
Method Article
Пробирного CFSE основе для изучения миграции мышиной кожи дендритных клеток в Опорожнение лимфатических узлов во время инфекции с
В этой статье
Резюме
An assay in mice to track cell migration from skin to draining lymph node is described which enables the characterization of skin Dendritic cells mobilized to the lymph node after footpad infection with Bacille Calmette-Guérin.
Аннотация
Дендритные клетки (ДК) являются важными для инициации иммунного ответа, в частности, благодаря их способности приобретать и антигена трансфер до дренажного лимфатического узла (DLN). Мобилизация контроллеров домена к DLN является сложным и остается полностью выяснены во время инфекции. Здесь описано , является использование инновационного, простого анализа , который опирается на флуорохрома 5- и 6-карбоксифлуоресцеина диацетат сукцинимидил эфира (CFSE) отслеживать миграцию контроллеров домена во время подушечку лапы инфекции Mycobacterium Bovis Кальметта-Герена (БЦЖ) в C57BL / 6 мышей. Этот анализ позволяет характеристику кожи DC подгруппах, активно передислоцировать к осушению, подколенной LN в ответ на БЦЖ. Этот протокол берет свое начало из модели BCG, где мигрирующие кожи были контроллеры домена идентифицированного с помощью проточной цитометрии. Анализ любезен к изучению и идентификации контроллеров домена или других клеток, которые домой к подколенной LN после инокуляции микробов, их метаболитов или других воспалительныхстимулов в подушечку лапы, и, следовательно, для изучения факторов, которые регулируют миграцию этих клеток.
Введение
Контроллеры домена локализованы в поверхности тела чувствуют микробы или их продукты, и при этом мобилизовать через лимфатические сосуды к дренажным LN (DLN) 1,2. Это перемещение необходим для транспортировки микробного антигена с DLN и последующего грунтования иммунного ответа на вторгающейся микроба. Действительно, блокада или отказ РС мигрировать из зараженной ткани к DLN приглушает Т-клеточный ответ 3-5. Соответственно, анализы, предназначенные для отслеживания миграции на уровне одной клетки полезны, чтобы помочь приписываем миграционную функцию данного подмножества DC.
Существует большой объем данных по мобилизации кожи ГЦ к DLN. Последние приходится большая часть знаний о миграции постоянного тока от воспаленных или зараженных сайтов 2. Это не удивительно, так как кожа находится большое население и ГЦ является весьма доступной поверхности для проведения экспериментов в лабораторных мышей. Несколько методов, таким образом, были разработаны с целью оценкимиграция мышиных ДК из кожи к DLN 2. FITC покраска кожи на сегодняшний день является наиболее часто используемым подходом. В этом анализе флуорохромом флуоресцеин-5-изотиоцианатом (ФИТЦ) получают в смеси с ацетоном и контактным раздражителем. Этот коктейль применяется к депилированной или бритой кожи и накопление FITC-меченых клеток, в свою очередь, проанализированные в DLN. Миграция также может быть изучен путем введения в кожу с флюорохром-меченый нано- или микрочастицы, что позволяет отслеживать фагоцитов, которые усваивают флуоресцентные частицы в коже. Лимфатические сосуды могут также быть канюлю непосредственно извлечь мигрирующие из контроллеров домена лимфы. Однако это технически сложно, особенно у мышей. Число ГЦ, полученных для последующего анализа также является ограничивающим фактором.
Несмотря на многие предыдущие исследования по миграции кожи DC, остается скудной информации о мобилизации кожи DC до DLN следующего vacciнация с Кальметта-Герена (БЦЖ), живой, ослабленный штамм Mycobacterium Bovis используется для вакцинации против M. туберкулез. BCG засевают в коже, вызывая ограниченную инфекцию, которая вызывает Т типа помощника-клеток 1 ответ. Оба DC субпопуляции, которые мобилизуют к DLN от БЦЖ-инфицированной кожи и факторы, которые регулируют их миграцию в ходе этого процесса до конца не изучены. В свете вышесказанного, простой , но новый метод был разработан где флуорохромом 5- и 6-карбоксифлуоресцеин диацетат сукцинимидил эфира (CFSE) впрыскивается на месте для отслеживания миграции ГЦ в DLN 3. C57BL / 6 вводили в заднюю лапу с инокулята БЦЖ и за день до умерщвления животным вводили в том же подушечку с высокой концентрацией CFSE. Двадцать четыре часа спустя животных умерщвляют и осушение подколенной LN (злотый) удалены и подготовлены для проточной цитометрии. Этот анализ позволяет язьntification клеток , которые мигрируют в течение ночи от подушечку кожи к DLN (рисунок 1).
Кроме того, у мышей ген-мишенью могут быть использованы, чтобы помочь определить факторы, необходимые для клеток для мобилизации в DLN. С помощью этого анализа, авторы настоящего доклада ранее обнаружили , что EpCAM низкая CD11b высокая миграционная кожи контроллеры домена транспорт BCG в DLN в процессе регулируемой интерлейкин-1 рецептора и внутриклеточного адаптера молекулы MyD88 3. Протокол данного CFSE на основе анализа миграции , который берет свое начало из приведенного выше доклада Bollampalli и др. 3 , где проточной цитометрии был использован для обнаружения и анализа мигрирующие кожи контроллеров домена, описано здесь. Преимущества и недостатки данного анализа обсуждаются.
протокол
Примечание: C67BL / 6 мышей использовали для экспериментов, показанных в этой рукописи. Эти животные были дом под конкретного патогена условий в виварии МТК, Каролинского института, Стокгольм, Швеция. Эксперименты на животных были проведены в соответствии с директивами и указаниями Шведского совета по сельскому хозяйству, Шведского агентства по охране животных и Каролинского института. Эксперименты были одобрены Советом по этике Север животных Стокгольм.
1. Подготовка CFSE запасов и хранения
- Приготовьте 5 мМ маточного 5- и 6-карбоксифлуоресцеин диацетат сукцинимидил эфира (CFSE) в диметилсульфоксиде. Vortex. Распределить 100 мкл аликвоты в стерильные 0,5 мл с завинчивающейся пробкой микропробирки. Хранить аликвот при -80 ° С.
2. Мыши
- Используйте врожденный мужские или женские мышей от 8 до 12-недельного возраста. Предпочтительными являются полу и возрасту соответствует животных. Убедитесь, что необходимые животные этические разрешения аре на месте до начала экспериментов.
