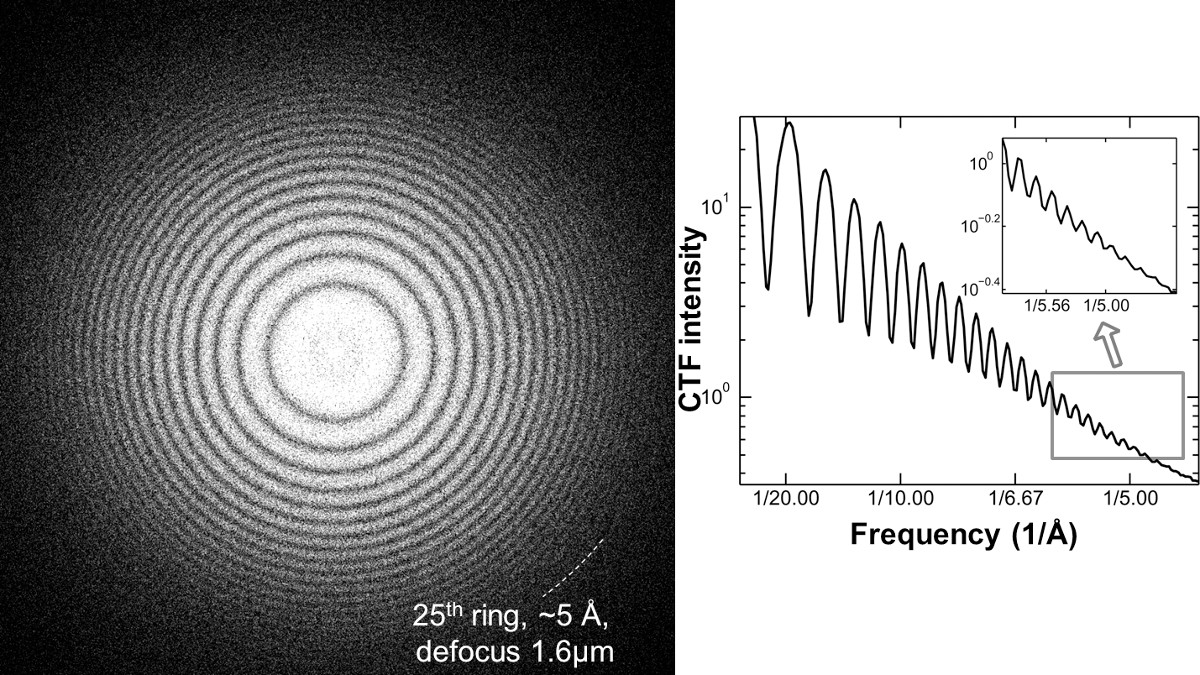
NOTA: Como um exemplo de um alinhamento adequado de uma MET LaB6 filamento operado sob alta tensão de 120 kV, mais do que 20 Thon-anéis podem ser visualizados, no qual, a resolução correspondente pode ser melhor do que 5,0 Å. Este Thon-anel foi conseguida por transferência de Fourier de uma película de carbono amorfo visualizados sob uma desfocagem de ~ 1,6 ^ m e uma dose de 20,4 electrónico / A2 (Figura 5).
NOTA: A observação da alta resolução Thon-ring é uma condição necessária, mas não uma condição suficiente para o alinhamento correto. Para conseguir efectivamente as imagens de alta resolução, muitos outros parâmetros são também importantes, tais como a qualidade da amostra, danos de radiação, e que deriva de carregamento, bem como a taxa de dose de iluminação.
- Traga a desfocagem de volta para quase Scherzer foco para imagens de pequenas proteínas em uma área ideal manchado.
- Pesquisar por área "nublado" a image. Uma área manchada idealmente está geralmente localizada perto da extremidade de uma área de mancha mais espessa, que se parece com uma área de "turvo" na grelha uma vez que a espessura da mancha é normalmente não uniformemente distribuídos na grelha revestida de carbono (Figura 6). Normalmente, um pequeno aumento (<100x) era ajudar a encontrar essas áreas nubladas antes de zoom in para geração de imagens.
Resultados
As implementações de OPNS incluem os exames estruturais e morfológicas de várias espécies de lipoproteínas, tais como: HDL nascente recombinante (rHDL), HDL recombinante esférica, o HDL no plasma, colesterol LDL, IDL e VLDL (Figura 2) 30; bem como proteínas tais como pequenas 53 kDa de CETP (entre as proteínas mais pequenas fotografados com sucesso por meio de EM) (Figura 3) 6 e anticorpos IgG 160kDa (uma das proteínas mais dinâmicos e heterogéneos) (Figura 4) 4,5,29,52 , até 28 kDa livre de lípidos apolipoproteína A-1 (Figura 2D), e GroEL e os proteossomas (Figura 2 M e N).
Outros exemplos de OPNS como um método de alto rendimento são examinar interacções proteína-proteína para descobrir os mecanismos da proteína, tais como a 53 kDa de CETP. Como descrito na introdução, como CETP interage com diferentes subclasses de lipoproteínas foram invested via examinando mais de 400 espécimes EM 6 Tanto quanto sabemos, não houve um caso de estudo crio-EM que envolve triagem quase uma centena de condições diferentes.
OPNS pode expandir fronteiras EM estudar pequeno e assimétrico estrutura da proteína em 3D e até mesmo expandir para alcançar estrutura 3D formar um único e individual proteína (sem média). Por exemplo, um anticorpo IgG é 160 kDa molécula com uma estrutura altamente flexível, em que a reconstrução 3D a resolução intermédia é difícil de ser conseguido. o método OPNS foi utilizado para a imagem de um anticorpo IgG a partir de um indivíduo série de ângulos de inclinação (Figura 4E e H). As imagens do OPNS permitiram a reconstrução bem sucedida de uma única proteína por tomografia eletrônica de partícula individual (Ipet) (Figura 4E - J) razoavelmente alta resolução e alto contraste de proteína 4,5. Na reconstrução ipet 3D 4, a série de inclinação 2D do anticorpo alvo foram gradualmente alinhada ao seu centro global calculado através de reconstrução iterativa, posteriormente, para atingir uma densidade 3D com uma resolução de ~ 14,1 Å (Figura 4F e G) 4. Pelo mesmo método, um único complexo de anticorpo-peptídeo foi reconstruído, e o péptido de domínio induzida a mudança conformacional interna de uma única partícula de anticorpo foi descoberto (Figura 4I e J, resolução ~ 16,6 Å) 4,5. Comparando-se as reconstruções 3D de diferentes partículas individuais, a análise estrutural pode permitir-nos para revelar proteínas flutuações termodinâmicas, e até mesmo "snap-shot" os estágios intermediários de uma reação química 4,5,29,53,54.
Em resumo, as proteínas com massa molecular entre 40 kDa - 200 kDa são um desafio para os exames estruturais EM padrão e reconstruções. Considerando-se que mais de 50%de proteínas têm massa molecular variando 40-200 kDa 1,2, relatamos um método OPNS como uma ferramenta robusta e de alto rendimento para empurrar a fronteira EM convencional para determinações estruturais pequenas e assimétricas e até mesmo estudos mecanicistas.

Figura 1 Rouleau artefato de lipoproteínas por coloração negativa (NS) EM convencional. Eletromicrografias (acima) e painéis de partículas selecionadas (abaixo de painéis) mostram várias lipoproteínas com rouleaux após NS com ácido fosfotúngstico (PTA) no âmbito do processo mix padrão. (A) reconstituído HDL (rHDL) com ApoE4, (B) apo AI contendo 9,6 nm discoidal rHDL, (C) contendo apoB LDL do plasma, (D) não contendo apolipoproteína lipossomas POPC. Bares: 50 nm. O tamanho da janela: A e C, 20 nm; B, 30 nm. The Journal of Lipid Research inicialmente publicado este trabalho 29,30. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 2 estrutura de proteínas por coloração negativa otimizado micrografias (OPNS) EM eletrônica mostram várias lipoproteínas e lipossomas sem rouleaux artefato por OPNS (A) apoE4 contendo rHDL;.. (B) 7,8 nm rHDL; (C) 8,4 nm rHDL; ( D) 9,6 nm rHDL, (E) 9,3 nm esférica rHDL, (F) de plasma humano α-HDL, (L) de plasma de HDL, (H) de LDL do plasma humano, (I), IDL de plasma humano, (J) de VLDL do plasma humano; (K) POPC lipossomas; (L) lipídico livre apo AI. Verificação adicional foi feito usando (M) GroEL e amostras (N) do proteassoma. Micrografias (painéis superiores) e partículas selecionadas (abaixo de painéis). Bares: 50 nm. O tamanho da janela: A - D, 20 nm; E e F, de 25 nm; L, 20 nm; H, 25 nm; I, 50 nm; J e K, de 100 nm; L, 80 nm., Exceto lipídico livre apo AI, GroEL e Proteasome, Esta pesquisa foi originalmente publicado no Journal of Lipid Research 29,30. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 3 micrografias eletrônicas OPNS de CETP. (A) micrografia Overview com CETP (tracejada círculos) partículas em forma de banana. (B) windowed Selecione partículas matérias. (C) Três únicas imagens de partículas CETP bruto mostrando claramente uma maior ponta globular, mais pesadas e maiores, incluindo a outra ponta mais leve e mais estreito (painel esquerdo) menor, vista de cima, mostrando regiões terminais semelhantes com menos curvatura global (média painel), para baixo e vista final ao longo do eixo da CETP (painel da direita) (D) A sobreposição da estrutura de cristal (PDB:. 2OBD) para estas imagens de partículas cruas de CETP, corresponde bem na forma e tamanho do domínio estrutural, mostrando a orientação pode ser quase directamente definida, mesmo sem o auxílio de computador (painéis correspondente a (C)) (E). Referência médias classe 2D livres (um processo ab initio para calcular as semelhanças (cálculo de correlação cruzada) dessas partículas orientadas aleatoriamente, tendo as partículas um elevada similaridade entre si foram agrupados, alinhados e depois feita a média para reduzir o ruído de fundo e melhorada a imagem das partículas contraste. As médias de classe 2D livre de referência são calculados pelo software refine2D.py no pacote EMAN, em que, sem qualquer modelo inicial humano feito foi envolvido nesta média da turma. As médias classe de referência pode ser utilizado como um método independente para validar a reconstrução 3D da reconstrução de uma única partícula). Referência (F) seleccionado médias livres classe 2D mostram uma extremidade distal maior em comparação com o outro. (L) da estrutura cristalina Sobreposta (APO : 2OBD) comparação com as médias gratuitos de referência, as imagens de sobreposição mostrar um perto de combinações perfeitas entre crystal-estrutura e média classe livre de referência, tanto em estrutura forma e tamanho domínio (baixo painel) (F) mapa de densidade 3D da CETP em um. Resolução de 13 Å foi reconstruído a partir de 8879 partículas fotografada por OPNS e método de reconstrução de uma única partícula (painel superior) e do corpo estriado atracou nesta reconstrução de uma única partícula pela estrutura cristalina (baixo do painel). O detinformações ailed do single reconstrução de partículas, incluindo o pré-processamento de imagem, modelo inicial, os procedimentos de refinamento, distribuição angular e Shell Fourier Correlação (FSC) foram publicados na seção de método e informações de apoio do trabalho original (H) Comparação final de reconstrução única partícula ( acima do painel) e ajuste comparação PDB adicional (painel inferior) 6. Bares: A, 50 nm; B - D, 10 nm; F, 3 nm. Alguns destes números foram publicados anteriormente na revista Nature Chemical Biology 6. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4 OPNS imagens e reconstruções de IgG humana partículas 1 de anticorpos. (A) Ao longovista de imagens anticorpo IgG1 OPNS humanos. Partículas mostrados em círculos brancos exibir claramente as partículas com 3 subdomínios. (B) Selecionado de alta resolução imagens cruas de cinco partículas individuais não conjugados de anticorpos e (c) as suas imagens com redução de ruído correspondentes ao reduzir manualmente ruído ambiente borda das partículas da matéria (sem contato nenhum densidade no interior das partículas) para visualizar facilmente. (D) A estrutura de cristal (APO 1IGT entrada) de anticorpo IgG1 que foi orientado para uma visualização da mesma forma para as imagens de EM. Comparação das imagens correspondentes mostra muitas características comuns entre a imagem EM e da estrutura cristalina, incluindo posições e formas de domínio. (E) O procedimento passo-a-passo de uma reconstrução ab initio 3D a partir de um único anticorpo exemplo IgG1 (sem média, um objeto individual) usando o método de tomografia elétron-partícula individual (Ipet). (F) A reconstrução 3D final de uma sipartícula anticorpo ngle na resolução de 14,1 Å mostrou três domínios em forma de anel formando um "Y". (L) de ancoragem da estrutura de cristal domínios em cada uma correspondente densidade de domínio mapeia respectivamente, e repetindo manualmente as laçadas entre os domínios. (H) O procedimento passo-a-passo de reconstrução 3D de um único complexo IgG-péptido (sem média, um objecto individual) por ipet. (I) A reconstrução 3D final de um complexo de péptido de IgG foi apresentado na resolução de 16,6 Å mostrou três haste domínios em forma, formando um "Y" (J), respectivamente, a estrutura de encaixe de cada cristal domínios de anticorpo de IgG (entrada PDB: 1IGT) em cada haste. densidades forma mostrou uma correspondência mal com estruturas cristalinas de domínio, sugerindo uma mudança conformacional domínio interno após a conjugação de péptidos. O procedimento detalhado, análise e estatísticas resolução foram descritas no artigo original 5 Bares: A, 50 nm; B - D, 10 nm. Este trabalho inicialmente publicado na PLoS ONE 4 e Scientific Reports 5. Clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 5. espectro de potência de filme de carbono amorfo fotografada sob 1,6 mM de desfocagem por uma Zeiss Libra 120 LaB6 TEM. Alta resolução de imagem requer provas suficientes para mostrar que EM tem a capacidade de alinhamento e de alta resolução apropriada. A análise do espectro de potência de carbono área nas proximidades da rede é um método eficiente para verificar a condição de coerência do feixe antes da aquisição de dados. Transferência de Fourier de uma imagem de uma área de carbono amorfo mostrar a função de transferência de contraste instrumento relacionado (C TF), chamado Thon-rings. Um alinhamento e coerência pode ser refletido pelo número de visíveis Thon-rings e a mais alta resolução de anéis Thon visíveis sob uma condição relativamente alta de desfocagem. Como exemplo de uma condição de alinhamento adequado perto de um laboratório 6 filamento TEM equipada, mais de ~ 20 Thon anéis (~ 5 a) pode ser gerado a partir de um filme de carbono são gravadas em 1,6 mM desfocar usando doses 20,4 e - / a 2. Os anéis Thon obtidos a partir de um TEM low-end têm uma semelhante ao do TEM high-end, como uma arma de emissão de campo (FEG) reforçada TEM operado sob a condição de alinhamento adequado. Este trabalho inicialmente publicada na revista Scientific Reports 5. Clique aqui para ver uma versão maior desta figura.
{kind=link}
"Width =" 6highres.jpg 500 "/>
Figura 6 Exemplo micrografias de áreas "ideais" para imagiologia. OPNS Três amostras de proteína, (A) GroEL (B) do proteassoma (C) esférico HDL, foram utilizadas como exemplos para demonstrar as OPNS ideais para imagiologia. Desde manchas são desigualmente distribuídos na amostra, a baixa ampliação da amostra geralmente aparece "nublado" (painel à esquerda). As áreas ideais para a imagem normalmente estão localizados nos limites dessas "nuvens". Um exemplo de passo-a-passo de zoom-in uma região "nublado" stain (painel médio esquerdo) mostram as imagens de alta ampliação apresentam alta resolução e imagens de alto contraste de partículas (painel do meio à direita). As imagens seleccionadas de partículas mostrar os detalhes da estrutura de proteínas (painel da direita). Bares:. 30nm Por favor, clique aqui para ver um grandeversão r desta figura.
{kind=link}

A Figura 7 um diagrama esquemático de procedimentos OPNS. (A) Estação grade EM incubação, estação simples para incubar a solução da amostra no grid brilho descarregada. (B) estação de trabalho de coloração, projetado para manter as gotas de água de lavagem, e gotas de manchas acima uma cama de gelo, além de minimizar a exposição mancha de luz. (C) Visão geral do processo de coloração, ilustrando: direcção mata-borrão, água de lavagem 3x, exposição mancha 3x UF, a incubação da amostra de coloração com grade invertida em gotículas de manchas, e parte traseira da amostra final de mata-borrão para assegurar uma camada fina de solução de corante de amostra antes da secagem por nitrogênio do ar. Clique aqui para ver uma versão maior do thé figura.
{kind=link}

Figura 8 alta - resolução de imagens de pequena proteína 53kDa CETP revelado a partir de um método OPNS modificado, crio-positivo-coloração (crio-PS) Cinco escolha de alta resolução e formato circular imagens cruas mascarados suaves de partículas CETP (com a reversão de contraste). e forma das partículas mascarado partículas (matérias-ruído reduzido manualmente cercam as bordas dos imagens das partículas da matéria, mas sem modificar qualquer densidade dentro das partículas) para visualizar facilmente. All-átomo e fita de cristal comparação estrutura para mascarado partículas matérias-alinhados com orientações semelhantes de cada partícula de imagens EM. Os detalhes de alta resolução das áreas de imagem RAW apresentar certa semelhança com a estrutura de cristal, enquanto nem todos os recursos de estrutura de cristal pode ser resolvido a partir de dados brutos. Remarka Bly, estas imagens matérias mostram semelhança em ambos os terminais acaba tendo nas proximidades pequenos orifícios circulares vistas na redução manualmente imagens de ruído (triângulos); Além disso, os filamentos nas regiões C-terminais de partículas dentro de imagens complementar estrutura cristalina β-folhas (setas) e, finalmente, laços salientes na região C-terminal de CETP são afins na estrutura cristalina (losangos). (A) Cinco seleccione de alta resolução e formato circular imagens suaves mascarados matérias de partículas CETP (com a reversão de contraste). (B) Filtro passa baixa para 10-A. (c) maior filtragem passa-baixo a 20 (D). mascarados imagens manualmente ruído reduzido. (E) Cristal comparação da estrutura de estrutura de fita. (f) comparação da estrutura de cristal por toda a representação átomo. Bar: 5 nm. Parte deste trabalho foi publicado na revista Nature Chemical Biology 6.escorrido "> Clique aqui para ver uma versão maior desta figura.

Figura 9 Eletromicrografias de lipossomas mostrando formação Rouleau por protocolo NS convencional (PTA mancha) sob diferentes salinidade. (A) de alta salinidade (~ 0,5 MNaCl). (B) salinidade regular (~ 0,25 M de NaCl). (C) Baixa salinidade (~ 0,1 M de NaCl). (D) A água pura. Micrografias (acima do painel) e selecione partículas janela (abaixo do painel) demonstrada. Bares: 100 nm. O tamanho da janela: 80 nm. The Journal of Lipid Research originalmente publicado este trabalho 30. Clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
Em comparação com as técnicas convencionais NS, OPNS pode evitar artefatos Rouleaux (Figura 1 e 9), fornecendo alta resolução confiável e razoável (~ 1 nm) detalhes estruturais de pequenas proteínas (Figura 2). Comparado com crio-EM, OPNS fornece um método de alto rendimento e pode examinar uma ampla variedade de proteínas e interacções proteína-proteína 6. No entanto, OPNS ainda tem suas desvantagens. Comparado a NS convencionais, OPNS implica: i) as medidas mais complicadas na preparação de amostras; ii) utilização de uma substância radioactiva, UF; iii) manter a mancha fresco devido à potência menor do que a mancha por causa da sua sensibilidade à luz UF; iv) para evitar a precipitação da solução de UF deve ser armazenado em -80 ° C e requer a descongelação antes da utilização; v) e, finalmente, tudo UF devem ser tratados como resíduos perigosos. Comparado com crio-EM, OPNS fornece imagens com resolução relativamente mais baixos e sem garantia de qualquer artefacto ainda não descobertono futuro.
Operação NS efeitos processuais sobre a formação de lipoproteínas rouleaux
NS protocolos para a preparação de proteínas para exame 16,29-36,55 podem ser classificados em três grandes grupos de métodos 50: mistura 55, gota-a-gota 16, e procedimentos de lavagem 29. i) O protocolo de mistura requer a mistura prévia da amostra e mancha de proteína a uma razão específica, e, em seguida, a aplicação da solução mista numa película de carbono revestido em uma grade, finalmente, removendo o excesso de solução com papel de filtro antes de secagem de ar para o EM exame 55. ii) O protocolo de gota-a-gota, envolve a aplicação de (~ 4 mL) gota amostra directamente a uma rede de EM revestido em carbono para deixar sit ~ 1 min, subsequentemente, removendo o excesso de solução da amostra, antes da aplicação de uma gota (~ 4 mL) de mancha solução no mesmo lado da grelha. Após cerca de 1 min de incubação, retirar os excess mancha através do contato com papel de filtro limpo, finalmente enviar para secagem ao ar 16,56-58. iii) O protocolo de lavagem (Figura 7) que requer a aplicação de uma solução de amostra grade EM revestido em carbono, durante 1 min, seguida de lavagem com água desionizada três vezes logo após a remoção do excesso de solução de cada vez por papel de filtro 3,29,30. OPNS protocolo foi modificado a partir deste procedimento de lavagem.
NS tipos de manchas e efeitos sobre a formação de lipoproteínas rouleaux
No NS, mais forte espalhamento de elétrons devido a manchas de metais pesados proporcionar contraste das proteínas 17-20,59. Amostras NS pode ainda sofrer doses maiores de radiação, e melhorar características de proteína por um contraste maior negativo 8,9,60. Manchas de metais pesados podem ser classificadas como: aniônicos, incluindo o ácido fosfotúngstico (PTA) 16, metilamina tungstate 14 e tungstato de silício 61; e cátionsic, tais como acetato de uranila (UA) 62, UF 3,29,30, e nitrato de uranilo (ONU) 63. Uma das manchas NS aniônicos que é mais comumente usado é PTA 33,64-67. PTA é um heteropoliácido, que é tipicamente utilizado em torno de um pH de 7,0-7,5. PTA, também pode ser utilizado para a coloração positiva 3,16,68. As cargas negativas de PTA podem mediar interações eletrostáticas com lipídios insaturados positivamente carregadas grupos de cabeça, como POPC, em ambas as superfícies de lipoproteínas e lipossomas. PTA pode causar a formação de rouleaux através desta interação 29,30, mesmo sob 29,30 salinidade tampão normal (Figura 1). Manchas de uranila, como UA, UF e da ONU são escolhas alternativas para catiônicos manchas de metais pesados e UA, em particular, é frequentemente usado para NS de uma variedade de espécimes biológicos 62,69,70. Experiências encontrado UF não causa rouleaux de lipoproteínas que contêm lípidos de ácido gordo insaturado, tal como POPC. No entanto, podem ainda induzir UF rouleformação aux lipoproteína que contêm lípidos de ácidos gordos saturados, tais como a fosfatidilcolina de dimiristoílo (DMPC) e 1-hexadecanoil-2-octadecanoil-sn-glicero-3-fosfocolina (PSPC). O mecanismo detalhado dessa interação é desconhecida. UA / UF / ONU podem trabalhar em menores valores de pH entre 3,5 e 4.6. Estes valores de pH mais baixos podem não adequado para certas macromoléculas biológicas que são sensíveis ao pH 14,71. Curiosamente, UA / UF pode corrigir a estrutura das proteínas dentro de poucos milésimos de segundo através de um mecanismo desconhecido 72. Ao contrário de PTA, UA / UF é mais semelhante a um sal de um ácido.
UF supostamente pode proporcionar melhores resultados do UA 73, em que, UF e da ONU manchas têm tamanhos de grãos menores do que UA (UF: 0,3 nm, da ONU: ~ 0,5 nm) 3,74. Um exemplo interessante, a estrutura semelhante a detalhes secundários de uma proteína pequena (53 kDa de CETP) podem ser visualizados directamente a partir das micrografias matérias quando UF foi utilizado como reagente corante negativo, seguindo orelatado anteriormente crio-coloração positiva (crio-PS) método (Figura 8) 6. Em crio-PS, a amostra foi manchado flash congelado, seguindo crio-EM procedimento de preparação da amostra em vez de processo de secagem em protocolo OPNS, o que pode causar o colapso estrutural secundário. A crio-PS é um método de alto contraste e imagens de alta resolução de pequenas proteínas 75. Desde que as imagens Cryo-PS reverteram contraste em comparação com os do protocolo de crio-NS relatou 22, mas tem contraste de imagem consistente para aqueles a partir de imagens de crio-EM convencionais, assim foi chamado o método de crio-PS. As imagens crio-PS mostram que o reagente de coloração, uranilo formato, penetra na superfície molecular, desafiando a visão convencional de que a coloração pode visualizar somente a estrutura da superfície exterior. O mecanismo de como uranilo formato penetra na superfície molecular não é conhecido. Uma possibilidade é que o catião de uranilo liga os grupos carboxilo da proteína, disponíveis, e assim, ocircundante gelo vítreo é de densidade mais baixa do que a coloração da proteína e os grupos de uranilo, actuando assim como uma mancha positiva. O método de crio-PS é semelhante ao método de substituição isomorfa múltipla (MIR), que era uma abordagem comum para resolver o problema de fase em cristalografia de raios X 76-78. No MIR, amostras de cristal são embebidos com uma solução de átomo pesado, incluindo urânio, para obter um formulário isomorfo a sua forma nativa. A adição do átomo pesado, por vezes, não afeta as dimensões celulares formação de cristais ou unidade, mas fornece informações adicionais de cristal estrutura determinação 76-78. Por beneficiando o pequeno grão de UF, crio-PS poderia fornecer uma imagem de alta resolução e alto contraste de uma única proteína, o que é importante para o estudo da estrutura de proteínas, especialmente quando se considera que quase todas as proteínas são naturalmente dinâmico e heterogêneo em solução. Para a validação do método de crio-PS, abrimos para um teste cego do campo EM.
Efeitos da concentração de sal na formação rouleaux nas lipoproteínas
Usando PTA tradicional reagente coloração negativa, formação rouleaux observado é mais provavelmente relacionados com interações lipídicas em vez de interação proteína. Imagiologia as amostras de lipossomas de POPC vesículas preparadas de acordo com considerável salinidade (NaCl 0,5 M), salinidade moderada (NaCl 0,25 M), relativamente fraca salinidade (NaCl 0,1 M), e água pura (Figura 9), as micrografias electrónicas de mostrar a formação foi rouleaux correlacionada com a concentração de sal (Figura 9A - D). Usando o reagente de coloração de UF como negativo, não foi observada em rouleaux POPC amostra lipossoma vesícula 30 (Figura 2D), o que sugere o processo de lavagem para reduzir rapidamente a concentração de sal é um direito antes de coloração necessária, mas não suficiente para o passo independentemente impedindo rouleaux em POPC relacionada amostras.
imagem TEM pequena proteína requer um estado quase-foco Scherzer.
Imagem TEM de sucesso de pequenas proteínas com maior resolução exige duas condições críticas, um alinhamento do feixe adequada e uma condição de aquisição foco quase Scherzer. i) Um alinhamento do feixe adequada pode ser julgado pelo número de toques Thon visíveis (espectro de energia) sobre a transferência de Fourier de uma imagem de filme de carbono adquiridos ao abrigo de um relativamente alto de desfocagem. Quanto melhor for a condição de alinhamento do feixe, mais anéis Thon pode ser visualizado e mensurável ii) Adquirir imagem sob uma quase foco Scherzer .. é outro estado crítico. Uma estratégia padrão tradicional para imagiologia de amostras biológicas normalmente utiliza alta desfocagem (1 - 2 ^ m e ainda mais), o que pode aumentar o contraste da imagem da amostra biológica 79. Esta estratégia é significativamente diferente da estratégia típica para geração de imagens estrutura resolução atômica de materiais duros, que muitas vezesutiliza um foco Scherzer próximo (~ 50 nm desfocar). Embora, a desfocagem maior poderia fortalecer biológica contraste da amostra, os sinais de alta resolução seria parcialmente eliminado. Como resultado, a cumplicidade de sinal de alta resolução seria inferior a uma condição superior de imagem desfocar. A razão é que, a imagem TEM é a convolução da estrutura da amostra e o CTF instrumento. O CTF é uma curva oscilante contra a frequência, na qual a curva frequentemente oscilado cruzando amplitude de zero a alta frequência. Cada momento em que CTF através de zero, o sinal de estrutura de amostra a esta frequência específica será eliminado (zero vezes qualquer número será zero). O sinal eliminado a esta frequência não pode ser recuperado por qualquer algoritmo de correcção de CTF (zero dividido por zero pode ser qualquer número aleatório, em vez de o sinal estrutural original). Sob maior imagem desfocar, o CTF irá oscilar muito mais agressiva, assim, a CTF atravessa amplitude zero, com mais freqüência, como resultado,os sinais estruturais a um maior número de frequências específicas serão eliminadas. Os mais sinais que são eliminadas, a menos cumplicidade a imagem terá. Notavelmente, a cumplicidade de imagem não pode ser recuperado ou corrigido por qualquer correção CTF. No entanto, um número de imagens adquiridas em diferentes incompletas desfoca pode ser usado para preencher os intervalos entre si para obter um sinal estrutural completa da estrutura de amostra através de um método de compensação, em que mesmo uma resolução atómica pode ser conseguida 9-12.
O método de cálculo da média é uma abordagem poderosa para conseguir a estrutura de algumas proteínas altamente simétricos e proteínas relacionadas estruturalmente rígidos. No entanto, continua a ser um desafio no estudo estrutural de pequenas proteínas e assimétricos, em especial para a dinâmica e proteínas estruturais flexíveis, tais como anticorpos e HDL. Média é baseada na suposição de que as partículas de proteína são idênticos em estrutura e conformação, mas different em orientações, no entanto, muitas proteínas são conhecidas por sofrer flutuação termodinâmico em solução. Ao aplicar o método de média sem antes conhecer as flutuações de flutuação termodinâmicas de proteínas, a estrutura média, muitas vezes pode causar falta domínios 10,80 ou variação local na resolução 81 em reconstruções crio-EM.
O sucesso na obtenção de reconstrução 3D da pequena proteína, tal como 53 kDa de CETP, e a estrutura 3D de uma única proteína, tal como o anticorpo 160 kDa IgG1, os benefícios de utilizar a condição de imagem foco Scherzer quase sob uma condição de alinhamento adequado. Embora o contraste da imagem parece baixo no curto Scherzer foco imagens, o contraste pode ser facilmente melhorada simplesmente aplicando uma filtragem passa-baixa para reduzir o ruído de alta freqüência em segundo plano. Acreditamos que, a curto-Scherzer focar imagens transportam os sinais estruturais máximo, e pode aumentar a precisão do alinhamento das partículas e aumentar a eficiência em de uma única partícula de reconstrução 3D e partícula individual tomografia eletrônica de reconstrução.
Divulgações
We have nothing to disclose.
Agradecimentos
Agradecemos ao Dr. Mickey Yang para edição e comentários, e as Dras. Lei Zhang e Peng Bo para obter assistência. Este trabalho foi financiado pela Secretaria de Ciência, Instituto de Ciências Básicas da Energia do Departamento de Energia dos Estados Unidos (contrato n. DE-AC02-05CH11231), o National Heart, Lung, and Blood Institute dos National Institutes of Health (sem . R01HL115153) e do Instituto Nacional de Ciências Médicas Gerais dos Institutos Nacionais de Saúde (sem. R01GM104427).
Materiais
| Name | Company | Catalog Number | Comments |
| Stain: Uranyl Formate | The SPI Supplies Division of Structure Probe, Inc. | 02545-AA | http://www.2spi.com/catalog/chem/Uranyl_Formate.shtml |
| Syringe: 1 ml NORM-JECT | VWR | 89174-491 | https://us.vwr.com/store/catalog/product.jsp?catalog_number=89174-491 |
| Syringe filter: 0.2 μm | VWR | 28145-487 | https://us.vwr.com/store/catalog/product.jsp?catalog_number=28145-487 |
| Syringe filter: 0.02 μm filter (Anotop 10) | Whatman | 6809-1002 | http://www.whatman.com/AnotopSyringeFilters.aspx |
| Dulbecco’s Phosphate Buffered Saline | SIGMA-Aldrich | D8537 | http://www.sigmaaldrich.com/catalog/product/sigma/d8537?lang=en®ion=US |
| Parafilm: 4 in. x 125 ft. | VWR | 470152-246 | https://us.vwr.com/store/catalog/product.jsp?catalog_number=WLS65710-A |
| Carbon film coated TEM grid: 300 mesh | Pacific Grid-Tech | Cu-300CN | http://www.grid-tech.com/catalog.htm#1-thincarbon |
| Carbon film coated TEM grid: 200 mesh | EMS (Electro Microscopy Sciences) | CF200-Cu | http://www.emsdiasum.com/microscopy/products/grids/support.aspx |
| Tweezer: Dumont Tweezer L5 | EMS (Electro Microscopy Sciences) | 72882-D | http://www.emsdiasum.com/microscopy/products/tweezers/dumont_clamping.aspx#02-L5 |
| Milli-Q Advantage A10 Ultrapure Water Purification System | EMD Millipore | Milli-Q Advantage A10 | http://www.millipore.com/catalogue/module.do?id=C10117 |
| Filter Paper: Grade 1 circles | Whatman | 1001-090 | http://www.whatman.com/QualitativeFilterPapersStandardGrades.aspx |
| Aluminum Foil | Any type | ||
| Filter Tips: Low-Retention Aerosol Filter Tips for 10µL Pipettors | VWR | 89174-520 | https://us.vwr.com/store/catalog/product.jsp?catalog_number=89174-520 |
| [header] | |||
| Glow Discharge Unit | Ted Pella | 91000 | http://www.tedpella.com/easiglow_html/Glow-Discharge-Cleaning-System.htm |
| Filter Tips: Low-Retention Aerosol Filter Tips for 40µL Pipettors | VWR | 89174-524 | https://us.vwr.com/store/catalog/product.jsp?catalog_number=89174-524 |
| Filter Tips: VWR Signature 200 µL Pipet Tips | VWR | 37001-528 | https://us.vwr.com/store/catalog/product.jsp?catalog_number=37001-528 |
| Pipet | Pipetman | F161201 and F167300 | http://www.pipetman.com/Pipettes/Single-Channel/PIPETMAN-Classic.aspx |
Referências
- Lipman, D. J., Souvorov, A., Koonin, E. V., Panchenko, A. R., Tatusova, T. A. The relationship of protein conservation and sequence length. BMC Evol Biol. 2, (2002).
- Tran, J. C., et al. Mapping intact protein isoforms in discovery mode using top-down proteomics. Nature. 480, 254-258 (2011).
- Ohi, M., Li, Y., Cheng, Y., Walz, T. Negative Staining and Image Classification - Powerful Tools in Modern Electron Microscopy. Biol Proced Online. 6, 23-34 (2004).
- Zhang, L., Ren, G. IPET and FETR: experimental approach for studying molecular structure dynamics by cryo-electron tomography of a single-molecule structure. PLoS ONE. 7, e30249 (2012).
- Tong, H., et al. Peptide-conjugation induced conformational changes in human IgG1 observed by optimized negative-staining and individual-particle electron tomography. Sci Rep. 3, 1089 (2013).
- Zhang, L., et al. Structural basis of transfer between lipoproteins by cholesteryl ester transfer protein. Nat Chem Biol. 8, 342-349 (2012).
- Henderson, R. Realizing the potential of electron cryo-microscopy. Q Rev Biophys. 37, 3-13 (2004).
- Sander, B., Golas, M. M. Visualization of bionanostructures using transmission electron microscopical techniques. Microsc Res Tech. 74, 642-663 (2011).
- Liu, H., et al. Atomic structure of human adenovirus by cryo-EM reveals interactions among protein networks. Science. 329, 1038-1043 (2010).
- Liao, M., Cao, E., Julius, D., Cheng, Y. Structure of the TRPV1 ion channel determined by electron cryo-microscopy. Nature. 504, 107-112 (2013).
- Baker, M. L., et al. Validated near-atomic resolution structure of bacteriophage epsilon15 derived from cryo-EM and modeling. Proceedings of the National Academy of Sciences of the United States of America. 110, 12301-12306 (2013).
- Bai, X. C., Fernandez, I. S., McMullan, G., Scheres, S. H. Ribosome structures to near-atomic resolution from thirty thousand cryo-EM particles. eLife. 2, e00461 (2013).
- Ren, G., et al. Model of human low-density lipoprotein and bound receptor based on cryoEM. Proc Natl Acad Sci U S A. 107, 1059-1064 (2010).
- Bremer, A., Henn, C., Engel, A., Baumeister, W., Aebi, U. Has negative staining still a place in biomacromolecular electron microscopy. Ultramicroscopy. 46, 85-111 (1992).
- Brenner, S., Horne, R. W. A negative staining method for high resolution electron microscopy of viruses. Biochim Biophys Acta. 34, 103-110 (1959).
- Forte, T. M., Nordhausen, R. W. Electron microscopy of negatively stained lipoproteins. Methods in enzymology. 128, 442-457 (1986).
- Colliex, C., et al., Prince, E., et al. . International Tables For Crystallography. , 259-429 (2006).
- Ren, G., Zuo, J. M., Peng, L. -. M. Accurate Measurements of Crystal Structure Factors Using a FEG Electron Microscope Using Digital Micrographs. Micron. 28, 459-467 (1997).
- Peng, L. M., Ren, G., Dudarev, S. L., Whelan, M. J. Robust parameterization of elastic and absorptive electron atomic scattering factors. Acta Crystallographica Section A. 52, 257-276 (1996).
- Peng, L. M., Ren, G., Dudarev, S. L., Whelan, M. J. Debye-Waller factors and absorptive scattering factors of elemental crystals. Acta Crystallographica Section A. 52, 456-470 (1996).
- Melchior, V., Hollingshead, C. J., Cahoon, M. E. Stacking in Lipid Vesicle Tubulin Mixtures Is an Artifact of Negative Staining. Journal of Cell Biology. 86, 881-884 (1980).
- De Carlo, S., Harris, J. R. Negative staining and cryo-negative staining of macromolecules and viruses for TEM. Micron. 42, 117-131 (2011).
- Allan, V., Vale, R. Movement of Membrane Tubules Along Microtubules In - Evidence for Specialized Sites of Motor Attachment. Journal of Cell Science. 107, 1885-1897 (1994).
- Catte, A., et al. Novel changes in discoidal high density lipoprotein morphology: a molecular dynamics study. Biophys J. 90, 4345-4360 (2006).
- Forester, G. P., Tall, A. R., Bisgaier, C. L., Glickman, R. M. Rat intestine secretes spherical high density lipoproteins. J Biol Chem. 258, 5938-5943 (1983).
- Gantz, D. L., Walsh, M. T., Small, D. M. Morphology of sodium deoxycholate-solubilized apolipoprotein B-100 using negative stain and vitreous ice electron microscopy. Journal of Lipid Research. 41, 1464-1472 (2000).
- Pentikainen, M. O., Lehtonen, E. M., Kovanen, P. T. Aggregation and fusion of modified low density lipoprotein. J Lipid Res. 37, 2638-2649 (1996).
- Tall, A. R., Green, P. H., Glickman, R. M., Riley, J. W. Metabolic fate of chylomicron phospholipids and apoproteins in the rat. J Clin Invest. 64, 977-989 (1979).
- Zhang, L., et al. An optimized negative-staining protocol of electron microscopy for apoE4 POPC lipoprotein. J Lipid Res. 51, 1228-1236 (2010).
- Zhang, L., et al. Morphology and structure of lipoproteins revealed by an optimized negative-staining protocol of electron microscopy. J Lipid Res. 52, 175-184 (2011).
- Gong, E. L., et al. Discoidal complexes containing apolipoprotein E and their transformation by lecithin-cholesterol acyltransferase. Biochim Biophys Acta. 1006, 317-328 (1989).
- Schneeweis, L. A., Koppaka, V., Lund-Katz, S., Phillips, M. C., Axelsen, P. H. Structural analysis of lipoprotein E particles. Biochemistry. 44, 12525-12534 (2005).
- Raussens, V., et al. Orientation and mode of lipid-binding interaction of human apolipoprotein E C-terminal domain. Biochem J. 387, 747-754 (2005).
- Li, X., Kan, H. Y., Lavrentiadou, S., Krieger, M., Zannis, V. Reconstituted discoidal ApoE-phospholipid particles are ligands for the scavenger receptor BI. The amino-terminal 1-165 domain of ApoE suffices for receptor binding. J Biol Chem. 277, 21149-21157 (2002).
- Innerarity, T. L., Pitas, R. E., Mahley, R. W. Binding of arginine-rich (E) apoprotein after recombination with phospholipid vesicles to the low density lipoprotein receptors of fibroblasts. J Biol Chem. 254, 4186-4190 (1979).
- Lu, B., Morrow, J. A., Weisgraber, K. H. Conformational reorganization of the four-helix bundle of human apolipoprotein E in binding to phospholipid. J Biol Chem. 275, 20775-20781 (2000).
- van Antwerpen, R., La Belle, M., Navratilova, E., Krauss, R. M. Structural heterogeneity of apoB-containing serum lipoproteins visualized using cryo-electron microscopy. J Lipid Res. 40, 1827-1836 (1999).
- van Antwerpen, R., et al. Cryo-electron microscopy of low density lipoprotein and reconstituted discoidal high density lipoprotein: imaging of the apolipoprotein moiety. J Lipid Res. 38, 659-669 (1997).
- Silva, R. A., et al. Structure of apolipoprotein A-I in spherical high density lipoproteins of different sizes. Proc Natl Acad Sci U S A. 105, 12176-12181 (2008).
- van Antwerpen, R. Preferred orientations of LDL in vitreous ice indicate a discoid shape of the lipoprotein particle. Arch Biochem Biophys. 432, 122-127 (2004).
- Davidson, W. S., Silva, R. A. Apolipoprotein structural organization in high density lipoproteins: belts, bundles, hinges and hairpins. Curr Opin Lipidol. 16, 295-300 (2005).
- Peters-Libeu, C. A., Newhouse, Y., Hatters, D. M., Weisgraber, K. H. Model of biologically active apolipoprotein E bound to dipalmitoylphosphatidylcholine. J Biol Chem. 281, 1073-1079 (2006).
- Peng, D. C., Song, C., Reardon, C. A., Liao, S. S., Getz, G. S. Lipoproteins produced by ApoE-/- astrocytes infected with adenovirus expressing human ApoE. Journal of Neurochemistry. 86, 1391-1402 (2003).
- Peters-Libeu, C. A., Newhouse, Y., Hall, S. C., Witkowska, H. E., Weisgraber, K. H. Apolipoprotein E*dipalmitoylphosphatidylcholine particles are ellipsoidal in solution. J Lipid Res. 48, 1035-1044 (2007).
- Jones, M. K., et al. Assessment of the validity of the double superhelix model for reconstituted high density lipoproteins: a combined computational-experimental approach. J Biol Chem. 285, 41161-41171 (2010).
- Carnemolla, R., et al. The specific amino acid sequence between helices 7 and 8 influences the binding specificity of human apolipoprotein A-I for high density lipoprotein (HDL) subclasses: a potential for HDL preferential generation. J Biol Chem. 283, 15779-15788 (2008).
- Sivashanmugam, A., et al. Structural basis of human high-density lipoprotein formation and assembly at sub nanometer resolution. Methods Cell Biol. 90, 327-364 (2008).
- Cavigiolio, G., et al. The interplay between size, morphology, stability, and functionality of high-density lipoprotein subclasses. Biochemistry. 47, 4770-4779 (2008).
- Chen, B., et al. Apolipoprotein AI tertiary structures determine stability and phospholipid-binding activity of discoidal high-density lipoprotein particles of different sizes. Protein Sci. 18, 921-935 (2009).
- Zhang, L., Tong, H., Garewal, M., Ren, G. Optimized negative-staining electron microscopy for lipoprotein studies. Biochim Biophys Acta. 1830, 2150-2159 (2013).
- Tong, H. M., Zhang, L., Huang, L. Q., Ren, G. Optimized negative-staining protocol for electron microscopy study of lipoprotein structure. Progress in Biochemistry and Biophysics. 39, 972-978, doi:Doi 10.3724/Sp.J.1206.2012.00224. 39, 972-978 (2012).
- Zhang, L., Kaspar, A., Woodnutt, G., Ren, G. Monitoring the Structural Changes of Conjugated Antibodies by High-Resolution Electron Microscopy and Individual-Particle Electron Tomography. Biophysical Journal. 98, 440-441 (2010).
- Ren, G., Zhang, L. Asymmetric Small Protein Structure Determination by Individual Particle Electron Tomography. Biophysical journal. 102, 394a (2012).
- Zhang, L., Ren, G. Structural Determination of Heterogeneous Protein by Individual-Particle Electron Tomography - Combination of Electron Tomography and Local Refinement Reconstruction Method for High-Resolution Structural Determination of Each Individual Protein Particle. Biophysical journal. 98, 441a (2010).
- Forte, T., Norum, K. R., Glomset, J. A., Nichols, A. V. Plasma lipoproteins in familial lecithin: cholesterol acyltransferase deficiency: structure of low and high density lipoproteins as revealed by elctron microscopy. J Clin Invest. 50, 1141-1148 (1971).
- Fang, Y., Gursky, O., Atkinson, D. Lipid-binding studies of human apolipoprotein A-I and its terminally truncated mutants. Biochemistry. 42, 13260-13268 (2003).
- Gursky, O., Ranjana, D. L., Gantz, Complex of human apolipoprotein C-1 with phospholipid: thermodynamic or kinetic stability. Biochemistry. 41, 7373-7384 (2002).
- Jayaraman, S., Gantz, D. L., Gursky, O. Effects of protein oxidation on the structure and stability of model discoidal high-density lipoproteins. Biochemistry. 47, 3875-3882 (2008).
- Ren, G., Peng, L. M. The Analytic Doyle-Turner Representation of High Energy Electron Absorptive Structure Factors. Acta Physica Sinica. 45, 1344-1349 (1996).
- Ren, G., Reddy, V. S., Cheng, A., Melnyk, P., Mitra, A. K. Visualization of a water-selective pore by electron crystallography in vitreous ice. Proc Natl Acad Sci U S A. 98, 1398-1403 (2001).
- Camejo, G., Suarez, Z. M., Munoz, V. The apo-lipoproteins of human plasma high density lipoprotein: a study of their lipid binding capacity and interaction with lipid monolayers. Biochim Biophys Acta. 218, 155-166 (1970).
- Kondo, A., Muranaka, Y., Ohta, I., Kanno, T. Dynamic reaction in a homogeneous HDL-cholesterol assay visualized by electron microscopy. Clin Chem. 45, 1974-1980 (1999).
- Tufteland, M., Ren, G., Ryan, R. O. Nanodisks derived from amphotericin B lipid complex. J Pharm Sci. 97, 4425-4432 (2008).
- Clay, M. A., Pyle, D. H., Rye, K. A., Barter, P. J. Formation of spherical, reconstituted high density lipoproteins containing both apolipoproteins A-I and A-II is mediated by lecithin : cholesterol acyltransferase. Journal of Biological Chemistry. 275, 9019-9025 (2000).
- Desilva, H. V., Masoliva, J., Taylor, J. M., Mahley, R. W. Identification of Apolipoprotein B-100 Low-Density Lipoproteins, Apolipoprotein B-48 Remnants, and Apolipoprotein E-Rich High-Density-Lipoproteins in the Mouse. Journal of Lipid Research. 35, 1297-1310 (1994).
- Fagan, A. M., et al. Unique lipoproteins secreted by primary astrocytes from wild type, apoE (-/-), and human apoE transgenic mice. Journal of Biological Chemistry. 274, 30001-30007 (1999).
- Musliner, T. A., et al. Dissociation of high density lipoprotein precursors from apolipoprotein B-containing lipoproteins in the presence of unesterified fatty acids and a source of apolipoprotein A-I. J Lipid Res. 32, 917-933 (1991).
- Silverman, L., Glick, D. The reactivity and staining of tissue proteins with phosphotungstic acid. J Cell Biol. 40, 761-767 (1969).
- Hamilton, R. L., Williams, M. C., Fielding, C. J., Havel, R. J. Discoidal bilayer structure of nascent high density lipoproteins from perfused rat liver. J Clin Invest. 58, 667-680 (1976).
- Pollard, H., Scanu, A. M., Taylor, E. W. On the geometrical arrangement of the protein subunits of human serum low-density lipoprotein: evidence for a dodecahedral model. Proc Natl Acad Sci U S A. 64, 304-310 (1969).
- Frasca, J. M., Parks, V. R. A Routine Technique for Double-Staining Ultrathin Sections Using Uranyl and Lead Salts. J Cell Biol. 25, 157-161 (1965).
- Zhao, F. Q., Craig, R. Capturing time-resolved changes in molecular structure by negative staining. Journal of structural biology. 141, 43-52 (2003).
- Knight, D. P. Negative staining of rat tail tendon collagen fibrils with uranyl formate. Tissue Cell. 7, 651-654 (1975).
- Dash, S., et al. Temperature programmed decomposition of uranyl nitrate hexahydrate. Journal of Nuclear Materials. 264, 271-282 (1999).
- Zhang, L., et al. Structural basis of transfer between lipoproteins by cholesteryl ester transfer protein. Nature Chemical Biology. 8, 342-349 (2012).
- Kartha, G. Combination of multiple isomorphous replacement and anomalous dispersion data for protein structure determination. 3. Refinement of heavy atom positions by the least-squares method. Acta crystallographica. 19, 883-885 (1965).
- Kartha, G., Parthasarathy, R. Combination of Multiple Isomorphous Replacement and Anomalous Dispersion Data for Protein Structure Determination. I. Determination of Heavy-Atom Positions in Protein Derivatives. Acta crystallographica. 18, 745-749 (1965).
- Kartha, G., Parthasarathy, R. Combination of Multiple Isomorphous Replacement and Anomalous Dispersion Data for Protein Structure Determination. Ii. Correlation of the Heavy-Atom Positions in Different Isomorphous Protein Crystals. Acta crystallographica. 18, 749-753 (1965).
- Taylor, K. A., Glaeser, R. M. Electron microscopy of frozen hydrated biological specimens. Journal of ultrastructure research. 55, 448-456 (1976).
- Correia, I., et al. The structure of dual-variable-domain immunoglobulin molecules alone and bound to antigen. mAbs. 5, (2013).
- Cardone, G., Heymann, J. B., Steven, A. C. One number does not fit all: mapping local variations in resolution in cryo-EM reconstructions. Journal of structural biology. 184, 226-236 (2013).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados