
Method Article
유전 조작
요약
여기에서 우리는 유형 I 및 유형 II를 사용하는 방법을보고 Δku80 변종 톡소 포자충 효율적으로 기능 게놈 분석 대상으로 유전자 삭제 및 유전자 교체를 생성합니다.
초록
상동 재조합을 사용하여 대상 유전자 조작 유전자 기능과 표현형 (들)의 상세보기를 얻을 수있는 기능 게놈 분석을 위해 선택의 방법입니다. 표적 유전자의 삭제, 대상 돌연변이 보완 유전자 기능 및 / 또는 태그 유전자 돌연변이 균주의 개발은 이러한 유전자 조작을 효율적으로 이중으로 통합 중재를 사용하여 그 유전자 현장을 대상으로 할 수있는 경우, 특히 유전자 기능을 해결하기 위해 강력한 전략을 제공합니다 상동 재조합을 통해 교차합니다.
톡소 포자충의 동종 재조합, 기능 게놈 분석의 매우 높은 속도로 인해 이전에 특정 유전자 좌위에 유전자 삭제 및 유전자 교체 대상에 대한 효율적인 방법의 부재로 제한되었습니다. 최근에, 우리는 유형 I 및 유형 II T의 변종 동종 재조합의 주요 통로를 폐지 유전자 encodin을 삭제하여 gondii의G KU80 단백질 1,2. Δku80 균주 tachyzoite (급성) 및 in vitro와 in vivo에서 bradyzoite (만성) 단계에서 일반적으로 행동과 상동 재조합 본질적으로 100 %의 빈도를 나타냅니다. Δ ku80 균주는 단일 유전자뿐만 아니라 게놈 규모의 1-4 페이지의 기능 게놈 연구는 가능합니다.
여기, 우리는 I 및 유형 II Δku80Δhxgprt 균주는 T.의 유전자 타겟팅 방식을 사전에 유형을 사용하는 방법을보고 gondii의. 우리는 산틴 - 크 산틴-구아닌의 phosphoribosyltransferase (HXGPRT) 선택 마커의 대상 삽입 또는 삭제에 의해 유전자 삭제, 유전자 교체 및 태그 유전자를 생성하기위한 효율적인 방법을 설명합니다. 프로토콜을 대상으로 설명하는 유전자는 기생충 게놈의 기능 분석을 진행하고 수행 한 균주를 개발하는 Δku80 변종 다양한 방법으로 사용할 수 있습니다여러 유전자 조작을 대상으로. 이 유전 적 방법과 이후의 표현형 분석의 응용 프로그램은 T.의 생물학의 기본적이고 독특한 측면을 보여줄 것입니다 gondii의 말라리아 (Plasmodium의 SP에.) 및 크립토스포리디움 (Cryptosporidium의)이 발생할 관련된 중요한 인간 병원체.
서문
톡소 포자충은 종종 일반적인 의무 세포 내 원생 기생충이며, 만성적으로 동물과 인간 5의 넓은 범위를 감염. 그것은이 병원균에 의해 10 억 이상의 인류가 현재 추정하고 만성 감염됩니다. T.에 의한 질병의 중요성에 추가 체외 성장과 우수한 마우스 모델에서 T. 만든의 gondii의 감염, 실험 도구, 강력한 게놈 자원 6의 증가 가용성, 편의성 gondii의 세포 진핵 병원체 및 말라리아 (Plasmodium의 SP에.) 및 크립토스포리디움 (Cryptosporidium의) 5,7과 같은 치명적인 질병을 일으키는 원인이 다른 중요한 apicomplexan 기생충의 광범위한 연구를위한 선도적 인 모델 시스템. 모델 생물로 톡소 포자충의 중요한 한계는 유전자 조작을 대상으로 나른다 자손의 비효율적 복구되었습니다. 이 proble 나유전자 타겟팅의 M은 T의 야생형 균주의 동종 재조합의 매우 높은 주파수를 기준으로 상동 재조합의 낮은 주파수로 인해 광범위한 DNA 상 동성이 유전자 연구 2에 사용되는 DNA의 표적 분자에 제공되는 gondii의 경우에도.
우리는 최근 유전자 타입 동종 재조합의 주요 통로를 차단 KU80 단백질 1,2 인코딩 유전자를 삭제하여 I 및 유형 II 톡소 포자충의 변종. I 및 유형 II Δ ku80 변종 정상적인 성장 속도, 크기 및 in vitro와 in vivo에서 체외 tachyzoite 및 bradyzoite 단계에서 생체의 두 동작을 결과 형식은, 그러나, 이러한 변형은 기본적으로 100 % 상동 재조합의 빈도와이 전시 표현형은 몇 칸토에 수백의 표적 유전자 조작으로 빠르게 분리 원하는 자손의 가능성을 증가USand에서 배 1,2,4. 최근 사회 전반 I 및 유형 II Δ ku80 긴장이 속도까지 크게 램핑이 유형의 사용, 다양한뿐만 아니라, T.의 표적 유전자 접근 방법의 성공률 gondii의 1-4,8-13. 여기에서 우리는 T.의 Δ ku80 변종 대상으로 유전자 삭제, 유전자 교체, 유전자 태그에 대한 포괄적 인 프로토콜을 설명 gondii의. 우리는 디자인하고 안정적으로 톡소 포자충의 Δ ku80 균주의 특정 유전자 또는 유전자 좌위에 유전자 조작을 대상으로하는 방법을 보여줍니다.
프로토콜
1. 관심의 유전자를 삭제하는 표적 유전자
이 프로토콜은 정의 유전 현장에서 표적 유전자의 삭제가 돌연변이 균주의 효율적인 생성을 위해 설계되었습니다. 앞에서 설명한 T. gondii의 산틴 - 크 산틴-구아닌의 phosphoribosyltransferase (HXGPRT) 선택 마커 mycophenolic 산 및 크 산틴 (MPA + X) 선택 14-16에 따라 안전하고 신뢰할 수있는 마커 삽입이 프로토콜에서 사용됩니다. 이 프로토콜은 효모 재조합 복제 17,18에 따라 DNA 표적 분자를 구성하기위한 검증하고 비용 효율적인 방법을 사용합니다. 몇 가지 다른 프로토콜은 체외 재조합 복제 방법, 또는 융합 PCR에 세균 재조합 복제 방법을 사용하여 재조합 표적 분자를 구성 상업적으로 사용할 수 있습니다. 아래에서 설명하는 프로토콜은 복제를 복구하는 항의 가장 높은 전반적인 효율성과 신뢰성을 제공합니다T의 Δ ku80 변종 타겟 유전자 삭제를 가진 ITES gondii의.
DNA를 대상으로 1.1 준비
- ToxoDB 6 (액세스 http://www.toxodb.org을 관심의 유전자 (GOI)에 대한 게놈 서열을 얻기 위해).
참고 : 게놈 시퀀스가 조작되고 변형에 가장 가까운 ToxoDB 데이터베이스 유형 I, II 또는 III 게놈 시퀀스에서 확인하셔야합니다. 예를 들면 : 저기, 대한 RH와 ME49를위한 GT1.
- 5 '과 3'효모 셔틀 버스로 33 bp의 중복을 통합 관심의 유전자 (GOI)의 게놈 대상 측면을 증폭 설계 특정 오버랩 프라이머 벡터 pRS416 (또는 양자 택일 pRS426)와 선택 마커로 33 bp의 중복을 통합 ( HXGPRT) (그림 1A-B). 5 '의 양쪽 끝에 각 프라이머의 독특한 제한 효소 사이트를 추가33 bp의 중복과 T. 사이에 차 3 '게놈 대상 측면, gondii의 특정 프라이머 시퀀스 (들) (그림 1A-B).
참고 :이 30 bp의 중복보다 큰 효율 효모 재조합 복제가 필요합니다. 5 '과 3'게놈 대상 측면은 800 타겟 삭제를 정의하는 1,400 bp의 사이에 특정 DNA 조각을 증폭하도록 설계되었습니다. 짧은 측면이 크게 Δ ku80 Δ hxgprt 배경 2의 효율성을 대상으로 감소된다는 점에 유의해야합니다.
- PCR는 5 '프라이머 F1과 R1을 사용하여 대상 측면을, 그리고 PCR은 3 증폭'증폭 게놈 DNA 3의 프라이머 F2와 R2를 (그림 1A-C)을 사용하여 대상 측면입니다. 따로, PCR은 관련 5 '와 3'DHFR이 프라이머 HXF 및 HXR를 사용하여 HXGPRT cDNA를 pminiHXGPRT 카세트 (14)의 측면 지역을 포함하여 1 ~ 2 KBP HXPGRT 유전자를 증폭(그림은 1B-C).
참고 : 형질 전환 할 부모의 변형으로부터 게놈 DNA를 사용하여 대상 측면을 증폭.
주 : 5 '와 3'DNA 등의 혼란 현장에서 HXGPRT의 대상 제거와 같은 후속 효율적인 유전자 조작을 용이하게하는 측면을 대상으로 상대 전진 방향으로 배치 HXGPRT 마커.
- 아가로 오스 겔 전기 영동에 의해 정확한 PCR 제품 크기를 확인하고 아가로 오스 겔 표준을 사용하여 DNA 단편 농도 또는 DNA 농도를 결정하기위한 다른 방법을 추정한다.
- 로 Gietz 및 Schiestly 17에서 설명 관할 효모의 aliquots를 생성합니다.
- 5 50 NG '게놈 대상 측면 3 50 NG'게놈 대상 측면, HXGPRT 선택 마커의 100 NG, 10의 최종 볼륨을 달성하기 위해 멸균 H 2 O와 선형 셔틀 벡터의 50 NG를 결합하십시오 -20 μL. 세 PCR 제품과 효모 E.와 관할 효모를 변환 Gietz 및 Schiestly (17)에 의해 기술 된 프로토콜을 사용하여 재조합 복제에 대한 대장균의 셔틀 벡터입니다. 플레이트 우라실 마이너스 최소 배지 한천 플레이트에 효모를 변환하고 2 30 ° C에서 알을 품다 - 6 일.
참고 : 플라스미드, 5 '목표 측면, HXGPRT 마커, 및 3'대상 측면의 주문 어셈블리는 DNA 조각 (그림 1A-B) 사이의 설계 33 bp의 중복에 의해 촉진되고있는 상동 재조합에 의해 중재됩니다 효모.
- 유라 마이너스 접시에 배 YPAD 2 ㎖를 추가하고 부드럽게 한천을 방해하지 않고 식민지를 꺼 내려 긁어 수확 효모. 5,000 XG에서 5 분 동안 효모 용액을 원심 분리하고 상층 액을 버린다.
- RNaseA를 포함하는 부유 버퍼 250 μL에 효모 펠렛을 resuspend을 50 추가 - 100 μl를솔루션 CID 씻어 유리 구슬. 효모 세포를 분쇄하는 최고 속도에서 5 분 소용돌이.
- 유리 구슬은 튜브의 바닥에 정착하고 2 ㎖ 에펜 도르프 튜브에 뜨는을 전송 할 수 있습니다.
- ~ 100 μL의 최종 볼륨 현탁, miniprep의 DNA 분리 키트를 사용하여 효모의 DNA를 분리합니다.
- DH10B 일렉트로 관할 E. 40 μL 2 μL 효모 DNA (~ 50 NG)를 혼합 대장균 얼음에 보관.
- 냉장 2mm 간격의 electroporation의 큐벳에 솔루션을 전송하고 2.4 kV의 및 129 Ω에서 박테리아 세포를 electroporate.
- 각 큐벳과 14 ML 스냅 캡 팔콘 튜브로 전송 용액에 0.8 ML SOC 국물을 추가하여 구조 세포. 60 분 - 40 롤러 드럼에 37 ° C에서 세포를 품어.
- 판 E. 2XYT + 암피실린 (AMP) 접시에 대장균 및 37 ° C에서 하룻밤 번호판을 품어.
- ~ 6 선택 - 8 하나의 식민지를하고 37 ℃에서 ~ 16 시간 동안 3 ML 2XYT + AMP 성장
- 각 E.의 30 % 글리세롤 냉장고 재고를 준비 대장균의 복제.
- E.로부터 플라스미드 pΔGOI 분리 대장균 클론 ~ 100 μL의 최종 볼륨 현탁, miniprep 키트를 사용하여.
- 제한 효소를 사용하여 pΔGOI가 DNA 플라스미드 크기를 측정하고 예상 pΔGOI의 DNA 패턴을 밴딩 확인 다이제스트 확인합니다.
- DNA는 5 '와 3'의 게놈 시퀀스 데이터를 기반으로 100 % DNA 시퀀스를 확인하는 게놈 대상 측면의 순서로 pΔGOI의 유효성 검사를 완료 http://www.ToxoDB.org을 .
참고 : 완벽에 가까운 순서 상동는 각 염기쌍의 차이 완벽한 동성의 길이에 해당 컷을 구성 이후 효율성 3을 대상으로 유전자를 극대화하기 위해 필수적입니다.
참고 : 저기, 부모의 성을 기준으로 유형 II Δku80 균주의 게놈 서열비는이 시간에 사용할 수 없습니다. 이 작품은 유형 II Δ ku80 변형에 대한 대리 게놈과 유형 II ME49 게놈 시퀀스를 사용합니다. 유전 좌위의 숫자에서 시퀀스 데이터를 기반으로, 그것은 ME49 게놈 저기, 게놈 ~ 1 만 당 뉴클레오티드, 이하의 주파수에서 단일 염기 다형성을 나타내는 것으로 추정된다.
- 검증 E.에서 글리세롤 주식과 ~ 250 ML 2XYT + AMP 하룻밤 문화를 접종하여 pΔGOI 주식의> 200 μg을 생성 대장균 miniprep 클론.
- ~ 1000 μL의 최종 볼륨 현탁 maxiprep의 DNA 분리 키트를 사용하여 대형 문화 pΔGOI 플라스미드 DNA를 분리합니다.
- 이전 형질에 대한 올바른 대상 DNA 분자를 확인 pΔGOI maxiprep DNA 시퀀싱 제한 효소 소화, 또는 DNA를 반복합니다.
- 독특한 제한 효소 X (는 re.X)를 사용하여 5 '끝에 ~ 15 μg의 pΔGOI을 선형화다이제스트 사이트 5 '대상 측면 (그림의 1B)에 내장.
참고 : T. 타겟팅 유전자 gondii의 관심 2 유전자 현장에서 이벤트를 대상으로하는 효율적인 주파수를 얻기 위해 DNA를 타겟팅 ~ 10 μg의 최소 필요합니다.
- pΔGOI의 선형화를 확인하고 아가로 오스 겔 전기 영동 또는 다른 방법으로 DNA 농도를 결정합니다.
주 : 5에서 제한 효소 소화 '측면이 완료 선형화를 얻을하지 않습니다, 설계 3'경우 고유 RE.Y 다이제스트 사이트가 pΔGOI의 선형화를위한 대안 사이트로 사용할 수 있습니다.
참고 : 완전 선형화 대상으로 DNA 분자는 Δ ku80 균주에서 상동 재조합에 의해 대상으로 성공적인 유전자 필수적이다.
- incubat에 의해 제한 효소를 중화15 68 ° C에서 ING - 20 분.
- 살균 H 2 O. 100 μL의 선형화 플라스미드의 적어도 15 μg을 준비 형질 전환의 때까지 -20 ° C에서 보관 선형 pΔGOI.
T. 1.2 준비 gondii의 기생충, 형질 전환, 선택, 서브 클로닝, 검증, 보관 및 유전자 조작 종자의 유지
T의 문화와 조작을위한 일반적인 방법 인간의 포피 섬유 아세포 (HFF) 세포 gondii의는 19-21 설명되어 있습니다. Δ ku80 Δ hxgprt의 변종 기생충 감염 매체 (이글 최소 필수 매체 (EMEM) 1 % 태아 소 혈청 (FBS), 1 % 항진균 항생제 100X 주식에서 희석으로 보충 성장 매체 1,2)에서 정상적으로 복제합니다. 톡소 포자충 모든 작업은 바이오 안전성 수준이 절차를 사용하여 수행해야합니다. T. gondii의 RHΔ ku80 2 깎다ntal 변형이 로스 연구소 14에서 RHΔ hxgprt 균주로부터 생성되었습니다. PruΔku80 1 부모의 변형이 안정적으로 통합 CAT 선택 마커 bradyzoite 단계 특정 발기인 LDH2의 통제하에 녹색 형광 단백질 (GFP) 기자를 포함 PruΔhxgprt (저기, gniaud 변형 BSG-4 22)로부터 생성되었습니다.
- 1 X 10 6 가능한 유형 I RH Δ ku80 Δ hxgprt 기생충에 합류 HFF 세포를 포함하는 25cm 2 플라스크에 접종 또는 2 × 10 6 가능한 유형 II Prugniaud (저기,) Δ ku80 Δ hxgprt 기생충 두 25cm 2 플라스크에 접종.
참고 : 실행 가능한 기생충 갓 밖으로 용해가 세포 외 기생충으로 정의됩니다.
참고 : 이 스케일은 세 가지 형질 전환 실험에 대한 가능한 기생충의 충분한 수를 제공합니다.
- Δ ku80 Δ hxgprt 감염 문화를 검사 ~ 68-72 시간이 포스트 감염. 형질의 정확한 타이밍을 도모하기 위해 HFF 세포의 95 % 용해 - 가능한 기생충 ~ 90의 존재를 확인합니다.
참고 : 낮은 기생충 생존 유전자 타겟팅 효율성을 폐지하기 때문에 갓 egressed 기생충의 분리는이 프로토콜의 성공을 위해 필수적입니다.
- 25cm 2 플라스크에 플러그 씰 뚜껑을 닫아 솔루션으로 가능한 기생충을 선동하고 적극적으로 뚜껑의 내부 또는 플라스크의 목에 액체가 튀는없이 성장 표면의 기생충을 선동 앞뒤로 플라스크를 흔들어 .
참고 : 기생충 수확 유형 I 또는 주사기 바늘 방출 프로토콜을 필요로하지 않습니다II Δ ku80 변종을 입력합니다.
- 3 μm의 멤브레인을 포함하는 필터 홀더에 부착 된 10 ㎖ 주사기에 기생충 솔루션을 전송하고 오픈 15 ML 스크류 캡 튜브의 상단에있는 필터 장치를 놓습니다. 주사기 15 ML 스크류 캡 튜브에 기생충을 필터링합니다.
참고 : 세포막을 통해 기생충을 필터링하여 감염된 세포와 세포 파편을 제거합니다.
- 혈구를 사용하여 기생충 농도 (ML 당 tachyzoite 형태)를 결정합니다.
참고 : 선택 사항 : 기생충 솔루션을 사용하여 플라크 형성 단위 (PFU) 7 읽 분석 21 설정 - 팔일 이후 형질 전환 기생충의 충분한 가능성 (PFU는 ≥ 0.2의 비율을 tachyzoite하기)를 결정합니다.
- 7 1,400 XG에 분, 기생충 펠렛을 방해하지 않고 ~ 0.4 ML에 뜨는을 대기음을위한 펠렛 기생충. 1,40에서 2 분 동안 회전0 XG와 대기음은 기생충 펠렛을 방해하지 않고 ~ 0.01 ML에 상층 남아.
- 튜브의 바닥을 쓸어 기생충 펠렛을 resuspend을 즉시 4 기생충 농도를 얻기 위해 기생충 펠렛 cytomix 23 버퍼 (1.33 배 농도)를 추가 - 5 × 10 7 기생충 / ㎖ cytomix합니다.
참고 : 이러한 추가는 유전자 타겟팅 효율성을 개선하지 않기 때문에 cytomix에 ATP와 글루타티온의 추가를 생략합니다.
- 프로토콜 1.1 (26 단계)에서 선형 pΔGOI의 100 μL 나누어 지는를 해동.
- 기생충 cytomix 솔루션 (~ 1.3-1.6 × 10 7 기생충)의 0.3 ML 전송 프로토콜 1.1 (26 단계)에서 선형 pΔGOI의 100 μL로 즉시 냉장 2mm 간격으로 전체 0.4 ML 기생충 + pΔGOI 믹스를 전송 electroporation의 쿠 베트.
- 1.4 kV의 24 Ω에서 기생충을 Electroporate.
- 해상도t ~ 5 분 다음 30 ML 감염 매체에 합류 HFF 세포를 포함 150cm 2 플라스크에 형질 큐벳의 전체 내용을 전송하고 하룻밤 감염된 문화를 품어 상온에서 형질 큐벳.
- mycophenolic 산에 선택하기 + 크 산틴 (MPA + X) ~ 20 시간으로 감염 매체를 대체하여 포스트 형질 (그림 2A) MPA + X 선택 배지 (MPA (25 ㎍ / ㎖) 및 크 산틴 (50 ㎍ / ml)에 감염 매체).
참고 : + X 선택 플라스크 잠복기 MPA를 방해하지 마십시오.
- 시각적 단층을 조사하여 MPA + X 선택 플라스크 ~ 8 일 포스트 - 형질의 PFUs의 총 수를 추정한다. 가벼운 현미경에 의해 감염 PFUs하고 건강한 지역 개발의 존재를 확인합니다.
주 : 패 하루 8으로 표시되지 않은 경우, 감염의 징후에 대한 선택과 모니터를 유지돌연변이 균주 이후 이온은 감소 복제 속도를 가질 수있다.
참고 : Δ ku80 Δ hxgprt 기생충의 변종이 매우 심한 가진 돌연변이 균주의 성공적인 분리를 허용 원하지 않는 nontargeted 이벤트의 매우 낮은 배경을 가지고 있지만, 치명적인없는 성장하자. 유전자가 필수적이며이를 삭제할 수없는 경우, 기본 형질 또는 이후 기생충 인구 전달, 인구는 대상 녹아웃에 해결로 기생충 선택시 복제를 중단 표현형을 보여줄 수 있습니다.
- 단층의 세포 기생충을 쫓아하는 3 단계로 플라스크를 흔들어 [보이는 패는 개발 한 후] 기생충 솔루션을 생성 할 수 있습니다. 전송 ~에서 기생충 솔루션의 0.5 ML MPA + 25cm 2 X 선택 플라스크 MPA + 합류 HFF 세포 (이 문화 패스 1A를 지정)를 포함하는 X 선택 플라스크.
- 기생충시키다의 원래 150cm 2 MPA + X 선택 2 ~ 3 일 후 플라스크 및 전송 ~ 두 번째 25cm 2-0.5 ML 기생충 솔루션 MPA + 합류 HFF 세포 (이 문화 패스 1B를 지정)를 포함하는 X 선택 플라스크.
- 감염의 영역이 패스 1A 및 / 개발하거나 2 ~ 5 일에 대한 1B 플라스크를 통과 할 수 있습니다. 시각적으로 패스 1A와 1B 플라스크 감염의 건강 영역이 25 가능한 PFUs의 최소 포함하는 광학 현미경으로 확인합니다.
- 25cm 2 플라스크에 MPA + X 선택 배지에서 계속 통과를 위해 패스 1A 또는 패스 1B 플라스크 중 하나를 선택합니다. MPA + X 선택에 따라 흐름을 유지한다.
참고 : 기생충은 사전에 서브 클로닝에 인구에서 비 통합 지속되고 episomes을 삭제하는 세대 충분한 수의 배양해야합니다. 유형 I RH Δ ku80 Δ hxgprt, 또는 그 이전 이십오일 후 형질에 대한 위하여 사전 이십일 후 형질에 서브 클로닝을 수행하지 마십시오유형 II 저기, Δ ku80 Δ hxgprt 기생충이 통합되지 않은 episomes에게 1,2를 희석 할 수있는 충분한 시간을 허용합니다.
- MPA + X 선택 배지로 합류 HFF 세포를 포함하는 96 - 웰 트레이에 Subclone 기생충. 1 기생 / 잘 2 기생충 / 잘 농도에서 두 번째 트레이의 농도에 하나의 트레이를 준비합니다.
참고 : 최근 (~ 90 - 25cm 2 플라스크에 HFF 세포의 95 %) 용해가 Subclone 기생충 기생충이 높은 가능성을 가지고 있는지 확인 할 수 있습니다.
참고 : 서브 클로닝을위한 최적의 기간 후 형질 전환이 성공적으로 목표 기생충 선택한 인구의 1에서 높은 주파수에서 발생하는 시간을 측정하여 설립되었다.
- 통로에 계속 감염 주간 통로 일정을 사용하여 MPA + X 선택 배지에서 형질 전환 기생충의 주요 인구기생충 용액 10 μL와 25cm 2 HFF 플라스크.
참고 : 대상 유전자 삭제 (녹아웃) 휴대 클론 18 단계, resubclone 지속적으로 유지 인구의 기생충의 첫 번째 서브 클로닝에서 얻을하지 않는 경우 ~ 10-12주 후 형질 전환.
참고 : 유전자의 삭제를 획득하지 않는 이유 중 하나는 일부 pΔGOI 플라스미드의 에피 솜 지속성에서 발생합니다. 에피 솜 지속성 5에 포함 된 시퀀스 '와 3'게놈 대상 측면에 의해 결정되며 HXGPRT 유전 적 요소에서 발생하는 나타나지 않습니다. 약 5 - 대상 플라스미드의 10 %가 기생충 인구에게 세대 수 차례 지속되고 episomes을 희석 주로 안정 integrants의 인구를 해결할 수 있도록하는 서브 클로닝의 이후 시간을 필요로 상당한 에피 솜 지속성을 나타냅니다.
- 점수를 96 - 웰 트레이 6 -에서 광학 현미경에 감염 한 지역으로 식별 한 PFU를 포함하는 우물 팔일 후 서브 클로닝 (유형 II 기생충) - 칠일 후 서브 클로닝 (유형 I 기생충) 또는 7 40X 또는 60X 파워 하나. 우물 PFU의 위치를 지정하는 96 잘 트레이 뚜껑에 작은 점을 표시합니다.
- 잘 잘 기생충을 분산 PFU 이상 유체의 흐름을 지시하여 50 μL (200 μL 팁)로 설정 피펫을 사용하여 단일 PFU를 포함하는 각의 내용을 섞는다.
참고 : 잘 혼합하면 HFF 단층의 기생충 용해를 가속화합니다. I RH 변종이 잘 용해됩니다 ~ 4 일 혼합 한 후, 타입 II 저기, 기생충은 잘 용해됩니다 2 ~ 5 일 혼합 한 후 입력하십시오.
- 10 μL 피펫 팁 기생충 솔루션을 동시에 드로잉 최대 6 μL 동안 - 0.5을 사용하여 이러한 용해 우물의 각각의 맨 아래에 우물을 (클론) 용해 다스와 스크래치를 선택합니다. 트랜스1 ML MPA + X 선택 배지에 합류 HFF 세포를 포함하는 24 잘 트레이에서 잘하는 기생충 솔루션의 6 μL FER.
참고 : 개방 우물의 바닥에 존재하는 기생충과 지속적인 접촉 피펫 팁의 구멍 계속 잘 바닥에 흠집을하는 것이 필수적입니다.
- HFF 세포의 용해를위한 24 잘 트레이를 모니터링 할 수 있습니다.
참고 : I RH 기생충은 일반적으로 1 ~ 4 일 24 잘 형식으로 잘 용해됩니다 입력합니다. 유형 II 저기, 기생충은 일반적으로 2 ~ 5 일에 잘 용해된다.
- + X 선택 매체 잘 당 합류 HFF 세포와 1 ML MPA를 포함하는 새로운 24 잘 트레이의 해당 well에 24도 형식으로 각 우물의 바닥에서 긁어 가능한 기생충 용액 2 μL를 전송합니다.
참고 : 모든 기생충 클론과 선이 계속 메인이 될 수 있습니다이 24 잘 트레이 형식으로 매 7 일마다 실행 가능한 기생충 용액 2 μL를 전달하여 의한 것.
- 확인하는 기생충이 성공적으로 시각 광학 현미경 ~ 18 시간이 포스트 흐름과 함께 검사하여 새 24 잘 트레이로 옮겨졌다.
- 도 1 ML, 10 ML 피펫과 에펜 도르프 튜브에 기생충 솔루션을 전송을 사용하여 24 - 23 단계에서 24 잘 트레이의 용해 우물에서 수확 기생충.
- 펠릿 7 분, 일단 1 ML 인산에 씻어 1,400 X g에서 기생충은 3 분 동안 1,400 X g에서 다시 식염수 (PBS)와 펠렛을 버퍼.
- 펠렛의 대기음 PBS 다음, 기생충을 resuspend을 위해 펠릿에 PBS 200 μl를 추가합니다. DNA 분리 될 때까지 -80에서 기생충 솔루션 ° C를 고정합니다.
- 조직 DNA 분리 minikit를 사용하여 각 클론의 기생충 DNA를 분리합니다.
- 검증 프라이머 (그림 2A-C)를 사용하여 PCR에 의한 목표 유전자 삭제 (녹아웃) 확인합니다. 에 대한 테스트그 유전자의 기능을 코딩 영역의 부재 상류 CXF와 하류 CXR 게놈 DNA 프라이머를 보여 게놈 대상 측면과 HXGPRT에 걸쳐 PCR 제품의 존재 (그림이 2A-B) 테스트 정확한 5 '및 3 사용 '삭제 된 유전자 현장에 HXGPRT 유전자의 통합을 목표로.
주 : 검증을위한 프라이머를 얻을 수있는 이익이나 거짓 긍정적 인 결과의 유전자 고유해야합니다. 프라이머 디자인에 유효성을 검사 할 수 http://www.toxodb.org 프라이머가 고유 확인 게놈 (들)에 프라이머 시퀀스를 폭파 있습니다.
- 주간 일정에 통과 기생충 클론은 24 단계의 설명. 대상 삭제가 확인되면, MPA + X에서 계속 선택 사항입니다.
- 냉동 주식을 준비하여 보관 기생충 클론. 펠렛 세포 기생충25cm 2 배양 플라스크에 용해 HFF 세포에서 기생충 농도에서 세포 배양 냉동 매체에 미디어를 제거하고 부드럽게을 Resuspend 기생충 펠릿> 4 × 10 ㎖ 당 7 기생충.에게 액체 질소에 무기한 cryovials 및 저장 기생충에 분주을 전송하거나, -80 ° C.
2. HXGPRT 삭제
이 프로토콜은 유전자 조작 Δ ku80 균주의 게놈에있는 그것의 통합 사이트에서 HXGPRT 마커를 제거하기 위해 설계되었습니다. HXGPRT 제거 마커 복구 및 HXGPRT 1-3에 따라 만 선택하여 여러 유전자 조작과 변형의 생성을 허용합니다. 아래에 설명 된 프로토콜은 다시 대상으로 현장 HXGPRT를 삭제하는 방법을 설명하지만, 그것은 관심의 유전자 현장에서 HXGPRT의 제거는 야생형 유전자 (C의 재 통합을 동시에 허용하는 주목해야한다omplementation), 유전자 조작의 다양한 수있는 돌연변이 유전자의 재 통합뿐만 아니라, 태그 유전자 (N-또는 C-말단 GFP, HA 태그, 등)의 재 통합. 작업 24의 메커니즘과 6 thioxanthine 선택을 사용하여 프로토콜을 대상으로는 이전에 1-3,15을 설명하고있다.
2.1 삭제 HXGPRT
- HXGPRT을 (그림 1B) 측면에서는 독특한 제한 효소 사이트에서 잘라 RE.Z으로 소화하여 pΔGOI에서 소비세 HXGPRT 선택 마커.
- 아가로 오스 겔 전기 영동으로 DNA의 완전한 소화를 확인합니다. 아가로 오스 겔에서 두 개의 밴드의 큰 분리합니다.
참고 : 두 밴드의 큰은 5 '와 3'DNA 목표 측면,하지만 HXGPRT 유전자와 함께 플라스미드를 포함합니다.
- 스핀 열 누운에 큰 DNA 밴드를 포함하는 아가로 오스 섹션을 배치G 멤브레인 젤 평면. RT에서 5 분 동안 13,000 X g에서 열을 회전. 100 μL의 볼륨을 가지고 흐름을 통해에 멸균 H 2 O를 추가합니다.
참고 : 다른 광고 방법은 아가에서 DNA를 분리 할 수 있습니다.
- 18 ㎕의 멸균 H 2 O의 최종 볼륨 DNA를 현탁, 에탄올 침전으로 DNA를 집중
- 7 ML H 2 O, 1 μL 10X 내고 버퍼 1 ㎕의 T4 DNA ligase를 (5 대)를 집중 DNA 1 μL를 혼합하고 4시에 반응을 배치 ° C pΔGOIc (pΔGOIclean) (그림 3A)를 생성 하룻밤.
참고 : RE.Z 포함하는 DNA의 끝 DNA 조각이 아니라 대상으로 돌연변이 유전자의 재 통합 대상 야생형 유전자 (보완)의 재 통합 대상에 적합한 플라스미드를 만들려면이 단계에서 설계 포함 할 수 있습니다 태그 유전자의 재 통합(N-또는 C-말단 GFP, HA 태그 등).
- 이전의 electroporation에 멸균 H 2 O로 반응 배 희석.
- E.에 pΔGOIc 변환 프로토콜 1.1으로 대장균, subclone 분리 및 대상 플라스미드를 확인합니다. 다음 (1.1.26 단계 1.1.11 참조) 형질 전환에 대비 pΔGOIc 선형화.
- 같은 프로토콜 1.2 (단 1-11)에 나와있는 기생충의 transfection 프로토콜을 반복합니다.
참고 : 이전 8 단계를 수행하기 HXGPRT의 대상 제거를 확인하는 돌연변이 균주에 가능합니다. 6 thioxanthine (6TX) 선택 배지 (200 ㎍ / ㎖ 감염 매체 6TX)와 PFU 분석에서 플라스크를 배치를 사용하여 돌연변이 균주 1 X 10 6 tachyzoites에 합류 HFF 세포의 150cm 2 플라스크를 감염. 더 (또는 매우 적은; <10) 있는지 확인 8 일 후 플라스크 검사 설정하는 것이 필수적입니다 PFU는 표시되지 않습니다가, 그 poten재판 유전자 효율성을 타겟팅 감소 HXGPRT 식 (6TX 저항 표현형)과 표현형 변이 균주의 잠재적 자연 복귀를 초과합니다.
주 : 약 5 - HXGPRT 통합 사이트의 10 %가 HXGPRT 마커가 아직 대상 사이트 (설명을 보시려면 대표 결과 섹션 참조)에 통합되어 있더라도 6TX 저항의 의미와 자연 주파수를 나타냅니다.
- 6TX 선택 배지에 150cm 2 플라스크에 매체를 변경하여 6TX 선택하기 ~ 20 시간의 포스트 형질을 시작합니다.
참고 : ~ 10 일이 후 형질 전환을위한 플라스크를 방해하지 마십시오.
- 12 포스트 형질 (유형 I 기생충) 또는 일 - 10 - 16 후 형질 (유형 II 기생충) 일 10시 PFU 형성에 플라스크를 검사합니다.
참고 : Parasi에HXGPRT의 삭제 타겟이되고 TES는 HXGPRT 유전자의 mRNA가 삭제 될 때까지 6TX 선택에서 성장을 시작하지 않으며, 잔류 HXGPRT 단백질이 비활성화됩니다.
- 합류 HFF 세포와 5 ML 6TX 선택 매체를 포함하는 새로운 25cm 2 플라스크에 기생충 솔루션의 1.0 ML - 기생충 솔루션 및 전송 0.5을 만들기 위해 선택 플라스크를 흔들어. 두 가지 이상의 일 동안 하루에 한 번 새 25cm 2 플라스크에 대한 기본 150cm 2의 전송을 반복합니다.
참고 : 다중 샘플링 HXGPRT 유전자를 잃은 가능한 대상 기생충을 캡처의 기회를 증가시킵니다.
- 건강한 복제 기생충의 영역이 PFUs 개발의 존재를 확인하기 위해 2 플라스크 ~ 5 일, 감염 후 25 센티미터 검사합니다. 감염의 영역을 포함하는 하나 또는 두 개의 플라스크를 선택합니다.
참고 : 기생충 6TX 선택의 정상적인 성장 속도로 HXGPRT 유전자 복제의 삭제.
- 6TX 선택 배지에서 통과 기생충을 계속 진행합니다.
- 선택 30 일 - 25 일 이후 기생충 인구 Subclone. 1 ~ 기생충 / 잘 / 잘 ~ 2 기생충과 다른 한 96 잘 트레이를 설정합니다.
- 고립 된 클론 기생충 DNA를 준비하고 (단계 19-29) 프로토콜 1.2에 따라 24 잘 포맷 문화의 클론을 유지한다.
- 그림 3A-B에 설명 된 전략을 사용하여 PCR로 HXGPRT 삭제를 확인합니다.
3. 단백질의 C-말단 태그
이 프로토콜은 MPA를 사용하여 유전자의 게놈 현장에서 상동 재조합을 통해 더블 크로스를 통해 통합을위한 태그를 대상으로 단백질의 C-말단에 태그를 위해 설계 + X 선택 2.4. 이 프로토콜은 효율적으로 작동하기 때문에의 5 'DHFR 순서 HXGPRT 마커는 다른 유전자 2,4에 대한 완전한 검증 기능 3 '비 번역 영역입니다.
내생 유전자 좌위에서 단백질의 3.1 직접 C-말단 태그
- 만들기 pΔGOItag 대상 플라스미드 효모 재조합 복제 (그림 4)와 프로토콜 1.1에 설명 된 방법을 사용하여 구성. 선택의 태그 (HA 태그, Myc와 태그, 그의 태그에 5 '종료 코돈은 3 위치로 이동 제외 게놈 대상 측면은 GOI의 코딩 영역의 마지막 800 1,200 BP (또는 게놈 DNA)를 포함' 등)
- (그림 4)에 설명 된 전략을 사용 프로토콜 1.1 및 1.2 단계를 수행하여 단백질 코딩 유전자의 내생 현장에 C-말단 태그의 타겟 삽입을 만듭니다.
- PCR 전략을 사용하여 내생 유전자 현장에서의 C-말단 태그의 삽입을 확인합니다.
- D에 프로토콜 1, 프로토콜 2에서 설명한 방법을 사용하여 대상을 변경할 유전자 현장태그 단백질을 표현 정확하게 조절 내생 유전자 현장을 만들 수 HXGPRT 선택 마커를 elete. 이 프로토콜은 태그 변형 (프로토콜 1 참조)에서 다른 궤적을 대상으로 다시 사용할 수 있습니다 HXGPRT 선택 마커를 복구합니다.
결과
자세한 템플릿은 제한 효소 사이트의 배치에 HXGPRT의 후속 삭제 유전 표적 유전자의 검증 대상뿐만 아니라, 플라스미드 건설을 촉진 프라이머의 생성을 포함, 유전자를 삭제 대상으로 플라스미드를 구축 제공됩니다 단일 단계 과정 (그림 1A-C, 그림 2A, 그림 3A). 일반 회로도 예를 들면, 목표 유전자 삭제 (그림 2A), 녹아웃의 삭제를 확인하는 데 사용되는 프라이머 쌍을 만들기 위해 제시, 유형 I rop18 (그림 2B) 및 PCR 검증의 대표적인 결과가 표시됩니다 (그림 2C). 이 대표 결과는 대상 상대적으로 어려운 유전자 좌위에서 얻을 수있는 결과의 범위를 설명하기 위해 표시됩니다. 성공적으로 목표 유전자 삭제에서의 유전자의 부재가 발생합니다관심이 늘어 PCR 제품 (PCR 1), (3)의 존재 '게놈 대상 측면 (PCR2), 제대로 5 사이의 통합 HXGPRT 선택 마커의 존재'와 (PCR3와 후 pCR4) 삭제를 정의 3 '게놈 대상 측면 . 클론 3, 4, 5, 6, 8, 9, 11 HXGPRT가 GOI (그림 2C)를 대체했다 타겟 유전자의 삭제를 (녹아웃) 확인됩니다.
유전자의 삭제를 포함하지 않는 클론은 밴딩 다양한 패턴으로 표현 될 수있다. 일반적으로 유전자의 삭제없이 복제는 PCR1 및 PCR2 관찰 밴드 부모의 변형 (부모 패턴)을 반영하지만 클론 1과 2 (그림 2C)를위한 것과 같이 PCR3 또는 후 pCR4합니다.합니다 이 "부모의 패턴은"몇 가지 잠재적 인 메커니즘에서 발생합니다. 때때로, MPA 저항 선택한 기생충 플라스미드 대상 pΔGOI의 비 통합 지속 episomes을 가지고, 우리는 계속 선택은 종종 이러한 episomes에게 강제주의통합 할 수 있습니다. 우리는 또한 내가 Δ는 pΔGOI의 의도 통합으로 인해 ku80 변형 DHFR 5 '와 3'발기인 요소 DHFR 유전자 좌도에 또는 부분적으로 삭제 HXGPRT에 통해 표현 HXGPRT의 cDNA를 포함하는 플라스미드 대상 형식 드문 배경을 준수 현장 14. 이 희귀 한 이벤트가 타겟 통합이지만, 그것은 의도 된 통합 아니었다 어떤 유전자 교체 전략을 설계 기억하는 핵심 역할을합니다. 타겟팅 pΔGOI에 실시 120 bp의보다 큰 DNA 상 동성이 플라스미드 원하지 않는 배경을 돌려 줄 수 Δ ku80 변종 2 재결합 다른 사이트를 제공 할 수 있습니다. HXGPRT 선택 마커 타입 II 형 DNA와 비교할 때 염기 다형성이 나는 순서를 기반으로하기 때문에 유형 II 저기, Δ ku80 변형이 배경이 감소 또는 존재하지 않는 나는을 입력 비교하는크게 유형 II 저기, Δ ku80 변형 1을 사용하여 실험이 드문 배경 줄어 듭니다. 유전자 현장은 (삭제할 수없는) 필수, 또는 현장에서 주파수를 대상으로, 또는 매우 낮은 유전자를 가지고 있다면 경우 episomes 쉽게 성장과 선택을 통해 제거되지 않습니다 【비 통합],이 부모 패턴 MPA에서 관찰 된 패턴을 지배하게 될 것이다 지속 저항 클론. 또한, 표적 DNA 분자는 때때로 단지 5 '또는 3'게놈 대상 측면과 밴드 (하지만 둘 다) 클론 10 같이 PCR1 및 PCR2과 함께 (5 PCR3 또는 후 pCR4 중 하나에 대한 관찰에 통합하는 관찰 '통합) 및 복제 7 (3'통합) (그림 2C). 이러한 패턴은 HXGPRT을 통합 만이 대상으로 통합이 관심의 유전자를 삭제하지 않는 현장에서 대상 플라스미드의 통합을 통해 단일 십자가의 드문 발생을 제안한다. 이 패턴 (레인 7 및 그림 2C 10) EM그 대상 통합 대신 발생 확인하는 PCR 데이터를보고 할 필요가 phasizes 하나는 동종 건너와 표적 분자의 동종 번째 통합. 드물게, 우리는 부모의 패턴 타겟 삭제 패턴을 모두 나타냅니다 클론 12 (그림 2C)로 표현 유전 적 혼합을 준수합니다. 이 패턴은 대부분 잘 복제에서 동일한 위치에 존재하는 두 개의 기생충 유전자형의 경우에 발생합니다.
Δ ku80Δgoi에서 HXGPRT의 제거를위한 일반적인 회로도 :: HXGPRT 긴장에서 HXGPRT을 삭제할 수는 (그림 3A) 표시됩니다. 클론 4, 7, 9, 11, 12 (그림 3C)에서 볼 수 있듯이 Δ ku80 Δ gra2 :: HXGPRT (그림 3B)에서 HXGPRT의 제거의 유효성을 검사하는 데 사용되는 프라이머 쌍은 PCR5에 고유 한 ~ 1.2 KBP 대역을 증폭. HXGPRT는 궤적, 2 ~ 3에서 제거되지 않은 경우.4 KBP 밴드는 (클론 1, 2, 3, 6, 8, 10 및 부모의 통제) 관찰된다. 몇 가지 메커니즘 HXGPRT의 제거 (6TX 선택) 더 도전적이고 HXGPRT의 통합 (MPA는 + X 선택)보다 효율적으로 만들기 위해 행동한다. MPA 선택은 6TX 선택 14,16 단순히 더 효율적입니다. 또한, HXGPRT 표현의 높은 수준은 MPA 선택 16,24보다 6TX 선택을 위해 필요합니다. 따라서, HXGPRT 효소 활동이나 돌연변이 또는 유전자의 현장에서 HXGPRT의 발현을 감소 성적인 변화를 줄일 수 변이는 잠재적 6TX 선택의 효과를 폐지 할 수 있습니다. 심지어 6TX 선택의 이러한 도전, Δ ku80 변종 6TX 선택의 성공률은 1-3의 첫 번째 시도에서 90 %보다 큽니다.
단백질 코딩 유전자의 직접 C-말단 태그에 대한 일반적인 구조는 (그림 4) 표시됩니다. 이방식은 태그 유전자의 HXGPRT 3 '의 상동 재조합을 통해 더블 크로스를 통해 직접 통합을 사용하며 위에서 설명한 것과 유사한 검증 전략을 사용합니다. 유전자는이 플라스미드 독립적으로 격리 대상으로 DNA와 두 번째 형질 전환을 사용하여 확인할 수있다 필수적인 유전자로 의심됩니다. 이러한 규제 유전자 발현에 대한 계획과 같은 다른 방법은 유전자가 25 중요한 경우 추가 검증에 사용할 수 있습니다.

그림 1. 플라스미드 대상 DNA의 설계의 개요. 설계. 도식은 PCR이 pRS416 셔틀 벡터와 HXGPRT의 minigene 카세트에 대한 33 bp의 겹침 5 '와 3'대상 측면을 증폭하는 데 사용되는 프라이머를 중복. 회로도에 나와 뇌관이었다5 '및 3'pΔROP18 10의 목표 측면. 표적 DNA 분자를 설계 B. 전반적인 전략을 생성하는 데 사용됩니다. pΔGOI의 백본 pRS416 셔틀 벡터를 포함하는 우라실 (URA) 및 암피실린 (AMP) 선택 마커입니다. ~ 1 KBP 5입니다 pRS416에 삽입 된 'F1과 R1 프라이머를 사용하여 HXGPRT의 minigene 카세트 및 pRS416에 대한 중복과 게놈 DNA에서 증폭 된 DNA 대상 측면은 1 ~ KBP 3'DNA 대상 측면을위한 중복과 게놈 DNA에서 증폭된다 HXGPRT의 minigene 카세트와 F2와 R2 프라이머 HXGPRT의 minigene 카세트를 사용 pRS416. 다음과 같은 고유 한 제한 효소 소화 사이트는 프라이머에 추가됩니다 F1 입문서는 re.X (제한 효소 X 컷 사이트) R2에 RE.Y, DNA 대상 측면 5 '5의 끝'에서 플라스미드를 잘라 소비세 HXGPRT 분에 R1 및 F2 프라이머의 DNA 대상 측면과 RE.Z 3 '3 끝'에서 플라스미드를 절단하는 프라이머rop18의 삭제 대상 구조를 생성하는 데 사용 igene 카세트입니다. C. 뇌관 시퀀스. 굵은 지역은 T.에 해당 gondii의 게놈 시퀀스와 굵은 글씨 이외의 지역 pRS416 및 HXGPRT의 minigene 카세트와 중복 제한 효소 사이트 및 시퀀스에 해당합니다. HX_F 및 HX_R 프라이머 쌍은 HXGPRT minigene 카세트 14 증폭. 프라이머는 5 ~ '3'읽어보십시오. 펜트 레스 등 10 일부터 적용 표. 큰 그림을 보려면 여기를 클릭하십시오 .
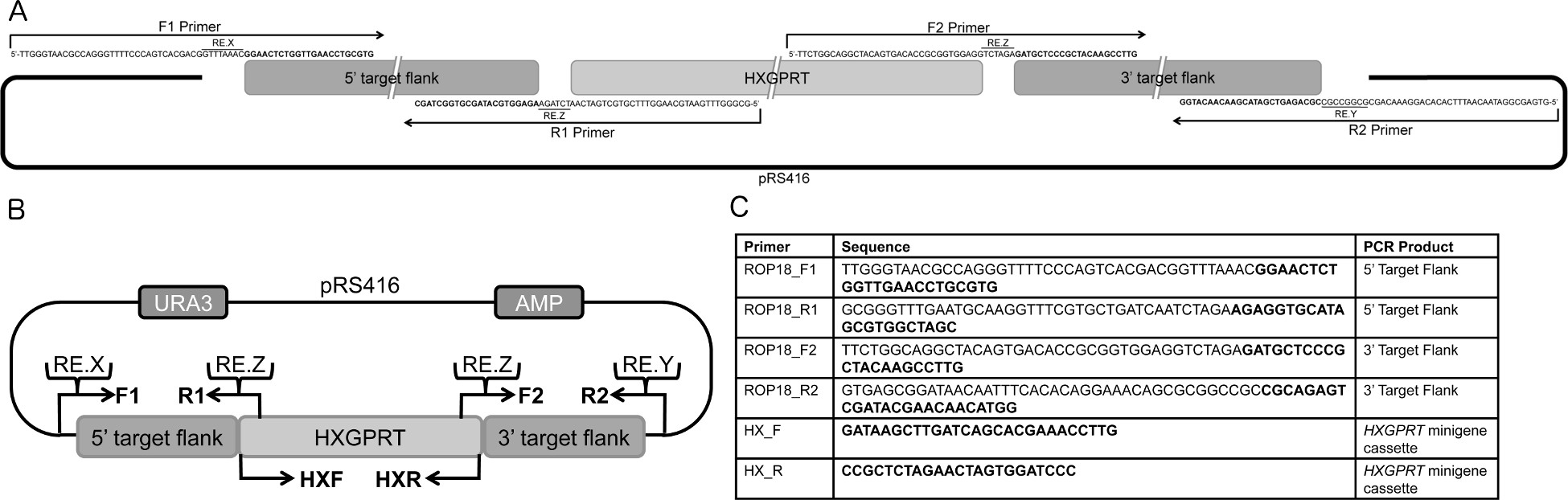{kind=link}

그림 2. HXGPRT를 사용하여 유전자의 삭제에 대한 프로토콜의 개요. . DisruptioN 번 교차하여 Δ ku80Δhxgprt 변형에 GOI ~ 1 KBP 5 'DNA 대상 측면과 ~ 1 KBP 3'플라스미드 pΔGOI에 DNA 대상 측면을 사용하여 상동 재조합 이벤트. PCR의 PCR1를 사용하여 전략 PCR2, PCR3 및 후 pCR4은 (눈금 표시) GOI 궤적의 대상 HXGPRT의 통합 및 삭제를 가진 복제 시스템의 유효성을 검사하는 유전자형 확인을 가능하게하는 표시됩니다. B. 대표 프라이머 쌍은 rop18의 삭제를 검증하기 위해 설계 . ROP18_DF 및 ROP18_DR는 PCR1, ROP18_ExF 및 ROP18_CxR이 PCR2, ROP18_CxF 및 DHFR_CxR이 후 pCR4을 (그림 2A 참조) 증폭 PCR3 및 DHFR_CxF 및 ROP18_CxR을 증폭 증폭 증폭. 프라이머는 5 ~ '3'읽어보십시오. 표 펜트 레스 등의 HXGPRT를 사용하여 목표 유전자 삭제 (rop18)의 검증 10. C. 대표 결과를 적응. MPA의 Δ ku80Δhxgprt 및 선택에 플라스미드 pΔROP18의 다음 형질 +X, MPA + X 저항하는 클론 rop18의 삭제을 분석 하였다. 부모의 변형 제어 PCR 3 (~ 1,200 BP)과 PCR을위한 PCR 1 (~ 400 bp의) 및 PCR 2 (~ 650 bp의) 제품에 대한 긍정적이고 부정적인 4 (~ 1,300 BP) 제품입니다. 대상 GOI 녹아웃은 PCR2, PCR3 및 후 pCR4 제품에 대한 긍정적이며 (그림 2A 참조) PCR 한 제품에 대한 음수입니다. 표시는 PCR1 및 PCR2 (상단 패널)의 결과를 대표 패널, PCR3 (중간 패널) 및 후 pCR4 (아래 패널)입니다. 클론 3, 4, 5, 6, 8, 9, 11은 복제의 타겟 유전자의 삭제 이벤트를 정의 PCR 1, PCR2, PCR3 및 후 pCR4의 정확한 밴딩 패턴을 나타냅니다. 클론 1, 2, 7, 10, 12 유전자 삭제하지 않습니다, 그리고 기타 잠재적 대표 결과에 해당합니다. (C = 부모의 통제 변형 Δ ku80Δhxgprt, M = 크기 마커). 큰 그림을 보려면 여기를 클릭하십시오 .
{kind=link}

그림 3. 관심의 유전자가 HXGPRT를 삭제하려면 다시 대상의 프로토콜의 개요. 더블 크로스 오버로 HXGPRT 선택 마커 제거 변형 Δ ku80Δgoi에서 상동 재조합 이벤트 :: HXGPRT은 1 ~ KBP 5 'DNA 대상 측면과 ~ 1 KBP 3'플라스미드 pΔGOIc에 DNA 대상 측면을 사용 A. 전략. 유전자형 확인을위한 PCR 전략은 (눈금 표시) 삭제를 걸쳐 PCR 제품 (PCR5)에 대한 분석에 프라이머 쌍을 사용하여 묘사된다. B.는 gra2에서 HXGPRT 선택 마커의 제거를 확인하기 위해 설계 담당자 프라이머 쌍 변형 Δ ku80Δgra2의 궤적 :: HXGPRT. 프라이머 GRA2 _CLF 및 GRA2_CxR는 PCR5를 증폭. 프라이머 5 ~ 6TX에 Δ ku80Δgoi :: HXGPRT 및 선택에 플라스미드 pΔGOIc의 다음과 같은 형질을 얻을 수 있습니다 6TX 저항 클론. C. 대표위원회 '3'읽어보십시오. 클론 4, 7, 9, 11, 12 HXGPRT 마커의 대상 삭제 (~ 1.2 KBP 제품 그 측면 '5에 약간의 중복에 측면'과 3 '외부 3에 걸쳐에 해당 PCR5의 정확한 밴딩 패턴을 전시 측면). 클론 1, 2, 3, 6, 8, 10 (이 밴드 빛)이 복제가 허용 6TX에 대한 저항의 정도를 전시에도 불구하고, 부모의 복제 유전자형에 해당하는 3.4 KBP ~ 주위의 예상 줄무늬 패턴을 나타냅니다 자신의 선택 (이 표현형은 가끔 (프로토콜 2.1 (8 단계)에있는 주 및 대표 결과 섹션을 참조하십시오) 셧다운 HXGPRT의 발현에 의해 자발적으로 발생할 수 있습니다. (C = 부모의 통제 변형 Δ ku80Δgoi :: HXGPRT, M = 크기 마커).w.jove.com/files/ftp_upload/50598/50598fig3large.jpg "대상 ="_blank "> 큰 그림을 보려면 여기를 클릭하십시오.

그림 4. 단백질의 C-말단에 태그를 대상으로 DNA 분자의 디자인. 전략은 또한 3 '하류 규제 영역으로 작동 할 수 HXGPRT 마커의 기능을 기반으로합니다. pΔGOItag의 백본 pRS416 셔틀 벡터 포함 우라실 (URA) 및 암피실린 (AMP) 선택 마커입니다. DNA 대상 측면에 대한 중복과 게놈 DNA에서 증폭 ~ 1 KBP 5 'Fgoi 및 Rtag 프라이머 ~ 1 KBP 3 사용 HXGPRT의 minigene 카세트 및 pRS416에 대한 중복과 게놈 DNA에서 증폭 된 DNA 대상 측면'은 pRS416에 삽입 F2와 R2를 사용하여 HXGPRT의 minigene 카세트 및 pRS416프라이머 및 HXGPRT의 minigene 카세트입니다. 5 'DNA 대상 측면은 대체 종료 코돈 다음 태그 (HA, Myc와 그의 등) 종료 코돈을 대체합니다. MPA + X 선택은 C-말단 태그와 하류 HXGPRT을 통합하고, HXGPRT 마커 다음 위치 3 'UTR 이동합니다. 다음과 같은 고유 한 제한 효소 소화 사이트는 프라이머에 추가됩니다 Fgoi 프라이머의는 re.X (제한 효소 X 컷 사이트) DNA 대상 측면 5 '5의 끝'에서 플라스미드를 잘라 RE.Y에 플라스미드 건설에 사용하는 다시 대상 태그 유전자 현장 삭제하려면 HXGPRT의 minigene 카세트 소비세에 Rtag 및 F2 프라이머의 DNA 대상 측면 3 '3 끝'에서 플라스미드 및 RE.Z을 잘라 R2 프라이머 3 'UTR의 위치를 복원합니다 HXGPRT 마커.
토론
여기에서 우리는 대상 유전자 삭제, 유전자 대체, 그리고 / 또는 태그 유전자를 가지고 유전자 조작 자손의 효율적인 복구를 가능하게 Δ ku80 기생충 변종 대상으로 효율적인 유전자에 대한 프로토콜을 제공합니다. 이러한 방법의 순차적 인 실행은 1-3 단일 또는 여러 대상으로 유전자 조작을 포함하는 기생충 돌연변이 체의 분리에 대한 안정적인 접근 방식을 제공합니다. 대상 삭제의 생성이 여러 요인에 따라 다르지만, 제시된 전략 및 프로토콜 Δ ku80 변형의 사용은 크게 T.를 정확히 정의 된 돌연변이 균주를 제작의 효율성과 편의성을 향상 기능 게놈 연구를위한 gondii의.
T.의 게놈 무성 생식 단계에서 gondii의이 반수체 있습니다. 따라서 유전자를 삭제할 계획 할 때 만들 수있는 고려 사항은 유전자가 생체에 필수적인해서는 안 것입니다. 그럼에도 불구하고, 타지의 효율성Δ ku80 변종 팅 유전자 녹아웃은 매우 높고, 이는 크게 손상 복제 속도로 돌연변이 균주의 분리 할 수 있습니다. 대상 유전자가 중요한 경우, 기생충은 종종 선택하는 동안 복제를 중단. 타겟 유전자 조작의 또 다른 중요한 요인은 유전 표적 측면의 적어도 620 BP가 검출 대상 통합 2를 얻기 위해 필요로 완벽한 동성이다. 동성 이상 지역 타겟팅 약 1,000 BP의 DNA의 측면을 대상으로하는 1-4 안정적이고 효율적인 방법입니다 사용하여 높은 효율을 생산하고 있습니다. 이러한 이유로 게놈 대상 측면은 T.의 동일한 균주에서 생성하는 것이 중요합니다 유전자 조작되고 변형 등 포자충 (RH, 저기,). 충분한 유사성의 부족은 종종 하나의 게놈 대상 측면의 대상으로 통합 될,하지만 다른 할 수 없습니다. 완전 통합 또는 녹아웃 엄마를 얻기 위해 실패Y는 에피 솜 지속성으로 인해합니다. 즉시 형질 전환 후, 표적 분자의 모든 episomes를 통합되지 않은 있으며, 때로는 표적 분자는 여러 세대 episomes로 유지할 수 있습니다. 1 ~ KBP 목표 DNA의 측면을 사용하기 전에 다시 서브 클로닝을 선택에서 기생충을 전달하기 위해 계속하는 것은 자주 에피 솜 지속성과 관련된 문제를 해결합니다.
한방의 유효성을 검사하고 HXGPRT 마커가 제거되면, 유전자 표적 전략은 1-3 단일 기생충 긴장에서 여러 유전자의 삭제를 생성하는 두 번째 GOI에 반복 할 수 있습니다. 삭제, 교체 또는 태그 유전자의 생성은 또한 다른 선택 마커 (블레오 마이신, CAT, pyrimethamine 저항 DHFR 등)를 사용하여 수행 할 수 있습니다. CAT 선택 마커가 현재 유형 II Δ ku80 저기, 게놈 1에 존재하지만,이 마커는 HXGPRT를 사용하여 제거 할 수 있습니다 선택 프로토콜은 여기에 설명. 우리 손에, HXGPRT 선택이 단일 복사본을 대상으로 삽입 MPA + X 중간. HXGPRT의 강력한 선택에 대한 충분한 곳 가장 높은 대상으로 효율성을 가장 안전하고 가장 효율적인 마커도의 타겟 삭제 현재 유일하게 유용한 방법을 제공합니다 6 thioxanthine (6TX) 선택 2,3를 사용하여 선택 마커 (그림 3). 따라서이 프로토콜은 여러 타겟 유전자 조작으로 정의 된 돌연변이 균주 개발을위한 현재의 유일한 방법을 제공합니다.
이 프로토콜은 톡소 포자충의 Δ ku80 변종 타겟 유전자 조작을위한 귀중하고 효율적인 방법을 제공하며, 하나의 유전자, 유전자의 가족, 또는 게놈 전체의 기능 게놈 연구의 분석에 적용 할 수 있습니다. 이전 Δ ku80 변종 1,2의 가용성에, 이러한 방식은 오프라인으로 인해 널리 실현되지 않았다이러한 방법 fficiency. 따라서,이 프로토콜은 톡소 포자충의 타겟 유전자 조작 광범위하게 적용하고 기생충 생물학, 호스트 응답 및 기능 유전체학에 대한 질문의 범위를 주소에 관심있는 모든 연구자에 액세스 할 수 있습니다.
공개
저자는 그들이 더 경쟁 재정적 이익이 없다는 것을 선언합니다.
감사의 말
이 작품은 DJB에 NIH에서 교부금 (AI041930, AI073142, AI075931, 및 AI091461)에 의해 지원되었다.
자료
| Name | Company | Catalog Number | Comments |
| Overlap Primers | Integrated DNA Technologies | 4 nmole Ultramer DNA oligos | |
| Validation Primers | Integrated DNA Technologies | 100 nmole DNA Oligo | |
| Yeast Strain #90845 | ATCC | Designation FY834 | |
| Shuttle Vector pRS416 | ATCC | 87521 | |
| DH10B E. coli | Invitrogen | 12033-015 | SOC broth in kit with E. coli |
| Resuspension Buffer | Qiagen | Buffer P1 in QIAprep Spin Miniprep Kit | |
| Miniprep Kit | Qiagen | 27104 | QIAprep Spin Miniprep Kit |
| Glass Beads | Scientific Industries | SI-BG05 | 0.5 mm acid-washed |
| Qiacube Automated Robotic Work Station | Qiagen | ||
| Electroporation Cuvette | USA Scientific | 9104-5050 | 2 mm-gap |
| BTX600 electroporator | BTX | ||
| Maxiprep Kit | Qiagen | 12662 | QIAprep Spin Maxiprep Kit |
| 25 cm2 Canted neck plug seal flask | Corning | 430168 | |
| 150 cm2 Canted Neck plug seal flask | Corning | 430823 | |
| Human Foreskin Fibroblasts (HFF) cells | ATCC | SCRC1041 | |
| Swin-lok Filter Holder | Whatman | 420200 | 25-mm-diameter |
| Membrane | Whatman | 110612 | Nucleopore Track-etched Membrane 3 mm pore size, 25-mm-diameter |
| Mycophenolic acid (MPA) | Sigma | ||
| Xanthine (X) | Sigma | ||
| 96-well Tissue Culture Tray | Corning Costar | ||
| 24-well Tissue Culture Tray | Corning Costar | ||
| MEM Eagle Media | Lonza Biowhittaker | 12-611F | |
| Fetal Bovine Serum | Gibco | 26140-111 | |
| Antibiotic-Antimycotic (Anti-Anti) | Gibco | 15240-062 | |
| Spin Column | Primm Labs | PAE-100 | Easy Clean DNA Extraction Spin Kit |
| T4 DNA Ligase | New England Biolabs | ||
| 6-thioxanthine | Acros Organics | ||
| Tissue DNA Minikit | Qiagen | 51104 | QIAamp DNA Blood Mini Kit |
| Cell Culture Freeze/Recovery Media | Gibco | 126-48-010 | |
| Phosphate Buffered Saline | Hyclone | SH30028.02 | minus calcium, minus magensium |
참고문헌
- Fox, B. A., et al. Type II Toxoplasma gondii KU80 knockout strains enable functional analysis of genes required for cyst development and latent infection. Eukaryot. Cell. 10, 1193-1206 (2011).
- Fox, B. A., Ristuccia, J. G., Gigley, J. P., Bzik, D. J. Efficient gene replacements in Toxoplasma gondii strains deficient for nonhomologous end joining. Eukaryotic Cell. 8, 520-529 (2009).
- Fox, B. A., Bzik, D. J. Avirulent uracil auxotrophs based on disruption of orotidine-5'-monophosphate decarboxylase elicit protective immunity to Toxoplasma gondii. Infection and Immunity. 78, 3744-3752 (2010).
- Hortua Triana, M. A., et al. Biochemical and molecular characterization of the pyrimidine biosynthetic enzyme dihydroorotate dehydrogenase from Toxoplasma gondii. Molecular and Biochemical Parasitology. 184, 71-81 (2012).
- Kim, K., Weiss, L. M. Toxoplasma gondii: the model apicomplexan. Int. J. Parasitol. 34, 423-432 (2004).
- Gajria, B., et al. ToxoDB: an integrated Toxoplasma gondii database resource. Nucleic Acids research. 36, 553-556 (2008).
- Kim, K., Weiss, L. M. Toxoplasma: the next 100 years. Microbes and infection / Institut Pasteur. 10, 978-984 (2008).
- Butcher, B. A., et al. Toxoplasma gondii rhoptry kinase ROP16 activates STAT3 and STAT6 resulting in cytokine inhibition and arginase-1-dependent growth control. PLoS pathogens. 7, e1002236(2011).
- Daher, W., Klages, N., Carlier, M. F., Soldati-Favre, D. Molecular characterization of Toxoplasma gondii formin 3, an actin nucleator dispensable for tachyzoite growth and motility. Eukaryot. Cell. 11, 343-352 (2012).
- Fentress, S. J., et al. Phosphorylation of immunity-related GTPases by a Toxoplasma gondii-secreted kinase promotes macrophage survival and virulence. Cell Host Microbe. 8, 484-495 (2010).
- Musiyenko, A., Majumdar, T., Andrews, J., Adams, B., Barik, S. PRMT1 methylates the single Argonaute of Toxoplasma gondii and is important for the recruitment of Tudor nuclease for target RNA cleavage by antisense guide RNA. Cellular microbiology. 14, 882-901 (2012).
- Straub, K. W., Peng, E. D., Hajagos, B. E., Tyler, J. S., Bradley, P. J. The moving junction protein RON8 facilitates firm attachment and host cell invasion in Toxoplasma gondii. PLos Pathog. 7, e1002007(2011).
- Szatanek, T., et al. Cactin is essential for G1 progression in Toxoplasma gondii. Molecular Microbiology. 84, 566-577 (2012).
- Donald, R. G., Carter, D., Ullman, B., Roos, D. S. Insertional tagging, cloning, and expression of the Toxoplasma gondii hypoxanthine-xanthine-guanine phosphoribosyltransferase gene. Use as a selectable marker for stable transformation. J. Biol. Chem. 271, 14010-14019 (1996).
- Donald, R. G., Roos, D. S. Gene knock-outs and allelic replacements in Toxoplasma gondii: HXGPRT as a selectable marker for hit-and-run mutagenesis. Molecular and Biochemical Parasitology. 91, 295-305 (1998).
- Pfefferkorn, E. R., Borotz, S. E. Toxoplasma gondii: characterization of a mutant resistant to 6-thioxanthine. Exp. Parasitol. 79, 374-382 (1994).
- Gietz, R. D., Schiestl, R. H. Frozen competent yeast cells that can be transformed with high efficiency using the LiAc/SS carrier DNA/PEG method. Nature Protocols. 2, 1-4 (2007).
- Oldenburg, K. R., Vo, K. T., Michaelis, S., Paddon, C. Recombination-mediated PCR-directed plasmid construction in vivo in yeast. Nucleic Acids Res. 25, 451-452 (1997).
- Fox, B. A., Gigley, J. P., Bzik, D. J. Toxoplasma gondii lacks the enzymes required for de novo arginine biosynthesis and arginine starvation triggers cyst formation. Int. J. Parasitol. 34, 323-331 (2004).
- Roos, D. S. Molecular genetic tools for the identification and analysis of drug targets in Toxoplasma gondii. Curr. Top. Microbiol. Immunol. 219, 247-259 (1996).
- Roos, D. S., Donald, R. G., Morrissette, N. S., Moulton, A. L. Molecular tools for genetic dissection of the protozoan parasite Toxoplasma gondii. Methods in Cell Biology. 45, 27-63 (1994).
- Singh, U., Brewer, J. L., Boothroyd, J. C. Genetic analysis of tachyzoite to bradyzoite differentiation mutants in Toxoplasma gondii reveals a hierarchy of gene induction. Molecular Microbiology. 44, 721-733 (2002).
- vanden Hoff, M. J., Moorman, A. F., Lamers, W. H. Electroporation in 'intracellular' buffer increases cell survival. Nucleic Acids Research. 20, 2902(1992).
- Pfefferkorn, E. R., Bzik, D. J., Honsinger, C. P. Toxoplasma gondii: mechanism of the parasitostatic action of 6-thioxanthine. Exp. Parasitol. 99, 235-243 (2001).
- Mital, J., Meissner, M., Soldati, D., Ward, G. E. Conditional expression of Toxoplasma gondii apical membrane antigen-1 (TgAMA1) demonstrates that TgAMA1 plays a critical role in host cell invasion. Mol. Biol. Cell. 16, 4341-4349 (2005).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유