3. Иммобилизация животных с изофлуран газа наркотизации
- Загрузите блок анестезии с изофлуран. Заполните 10 мл газонепроницаемые стеклянный шприц с изофлуран. Избегайте введения пузырьков воздуха. Соедините шприц и жидкости на входе с помощью прилагаемого трубки и перемещения толкателя вперед, пока жидкость в соединительной трубе не заметно только о том, чтобы войти в испаритель.
ПРИМЕЧАНИЕ: Не храните изофлуран в шприце, когда он не используется. - Обезболить животных с изофлуран в индукционной камере, с воздуха поддерживали на уровне приблизительно 400 - 500 мл / мин и концентрации изофлуран на уровне 3,5%.
ПРИМЕЧАНИЕ: Устройство не будет работать без воздушного потока. - После того, как животные спят, поверните запорный кран на блоке анестезии маршрут изофлуран газа в маске части. Убедитесь, что маска часть не повреждена, чтобы избежать утечки газа. Воздушный поток может поддерживаться на уровне 400 мл / мин, альтернативно снизилась до 200 мл / мин. Снижение концентрации изофлуран до 2,6%.
Примечание: обезболивающий эффект может быть увеличено и / или уменьшена путем регулирования скорости потока.
4. БЦЖ Инъекции и восстановление
- Визуально подтвердить и выполнить тесты, такие как тест щепотку рефлекс пальца ноги, чтобы убедиться, что у мышей обездвижены / спящим на изофлурановым маски кусок перед выполнением инъекции.
- Привить в заднюю лапу, 1 × 10 6 колониеобразующих единиц Mycobacterium Bovis Кальметта-Герена (БЦЖ) в 30 мкл PBS с использованием шприца , снабженного 29 G х ½ дюйма иглы.
Примечание: Генерирование микобактерий запасов кратко описана в первоначальном описании этой процедуры 3, и в существенной подробно в другом месте 6. БЦЖ также легко доступны из коммерческих источников. Авторы рекомендуют BCG быть импульсным или ультразвуком пропускали через шприц, чтобы разрушить комки до его inoculatiна в организм мышей. BCG повторяется здесь как микробные раздражители учитывая его занятость в первоначальном описании этого протокола 3 , но авторы , безусловно , не поощрять испытания других микробных стимулов. - Выполните ту же процедуру, описанную выше, для инъекции PBS в контрольных мышей.
- Монитор животных после инъекции, чтобы подтвердить, что восстановление после анестезии прошла успешно, и что животные могут поддерживать себя на кололись, заднюю лапу.
5. CFSE Инъекции
- Приготовление CFSE для инъекций:
- Оттепель флакон 5 мМ CFSE исходного раствора от -80 ° C. Развести CFSE 10 раз в PBS. Ресуспендируют раствор тщательно перед заполнением шприца в рамках подготовки к инъекции.
- CFSE Injection и восстановление:
- Повторите эту процедуру для обезболивающее животных в разделе 3. Вводят 20 мкл 0,5 мМ CFSE в заднюю лапу, ранее полученного БЦЖ или PBS.
Примечание: инъекция CFSE следует проводить за день до этого (24 ч) плановую дату для жертвоприношения.
- Повторите эту процедуру для обезболивающее животных в разделе 3. Вводят 20 мкл 0,5 мМ CFSE в заднюю лапу, ранее полученного БЦЖ или PBS.
6. Хирургическое удаление подколенного лимфатического узла (злотый)
Примечание: Использование стерилизованных хирургических инструментов и их повторной дезактивации в 70% этаноле рекомендуется на протяжении всего этого шага.
- Усыпить мышей. Спрей животное с 70% -ным этанолом. Зафиксируйте животное в положении лежа на спине, на рассечение борту.
Примечание: Авторы могут рекомендовать эвтаназии через цервикальной дислокации. - Обязательно сделайте вертикальный разрез в задней части бедра. Очистить мышцы и жир с помощью ножниц и пинцета, чтобы разоблачить и, следовательно, иссечь злотый, расположенные глубоко в жировой мешочек, в подколенной ямке.
- Перенести Pln в 0,5 мл стерильного PBS, на льду.
7. Генерация одноклеточных подвески из злотый
- Гомогенизируйте злотый через 70 мкм ячейки фильтра НОАКе изд в 6-луночный планшет, содержащий-5 мл PBS, или флуоресцентной сортировкой активированных клеток (FACS) буфер (PBS, содержащий 2% фетальной телячьей сыворотки (FCS), 5 мМ этилендиаминтетрауксусной кислоты (ЭДТА) и 1 мМ азида натрия). Перемешать через сетчатый фильтр, используя задний конец 3 мл шприца.
Примечание: Азид натрия токсичен и следует обращаться с осторожностью. - Промыть фильтр грубой очистки с еще 5 мл PBS или FACS буфером, собирают материал в 15 мл пробирку и осаждения клеток при 277 мкг (1200 оборотов в минуту, R = 172 мм) в течение 5 мин, 4 ° С. В качестве альтернативы, гомогенизируют pLNs непосредственно в микропробирок с использованием полипропилена гомогенизаторы.
- В зависимости от размера гранул, ресуспендируют злотый суспензии в соответствующем объеме PBS или буфера FACS, например, 200 - 1000 мкл. Использование счетнокамерное для количественного определения общего числа клеток, полученной из каждой суспензии LN. Используйте трипановым синим, чтобы исключить мертвые / умирающие клетки.
8. Ячейка Окрашивание и проточной цитометрии
- Трansfer 1 - 10 × 10 6 клеток в 5 мл с круглым дном полистирола трубки. Промывают FACS буфером, и окатышей центрифугированием при 277 х г (1200 оборотов в минуту, R = 172 мм) в течение 5 мин, 4 ° С.
- Удалите супернатант и ресуспендируют осадок в 50 - 100 мкл коктейля антител (см дендритных клеток [DC] маркеры используются для поверхностного окрашивания, Материалы таблицы) , содержащий 0,5 - 1 мкг анти-мыши CD16 / CD32 (блокировать неспецифическую Fc-опосредованной взаимодействия) в FACS буфере в течение 30 - 45 мин, на льду. Защита образцов от света.
- После инкубации промыть клетки с FACS буфером и окатышей центрифугированием при 277 х г (1200 оборотов в минуту, R = 172 мм) в течение 5 мин, 4 ° С.
- Ресуспендируют клеток в 50 - 100 мкл буфера FACS. Приобретать данные на проточном цитометре 7-10.
Примечание: Для получения превосходных обзоров по проточной цитометрии мы отсылаем читателя к следующим публикациям, которые передают теоретические и практические сведения об этом методе.
Результаты
Анализа миграции CFSE на основе (рис 1) использовали вместе с проточной цитометрии для обнаружения и характеризуют CFSE-меченых клеток , которые появляются в злотый после БЦЖ - инфекции в подушечку. Большая часть маркировки CFSE в PLn находится на MHC-II высоко и CD11c + / низкий клетки (рис 2A), в соответствии с ГЦ кожи, которые мигрировали к коже DLN 11. И частота и общее количество CFSE-меченого, высокая CD11c MHC-II + / низкие клетки увеличиваются в pLNs от БЦЖ-инфицированных мышей по сравнению с PBS-инъецированных контрольной группы (рис 2B-C), предполагая , что БЦЖ усиливает перемещение этих клетки к DLN. Поскольку это меры анализа миграции в течение периода 24-часовой, можно было установить , что контроллеры домена кожи по - прежнему входя в DLN 5 дней после заражения (рис 2C). Это расширяет миграцию постоянного тока в текущей модели далеко за пределы шкалы времени для первоначального activatiна антиген-специфических CD4 + Т - клеток в PLn 3. Кроме того, миграционные контроллеры домена кожи выражают повышенные уровни со-стимулирующими молекулами CD80 и CD86 (Рисунок 2D). Это согласуется с предыдущими наблюдениями из эксплантов кожи культур и FITC кожи картины 11, поддерживающей концепцию , что мигрирующие контроллеры домена активируются клетки. Важно отметить, что как LN-резиденты РС (MHC-II + CD11c высокие клетки) и плазмоцитов контроллеры домена (MHC-II с низким CD11c низким PDCA 1-+ клеток), которые входят воспаленные LNS через высокие венулах и не афферентные лимфатической 12, где отрицательный для CFSE (рис 2EF), предполагая , что на самом деле CFSE маркировка произошло в основном в подушечку.
Учитывая , что высокая CD11c MHC-II + / низкие клетки представляют собой не один , а несколько DC субпопуляции 13, дополнительные поверхностные маркеры CD103, EpCam (CD326) и CD11b были включены в панель с использованием меченых антител , используемых для обнаружения проточной цитометрии (Материалы таблицы) для дальнейшей характеристики популяции мигрирующих РС (Рисунок 3). При этом, к югу от населения EpCAM низких высоких клеток CD11b был идентифицирован как основной миграционный кожи DC подмножество в ответ на БЦЖ - инфекции (рисунок 3).
Данные цитометрии потока, показанные здесь, был приобретен на поток цитометр оснащен 4-мя лазерами (488, 532, 640 и 405 нм). Данные были проанализированы с помощью программного обеспечения для анализа клеток. Другие цитометры многоцветной потока , чем указано в таблице материалы могут быть использованы для успешного обнаружения маркеров , указанных в приведенных выше исследованиях, или выявления других маркеров, на других типах клеток по этому вопросу. Для протокола, описанного здесь, цитометр способен обнаруживать 6 различных флуорохромии является neceмируется необходимый. Важно то , что эти клоны для каждого антитела , используемые указаны (Материалы Таблица). Выбор флюорохром конъюгата следует рассматривать, как это может зависеть от лазеров и каналов обнаружения на проточном цитометре доступны. Аналогичным образом, антитела должны подбираться для определения наиболее подходящих разведений для окрашивания.
Частоты народонаселения в BCG- и PBS инъецировали образцы рентгенографического и используются вместе с общим количеством клеток для вычисления абсолютного числа определенных, закрытых подмножеств контроллеров домена. Достоверность различий в группе данных средств анализировали с помощью Студента го испытания или ANOVA, где это уместно, с отсечкой р <0,05. Выпадающие были исключены из анализа следующие испытания Граббса s для недопустимое.

Рисунок 1. Схема: БЦЖ подушечку инфекцииМодель и CFSE-основе анализа миграции. БЦЖ засевают в заднюю лапу от мышей C57BL / 6. В разные моменты времени после инфицирования, и 24 ч до умерщвления, флуорохромом CFSE вводят в той же подушечку, ранее полученной БЦЖ. Осушение, подколенных лимфатических узлов выделяют и CFSE-меченых клеток , характеризующиеся проточной цитометрии. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
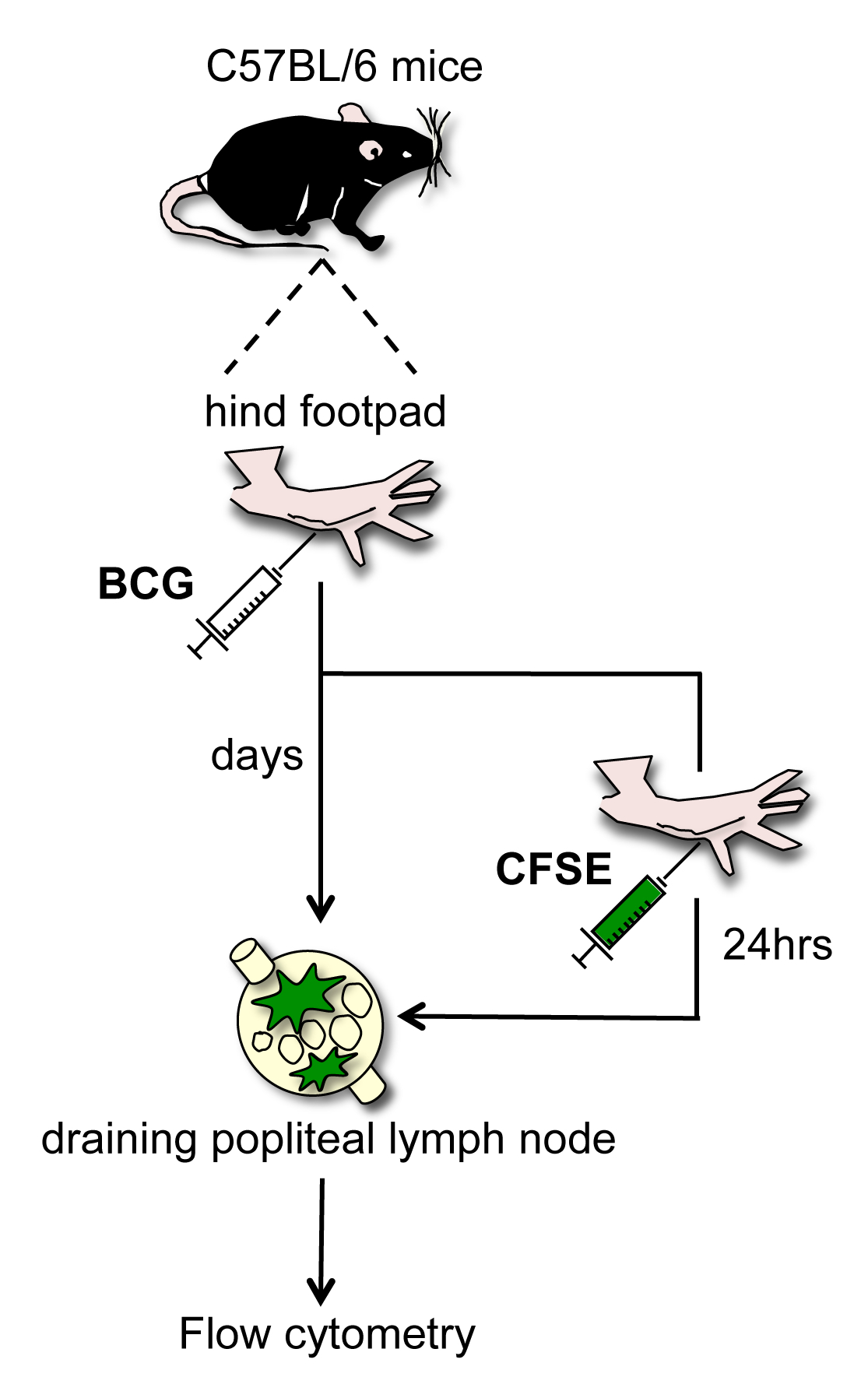{kind=link}

Рисунок 2. Миграционные контроллеры домена кожи являются большинство населения Переселение к DLN После БЦЖ подушечку инфекции. C57BL / 6 мышей инфицировали 1 × 10 6 КОЕ БЦЖ в подушечку. За двадцать четыре часа до умерщвления, животным вводили 0,5 мМ CFSE в том же подушечку. Подколенной LNS собирали, гомогенизировалив одноклеточных суспензий и подвергали проточной цитометрии. Для измерений, выполненных через 1 день после БЦЖ, CFSE вводили 2ч после БЦЖ. (А и Б) Зебра участки , показывающие доминирующую экспрессию CFSE на MHCII высоких и CD11c + / низких клеток в БЦЖ-осушение Pln через 3 дня после заражения. (C) Частота и общее количество CFSE-меченого MHCII высокой CD11c + / низкие клетки в зл из BCG- и PBS-инъецированных мышей рентгенографического в различные моменты времени. (D) средней интенсивности флуоресценции (MFI) для CD80 и CD86 определяли на CFSE + и CFSE нег высокой CD11c MHC-II + / низкие кожи в DCs злотый 3 дня после БЦЖ - инфекции. (Высокая CD11c MHC-II + / низкий) (E) CFSE выражение в ГЦ кожи, LN-резидент РС (MHC-II + CD11c высокий) и (F) плазмоцитов РС (MHC-II с низким CD11c низким PDCA-1 +)определяли с помощью проточной цитометрии через 3 дня после БЦЖ. Для каждого эксперимента, по меньшей мере, 5 мышей использовали для БЦЖ-инфицированных групп и 3 для ПБС-инъецированных контролей. Полоски показывают стандартную ошибку среднего значения. Общее количество CFSE-меченых клеток внутри каждой подгруппы приведены в Bollampalli и др, PLoS Pathog 2015, 11:. E1005206 3. * Обозначает статистически значимых различий между БЦЖ-инфицированных и PBS контрольной группы. Один из двух независимых экспериментов, показанных. Рисунок адаптирован из Bollampalli и др, PLoS Pathog 2015, 11:... E1005206 3, с разрешения издателя Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
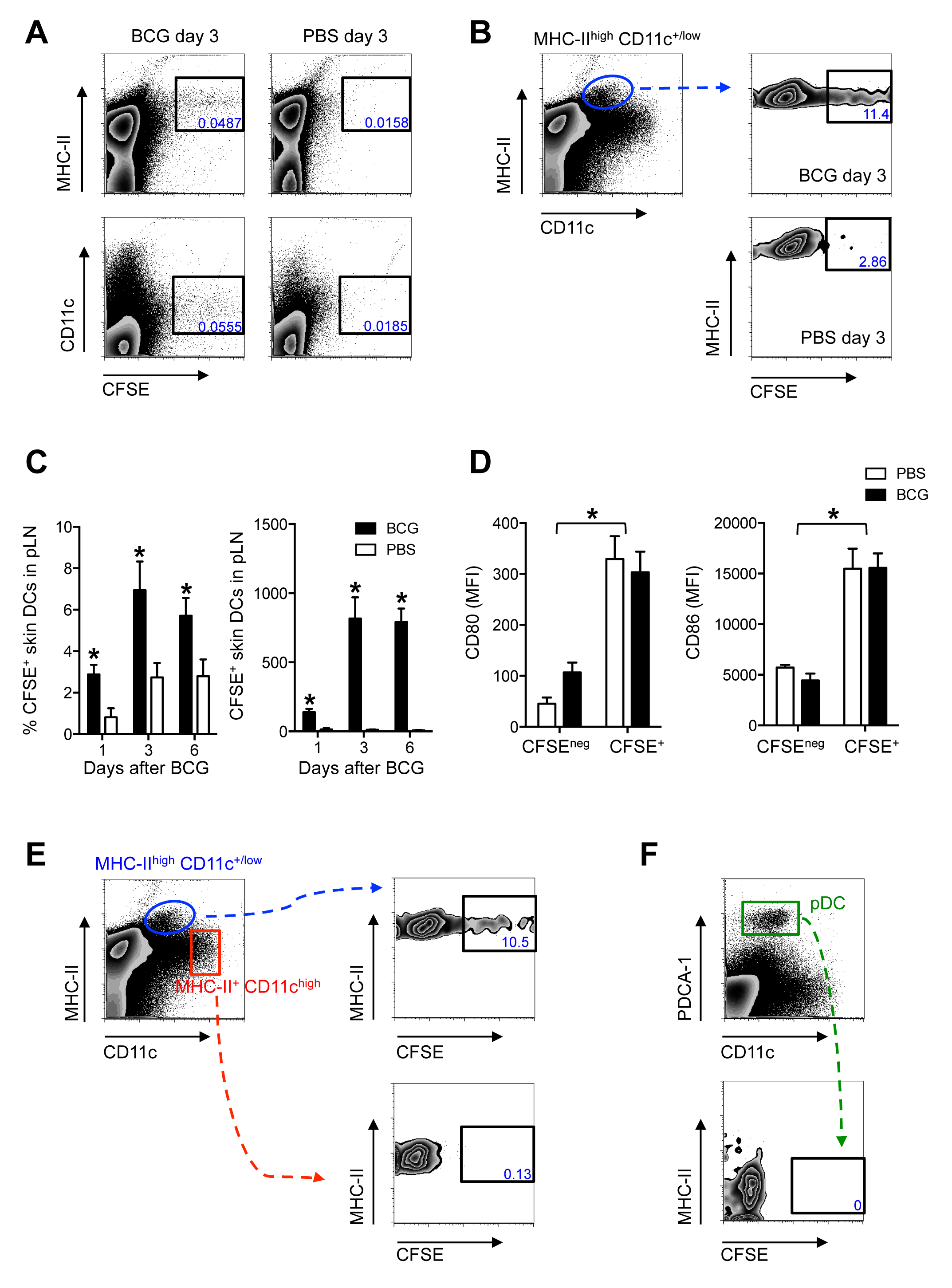{kind=link}

Рисунок 3. EpCAM низкий CD11b высокие контроллеры домена являются основной кожи DC Subset TraffickING к DLN После БЦЖ подушечку инфекции. C57BL / 6 мышам вводили BCG и CFSE , как на рисунке 2. (А) Zebra участки , показывающий стробирования стратегию использовавшимися (верхняя панель) и частоту CFSE + клеток внутри каждого, определенное подмножество в разные моменты времени после БЦЖ-инфекции (нижних панелей). Частоты в пределах каждого подмножества могут варьироваться в зависимости от момента времени после заражения. После 3 -х дней БЦЖ можно ожидать , чтобы найти около 0,2% высокого CD11c MHC-II + / низкие клетки среди всех клеток LN. Из них примерно 6% CD103 + клетки. Для подмножеств , найденных в пределах высоких CD103 отрицательного ворота MHC-II, можно найти около 42% CD11b высокой EpCam низкие клетки, 27% CD11b низким EpCam низкие клетки и 7% ЛЦ. (Б) Всего числа CFSE + клеток внутри каждой определенной подгруппы в различные моменты времени после БЦЖ - инфекции показывается. Для каждого experimeнт, по меньшей мере, 5 мышей использовали для БЦЖ-инфицированных групп и 3 в контрольной группе PBS. Полоски показывают стандартную ошибку среднего значения. * Обозначает статистически значимые различия по сравнению с PBS-инъецированных контрольной группы. Один из трех независимых экспериментов, показанных. Рисунок перепечатали из Bollampalli и др, PLoS Pathog 2015, 11:.. E1005206 3, с разрешения издателя Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
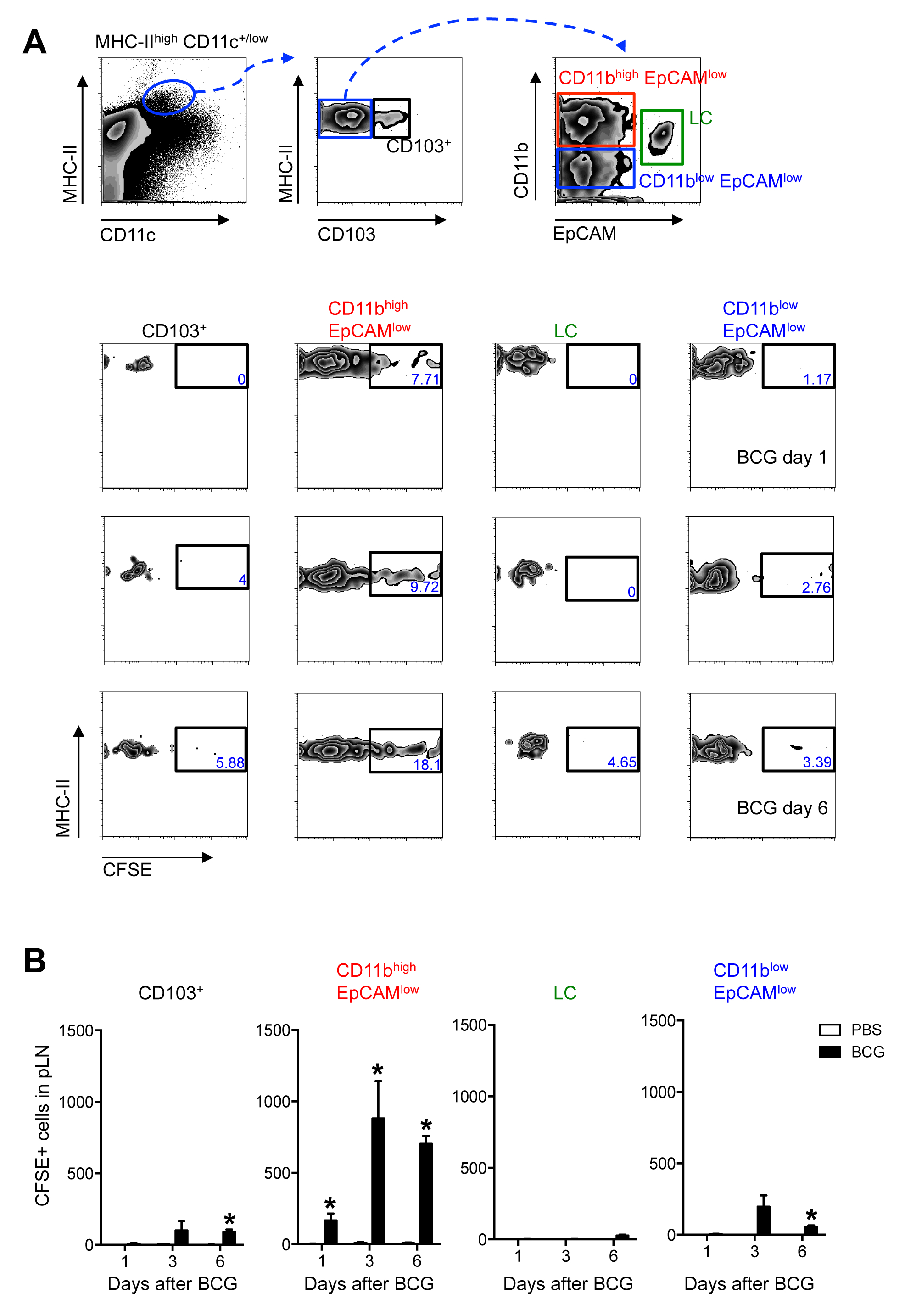{kind=link}
Обсуждение
НЦОД являются специализированными антиген-представляющих клеток, которые могут реагировать на микробную вызов на поверхности тела путем приобретения микробного антигена и миграции на DLN через лимфатическую систему. Там, контроллеры домена представить этот антиген Т-клеток способом, который приводит в реакции первичных Т-клеток. Процесс миграции DC от ткани до DLN Поэтому очень важно для понимания того, как инициируется продуктивный Т-клеточный ответ. Методы измерения миграции постоянного тока, следовательно, очень полезны в разворачивание выше.
Мышиная модель была использована для изучения кожи DC миграцию к DLN после заражения БЦЖ, живой вакцины, которая впрыскивается клинически в коже. В строке, БЦЖ вводится в заднюю лапу, чтобы имитировать этот кожный маршрут вакцинации. Простой анализ был разработан и, следовательно, включены в эту модель для изучения миграции кожи DC подгруппах из прививка БЦЖ сайта в подушечку кожи к дренажным злотый. Этот протокол RELIЕ. С. по инжекции Флуорохром CFSE в ту же лапу, что ранее полученного БЦЖ, а затем выделяют водосточные Pln 24 ч позже для анализа присутствия CFSE-меченых клеток с помощью проточной цитометрии. Хотя это и не описано в протоколе, LNS из этой установки также могут быть получены для анализа с помощью конфокальной микроскопии, для изучения процессов миграции , таких как позиционирование миграционных клеток в LN 3. Кроме того, аналогичные результаты по БЦЖ-триггерным кожи миграции постоянного тока были получены с использованием обоих BCG Пастера 1173P2 3 и БЦЖ SSI 1331 14.
CFSE обычно используется для обозначения клеток в пробирке и отслеживать такие меченых клеток в естественных условиях после клеточных передач 15. В текущем протоколе , хотя, он был использован непосредственно маркировать клетки кожи на месте. Молекулярные основы маркировки CFSE подобна FITC, который также амин-реактивным флуоресцентный краситель. CFSE Однако предпочтительно над FITC для конъюгации, как это создАтес очень стабильные карбоксамидных облигации 16. Кроме того, CFSE сильно мембрана проницаема и пассивно диффундирует в клетки. Оказавшись внутри, он образует внутриклеточное амин (флуоресцентные) конъюгаты, сохраняющиеся в меченую клетку 15.
CFSE на основе анализа миграции, разработанная авторами позволяет идентифицировать клетки, перемещающих от кожи до DLN в течение 24-ч в ответ на БЦЖ. Таким образом, он измеряет острую миграцию в течение определенного периода времени и позволяет исследовать способность различных, впрыскивается стимулы, чтобы вызвать миграцию. Этот анализ отличается от классической методики FITC живописи кожи, который измеряет кумулятивный событие миграции, спровоцированные кожное, местного применения контактного сенсибилизирующее агента. Таким образом, анализа миграции CFSE на основе могут быть использованы для идентификации мигрирующие кожи или DCs другие клетки, которые домой к Pln после инъекции микробов, микробные продукты или другие воспалительные стимулы в диафрагменноеootpad. Действительно, оба нейтрофилы и γ: d Т - клетки , было показано , что переселиться из кожи DLN в ответ на БЦЖ 17,18. На самом деле, анализ недавно был использован в условиях ко-инфекции , чтобы показать , что кишка локализованы нематодозы может приглушить БЦЖ-триггерным кожи миграции постоянного тока к DLN 14.
Миграция DC часто оценивается от 24 до 72 ч в FITC живописи кожи , так как FITC-меченого контроллеры домена трудно обнаружить от 4 до 5 дней после покраски 19. Потеря сигнала CFSE на меченой РС не является проблемой в анализе, представленном здесь, так как анализ миграции был ограничен до 24 часов; причина этого в том, что авторы хотели специально изучать «овернайт» миграционные события. Другие заинтересованные в этом анализе, однако, рекомендуется изучить более ранние или даже более поздние моменты времени для отслеживания CFSE основе. К тому же, так как CFSE вводят путем инъекции может быть дано в любое заданное время. Такая гибкость позволила Authors для оценки миграции постоянного тока между 5 -й день и 6 после БЦЖ - инфекции (рис 2C) 3. Инъекционное CFSE потенциально может ограничить типы клеток , доступные на месте по отношению к реакции мечения. Например клеток Лангерганса (LCS), которые являются контроллеры домена , которые находятся в верхнем слое кожи, эпидермис, мигрировать плохо в ответ на БЦЖ анализа миграции CFSE основе (рисунок 3) 3. Тем не менее, во время кожи картины FITC, контроллеры домена , расположенных в дерме, слой под эпидермисом, легко маркировать и мигрируют к DLN быстрее , чем LCs 20,21. Это говорит о том, что клетки могут стать меченное флуорохромом не обязательно необходимости локализовать к слою кожи, где применяется раствор маркировки.
В качестве модели для БЦЖ - инфекции, ухо мыши обеспечивает добросовестное внутрикожный прививки , который является более в соответствии с клинического применения БЦЖ в качестве вакцины туберкулеза. Foотпад инъекции с другой стороны, это, вероятно, сочетание внутрикожного и подкожный пути. Тем не менее, это требует навыков и подготовки , чтобы правильно и воспроизводимо доставить БЦЖ в дерму 22, так что в клинической практике это не всегда может быть дано по- настоящему внутрикожное , а скорее подкожное или комбинацией обоих. В качестве сайта инокуляции подушечку имеет два очень явные преимущества над ухом, которые заслуживают упоминания. Во-первых, иммунный ответ сосредоточен на дренажного Pln следующей инъекции в подушечку лапы и это облегчает изучение ответов. Авторы этого доклада нашли злотый быть первым и основным LN сливе подушечку 3. Другие предложили раскол дренаж между подколенной и LNs после того, как расположенный под подвздошной костью инъекции Эван синий 23. Тем не менее, в отношении уха, есть более чем одно ухо DLN у мышей , и они не могут быть регулярно присутствовать или легко определить местонахождение 24. Последнее явно вводит liability в исследованиях, стремящихся расследовать ответы в DLN. Во-вторых, большие объемы могут быть привиты в подушечку лапки (10 - 50 мкл) по сравнению с ухом. Это, в свою очередь, сводит к минимуму как ответственность введения основных изменений в лимфоток и того, чтобы работать с очень малыми объемами впрыска, что само по себе является проблемой, когда речь идет о посеве большого количества микобактерий. В ухе, где объем вводимого препарата должно быть небольшим (1-10 мкл), даже в 5 мкл может изменять лимфатического потока и потенциально заставить большее количество впрыскиваемого материала непосредственно в DLN бесконтрольно. Поэтому важно учитывать объемов закачки. Это особенно важно в экспериментах, касающихся вклада клеток в перегона бактерии к LN. В модели подушечку лапы БЦЖ, некоторые БЦЖ, возможно, достиг дина в отсутствии клеточного транспорта. Это основано на наблюдении, что можно обнаружить еще БЦЖ в DLN мыши, где миграцию клеток от стопынакладка была полностью удалена с помощью местной инъекции токсина коклюша 3. Является ли это указывает на то, что БЦЖ может непосредственно получить доступ к лимфатические сосуды и достигают дина в истинно бесклеточной способом остается определить, так как оно не может быть исключено, что микобактерии в данном случае, когда предоставляется доступ к лимфатическим сосудам силой Вводимый объем.
Практическое рассмотрение для вакцинаций в ухе или подушечку в том, что животные должны быть надлежащим образом иммобилизованные для инъекций, чтобы быть выполнена правильно, и воспроизводимым образом. Isoflurane газа описан в текущем протоколе в качестве способа сдерживать животных в рамках подготовки к подушечку инъекций. Относительно быстрая индукция и время восстановления для изофлурановой-опосредованной анестезии делают его популярным методом, но по аналогии с другими анестетиками, это влияет на физиологию, которая может помешать с экспериментальными результатами 25. Действительно, как летучие и инъекционные анестетики уменьшают лимфоток, упразднивпроизвольные движения мышц, снижение мышечного тонуса и уменьшение лимфангиона сжатия 26. Кроме того, сокращение в 5 раз в миграции к DLN ДК адаптивно переносили в подушечку было сообщено в наркотизированных животных 27. С учетом вышеизложенного, и поскольку процедуры обработки до начала анестезии, вероятно, вызовет больше напряжения в организме животных, чем сама инъекция, которая является одновременно быстрым для администрирования и не требует стадии восстановления, возможность адекватно сдерживать животное без наркоза должен быть на рассмотрении. Удерживающим мышь настроена для подушечку инъекции, где животное может быть быстро доведена до иммобилизованной положение и инокулировали в подушечку лапы в пределах от 30 до 40 сек будет идеальным впрыснуть мышь в заднюю лапу, не изменяя другие аспекты его физиологии в процессе , и было бы чрезвычайно полезным при изучении адресации миграции клеток через лимфатическую систему.
В заключение, в настоящем докладевыдвигает на первый план новый протокол для отслеживания контроллеров домена кожи во время их мобилизации в DLN с использованием анализа, который контролирует миграцию в течение определенного окна 24-часовой. Эта CFSE на основе анализа миграции позволяет обнаруживать и анализировать мигрирующие клетки с помощью проточной цитометрии, как подробно описано здесь, а также с помощью конфокальной микроскопии 3. Он обеспечивает гибкую платформу для изучения разнообразных инжектированных раздражителей и их способность не вызывают миграцию постоянного тока, и, что важно, может быть использован для отслеживания не только РС, но и другие группы населения, перемещающихся с кожи к DLN во время различных экспериментальных установок.
Раскрытие информации
The authors declare that they have no competing financial interests.
Благодарности
This work was supported by Karolinska Institutet, a project grant from the Swedish Research Council VR (Dnr 2014-2794) and a KID Doctoral Grant from Karolinska Institutet (All to A.G.R). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Материалы
| Name | Company | Catalog Number | Comments |
| Isoflurane | Baxter | 1001936060 | |
| 5- and 6-carboxyfluorescein diacetate succinimidyl ester (CFSE) | Invitrogen | C1157 | |
| Dimethyl sulphoxide (DMSO) | Sigma | D2650 | |
| Fetal bovine serum (FBS) | Sigma | F7524 | |
| Bacille Calmette-Guérin (BCG) | strain Pasteur, in house | ||
| Trypan blue solution | Sigma | T8154 | |
| EDTA | In house | ||
| Sodium azide | Sigma | S2002 | |
| PBS | In house | ||
| Filtered water | In house | ||
| BD Plastipak 3 ml syringe | BD Medical Surgical Systems | 309658 | alternative: 5 ml syringe |
| 6 well plates | TPP | 92006 | |
| 15 ml centrifuge tubes | TPP | 91015 | Polypropylene |
| 5 ml round bottom tubes | BD Falcon | 3272948 | Polystyrene |
| Univentor 400 anesthesia unit | Univentor | 8323001 | |
| Monojet 3/10 ml syringe | Medicarrier | 8881511144 | 29 G x 1/2 in. |
| Sterile filter unit | TPP | 99500 | PES membrane |
| EPPI-Pistill homogenizer | Schuett biotech | 3.200 512 | Polypropylene |
| Allegra X-30R centrifuge | Beckman Coulter | B01145 | |
| Bürker counting chamber | VWR | 631-0920 | |
| 0.5 ml screw cap microtube | Sarstedt | 72.730 | Polypropylene |
| 1.5 ml Eppendorf microcentrifuge tube | VWR | 700-5239 | Polypropylene |
| 70 micron cell strainers | VWR | 734-0003 | |
| BD LSR-II flow cytometer, operated with FACSDiva software | BD Biosciences | Discontinued. Other Flow cytometers available, minimum detection of 6 fluorochromes needed | |
| FlowJo cell analysis software | Tree Star | ||
| Antibody | Clone | Company | |
| MHC-II I-A/I-E | M5/114.15.2 | BD Biosciences | |
| CD11b | M1/70 | BD Biosciences | |
| CD11c | HL3 | BD Biosciences | |
| CD16/CD32 (Fc-block) | 2.4G2 | BD Biosciences | |
| CD326/EpCAM | G8.8 | Biolegend | |
| CD103 | 2E7 | Biolegend | |
| CD80 | 16-10A1 | Biolegend | |
| CD86 | GL-1 | Biolegend |
Ссылки
- Platt, A. M., Randolph, G. J. Dendritic cell migration through the lymphatic vasculature to lymph nodes. Adv Immunol. 120, 51-68 (2013).
- Randolph, G. J., Angeli, V., Swartz, M. A. Dendritic-cell trafficking to lymph nodes through lymphatic vessels. Nat Rev Immunol. 5, 617-628 (2005).
- Bollampalli, V. P., et al. BCG Skin Infection Triggers IL-1R-MyD88-Dependent Migration of EpCAMlow CD11bhigh Skin Dendritic cells to Draining Lymph Node During CD4+ T-Cell Priming. PLoS Pathog. 11, e1005206 (2015).
- Allan, R. S., et al. Migratory dendritic cells transfer antigen to a lymph node-resident dendritic cell population for efficient CTL priming. Immunity. 25, 153-162 (2006).
- Itano, A. A., et al. Distinct dendritic cell populations sequentially present antigen to CD4 T cells and stimulate different aspects of cell-mediated immunity. Immunity. 19, 47-57 (2003).
- Larsen, M. H., Biermann, K., Jacobs, W. R. Laboratory maintenance of Mycobacterium tuberculosis. Curr Protoc Microbiol. Chapter 10, Unit 10A 11 (2007).
- Tung, J. W., et al. Modern flow cytometry: a practical approach. Clin Lab Med. 27, 453-468 (2007).
- Perfetto, S. P., Chattopadhyay, P. K., Roederer, M. Seventeen-colour flow cytometry: unravelling the immune system. Nat Rev Immunol. 4, 648-655 (2004).
- Sharrow, S. O. Overview of flow cytometry. Curr Protoc Immunol. Chapter 5, Unit 5.1 (2002).
- Holmes, K., Lantz, L. M., Fowlkes, B. J., Schmid, I., Giorgi, J. V. Preparation of cells and reagents for flow cytometry. Curr Protoc Immunol. Chapter 5, Unit 5.3 (2001).
- Henri, S., et al. The dendritic cell populations of mouse lymph nodes. J Immunol. 167, 741-748 (2001).
- Diacovo, T. G., Blasius, A. L., Mak, T. W., Cella, M., Colonna, M. Adhesive mechanisms governing interferon-producing cell recruitment into lymph nodes. J Exp Med. 202, 687-696 (2005).
- Henri, S., et al. Disentangling the complexity of the skin dendritic cell network. Immunol Cell Biol. 88, 366-375 (2010).
- Obieglo, K., et al. Chronic Gastrointestinal Nematode Infection Mutes Immune Responses to Mycobacterial Infection Distal to the Gut. J Immunol. 196 (5), (2016).
- Parish, C. R., Glidden, M. H., Quah, B. J., Warren, H. S. Use of the intracellular fluorescent dye CFSE to monitor lymphocyte migration and proliferation. Curr Protoc Immunol. Chapter 4, Unit4.9 (2009).
- Banks, P. R., Paquette, D. M. Comparison of three common amine reactive fluorescent probes used for conjugation to biomolecules by capillary zone electrophoresis. Bioconjug Chem. 6, 447-458 (1995).
- Nakamizo, S., et al. Dermal Vgamma4(+) gammadelta T cells possess a migratory potency to the draining lymph nodes and modulate CD8(+) T-cell activity through TNF-alpha production. J Invest Dermatol. 135, 1007-1015 (2015).
- Abadie, V., et al. Neutrophils rapidly migrate via lymphatics after Mycobacterium bovis BCG intradermal vaccination and shuttle live bacilli to the draining lymph nodes. Blood. 106, 1843-1850 (2005).
- Thomas, W. R., Edwards, A. J., Watkins, M. C., Asherson, G. L. Distribution of immunogenic cells after painting with the contact sensitizers fluorescein isothiocyanate and oxazolone. Different sensitizers form immunogenic complexes with different cell populations. Immunology. 39, 21-27 (1980).
- Shklovskaya, E., Roediger, B., Fazekasde St Groth, B. Epidermal and dermal dendritic cells display differential activation and migratory behavior while sharing the ability to stimulate CD4+ T cell proliferation in vivo. J Immunol. 181, 418-430 (2008).
- Kissenpfennig, A., et al. Dynamics and function of Langerhans cells in vivo: dermal dendritic cells colonize lymph node areas distinct from slower migrating Langerhans cells. Immunity. 22, 643-654 (2005).
- Kim, Y. C., Jarrahian, C., Zehrung, D., Mitragotri, S., Prausnitz, M. R. Delivery systems for intradermal vaccination. Curr Top Microbiol Immunol. 351, 77-112 (2012).
- Harrell, M. I., Iritani, B. M., Ruddell, A. Lymph node mapping in the mouse. J Immunol Methods. 332, 170-174 (2008).
- Van den Broeck, W., Derore, A., Simoens, P. Anatomy and nomenclature of murine lymph nodes: Descriptive study and nomenclatory standardization in BALB/cAnNCrl mice. J Immunol Methods. 312, 12-19 (2006).
- Gargiulo, S., et al. Mice anesthesia, analgesia, and care, Part I: anesthetic considerations in preclinical research. ILAR J. 53, E55-E69 (2012).
- Schmid-Schonbein, G. W. Microlymphatics and lymph flow. Physiol Rev. 70, 987-1028 (1990).
- Tal, O., et al. DC mobilization from the skin requires docking to immobilized CCL21 on lymphatic endothelium and intralymphatic crawling. J Exp Med. 208, 2141-2153 (2011).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены