
Method Article
Генетические манипуляции
В этой статье
Резюме
Мы сообщаем способ использования типа I и типа II Δku80 Штаммов Токсоплазма Эффективно генерировать целевой ген делеции и замены генов для функционального геномного анализа.
Аннотация
Целевые генетической манипуляции с использованием гомологичной рекомбинации методом выбора для функционального геномного анализа, чтобы получить детальное представление о функции генов и фенотипа (ов). Развитие мутантные штаммы с целевыми делеции гена, целевые мутации, дополненные функции гена и / или на ярлыке генов обеспечивает мощный стратегии для решения функции генов, особенно если эти генетических манипуляций могут быть эффективно ориентированы на локус гена интереса с помощью интегрирования опосредованного двойной пересечь гомологичной рекомбинации.
В связи с очень высокими темпами негомологичного рекомбинации, функциональный геномный анализ Токсоплазма ранее была ограничена отсутствием эффективных методов генной ориентации делеции и замены генов конкретных генетических локусов. В последнее время мы отменили основной путь негомологичных рекомбинации в тип I и тип II штаммов T. гондий, удалив ген encodinг Ku80 белка 1,2. Δku80 штаммы ведут себя нормально в течение тахизоит (острый) и брадизоитного (хронический) этапов в пробирке и в естественных условиях и проявляют по существу 100% частота гомологичной рекомбинации. Δ Ku80 штаммов сделать функциональные геномные исследования возможно на одном гене, а также на геном шкале 1-4.
Здесь мы сообщаем способы использования типа I и типа II Δku80Δhxgprt штаммов для продвижения генного таргетинга подходы в Т. гондий. Мы выделяем эффективных методов генерации генной делеции гена замен, и отмеченных генов целевых вставка или удаление гипоксантин ксантин-гуанинфосфорибозилтрансферазы (HXGPRT) селективный маркер. Описанные целенаправленное изменение генов протокол может быть использован в различных способов Δku80 штаммов для продвижения функционального анализа генома паразита и разработать один штаммы, несущиенесколько целевых генетических манипуляций. Применение этого метода генетических и фенотипических последующие анализы покажут фундаментальные и уникальные аспекты биологии Т. гондий и связанных с ними значительных человеческих патогенов, вызывающих малярию (Plasmodium Sp.) и криптоспоридиоз (Cryptosporidium).
Введение
Токсоплазма является общей облигатными внутриклеточными паразитами простейшими, что часто и длительно заражает широкий спектр животных и человека 5. Подсчитано, что более 1 миллиарда людей в настоящее время и хронически инфицированных этим возбудителем. В дополнение к важности заболеваний, вызванных T. гондий инфекции, повышение доступности инструментов экспериментального, мощные ресурсы геномики 6, простота в пробирке рост и отличные модели мыши сделали Т. гондий ведущей моделью системы для более широкого изучения внутриклеточных патогенов эукариотических и других значимых apicomplexan паразитов, которые вызывают тяжелых заболеваний, таких как малярия (Plasmodium Sp.) и Криптоспоридиоз (Cryptosporidium) 5,7. Существенным недостатком Токсоплазма в качестве модельного организма была неэффективной восстановления потомства, продающие целевых генетических манипуляций. Это проблем в генного таргетинга связано с низкой частотой гомологичной рекомбинации по отношению к очень высокой частоте рекомбинации в негомологичного штаммы дикого типа Т. гондий даже при обширных гомологии ДНК содержится в ДНК-мишени молекул, используемых в генетических исследованиях 2.
Недавно мы генетически блокировали основной путь негомологичной рекомбинации в тип I и тип II штаммов Токсоплазма путем удаления гена, кодирующего белок Ku80 1,2. В результате я типа и типа II Δ Ku80 штаммы проявляют нормальные темпы роста, размер и поведение как в пробирке и в естественных условиях в течение тахизоит и брадизоитного этапов в пробирке и в естественных условиях, однако, эти штаммы обладают по существу 100% частота гомологичной рекомбинации и это фенотип увеличивает вероятность быстрого выделения желаемого потомства целевых генетических манипуляций нескольких сотен до нескольких ТхоUSand раза 1,2,4. Последние всей общины, использование типа I и типа II Δ Ku80 штаммы существенно увеличили темп, разнообразие, а также вероятность успеха целевые генетические подходы в Т. гондий 1-4,8-13. Здесь мы опишем всеобъемлющего протокола для целевого гена удаления, замены генов и генных пометки в Δ Ku80 штаммов T. гондий. Мы показываем, как проектировать и надежно целевых генетических манипуляций конкретных генов или генетических локусов в Δ Ku80 штаммов Токсоплазма.
протокол
1. Генного таргетинга Удаление гена
Этот протокол предназначен для эффективной генерации мутантного штамма, который имеет целевой удаление гена в определенном генетическом локусе. Ранее описанные Т. гондий гипоксантин ксантин-гуанинфосфорибозилтрансферазы (HXGPRT) селективного маркера используется в данном протоколе для безопасного и надежного маркера вставки после микофенолокислота и ксантин (MPA + X) отбор 14-16. Этот протокол использует проверенную и экономически эффективный способ построения ДНК ориентации молекул на основе дрожжей рекомбинационная клонирования 17,18. Некоторые альтернативные протоколы являются коммерчески доступными для построения рекомбинантных молекул, имеющих мишень с использованием бактериальной рекомбинационной методами клонирования в пробирке рекомбинационной методами клонирования, или сплав ПЦР. Протокол, описанный ниже обеспечивает самую высокую общую эффективность и надежность восстановления клонированного пунктыITES обладающих целевой делеция гена в Δ Ku80 штаммов T. гондий.
1.1 Подготовка улавливающей ДНК
- Доступ ToxoDB 6 ( http://www.toxodb.org ), чтобы получить геномных последовательностей для представляющего интерес гена (ПИ).
Примечание: геномные последовательности должны быть получены от типа I, II или III генома в ToxoDB базу данных, которая находится ближе всего к деформации манипулируют. Например: GT1 для RH и ME49 для Пру.
- Дизайн специфических праймеров перекрытия, которые усиливают 5 'и 3' геномной ориентации склонов интерес ген (ПИ), включающий 33 п.н. перекрытия с дрожжевой челночный вектор pRS416 (или альтернативно pRS426) и включают 33 п.н. перекрытия с селективным маркером ( HXGPRT) (фиг.1А-В). Добавить уникальный сайт рестрикционного фермента в каждый праймер на обоих концах 5 'Nd 3 'геномной таргетинга флангах, помещается между перекрытием 33 б.п., а Т. гондий конкретной последовательности праймера (ов) (фиг.1А-В).
Примечание: более 30 перекрытия п.о. необходим для эффективной рекомбинации клонирование дрожжей. 5 'и 3' геномной ориентации бокам предназначены для амплификации специфических фрагментов ДНК от 800 до 1400 пар оснований, которые определяют целевой удаления. Имейте в виду, что более короткие флангах позволит значительно снизить эффективность адресности в Δ Δ Ku80 hxgprt фоне 2.
- ПЦР усиливают 5 'целевой фланг с использованием праймеров F1 и R1, и ПЦР усиливают 3'-мишень фланг с использованием праймера F2 и R2 (фиг.1А-С), из геномной ДНК 3. Отдельно ПЦР усиливают ~ 2 т.п.н. HXPGRT ген, включая соответствующие 5 'и 3' фланкирующие области DHFR из HXGPRT кДНК pminiHXGPRT кассета 14 с использованием праймеров HXF и HXR(Фиг.1В-C).
Обратите внимание: Усиление целевой бока с использованием геномной ДНК из родительского штамма для трансфекции.
Примечание: Место HXGPRT маркер в прямом ориентации по отношению к 5 'и 3' ДНК ориентации бока чтобы облегчить последующее эффективное генетических манипуляций, таких как целевые удаление HXGPRT с нарушенной локуса.
- Проверьте правильность размеров продукта ПЦР с помощью электрофореза в агарозном геле и оценить концентрацию ДНК-фрагмент использованием агарозном геле стандартов или другой способ определения концентрации ДНК.
- Создание компетентных аликвотам дрожжей, как описано в Gietz и Schiestly 17.
- Объединение 50 нг 5 'геномной ориентации фланг, 50 нг 3' геномной ориентации фланг, 100 нг маркер селекции HXGPRT и 50 нг линеаризованной челночный вектор стерильной H 2 O для достижения конечного объема 10 -20 мкл. Трансформации компетентных дрожжи с три ПЦР-продуктов и дрожжей E. палочка челночный вектор для клонирования рекомбинационная использованием протокола описывается Gietz и Schiestly 17. Пластина трансформированных дрожжей на урацил-минус чашках с минимальной агаризованной средой и инкубируют при 30 ° С в течение 2 - 3 дней.
Примечание: заказать сборку плазмиды, 5 'целевой фланг, маркер HXGPRT и 3' целевой фланге способствует инженерии 33 б.п. перекрытия между фрагментами ДНК (рис. 1A-B) и опосредовано гомологичной рекомбинации в дрожжей.
- Урожай дрожжи, добавив 2 мл 2x YPAD на урацил-минус пластин и осторожно выскабливание, чтобы сместить колоний, не нарушая агара. Центрифуга дрожжевой раствор в течение 5 мин при 5000 х г и отбросить супернатант.
- Ресуспендируют осадок дрожжей в 250 мкл буфера, содержащего ресуспендирования РНКазы А и добавляют 50 - 100 мклCID промытые стеклянные шарики в раствор. Vortex в течение 5 минут на самой высокой скорости, чтобы разбить клетках дрожжей.
- Разрешить стеклянные шарики оседают на дно пробирки и передачи супернатант в 2 мл пробирку Эппендорфа.
- Изолят дрожжей ДНК с использованием ДНК изоляции минипрепаративных комплект, ресуспендирования в конечном объеме ~ 100 мкл.
- Смешайте 2 мкл ДНК дрожжей (~ 50 нг) с 40 мкл DH10B электропорации компетентных E. палочка держится на льду.
- Передача раствора в охлажденной 2 мм кювету для электропорации разрыв и электропорации клетки бактерий при 2,4 кВ и 129 Ω.
- Спасательная клетки путем добавления 0,8 мл бульона SOC в каждую кювету и передачи решения 14 мл пробирку с защелкивающейся крышкой Сокола. Клетки инкубируют при 37 ° С на каток в течение 40 - 60 мин.
- Пластина E. палочки на 2ХУТ + ампициллин (АМФ) пластины и инкубировать пластин при 37 ° С в течение ночи.
- Выбор ~ 6 - 8 отдельных колоний и расти в 3 мл 2ХУТ + AMP в течение ~ 16 часов при 37 ° С.
- Подготовьте 30% запаса глицерина морозильник каждого E. палочка клона.
- Выделяют плазмиду pΔGOI из E. кишечной клонов с использованием набора минипрепаративных, ресуспендирования в конечном объеме ~ 100 мкл.
- Подтвердить pΔGOI использованием рестриктазы дайджесты для измерения ДНК плазмиды размером и проверить ожидаемое ДНК pΔGOI исчерченности.
- Завершение проверки pΔGOI секвенированием ДНК 5 'и 3' геномной ориентации бока, чтобы проверить 100% ДНК последовательности на основе геномной последовательности данных в http://www.ToxoDB.org .
Примечание: Около прекрасного гомологии последовательности необходимо для максимизации эффективности генного таргетинга 3, так как каждая пара оснований разница представляет собой соответствующее сокращение длины идеально гомологии.
Примечание: последовательность генома типа II Δku80 штамм на основе родительских ул Прудождь не доступна в данный момент. Эта работа использует тип II ME49 последовательность генома, поскольку суррогатная генома для типа II штамма Δ Ku80. На основании данных о последовательности на количество генетических локусов, предполагается, что ME49 генома и Pru геном обладают полиморфизмы одного нуклеотида на частоте ~ 1 на 10000 нуклеотидов или менее.
- Создать> 200 мкг со pΔGOI путем посева ~ 250 мл 2ХУТ + AMP ночной культуры с глицерином акции о проверенной E. Клон минипрепаративных палочка.
- Изолят pΔGOI плазмидной ДНК из большой культуры с использованием изоляции Maxiprep ДНК комплекта ресуспендирования в конечном объеме ~ 1000 мкл.
- Повтор рестриктазой дайджесты, или секвенирования ДНК pΔGOI Maxiprep ДНК, чтобы проверить правильность ориентации молекулы ДНК до трансфекции.
- Линеаризовать ~ 15 мкг pΔGOI на 5'-конце использованием уникального рестрикционного фермента Х (RE.X)Дайджест сайт построен в 5 'целевой фланг (рис. 1В).
Примечание: генного таргетинга в Т. гондий требует минимум ~ 10 мкг улавливающей ДНК для получения эффективной частоты ориентации событий на генном локусе интерес 2.
- Подтвердить линеаризации pΔGOI и определяют концентрацию ДНК в агарозном геле или альтернативный метод.
Примечание: Если ограничения ферментативного расщепления на 5 'фланге не дает полной линеаризации, разработаны 3' уникальный RE.Y дайджест сайта можно использовать в качестве альтернативного места для линеаризации pΔGOI.
Примечание: полностью линеаризованы ориентации молекулы ДНК имеет важное значение для успешного генного таргетинга путем гомологичной рекомбинации в Δ штаммов Ku80.
- Нейтрализовать ограничение фермента incubatния при 68 ° С в течение 15 - 20 мин.
- Подготовьте по крайней мере 15 мкг линеаризованной плазмиды в 100 мкл стерильной H 2 O. Магазин линеаризованной pΔGOI при -20 ° С до времени трансфекции.
1.2 Подготовка Т. гондий паразитов, трансфекции, отбор, субклонировании, проверки, архивных и поддержания генетически модифицированных штаммов
Общие методы культуры и манипулирование T. гондий в фибробласта крайней плоти человека (HFF) клетки описаны 19-21. Δ Δ Ku80 hxgprt штаммов репликации обычно в паразитом инфекции среды (орлов минимальной поддерживающей среде (ЕМЕМ) питательной среде с добавлением 1% фетальной бычьей сыворотки (FBS) и 1%-противогрибковые антибиотики разбавляют с 100x акции) 1,2. Вся работа с Токсоплазма должна быть выполнена с использованием уровня биобезопасности 2 процедуры. Т. гондий RHΔ Ku80 2 пареntal штамм был сгенерирован из штамма RHΔ hxgprt от Роос Лаборатории 14. PruΔku80 один родительский штамм был сформирован из PruΔhxgprt (Pru gniaud штамм BSG-4, 22), который содержит стабильно интегрированную CAT селектируемый маркер и зеленый флуоресцентный белок (GFP) репортер под контролем брадизоитного этап LDH2 конкретного промотора.
- Привить 25 см 2 колбы, содержащей клетки HFF сливающийся с 1 х 10 6 жизнеспособных типа RH Я Δ Δ Ku80 hxgprt паразиты, или привить два 25 см 2 колбы с 2 х 10 6 жизнеспособных типа II Prugniaud (Пру) Δ Δ Ku80 hxgprt паразитов.
Примечание: жизнеспособных паразитов определяются как внеклеточных паразитов, которые недавно лизировались вне.
Примечание: Эта шкала обеспечивает достаточное количество жизнеспособных паразитов для трех экспериментов трансфекции.
- Осмотрите Δ Δ Ku80 hxgprt Инфицированные культуры ~ 68 - 72 часа после заражения. Убедитесь в наличии жизнеспособных паразитов и ~ 90 - 95% лизис клетки HFF планировать точные сроки трансфекции.
Примечание: изоляция недавно egressed паразитов имеет важное значение для успеха этого протокола, так как низкая жизнеспособность паразита отменят ген-таргетинга эффективности.
- Перемешивают жизнеспособных паразитов в раствор путем закрытия плагин уплотнение крышки на 25 см2 колбу и энергично встряхивать колбу назад и вперед для перемешивания паразитов от ростовой поверхности без разбрызгивания жидкости на внутренней стороне крышки или горло колбы .
Примечание: паразитов урожай не требует шприца игла выпуска протокол для типа I илитип II Δ Ku80 штаммов.
- Передача паразита раствора в 10 мл шприца прикреплен к держателю фильтра, содержащих 3 мкм мембрану и поместите фильтр в верхней части открытого 15 мл завинчивающейся крышкой трубки. Шприцевой фильтр паразитов в 15 мл с завинчивающейся крышкой трубки.
Примечание: фильтрация паразитов через мембрану удаляет инфицированных клеток и клеточного дебриса.
- Определение концентрации паразитов (тахизоит формы на мл) с использованием гемоцитометра.
Примечание: Дополнительно: Использование паразита решения, создана бляшкообразующие блока (PFU) анализа 21, который будет прочитан 7 - 8 дней, чтобы определить адекватную жизнеспособность трансфицированным паразиты (ОРП тахизоит отношением ≥ 0.2).
- Гранула паразитов в течение 7 минут при 1400 х г и супернатант до ~ 0,4 мл, не нарушая паразита гранул. Вращаться в течение 2 мин при 1,400 XG и аспирации супернатант оставшееся до ~ 0,01 мл, не нарушая паразита гранул.
- Ресуспендируют осадок паразита, щелкая нижней части трубы и сразу же добавить cytomix 23 буфера (1.33x концентрация) для паразита гранул для получения паразита концентрации 4 - 5 × 10 7 паразитов / мл cytomix.
Примечание: пропустить добавлением АТФ и глутатиона cytomix поскольку эти дополнения не повышение эффективности генного таргетинга.
- Разморозить 100 мкл аликвоты линеаризованному pΔGOI из протокола 1.1 (шаг 26).
- Передача 0,3 мл раствора паразита cytomix (~ 1,3 - 1,6 · 10 7 паразитов) в 100 мкл линеаризованной pΔGOI с протоколом 1,1 (шаг 26), и немедленно передать всю 0,4 мл паразит + pΔGOI смесь к охлажденному 2 мм зазор кювету для электропорации.
- Электропорации паразитов на 1,4 кВ и 24 Ω.
- Resт трансфекции кювете при комнатной температуре в течение ~ 5 мин, затем передавать все содержимое трансфекции кювету в 150 см 2 колбу, содержащую сливной клетки HFF с 30 средних инфекции мл и инкубируют инфицированной культуры в течение ночи.
- Начать выбор в микофеноловая кислота + ксантина (MPA + Х) ~ 20 часов после трансфекции (фиг. 2A), заменив инфекции среде с MPA + X селективной среде (MPA (25 мкг / мл) и ксантина (50 мкг / мл) в инфекции среды).
Примечание: Не беспокоить инкубации MPA + X выбор колбу.
- Оценить общее количество блоков PFU в MPA + X выбор колбу ~ 8 дней после трансфекции путем визуального осмотра монослоя. Убедитесь в наличии разработке блоков PFU и здоровой зоны инфекции с помощью световой микроскопии.
Примечание: Если бляшки не видны днем 8, сохраняют выбор и следить за признаками заразитьиона, так как мутантные штаммы могут иметь льготный тариф репликации.
Примечание: Δ Δ Ku80 hxgprt штаммов паразита имеют чрезвычайно низкую фоне нежелательных нецелевые событий, что позволяет для успешного выделения мутантных штаммов с довольно тяжелой, но не смертельной дефекты роста. Если ген имеет важное значение и она не может быть удалена, первичный трансфекции или впоследствии передается паразит населения, может выявить фенотип, где паразитов прекращают репликацию в процессе выбора, поскольку население решает целевых нокаутом.
- Встряхните колбу как в шаге 3, чтобы выбить внеклеточных паразитов из монослоя [после видимого бляшек разработали] для генерации паразита решение. Передача ~ 0,5 мл паразита раствор из MPA + X выбор колбу на 25 см 2 MPA + X выбор колбу, содержащую сливной клетки HFF (назначают этой культуры проход 1А).
- Выбить паразитаы в оригинальной 150 см 2 МПа + X выбор колбу ~ 3 дня спустя и передачи ~ 0,5 мл паразита решение второй 25 см 2 МПа + X выбор колбу, содержащую клетки HFF сливающийся (1B обозначения этой культуры проход).
- Разрешить зонах инфекции развиваться в проходе 1А и / или пройти 1B для колб ~ 5 дней. Визуально проверить с помощью световой микроскопии, что проход 1А и 1В колб содержать минимум 25 жизнеспособных блоков PFU со здоровыми зон заражения.
- Выберите либо 1А проход или колбе проход 1B для продолжения проход в MPA + X селективную среду в 25 см 2 колбы. Поддерживать проход под MPA выбор + X.
Примечание: Паразиты должны быть выращены в течение достаточного числа поколений, чтобы удалить неинтегрированных сохраняющейся эписом из популяции до субклонирования. Не выполняйте субклонировании до 20 дней после трансфекции для типа RH Я Δ Δ Ku80 hxgprt, или до 25 дней после трансфекции дляТип II Пру Δ Δ Ku80 hxgprt паразитов чтобы было достаточно времени, чтобы разбавить неинтегрированные эписом 1,2.
- Субклон паразитов в 96-луночный лоток, содержащий сливной клетки HFF с MPA + селективной среде X. Подготовьте один лоток в концентрации 1 паразита / лунку и второй лоток при концентрации 2 паразитов / лунку.
Примечание: Субклон паразитов, которые недавно лизируют (~ 90 - 95% от HFF клеток в 25 см 2 колбы) для того, чтобы паразиты имеют высокую жизнеспособность.
Примечание: оптимальный срок после трансфекции для субклонирования была создана путем измерения, как быстро успешно целевых паразитов возникают при высокой частоте в выбранном населения 1.
- Продолжайте прохождение первичного населения трансфицирован паразитов в MPA + селективной среде X Использование еженедельный график прохождения заражение25 см 2 HFF колбу с 10 мкл паразита раствора.
Примечание: если клонов проведение целевых делеции гена (нокаутом) не получается из первого субклонирования в шаге 18, resubclone паразитов из непрерывно поддерживается население ~ 10 - 12 недель после трансфекции.
Примечание: Одна из причин, делеция гена не получено возникает из эписомными сохранением некоторых плазмид pΔGOI. Эписомными стойкость определяется последовательностей, содержащихся в 5'-и 3'-геномной ориентации боках и видимому, не возникают из HXGPRT генетического элемента. Около 5 - 10% от ориентации плазмиды обладают значительной эписомными стойкость, которая требует более позднее время субклонирования, чтобы позволить паразита населения достаточным количеством поколения раз, чтобы разбавить сохраняющейся эписом и решать население в основном стабильны интегрантов.
- Оценка 96-луночный лоток 6 - 7 дней после субклонирования (тип I паразитов) или 7 - 8 дней после субклонирования (тип II паразитов) для лунок, содержащих одну PFU, определены как одну зону инфекции в световой микроскопии при либо 40X или 60X власти. Отметить небольшой точкой на 96-луночных лоток крышкой для обозначения расположения PFU в скважине.
- Смешайте содержимое каждой лунки, содержащие один PFU с помощью пипетки установлен в 50 мкл (200 мкл наконечник) путем направления потока текучей среды по PFU разойтись паразитов в скважине.
Примечание: Смешивание также ускоряет паразита лизис монослой HFF. Тип I RH штаммов растворит хорошо ~ 4 дня после смешивания, тип II Пру паразитов растворит хорошо ~ 5 дней после смешивания.
- Выбор десятков лизировали скважины (клоны) и царапин в нижней части каждого из этих лизированных скважин с использованием 0,5 - 10 мкл кончиком пипетки одновременно составление 6 мкл раствора паразита. ТрансТЭР 6 мкл паразита решений для хорошо известных в 24-и сливной лоток, содержащий клетки HFF в 1 мл MPA + X селективной среде.
Примечание: Важно, чтобы царапать нижней части так, чтобы держать отверстие отверстие наконечника пипетки в постоянном контакте с паразитами, проживающих в нижней части скважины.
- Монитор 24-луночных лоток для лизиса клетки HFF.
Примечание: тип I RH паразиты, как правило, хорошо растворяя в 24-луночный формат в ~ 4 дней. Тип II Пру паразиты, как правило, хорошо растворяя в ~ 5 дней.
- Передача 2 мкл эффективным решением паразита поцарапаны от нижней части каждой лунки в 24-луночный планшет в соответствующую лунку нового 24-луночных сливной лоток, содержащий клетки HFF и 1 мл MPA + X селективной среде на лунку.
Примечание: Все паразиты клонов и линий можно непрерывно основныхполученные путем пропускания 2 мкл жизнеспособным решением паразита каждые 7 дней в этой 24-и лоток формата.
- Убедитесь, что паразиты были успешно перенесены на новый 24-а лоток визуальном осмотре световой микроскопии ~ 18 ч после прохода.
- Урожай паразитов из лизированных лунки 24-луночного лоток с шага 23 - 24 с использованием либо 1 мл или 10 мл пипетки и перенос паразита раствор в пробирку Эппендорфа.
- Гранул паразитов при 1400 х г в течение 7 мин, промывают один раз в 1 мл забуференного фосфатом физиологического раствора (PBS), и осадок снова при 1400 х г в течение 3 мин.
- Aspirate PBS от осадка, затем добавьте 200 мкл PBS к осадку, чтобы ресуспендировать паразитов. Зафиксировать паразита раствор при -80 ° С до выделения ДНК.
- Изолировать паразита ДНК из каждого клона использованием изоляции ткани ДНК MINIKIT.
- Проверка целевого удаление гена (нокаут) методом ПЦР с использованием праймеров проверки (Фиг.2А-С). ИспытаниеОтсутствие функциональной кодирующей области гена и использование CxF вверх по течению и ниже по течению CXR геномной ДНК праймеров (фигуры 2A-B), тест на наличие ПЦР-продукты, которые охватывают геномной ориентации боках и HXGPRT описывает правильное 5'-и 3' "направленной интеграции в HXGPRT гена в локусе гена удален.
Примечание: Праймеры для проверки должен быть уникальным для представляющего интерес гена или ложный положительный результат может быть получен. Грунтовка дизайн может быть утверждена http://www.toxodb.org взрывным последовательности праймеров к геному (ы), чтобы проверить праймеров являются уникальными.
- Прохождение паразита клонов на недельном графике как описано в пункте 24. После удаления целевых подтверждена, продолжается отбор в MPA + X не является обязательным.
- Архив клонов паразита путем подготовки морозильник запасов. Гранула внеклеточных паразитовиз лизированных клеток HFF в 25 см 2 колбы культуры, средств массовой информации и удалить аккуратно ресуспендируйте паразита осадок в среде клеточной культуры при замораживании паразит концентрации> 4 х 10 7 паразитов на мл. Передача аликвоты и хранят криопробирок паразитов до бесконечности в жидкий азот, или при -80 ° С.
2. Удаление HXGPRT
Этот протокол предназначен для удаления HXGPRT маркер от сайта интеграции в геноме генетически модифицированных Δ Ku80 штамма. Удаление HXGPRT позволяет извлекать маркер и генерации штаммов с множественной генетических манипуляций с использованием только выбор на основе HXGPRT 1-3. В то время как протокол подробно описаны ниже описывается метод повторной целевой локус, чтобы удалить HXGPRT, следует отметить, что удаление HXGPRT на генном локусе интерес также позволяет одновременно повторной интеграции гена дикого типа (сomplementation), повторное интегрирование мутантный ген, а также повторное интегрирование меченый ген (N-или С-концевых GFP, HA тег, и т.д.,), что позволяет для различных генетических манипуляций. Механизм действия 24 и ориентации протоколов с использованием 6-тиоксантина выбора были ранее описаны 1-3,15.
2.1 Удалить HXGPRT
- Акцизный HXGPRT селективный маркер от pΔGOI расщеплением с RE.Z чтобы разрезать по уникальной сайты рестрикции фланговые HXGPRT (рис. 1В).
- Проверка полное переваривание ДНК с помощью электрофореза в агарозном геле. Изолировать большей из двух полос из геля агарозы.
Примечание: больший из двух полос содержит плазмиду вместе с 5 'и 3' бока ДНК-мишени, но не HXGPRT гена.
- Поместите разделе агарозном содержащих более значительные полосы ДНК в Layin колонке спинаг геля плоский на мембране. Спин колонку при 13000 х г в течение 4 мин при комнатной температуре. Добавить стерильной Н2О в проточном, чтобы довести объем до 100 мкл.
Примечание: другие коммерческие методов для выделения ДНК из агарозы.
- Концентрат ДНК путем осаждения этанолом, ресуспендирования ДНК в конечном объеме 18 мкл стерильной H 2 O.
- Смешать 1 мкл концентрированной ДНК с 7 мл H 2 O, 1 мкл 10х буфера лигирования и 1 мкл Т4 ДНК-лигазы (5 единиц) и поместить реакции при 4 ° С в течение ночи для получения pΔGOIc (pΔGOIclean) (фиг. 3А).
Примечание: фрагменты ДНК с RE.Z содержащие концы ДНК могут быть разработаны и включены в это шаг, чтобы создать плазмиды подходит для целевого повторной интеграции гена дикого типа (дополнения), целевые повторной интеграции мутантный ген, а также целевые реинтеграции отмеченных гена(N-или С-концевых GFP, HA тег и т.д.).
- Разбавляют реакционную 2x стерильной Н2О до электропорации.
- Преобразование pΔGOIc в E. палочки, как в протоколе 1,1 субклонировать, изолировать и проверять адресности плазмиды. Тогда линеаризуем pΔGOIc в рамках подготовки к трансфекции (см. шаги 1.1.11 к 1.1.26).
- Повторите паразита протокол трансфекции, как указано в протоколе 1.2 (шаги с 1 по 11).
Примечание: Перед проведением шаги с 1 по 8, убедитесь, что целевые удаление HXGPRT возможно в мутантный штамм. Заразить 150 см 2 колбы вырожденные клетки HFF 1 × 10 6 тахизоиты мутантного штамма с использованием 6-тиоксантин (6TX) Выбор среды (200 мкг / мл 6TX инфекции в среде) и колбу помещают в анализе PFU. Осмотрите колбу 8 дней, чтобы убедиться, что нет (или очень мало; <10) PFU не видно, что очень важно установить, что потенциалпространственной ориентации генных эффективность превысит любые потенциальные спонтанной реверсии мутантного штамма к фенотипу с пониженной экспрессией HXGPRT (6TX фенотип сопротивление).
Примечание: примерно 5 - 10% сайтов HXGPRT интеграции проявляют значительный и спонтанной частоты 6TX сопротивления хотя HXGPRT маркер еще интегрированы на целевом сайте (см. раздел Представитель Результаты пояснения).
- Начать 6TX выбор ~ 20 часов после трансфекции путем изменения среды в 150 см 2 колбу 6TX селективной среде.
Примечание: Не беспокоить колбу в течение ~ 10 дней после трансфекции.
- Осмотрите колбу для формирования PFU на 10 день - 12 после трансфекции (тип I паразитов) или 10-й день - 16 после трансфекции (тип II паразитов).
Примечание: ParasiTES мишенью для удаления HXGPRT не начнет расти под 6TX выбора, пока HXGPRT генов и мРНК не будет удалено, а остаточный белок HXGPRT инактивируется.
- Встряхнуть выбор колбу создать паразита раствора и передачи 0,5 - 1,0 мл паразита решения новой 25 см2 колбу, содержащую сливной клетки HFF и 5 мл 6TX селективной среде. Повтор передачи от первичного 150 см 2 до нового 25 см 2 колбы один раз в день в течение еще двух дней.
Примечание: Несколько выборки повышает шансы на захват целевых жизнеспособные паразиты, которые потеряли HXGPRT гена.
- Проверьте 25 см 2 колбы ~ 5 дней после заражения, чтобы проверить наличие разработке блоков PFU с зонами здоровых репликации паразитов. Выбор одного или двух колбах, содержащих зоны инфекции.
Примечание: Паразиты удален из дублирующих генов HXGPRT с нормальной скоростью роста 6TX выбора.
- Продолжайте прохождении паразитов в 6TX селективной среде.
- Субклоном населения паразита через 25 - 30 дней после определения. Установить с 96 лунками, поднос с ~ 1 паразита / лунку, а другой с ~ 2 паразитов / лунку.
- Подготовка паразита ДНК из выделенных клонов и клонов поддерживать в 24-луночный планшет культуры в соответствии с протоколом 1,2 (шаги 19 до 29).
- Подтвердить удаление HXGPRT помощью ПЦР с использованием стратегии, изложенной на фиг.3А-B.
3. С-концевой Мечение белков
Этот протокол предназначен для С-концевого мечения белков путем воздействия на тег для интеграции через двойной крест над гомологичной рекомбинации в геномного локуса гена использованием MPA + X выбор 2,4. Этот протокол работает эффективно, потому что 5'-DHFR последовательность HXGPRT маркером является полностью проверены функциональные 3 'нетранслируемой области для других генов 2,4.
3.1 Прямое С-концевого мечения белков при эндогенных генетических локусов
- Создать таргетинга pΔGOItag плазмиды построить использованием дрожжей рекомбинационная клонирования (рис. 4) и методы, описанные в протоколе 1.1. 5 'геномной ориентации фланг содержит последние 800 до 1200 пар оснований кодирующей области (или геномной ДНК) ПИ, за исключением кодона терминации перемещается в положение 3' к тегу выбора (HA тег, Myc тег Его тег и т.д.)
- Создание целевого введения C-концевой метки с эндогенным локусом белок-кодирующих генов, следуя инструкциям в протоколе 1.1 и 1.2 с использованием стратегии, изложенные (рис. 4).
- Убедитесь, что вставка С-концевой метки на эндогенный локус гена с использованием стратегии ПЦР.
- Retarget локуса гена использованием методов, описанных в протоколом 1 и 2 до Delete HXGPRT селективный маркер для создания точно регулировать эндогенный локус гена, который выражает меченый белок. Этот протокол также восстанавливает HXGPRT селектируемый маркер, который может быть использован повторно целевой другом локусе помеченный деформации (см. протокол 1).
Результаты
Подробный шаблон предоставляется для построения ориентации плазмида, чтобы удалить ген, включая размещение сайты рестрикции и генерации праймеров, которые облегчают генетический ориентации и проверки целенаправленное изменение генов, а также конструкции плазмиды для последующего удаления HXGPRT в пошаговый процесс (рис. 1A-C, 2А, 3А). Общая схема представлена для изготовления целевого удаление гена (фиг. 2А), пар праймеров используется для проверки удаление нокаут, например, типа I rop18 (2В), а репрезентативные результаты проверки ПЦР показаны (фиг. 2С). Этот представитель Результат показан на иллюстрации диапазон результатов, которые могут быть получены при генетических локусов, которые относительно трудно цели. Успешно целевых делеции гена приведет к отсутствию гена винтерес продукта ПЦР (PCR 1), присутствие 3'геномной ориентации фланг (ПЦР2), а также наличие HXGPRT селектируемый маркер образом интегрированы между 5 'и 3' геномной ориентации бока, которые определяют удаления (PCR3 и pCR4) . Клоны, 3, 4, 5, 6, 8, 9 и 11, проверяются делеции гена-мишени (заглушек) где HXGPRT заменил ПИ (фиг. 2С).
Клон, который не содержит делеции гена может быть представлен различными исчерченности. Как правило, клон без делеции гена будет отражать родительским штаммом (родительский шаблон) с полосы, наблюдаемые для ПЦР1 и ПЦР2, но не для PCR3 или pCR4, как показано на клоны, 1 и 2 (фиг. 2С). Это «Родительский шаблон" возникает от нескольких потенциальных механизмов. Иногда, MPA устойчивостью отдельных паразитов нести неинтегрированные сохраняющиеся эписом из pΔGOI таргетинга плазмиды, и отметим, что продолжение выбора часто вынуждает этих эписоминтегрироваться. Отметим также некоторые редкие фона в типа I Δ Ku80 деформации из-за непреднамеренного интеграции pΔGOI ориентации плазмиду, содержащую кДНК HXGPRT выражается через DHFR 5 'и 3' промоторные элементы, либо в локусе DHFR или в частично удален HXGPRT локус 14. Хотя это редкое событие является целевой интеграции, это не было предназначено интеграции и служит Важно помнить при разработке любой стратегии замены генов. Гомологии ДНК более 120 б.п. осуществляется на pΔGOI таргетинга плазмиды может обеспечить альтернативное место для рекомбинации в Δ Ku80 2 штаммов, которые могут отдать нежелательного фона. В типе II Pru Δ Ku80 штамм этом фоне снижается или отсутствует по сравнению с типа I, поскольку HXGPRT селектируемого маркера на основе типа I последовательности, которые имеют нуклеотидные полиморфизмы в сравнении с ДНК типа II которыхзначительно снижает эту редкую фона в экспериментах с использованием типа II Пру Δ Ku80 штамм 1. Если генный локус существенно (не может быть удален), или имеет чрезвычайно низкое генного таргетинга частота на локус, или если сохраняется [неинтегрированные] эписом не легко устраняется с помощью роста и выбора, эта родительская модель будет доминировать в картине, наблюдаемой в MPA резистентных клонов. Альтернативно целевой молекулы ДНК, иногда наблюдается интегрировать только в 5'-или 3'-геномной ориентации фланг и полоса наблюдается либо PCR3 или pCR4 (но не оба) вместе с ПЦР1 и ПЦР2, как показано на клон 10 (5 "интеграция) и клон 7 (3 'интеграция) (рис. 2С). Эти модели предполагают, редкие появления одного крест над интеграцией адресности плазмиды в локусе, который объединяет HXGPRT но эта направленной интеграции не удаляет интересующего гена. Эта модель (дорожки 7 и 10 на рис 2С) EMphasizes необходимости сообщать ПЦР данных, чтобы проверить, что произошло направленной интеграции вместо одного гомологичных пересекать и второй негомологичного интеграции целевой молекулы. Редко, мы наблюдаем генетической смеси, содержащей клон 12 (рис. 2С), которая представляет собой как родительская модель и целевую модель удаления. Эта картина, скорее всего, возникает в некоторых случаях от двух генотипов паразитов, которые присутствуют в том же месте в клонирования хорошо.
Общая схема для удаления HXGPRT от Δ ku80Δgoi :: HXGPRT удалить HXGPRT из штамма показан (фиг. 3А). Пара праймеров использоваться для подтверждения удаления HXGPRT от Δ Δ Ku80 gra2 :: HXGPRT (фигура 3В) усиливает уникальную ~ 1,2 т.п.н. полосу в PCR5 как показано на клоны 4, 7, 9, 11 и 12 (фиг. 3C). Если HXGPRT не удаляется из локуса, ~ 3.4 Kbp наблюдается полоса (клоны 1, 2, 3, 6, 8, 10, и родительский контроль). Несколько механизмов действовать так, чтобы удаление HXGPRT (6TX выбор) более сложной и менее эффективно, чем интеграция HXGPRT (MPA + X выбора). MPA выбора просто более эффективны, чем выбор 6TX 14,16. Кроме того, более высокие уровни HXGPRT выражения, необходимые для 6TX выбор, чем для MPA выборы 16,24. Следовательно, мутации, которые могут уменьшить HXGPRT активность фермента или мутации или эпигенетические изменения, которые снижают уровень экспрессии HXGPRT в локусе гена потенциально может отменить эффективность 6TX выбора. Даже с этими вызовами 6TX выбора, успеха 6TX выбор в Δ Ku80 штаммов составляет более 90% с первой попытки 1-3.
Общая схема прямого С-концевого мечения белок-кодирующих генов показано (рис. 4). ЭтоСхема использует прямую интеграцию через двойной крест над гомологичной рекомбинации из HXGPRT 3 'из помеченных геном и также использует стратегию проверки, аналогичные описанным выше. Когда ген предположительно основного гена этого может быть проверена с использованием второй трансфекции независимо выделенных улавливающей ДНК плазмиды. Альтернативные методы, такие как схемы для регулируемой экспрессии генов, которые доступны для дальнейшей проверки, если гена необходима 25.

Рисунок 1. Обзор конструкции улавливающей ДНК плазмиды. . Схемы для проектирования перекрываются праймеров для ПЦР-амплификации 5 'и 3' флангах цель с 33 б.п. перекрывается для pRS416 челночный вектор и HXGPRT миниген кассету. Праймеры, представленной на схематической былииспользуется для генерации 5 'и 3' целевой склонах pΔROP18 10. B. Общая стратегия для проектирования ориентации молекулы ДНК. Основу pΔGOI является pRS416 трансфер вектор, содержащий урацил (URA) и ампициллин (AMP) селективные маркеры. Извините, в pRS416 является ~ 1 кб, 5 'ДНК-мишени фланг усиливается из геномной ДНК с перекрытиями для кассеты HXGPRT миниген и pRS416 используя клавиши F1 и R1 праймеров, ~ 1 кб, 3' ДНК-мишени фланг усиливается из геномной ДНК с перекрытиями для Кассета HXGPRT миниген и pRS416 помощью F2 и R2 праймеров и кассета HXGPRT миниген. Следующие уникальных рестрикционных ферментов дайджест сайтов добавляются к праймеров: RE.X (рестриктазы Х места среза) в F1 грунтовки, чтобы вырезать плазмиду в 5'-конце 5 'фланг ДНК-мишени, RE.Y в R2 Грунтовка сократить плазмиды в 'конце 3' 3 ДНК фланг цели и RE.Z в R1 и F2 праймеров для акцизных мин HXGPRTigene кассету. C. Последовательности праймеров, используемый для создания ориентации конструкцию для удаления rop18. смелое области соответствуют T. гондий геномной последовательности и полужирный области соответствуют сайты рестрикции, и последовательности, которые пересекаются с pRS416 и кассета HXGPRT миниген. Пара HX_F и HX_R грунтовка усиливает HXGPRT кассеты миниген 14. Праймеры считывается из 5 'к 3'. Таблица адаптирована из Fentress и др. 10. Нажмите здесь, чтобы увеличить рисунок .
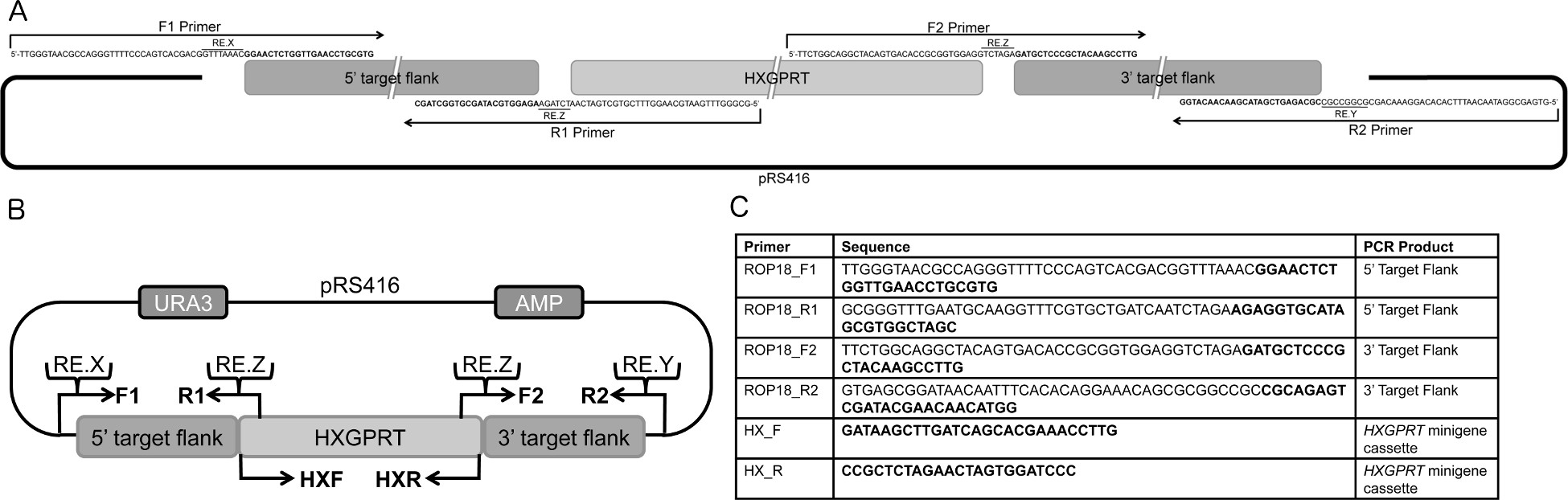{kind=link}

Рисунок 2. Обзор протокола для удаления гена с использованием HXGPRT. . Disruptioп ГОИ в штамме Δ ku80Δhxgprt помощью двойного кроссовера гомологичной рекомбинации использованием ~ 1 кб, 5 'ДНК фланг цели и ~ 1 кб, 3' ДНК-мишени фланг плазмиды pΔGOI. Стратегия ПЦР с использованием ПЦР1, ПЦР2, PCR3 и pCR4 (не в масштабе) показано, что позволяет генотип проверки для проверки клонов с целевой интеграции HXGPRT и удаление локуса ПИ. B. пар представитель праймер, сконструированный для подтверждения удаления rop18 . ROP18_DF и ROP18_DR усиливают ПЦР1, ROP18_ExF и ROP18_CxR усиливают ПЦР2, ROP18_CxF и DHFR_CxR усиливают PCR3 и DHFR_CxF и ROP18_CxR усиливают pCR4 (рис. 2А). Праймеры считывается из 5 'к 3'. Таблица адаптирована из Fentress и др. 10. C. Представитель Результаты проверки целевого удаления гена (rop18) с использованием HXGPRT. После трансфекции плазмиды pΔROP18 в Δ ku80Δhxgprt и отбор в MPA +X, MPA + X резистентные клоны анализировали на удаление rop18. Родительский контроль деформации положительна для ПЦР-1 (~ 400 б.п.) и ПЦР-2 (~ 650 б.п.) продукта и отрицательные для ПЦР-3 (~ 1200 пар оснований) и ПЦР 4 (~ 1300 пар оснований) продукта. Целевых нокаутом ГОИ является положительным для ПЦР2, PCR3 и pCR4 продукты и отрицательное для ПЦР 1 продукт (см. рисунок 2А). Показаны представитель панели результатов ПЦР1 и ПЦР2 (верхняя панель), PCR3 (средняя панель) и pCR4 (нижняя панель). Клоны, 3, 4, 5, 6, 8, 9 и 11 демонстрируют правильное исчерченности 1 ПЦР ПЦР2, PCR3 и pCR4, который определяет целевые событие делеция гена в клоне. Клоны, 1, 2, 7, 10 и 12 не являются ген делеции и соответствуют других потенциальных результатов представителя. (C = родительский штамм управления Δ ku80Δhxgprt, M = маркеры размера). Нажмите здесь, чтобы увеличить рисунок .
{kind=link}

Рисунок 3. Обзор протоколов для повторной ориентации интересующего гена, чтобы удалить HXGPRT. А. стратегии для удаления HXGPRT селектируемого маркера путем двойного кроссинговера гомологичной рекомбинации в штамме Δ ku80Δgoi :: HXGPRT использованием ~ 1 т.п.н. 5'-ДНК фланг цели и ~ 1 т.п.н. 3 'ДНК-мишени бок о плазмиды pΔGOIc. Стратегия ПЦР для проверки генотип изображен с использованием пары праймеров для анализа на продукт ПЦР (PCR5), который охватывает удаления (не в масштабе). B. представитель пара праймер, сконструированный для проверки удаление HXGPRT селектируемого маркера из gra2 локус в штамме Δ ku80Δgra2 :: HXGPRT. Грунтовки GRA2 _CLF и GRA2_CxR усиливают PCR5. Грунтовки считываются из 5 'к 3'. С. представителем панели 6TX-резистентные клоны, которые могут быть получены после трансфекции плазмиды pΔGOIc в Δ ku80Δgoi :: HXGPRT и выбор в 6TX. Клоны, 4, 7, 9, 11 и 12 демонстрируют правильное исчерченности из PCR5, который соответствует целевой удаления маркера HXGPRT (~ 1,2 т.п.н. продукт, который охватывает "бок с небольшим перекрытием с 5'-3 фланг и снаружи 3 ' фланг). Клоны, 1, 2, 3, 6, 8 и 10 (эта полоса света) представляют собой ожидаемый исчерченности около ~ 3,4 т.п.н., соответствующий родительский генотип клон, хотя эти клоны показали степень устойчивости к 6TX, которые позволили их выбора (этот фенотип может иногда возникать спонтанно остановке HXGPRT выражения (см. примечание в протоколе 2.1 (этап 8), а в разделе Представитель Результаты). (C = родительский штамм управления Δ ku80Δgoi :: HXGPRT, M = маркеры размера).w.jove.com/files/ftp_upload/50598/50598fig3large.jpg "целевых =" _blank "> Нажмите здесь, чтобы увеличить рисунок.

Рисунок 4. Конструкция ориентации молекулы ДНК, чтобы пометить С-конце белка. Стратегия основана на способности HXGPRT маркер также функционировать как вниз по течению регуляторная область 3 '. Основу pΔGOItag является pRS416 трансфер вектор, содержащий урацил (URA) и ампициллин (AMP) селективные маркеры. Извините, в pRS416 является ~ 1 кб, 5 'ДНК-мишени фланг усиливается из геномной ДНК с перекрытиями для кассеты HXGPRT миниген и pRS416 использованием Fgoi RTAG и праймеры, ~ 1 кб, 3' ДНК-мишени фланг усиливается из геномной ДНК с перекрытиями для Кассета HXGPRT миниген и pRS416 помощью F2 и R2Праймеры и кассета HXGPRT миниген. 5 'ДНК-мишени фланг заменяет терминирующий кодон с тегом (HA, Myc, His, и т.д.), а затем кодон терминации замены. MPA + X выбор интегрирует С-концевой метки и вниз по течению HXGPRT и перемещает 3 'UTR в положение, после HXGPRT маркера. Следующие уникальных рестрикционных ферментов дайджест сайтов добавляются к праймеров: RE.X (рестриктазы Х места среза) в грунтовке Fgoi, чтобы вырезать плазмиду в 5'-конце 5 'фланг ДНК-мишени, а в RE.Y R2 грунтовки, чтобы вырезать плазмиду в-конец 3 '3 фланг ДНК-мишени, и RE.Z в RTAG и F2 праймеры, чтобы вырезать кассета HXGPRT миниген для использования в конструкции плазмиды повторно целевой помеченный локуса гена удалить Маркер HXGPRT который будет восстанавливать размещения 3 'UTR.
Обсуждение
Здесь мы предлагаем протокол для эффективного целенаправленное изменение генов в Δ Ku80 штаммов паразита чтобы позволить эффективное восстановление генетически модифицированных потомков, которые обладают гена-мишени делеции, замены гена и / или на ярлыке генов. Последовательное выполнение этих способов обеспечивает надежный подход для выделения паразита мутанты, которые содержат одну или несколько целевых генетических манипуляций 1-3. При генерации целевых удаления зависит от множества факторов, использование Δ Ku80 штамма представлены стратегии и протокол значительно повышает эффективность и простоту делает точно определены мутантных штаммов T. гондий для функциональных исследований геномных.
Генома T. гондий во бесполых стадий гаплоидным. Таким образом, рассмотрение, чтобы сделать при планировании удалить ген, что ген не должно быть необходимым в пробирке. Тем не менее, эффективность круглый щитTing гена нокаутом в Δ Ku80 штаммов чрезвычайно высока, и это делает возможным выделение мутантных штаммов со значительным нарушением цены репликации. Если гена-мишени необходимо, паразиты часто перестают повторить при выборе. Другим важным фактором в целевых генетических манипуляций в том, что идеальный гомологию по крайней мере с 620 б.п. геномной таргетинга фланге, необходимых для получения целевых обнаруживаемых интеграции 2. Дольше областей гомологии производят более высокие эффективность таргетинга и таргетинга использованием ДНК флангах около 1000 BP является надежным и эффективным подходом 1-4. По этой причине важно, чтобы геномных таргетинга флангах, сгенерированы исходя из того же штамма T. гондий (RH, Пру) как штамм, который в настоящее время генетически модифицированные. Отсутствие достаточной гомологии часто приводит в целевых интеграцию одного геномного таргетинга фланга, но не других. Неполное интеграции или неполучение нокаутом маY быть также связано с эписомными настойчивости. Сразу же после трансфекции, и все ориентации молекул неинтегрированных эписом, а иногда и молекул, имеющих мишень может сохраняться как эписомы в течение многих поколений. Использование ДНК-мишени бока ~ 1 т.п.н. и продолжает передавать паразитов при выборе до повторного субклонирования часто решает проблемы, связанные с эписомными настойчивости.
После того, как нокаут проверяется и HXGPRT маркер снимается, гена-мишени стратегия может быть повторен на второй ПИ для создания нескольких делеции гена в одного штамма паразита 1-3. Поколения делеции, замены, или на ярлыке гены также могут быть выполнены с использованием различных селективных маркеров (блеомицин, CAT, пириметамин устойчивы DHFR и т.д.). Хотя CAT селектируемый маркер в настоящее время присутствует в тип II Δ Ku80 Pru геном 1, этот маркер можно удалить с помощью HXGPRT Выбор Протокол, описанный здесь. В наших руках, HXGPRT выбор является самым безопасным и наиболее эффективным маркером с самой высокой эффективности адресности, где один-копии целевых вставки достаточно для надежного выделения в MPA + X среды. HXGPRT также предоставляет полезные только в настоящее время подход к целевой удаление селективный маркер (рис. 3) с помощью 6-тиоксантин (6TX) выбор 2,3. Таким образом, этот протокол обеспечивает только текущий подход к разработке определены мутантных штаммов с несколькими целевыми генетических манипуляций.
Этот протокол представляет собой ценный и эффективный метод для целевого генетические манипуляции в Δ Ku80 штаммов Токсоплазма и применима для анализа одного гена, семейство генов, или генома функциональных исследований геномных. До наличие Δ Ku80 1,2 штаммов, эти подходы не были широко представляется возможным из-за неравенстваfficiency этих методов. Следовательно, этот протокол широко применяется для целевых генетических манипуляций Токсоплазма и доступной для всех исследователей заинтересованы в решении ряда вопросов по биологии паразита, ответа хозяина и функциональной геномики.
Раскрытие информации
Авторы заявляют, что у них нет конкурирующих финансовых интересов.
Благодарности
Работа выполнена при поддержке грантов от NIH для DJB (AI041930, AI073142, AI075931 и AI091461).
Материалы
| Name | Company | Catalog Number | Comments |
| Overlap Primers | Integrated DNA Technologies | 4 nmole Ultramer DNA oligos | |
| Validation Primers | Integrated DNA Technologies | 100 nmole DNA Oligo | |
| Yeast Strain #90845 | ATCC | Designation FY834 | |
| Shuttle Vector pRS416 | ATCC | 87521 | |
| DH10B E. coli | Invitrogen | 12033-015 | SOC broth in kit with E. coli |
| Resuspension Buffer | Qiagen | Buffer P1 in QIAprep Spin Miniprep Kit | |
| Miniprep Kit | Qiagen | 27104 | QIAprep Spin Miniprep Kit |
| Glass Beads | Scientific Industries | SI-BG05 | 0.5 mm acid-washed |
| Qiacube Automated Robotic Work Station | Qiagen | ||
| Electroporation Cuvette | USA Scientific | 9104-5050 | 2 mm-gap |
| BTX600 electroporator | BTX | ||
| Maxiprep Kit | Qiagen | 12662 | QIAprep Spin Maxiprep Kit |
| 25 cm2 Canted neck plug seal flask | Corning | 430168 | |
| 150 cm2 Canted Neck plug seal flask | Corning | 430823 | |
| Human Foreskin Fibroblasts (HFF) cells | ATCC | SCRC1041 | |
| Swin-lok Filter Holder | Whatman | 420200 | 25-mm-diameter |
| Membrane | Whatman | 110612 | Nucleopore Track-etched Membrane 3 mm pore size, 25-mm-diameter |
| Mycophenolic acid (MPA) | Sigma | ||
| Xanthine (X) | Sigma | ||
| 96-well Tissue Culture Tray | Corning Costar | ||
| 24-well Tissue Culture Tray | Corning Costar | ||
| MEM Eagle Media | Lonza Biowhittaker | 12-611F | |
| Fetal Bovine Serum | Gibco | 26140-111 | |
| Antibiotic-Antimycotic (Anti-Anti) | Gibco | 15240-062 | |
| Spin Column | Primm Labs | PAE-100 | Easy Clean DNA Extraction Spin Kit |
| T4 DNA Ligase | New England Biolabs | ||
| 6-thioxanthine | Acros Organics | ||
| Tissue DNA Minikit | Qiagen | 51104 | QIAamp DNA Blood Mini Kit |
| Cell Culture Freeze/Recovery Media | Gibco | 126-48-010 | |
| Phosphate Buffered Saline | Hyclone | SH30028.02 | minus calcium, minus magensium |
Ссылки
- Fox, B. A., et al. Type II Toxoplasma gondii KU80 knockout strains enable functional analysis of genes required for cyst development and latent infection. Eukaryot. Cell. 10, 1193-1206 (2011).
- Fox, B. A., Ristuccia, J. G., Gigley, J. P., Bzik, D. J. Efficient gene replacements in Toxoplasma gondii strains deficient for nonhomologous end joining. Eukaryotic Cell. 8, 520-529 (2009).
- Fox, B. A., Bzik, D. J. Avirulent uracil auxotrophs based on disruption of orotidine-5'-monophosphate decarboxylase elicit protective immunity to Toxoplasma gondii. Infection and Immunity. 78, 3744-3752 (2010).
- Hortua Triana, M. A., et al. Biochemical and molecular characterization of the pyrimidine biosynthetic enzyme dihydroorotate dehydrogenase from Toxoplasma gondii. Molecular and Biochemical Parasitology. 184, 71-81 (2012).
- Kim, K., Weiss, L. M. Toxoplasma gondii: the model apicomplexan. Int. J. Parasitol. 34, 423-432 (2004).
- Gajria, B., et al. ToxoDB: an integrated Toxoplasma gondii database resource. Nucleic Acids research. 36, 553-556 (2008).
- Kim, K., Weiss, L. M. Toxoplasma: the next 100 years. Microbes and infection / Institut Pasteur. 10, 978-984 (2008).
- Butcher, B. A., et al. Toxoplasma gondii rhoptry kinase ROP16 activates STAT3 and STAT6 resulting in cytokine inhibition and arginase-1-dependent growth control. PLoS pathogens. 7, e1002236(2011).
- Daher, W., Klages, N., Carlier, M. F., Soldati-Favre, D. Molecular characterization of Toxoplasma gondii formin 3, an actin nucleator dispensable for tachyzoite growth and motility. Eukaryot. Cell. 11, 343-352 (2012).
- Fentress, S. J., et al. Phosphorylation of immunity-related GTPases by a Toxoplasma gondii-secreted kinase promotes macrophage survival and virulence. Cell Host Microbe. 8, 484-495 (2010).
- Musiyenko, A., Majumdar, T., Andrews, J., Adams, B., Barik, S. PRMT1 methylates the single Argonaute of Toxoplasma gondii and is important for the recruitment of Tudor nuclease for target RNA cleavage by antisense guide RNA. Cellular microbiology. 14, 882-901 (2012).
- Straub, K. W., Peng, E. D., Hajagos, B. E., Tyler, J. S., Bradley, P. J. The moving junction protein RON8 facilitates firm attachment and host cell invasion in Toxoplasma gondii. PLos Pathog. 7, e1002007(2011).
- Szatanek, T., et al. Cactin is essential for G1 progression in Toxoplasma gondii. Molecular Microbiology. 84, 566-577 (2012).
- Donald, R. G., Carter, D., Ullman, B., Roos, D. S. Insertional tagging, cloning, and expression of the Toxoplasma gondii hypoxanthine-xanthine-guanine phosphoribosyltransferase gene. Use as a selectable marker for stable transformation. J. Biol. Chem. 271, 14010-14019 (1996).
- Donald, R. G., Roos, D. S. Gene knock-outs and allelic replacements in Toxoplasma gondii: HXGPRT as a selectable marker for hit-and-run mutagenesis. Molecular and Biochemical Parasitology. 91, 295-305 (1998).
- Pfefferkorn, E. R., Borotz, S. E. Toxoplasma gondii: characterization of a mutant resistant to 6-thioxanthine. Exp. Parasitol. 79, 374-382 (1994).
- Gietz, R. D., Schiestl, R. H. Frozen competent yeast cells that can be transformed with high efficiency using the LiAc/SS carrier DNA/PEG method. Nature Protocols. 2, 1-4 (2007).
- Oldenburg, K. R., Vo, K. T., Michaelis, S., Paddon, C. Recombination-mediated PCR-directed plasmid construction in vivo in yeast. Nucleic Acids Res. 25, 451-452 (1997).
- Fox, B. A., Gigley, J. P., Bzik, D. J. Toxoplasma gondii lacks the enzymes required for de novo arginine biosynthesis and arginine starvation triggers cyst formation. Int. J. Parasitol. 34, 323-331 (2004).
- Roos, D. S. Molecular genetic tools for the identification and analysis of drug targets in Toxoplasma gondii. Curr. Top. Microbiol. Immunol. 219, 247-259 (1996).
- Roos, D. S., Donald, R. G., Morrissette, N. S., Moulton, A. L. Molecular tools for genetic dissection of the protozoan parasite Toxoplasma gondii. Methods in Cell Biology. 45, 27-63 (1994).
- Singh, U., Brewer, J. L., Boothroyd, J. C. Genetic analysis of tachyzoite to bradyzoite differentiation mutants in Toxoplasma gondii reveals a hierarchy of gene induction. Molecular Microbiology. 44, 721-733 (2002).
- vanden Hoff, M. J., Moorman, A. F., Lamers, W. H. Electroporation in 'intracellular' buffer increases cell survival. Nucleic Acids Research. 20, 2902(1992).
- Pfefferkorn, E. R., Bzik, D. J., Honsinger, C. P. Toxoplasma gondii: mechanism of the parasitostatic action of 6-thioxanthine. Exp. Parasitol. 99, 235-243 (2001).
- Mital, J., Meissner, M., Soldati, D., Ward, G. E. Conditional expression of Toxoplasma gondii apical membrane antigen-1 (TgAMA1) demonstrates that TgAMA1 plays a critical role in host cell invasion. Mol. Biol. Cell. 16, 4341-4349 (2005).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены