
Method Article
Изображение иммунологического синапса человека
* Эти авторы внесли равный вклад
В этой статье
Резюме
В этом протоколе образуется как иммунологическое синапсовое образование, так и последующий поляризованный секреторный трафик в сторону иммунологического синапса. Сотовые конъюгаты были сформированы между суперантиген-пульсированной клеткой Раджи (действуя как антиген-представляя клетка) и клоном Jurkat (действуя в качестве эффектора-помощника Т-лимфоцитов).
Аннотация
Целью метода является создание иммунологического синапса (ИС), пример ассоциирования клеток к клетке, образованной антигенообразующей клеткой (APC) и эффективным помощником T-лимфоцитов (Т) и записи изображений, соответствующих первым стадиям Формирование ИГ и последующие события, связанные с торговлей людьми (происходящие как в БТР, так и в ячейке Th). Эти события в конечном итоге приведут к поляризованной секреции в ИГ. В этом протоколе, клетки Jurkat оспаривается с стафилококкэн энтеротоксин Е (SEE)-импульсные клетки Раджи, как модель синапса клетки был использован, из-за близости этой экспериментальной системы к биологической реальности (Th cell-APC синаптических конъюгированных). Представленный здесь подход включает в себя спряжение от клеток к клетке, приобретение покадровой промежутки, широкополевую флуоресценционную микроскопию (WFFM), за которым следует обработка изображений (после приобретения деконволюции). Это улучшает соотношение сигнала к шуму (SNR) изображений, улучшает временное разрешение, позволяет синхронизировать приобретение нескольких фторхромов в возникающих синапических конъюгированных и уменьшает отбеливание флуоресценции. Кроме того, протокол хорошо сочетается с протоколами фиксации конечных ячеек (параформальдегид, ацетон или метанол), что позволит дополнительно оформить и анализировать иммунофлуоресценцию. Этот протокол также совместим с лазерной сканирующей конфокальной микроскопией (LSCM) и другими самыми современными методами микроскопии. В качестве основного предостережения, только те T-ячейки-APC границ (так называемые интерфейсы IS), которые были на правом 90 "угол фокус плоскости вдоль оси может быть правильно отображаются и проанализированы. Существуют и другие экспериментальные модели, которые упрощают визуализацию в измерении и следующий анализ изображений, но эти подходы не эмулируют сложную нерегулярную поверхность БТР и могут способствовать нефизиологическому взаимодействию в ИС. Таким образом, экспериментальный подход, используемый здесь, подходит для воспроизведения и противостояния некоторым биологическим сложностям, возникающим в ИГ.
Введение
Основной целью метода является создание иммунологических синапсов (ИС) конъюгированных клеток к клетке, образованной антиген-представляя клеткой (APC), импульсной с суперантигенами SEE и эффектором Th-клеток, и регистрировать изображения, соответствующие первым стадиям формирования иммунологического синапса и последующих событий, связанных с торговлей людьми (происходящих как в APC, так и в Т.К.), что в конечном итоге приведет к поляризации. Создание IS Т-лимфоцитов при связывании их клеточного рецептора (TCR) к антигенам, связанным с MHC-II на APC организует чрезвычайно динамичный, податливый и критический экземпляр, участвующий в антиген-специфических, гуморальных и клеточных иммунных реакциях1,2. IS определяется формированием специального супрамолекулярного активированного комплекса (SMAC) шаблон характеризуется актина процесса реорганизации3. После строительства ИГ Т-лимфоцитами с БТР, поляризация секреторных пузырьков по отношению к ИГ, по-видимому, замешана в поляризованной секреции в синаптической пропасти. Этот сфокусированный механизм, как представляется, недвусмысленно поставлять иммунную систему с великолепно регулируется хитрость для повышения эффективности критических секреторных эффектор роли Т-лимфоцитов, при одновременном снижении неспецифических, цитокино-арбитражная стимуляция клеток-прохожих, убийство нерелевантных клеток-мишеней и апоптотического самоубийства через активацию индуцированной смерти клеток (AICD)4.
Последствия IS меняя на природе и T лимфоцитов и APC. Синаптический контакт Т-клеток (обычно CD4" клеток) с APC отображения антигена, связанных с MHC-II производит активацию Т-клеток (цитокин секреция, пролиферация и т.д.) и, в некоторых случаях, апоптоз через AICD4. Для цитотоксических Т-лимфоцитов (CTL) (главным образом клеток CD8), взаимодействующих с APC, представляющим антиген, связанный с MHC-I, результат отличается от предварительной стимуляции или нет CTLs с антигеном. Таким образом, наивные CtL, идентифицирующие комплексы антигена-MHC-I на БТР, «прикораются» для уничтожения клеток-мишеней и разделения. Primed CTLs также установить синапсы с клетками-мишенями (т.е., клетки, инфицированные вирусами или опухолевыми клетками) производя антиген-специфические выращения клеток5,6.
Поляризованная секреция экзосом в иммунологическом синапсе является развивающейся и сложной областью исследований, участвующих в соответствующих иммунных реакциях7. Было продемонстрировано, что многовезикулярные тела (MVB) с внутрилюмичными пузырьками (ILVs) опыт поляризованной транспортировки к IS8,9 ( Видео1) на стимуляции TCR с антигеном. Слияние этих MVB на синаптической мембране индуцирует их дегранизацию и высвобождение ILVs как экзосомы к синаптической расщелине8,10. Это происходит в IS формируется Th-типа Jurkat клеток, которые были оспорены с SEE суперантигена покрытием Раджи клетки, действующие в качестве APC11, TCR-стимулировали CD4и лимфобласты, и загрунтованные CTL. Таким образом, синапсы, изготовленные клетками Jurkat, представляют собой ценную модель для изучения поляризованного секреторного движения экзосом. Кроме того, несколько десятилетий исследования показали, что многие фундаментальные идеи в TCR сигнализации пришли из исследований с преобразованными Т-клеточных линий, и действительно наиболее известным из этих моделей систем является Jurkat лейкемии Т-клеточной линии12.
Формирование полностью развитой IS производит несколько важнейших биологических результатов, в том числе активации Т-клеток, активации наивных CTLs или убийство целевых клеток загрунтованные CTLs, кроме анергии или AICD5. Таким образом, Есть два основных типа секреторных IS создана Т-лимфоцитов, которые приводят к очень разнообразны, но так же критически, иммунный эффектор функции1,6,13. С одной стороны, ИГ из загрунтованное цитотоксические Т-лимфоциты (КТЛ) вызывает быструю поляризацию (от секунд до нескольких минут) литических гранул (называемых "секретными лизосомыми") по отношению к ИГ. Дегрануляция литические гранулы индуцирует секрецию перфорина и гранзимков в синаптической расщелине14, которые являются про-апоптотические молекулы. Выделяемые перфорин и гранзимы впоследствии вызывают убийство целевых ячеек15,16. CTL развиваются временные синапсы, продолжительностью всего несколько минут, так как целевые ячейки убивают3,17. Это, вероятно, связано с обстоятельством, что оптимальная задача CTL требует быстрого и временного контакта для того, чтобы распределить как можно больше смертельных ударов по многочисленным ячейкам3,17. В отличие от этого, Т лимфоцитов, таких как клетки Jurkat генерировать стабильный, давний IS (от 10-30 мин до часов), так как это, как представляется, необходимо как для направленного и непрекращающейся секреции стимулирования цитокинов3,17. Цитокины также заключены в секреторные пузырьки, и некоторые из них (напротив, ИЛ-2, ИФН-З) испытывают поляризованный транспорт в ИС17 и секрецию. Одной из основных характеристик ИГ является формирование поисковых, слабых и переходных контактов между Т-ячейкой и БТР(видео 1),которые могут привести к более сильному взаимодействию и созданию зрелого ИГ, при условии, что ТКР определяет коньячные комплексы антиген-МХК и что установлены соответствующие костимулирующие соединения5. И начало первоначальных контактов, и основание зрелого, полностью продуктивного ИГ, по своей сути стохастические, быстрые и асинхронные процессы5,18. Кроме того, существует скудная частота в создании клеток к клетке конъюгирует19, которые могут представлять собой проблему для методов визуализации (пожалуйста, обратитесь к результатам и обсуждения разделов).
Еще одна основная проблема при изучении поляризации микротубуле-организационного центра (MTOC) и секреторных гранул в Т-лимфоцитах заключается в том, что весь процесс происходит быстро (секунд до нескольких минут), особенно в КТЛ. Учитывая эти факты, большинство ранних подходов сталкиваются с конечная точка стратегии, в которой APC / целевых клеток и Т-лимфоцитов смешиваются совместно и сходятся низкой скорости центрифугирования, в пользу клетки к клетке сопряженного создания, инкубируется в течение нескольких минут, фиксированной и впоследствии оценивается для передислокация MTOC и / или секреторные пузырьки к IS20. Этот подход имеет два существенных ограничения: никаких живых данных о торговле людьми не было достигнуто и высокий уровень фоновой MTOC / секреторные гранулы поляризации были получены, вероятно, из-за стохасического характера создания ИГ18. Кроме того, любая корреляция между TCR-стимулировали, первоначальные сигнальные события (т.е. внутриклеточного кальция поднимается, актин реорганизации) и секреторной поляризации везикул является проблематичным для исследования. Таким образом, императивные положения для надлежащей визуализации ИС в живых клетках объединяются для повышения конъюгированной материализации от клетки к клетке, синхронизации генерации ИС и, по возможности, для обеспечения создания клеточных конъюгированных веществ на определенных полях микроскопа XY и позициях. Для того чтобы избежать всех этих проблем, было разработано несколько стратегий. Вне рамок настоящего документа объясняется эти методы, их преимущества и недостатки. Пожалуйста, обратитесь к ранее опубликованным обзорам, которые касаются этих важных пунктов1,4,5,21.
Тот факт, что IS, сделанные Th лимфоцитов являются долгоживущими, и то обстоятельство, что в Th лимфоцитов MTOC, лимфокин-содержащих секреторных гранул и MVB занять от нескольких минут до часов, чтобы двигаться и доск IS22 делает Th-APC является идеальным кандидатом для визуализации с помощью протокола, описанного здесь.
протокол
1. Подготовка слайдов для соблюдения Raji клетки
- Добавьте 150 л фибронектина (100 мкг/мл) на скважину к 8-микроколодцу камерной горки (пластиковая нижняя камера слайд) и инкубировать его в течение 30 мин до 1 ч при 37 градусах Цельсия. Этот субстрат адгезии позволит привязывать клетки Раджи к дну скважины (Шаг 2), образование живых конъюгированных с клетками Jurkat (Шаг 4) и захват микроскопии замедленного периода (шаг 6), а также совместим впоследствии с факультативной параформадгидом (PFA) фиксацией (шаг 7).
ПРИМЕЧАНИЕ: Для фиксации ацетона используйте стеклянные слайды нижней камеры и поли-Л-лизин (20 мкг/мл) вместо фибронектина, так как ацетон растворяет пластик. 8 микровелл камеры слайд (1 см2 хорошо, 300 л максимального объема) или эквивалент является гибким и соответствующим форматом. - Аспир фибронектин с помощью 200 л автоматической пипетки и мыть каждый колодец с 200 Л ПБС в течение 2 минут с нежной встряхивания. Повторите эту стирку еще раз. Камера слайд может храниться на данном этапе с PBS в течение 1-2 недель при 4 градусах Цельсия.
2. Прилипание клеток Раджи к камерным слайдам и 7-амино-4-хлоретилкумарин (CMAC) Маркировка
- Передача 10 мл конфлюента (1-2 х 106 ячеек/мл) предварительной культуры клеток Раджи в 15 мл, V-нижняя трубка. Хорошо перемешайте и используйте 10 кЛ для подсчета клеток на камере Нойбауэр или эквиваленте.
- Центрифуги оставшиеся клетки на 300 х г в течение 5 мин при комнатной температуре. Аспирируй и отбрасывай супернатант.
- Аккуратно приостановите действие клеточных гранул в теплой полной культуре среды (RPMI 1640 дополнен 10% FCS, 2 мМ глутамин, 10 мМ HEPES, 100 U/mL пенициллин, и 100 мкг /mL streptomycin) в концентрации 106 клеток / мл. Используйте приведенное ниже уравнение:
(начальныйх Vначальный , -окончательный х Vfinal), гдеинициал представляет начальную концентрацию ячейки, Vначальный объем подвески клетки, -окончательный - окончательная концентрация ячейки, Vокончательный том подвески клетки.
ПРИМЕЧАНИЕ: В зависимости от концентрации клеток начальной культуры, можно собрать больше клеток, чем необходимо, но важно поддерживать оставшиеся клетки в культуре (37 градусов по Цельсию) до конца эксперимента, чтобы предотвратить потенциальные проблемы (см. Шаг 2.6). - Этикетка Raji клетки, чтобы их идентификация во время синаптической конъюгации формирования. В этом эксперименте маркировка 7-амино-4-хлоретитилкумарина (CMAC) выполняется в шаге 2.4.2.
- Перенесите необходимое количество клеток Раджи в среде культуры в трубку 2 мл. Для 8 микроколодца камерной горки необходимо в общей сложности 1,6 мл клеточной подвески (200 л на скважину).
- Добавьте CMAC к конечной концентрации 10 мкм. Держите клетки в темноте, покрывая трубку алюминиевой фольгой, так как CMAC светочувствительный. 200 кЛ, содержащие 2 x 105 Раджи клетки необходимы на 1 см2 хорошо. Таким образом, если 8 скважин должны быть подготовлены, 1,6 х 106 Раджи клетки не требуется.
ПРИМЕЧАНИЕ: Маркировка клеток Raji с синим синяя клетка (CMAC, УФ возбуждение и синие выбросы) отличает их от Т-клеток, когда синаптические конъюнты образуются. Этот краситель совместим с PFA и ацетон фиксативов и позволяет дальнейшие процедуры иммунофлуоресценции. Старайтесь избегать легкой экспозиции. Маркировка клеток Раджи в бассейне с CMAC с последующей повторной подвеской перед присоединением клеток Раджи к фибронектин-покрытым камерными слайдами обеспечивает однородную маркировку клеток Раджи CMAC между различными скважинами.
- Приготовьте CMAC-окрашенные клетки и, после аспирации PBS в камере слайд от шага 1.2., передача 200 злилок подвески к каждой скважине фибронектина покрытием камеры слайды подготовлены в шаге 1.1-1.2. Инкубировать камерную горку при 37 градусах Цельсия, 5% CO2 на 30 мин-1 ч.
ПРИМЕЧАНИЕ: На этом этапе одновременно происходит стегея и маркировка CMAC, что экономит время. Пожалуйста, имейте в виду, что клетки Раджи будет осадок быстро и осторожность должна быть принята для поддержания однородной концентрации в клеточной подвески перед посевом. На данном этапе не обязательно мыть ЦМАКП, так как мытье CMAC будет легче осуществляться на шаг 2.7 (когда помеченные клетки Раджи уже прилипают к камерным слайдам). Так как CMAC присутствует в суспензии клетки в большом избытке, синий фон флуоресценции слишком высок, чтобы различать сине-окрашенные клетки. Проверьте ячейку CMAC флуоресценции в шаге 2.7 после мытья CMAC. - Убедитесь, что клетки Раджи прилипают к нижней части колодцев нежным встряхиванием камерных горок на микроскопе. Убедитесь, что ячейки отображают зазоры между собой и не являются сливаться(рисунок 1,средняя панель). 50-60% клеточного слияния является целесообразным.
- Если большинство клеток эффективно придерживаются камерной горки и не наблюдаются клеточные зазоры, промойте каждую скважину теплой полной средой и переостановите среду с 200-м м л автоматической трубой, чтобы отсоединить избыток клеток. Проверьте слияние после каждого шага отсрочки.
- Если клетки не придерживаются, повторите шаг слипа снова и увеличьте время адгезии и/или число клеток.
ПРИМЕЧАНИЕ: Здесь можно остановиться, инкубировать камерную горку при 37 градусах Цельсия, 5% CO2 за ночь (O/N), и продолжить с протоколом на следующий день. Пожалуйста, подтвердите на следующий день, что клетки Raji остаются прилипшими и CMAC-маркированные с помощью флуоресценционной микроскопии.
- Вымойте каждый хорошо снова тщательно с теплой дополненной RPMI для устранения избыточного CMAC и проверить на голубое излучение с флуоресцентным микроскопом (Рисунок 1).
ПРИМЕЧАНИЕ: Чтобы избежать использования масла погружения (липкое и вязкое) и высокочисленных целей масла диафрагмы, цели экстра-длинней (т.е. 20x или 40x) цели могут быть использованы для быстрой проверки маркировки CMAC с помощью флуоресцентного микроскопа.
3. Пульс CMAC - Приклеенные этикетку клетки Раджи со стафилококковым энтеротоксином E
- Добавить стафилококковый энтеротоксин Е (SEE, 1 мкг/мл) к каждой скважине. SEE можно удобно разбавлять в среде культуры клетки (рабочее решение на 100 мкг/мл) от замороженных запасов SEE (1 мг/мл в PBS). Используйте 2 злитроподнина 100-x рабочего раствора на 200 микроскважин.
ПРЕДЕКТО: Используйте перчатки для этого шага и распоряжаться использованный наконечник в коробку биоопасности. - Инкубировать камерную горку при 37 градусах Цельсия, 5% CO2 не менее 30 мин. Эффект SEE длится не менее 3-4 ч.
ПРИМЕЧАНИЕ: SEE может быть добавлен к скважинам в разные моменты времени, когда это необходимо, если запланированы различные установки замедленного замедленного заключения (шаг 5) и/или в зависимости от точки начала для экспериментов конечной точки (шаг 6).
4. Подготовка ячеек джурката
- Используйте для этого эксперимента ранее растущую культуру клеток jurkat (1-2 x 106 клеток/мл). Используйте клетки из стандартной культуры колбу или из предыдущего трансфекции следующие стандартные протоколы электропорации, как ранее описано23. Трансфекция клеток Jurkat позволит промежуток времени визуализации трафика секреторных гранул в живых клетках. Например, когда GFP-CD63 (маркер MVB) выражается движение GFP-CD63-украшенные пузырьки могут быть записаны (Видео 1).
- Наблюдайте за клетками под фазовик-контрастным микроскопом. При превышении мертвых клеток (20-30%) наблюдаются, выполняют Филколл плотности градиент центрифугирования с использованием стандартных протоколов24, чтобы устранить избыток мертвых клеток (мертвые клетки обладают более высокой плотностью, чем живые клетки) до использования (см. Обсуждение).
- Перенесите клетки в трубку 15 мл, V-нижнюю трубку и используйте 10 л для подсчета с помощью гемоцитометра.
- Центрифуги остальные клетки, как описано в шаге 2.2. Откажитесь от супернатанта и отрепримите клетки втой же концентрации, что и клетки Раджи (1 х 10 6/мл), используя свежую, теплую культурную среду. Выполните шаги 2.2-2.3.
- Поддерживайте клетки Jurkat в культуре (37 градусов по Цельсию, 5% CO2)в ожидании Шага 4.
ПРИМЕЧАНИЕ: Во втором варианте (трансфекция) количество живых клеток будет значительно ниже, чем в первом. Таким образом, рассмотрите возможность использования более высокого объема стартовой культуры клеток, чтобы иметь достаточное количество ячеек для эксперимента. Из 10 х 106 юркат ныхсоцитов на кювет и трансфекцию, только 2-4 х 106 юркатных клеток выживут после 48 ч трансфекции, и некоторые из этих клеток будут потеряны во время шага Фиколла. Таким образом, одного квета электропорации, как правило, достаточно, чтобы бросить вызов присоединенным СЭЭ-импульсных клеток Раджи из 8 микроскважин (1,6 х 106 трансфицированных клеток Jurkat необходимо).
5. Совместное посев ячеек Раджи и Джуркат
- Возьмите камеры слайды, содержащие CMAC-помечены, SEE-пульсированные, придерживаются Raji клетки из инкубатора от шага 3.2. На данном этапе нет необходимости мыть CMAC, так как это было ранее сделано в шаге 2.7.
- Тщательно аспирируйте культурную среду каждой скважины, один за 1, из одного угла колодца, используя автоматический пипетку 200 л. Не позволяйте среде в хорошо высохнуть полностью.
- Немедленно замените среду 200 qL перекрышенных клеток Jurkatв среде культуры клетки (1 x 10 6/mL) подготовленных в шаге 4.5. Если время промежуток изображения выполняется, перейдите на шаг 6 сразу после этого шага, так как клетки Jurkat, как правило, осадок и образуют синаптические конъюгирует очень быстро. Для удобства, микровеллы, содержащие SEE-импульсные, придерживались клетки Раджи, которые не получают посева с клетками Jurkat на данном этапе должны поддерживаться с средой клеточной культуры до последующего вызова с клетками Jurkat. Это гибко позволит последующему испытанию с клетками Jurkat для дополнительного промежуток времени или обратного кинетической, конечных экспериментальных подходов.
- Если промежуток времени будет выполнен, быстро перейти к шагу 6. Это включает в себя со-культуру для 1-2 ч на микроскоп этапе инкубатора или эквивалент на 37 КС, 5% CO2, чтобы синаптические конъюгированные формирования и одновременного приобретения изображения. Если анализ конечных точек, но не промежуток времени, предусмотрено проверка конъюгированного образования после периода совместной культуры с помощью микроскопа (как на рисунке 1) перед фиксацией клеток (шаг 7).
6. Время Lapse Изображение новых синаптические конъюгирует
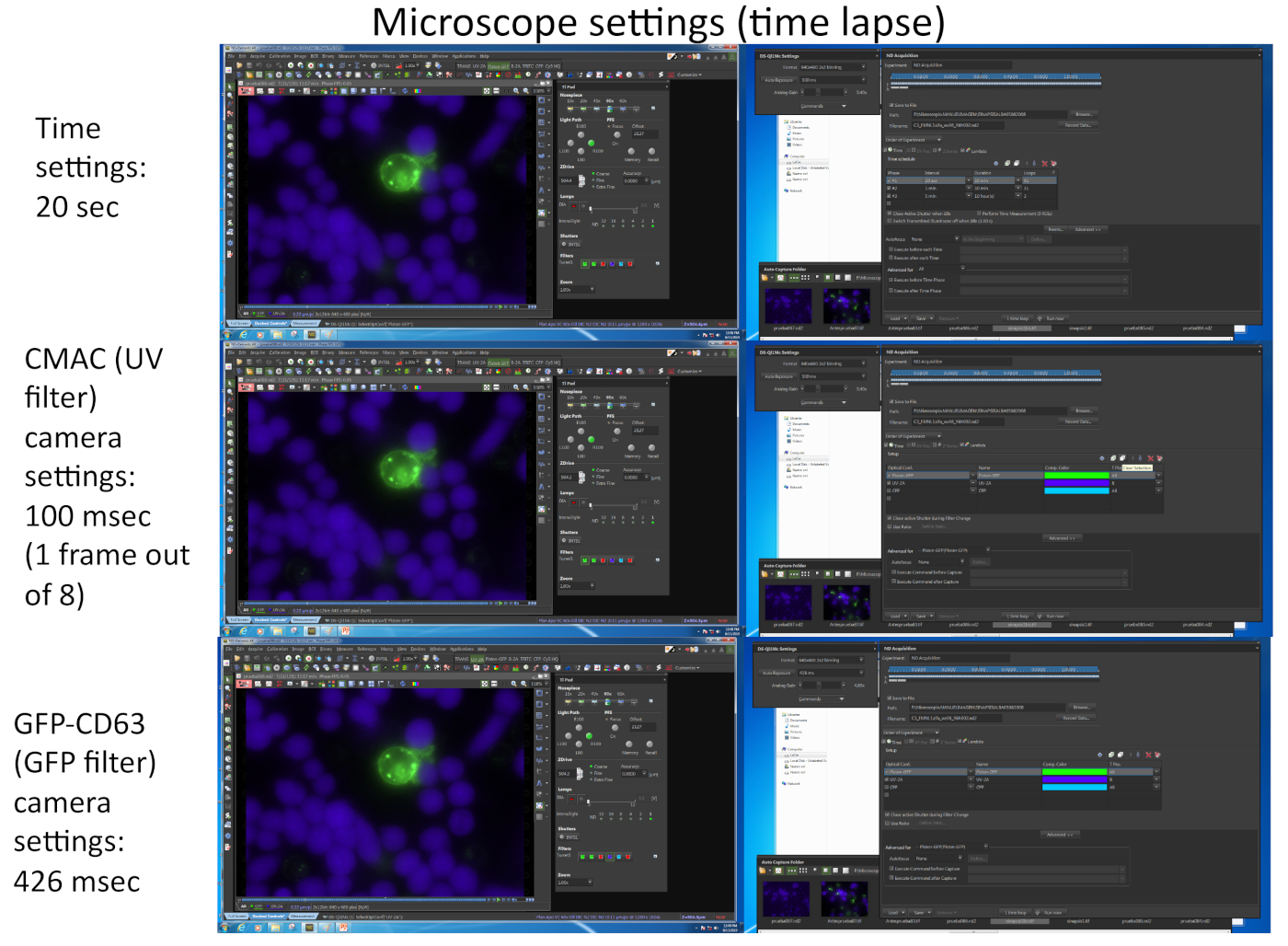
- Подготовьте микроскоп и инкубационную камеру перед визуализацией. На примере, показанном в видео 1, подробные настройки микроскопии отображаются на рисунке 2.
ПРИМЕЧАНИЕ: Если запланирован эксперимент по промежуток времени, все настройки и дополнения микроскопа (камера культуры эмбиента и т.д.) должны быть подготовлены перед добавлением подвески Jurkat к камерной горке с присоединенными клетками Раджи. Следующие шаги описаны для коммерческого микроскопа(Таблица материалов). Тем не менее, любой перевернутый флуоресцентный микроскоп, оснащенный инкубатором клеточной культуры, может быть использован.- Используйте микроскоп с 60-кратным погружением в масляное, высокочисленной диафрагмой при визуализации поляризованного трафика.
- Убедитесь, что автоматическая система фокусировки включена и отрегулируйте смещение, чтобы сфокусировать ячейки Raji, привязанные к нижней поверхности. Пожалуйста, обратитесь к Рисунку 1, Видео 1 и Видео 2.
- После того, как Jurkat дополнение к каждому хорошо, содержащий присоединенные клетки Raji в шаге 5.3, быстро найти microwell камеры слайд на предварительно нагретый (1-2 h) микроскоп этап инкубатор (т.е., OKOlab) и выбрать некоторые позиции XY с микроскопом, поля, в которых он находится вероятно, чтобы зафиксировать формирующееся образование ИГ, сделанное, например, юркат-транс-трансативной ячейкой, попадающей в фокус микроскопа.
- Используйте предварительно нагретый инкубатор стадии микроскопа, так как было отмечено, что стабилизированная по температуре стадия поддерживает стабильные позиции X,Y,. Критериями для удобного поля XY являются: хорошо сфокусированные и несовместимые клетки Раджи (т.е. отображение зазоров между клетками) и наличие трансинфицированных клеток Jurkat (это можно проверить путем объединения каналов передачи и УФ или GFP). Клетки Jurkat будут осадок очень быстро (несколько минут) на камере слайд, и шансы на изображение возникающих синапсов будет уменьшаться со временем (Рисунок 1). Можно либо закончить эксперимент после определенного промежуток времени, либо перейти к Шагу 7 и исправить конъюги для последующего иммунофлуоресценции и анализов.
ПРИМЕЧАНИЕ: Можно выбрать до 16 различных полей микроскопа от до 4 различных микровелл для одновременного, многоскважины промежуток времени приобретения с надлежащим временным разрешением (1-2 мин на кадр). Ограничение зависит как от количества и интенсивности (влияющего на экспозицию камеры) различных флюорохромов, которые должны быть изображены (в зависимости от количества выраженных флуоресцентных белков, кроме CMAC). Одним из способов увеличения частоты кадров является запись для канала CMAC только в одном из каждого "n" временных рамок для GFP (т.е. n No 8, как показано на рисунке 2),так как клетки Raji придерживаются скважины дно и не легко перемещаться как клетки Jurkat. Кроме того, это благотворно для жизнеспособности клеток, так как частое воздействие УФ-излучения может повредить клетки. Попробуйте настроить частоту таймфрейма до 1 кадра каждые 1 минуту или меньше (т.е. 20 с на кадр в видео 1, Рисунок 2) так как поляризация MVB занимает несколько минут до нескольких часов для завершения. Микроскоп, оснащенный моторизованной башней эпифлуоресценции и соответствующими фильтрами флуоресценции или эквивалентами диапазона.
7. Формирование конечных точек синаптической конъюги и фиксации
- Если только эксперимент конечная точка запланирована (1-2 часа инкубации, чтобы синаптические конъюгированные формирования является целесообразным), инкубировать камерный слайд при 37 КС, 5% CO2 для 1-2 ч. Проверьте конъюгированное образование после периода культуры (как в рисунке 1) и, впоследствии, исправить конъюгты с ацетоном или PFA (фиксация будет зависеть от антигенов и антител, которые будут использованы). В этом случае инкубация не требует микроскопа стадии инкубатора. Пожалуйста, обратитесь к Видео 3 для примера.
- Чтобы исправить клетки, мыть колодец нежной тряской с теплой RPMI (37 градусов по Цельсию) среды без FCS (альбумин из сыворотки может осадок с ацетоном фиксации). Аспирировать и добавлять 200 КЛ PFA или предварительно охлажденный ацетон к каждому колодцу. Инкубировать камеру слайд при комнатной температуре (RT) или на льду, соответственно, в течение 20 минут.
ПРИМЕЧАНИЕ: Для фиксации ацетона, предварительно охладить ацетон при -20 градусов по Цельсию и предварительно охладить камеру слайд при 4 градусах Цельсия. Удалите пластиковые крышки, когда ацетон используется для фиксации клеток, культивированных в 8 хорошо стеклянных нижних слайдов камеры. - Вымойте каждый колодец дважды с ПОМОЩЬю PBS и добавьте 200 qL раствора для закалки (PBS, 50 mM NH4Cl). Инкубировать камерную горку при 4 градусах Цельсия.
ПРИМЕЧАНИЕ: На этом этапе камерная горка может оставаться в течение по крайней мере одного месяца при 4 градусах Цельсия, прежде чем выполнять протокол иммунофлуоресценции, как описано8. Крышка предотвращает испарение.
8. Обработка изображений
- Выполните после приобретения изображения деконволуции (т.е. Гюйгенс деконволюции или эквивалент, Таблица материалов) замедленного серии и / или еще фотографии фиксированных клеток. Deconvolute с использованием соответствующего программного обеспечения (т.е., используя "широкое поле" оптический вариант в Гюйгенс) и правильные оптические параметры. Deconvolution требует для обработки изображения измеренной функции распространения точки (PSF) микроскопа4.
- Кроме того, используйте программное обеспечение для расчета идеализированного PSF путем автоматической загрузки оптических параметров, включенных в метаданные из файлов микроскопа. Эти оптические параметры включают в себя флюорохромную длину волны, индекс преломления, численное отверстие цели и технику визуализации (конфокальный, широкоугольный и т.д.) 4.
- Впоследствии, программное обеспечение для визуализации использует PSF и различные алгоритмы деконволуации (т.е. qMLE и CMLE в программном обеспечении Гюйгенса) в пошаговом накопительном процессе, процесс расчета которого, по мере необходимости, может быть постоянно визуализирован и остановлен (или возобновлен) при необходимости пользователем. На этом этапе пользователь может изменить количество сверток и/или отношение сигнала к шуму и возобновить деконволюцию. Деконволюция программное обеспечение хорошо работает с промежуток времени серии (X, Y, T) (Видео 2) и стеки (X, Y, ) (Видео 3). Деконволюционные каналы были впоследствии объединены в CMAC, сырой канал, так как цитозольные, диффузные фторхромы не улучшаются путем деконволюции4,9.
Результаты
Мы следовали описанному протоколу, чтобы создать спряжение иммунного синапса Jurkat-Raji и правильно изобразить ранние стадии формирования ИГ. Наша цель состояла в том, чтобы улучшить ранние подходы20 ранее последовали для изучения поляризации MTOC и секреторного механизма по отношению к ИГ. Эти подходы были основаны на стратегии конечной точки, которая не позволяла сотнизить образование ИГ или ранние синаптические события, так как в этих стратегиях обязательное формирование ИГ происходило в гранулах смешанных, центрифугированных клеток, но не под микроскопом. Наш протокол был разработан, чтобы избежать этого основного предостережения, так как подход был основан на использовании соответствующей концентрации клеток (Шаги 2 и 3), в пользу формирования клетки к клетке IS спрягает на собственных слайдах камеры микроскопа (8 микро-хорошо камеры слайд), установленный на предварительно подогретой микроскоп этап инкубатора (в шаге 4). Эта стратегия индуцирует формирование ИГ, одновременно с замедленной захвата изображений(видео 1).
Рисунок 1 представляет собой синаптические конъюгаты Jurkat-Raji, полученные по протоколу (шаг 5.2). Изображение представляет собой первый кадр из репрезентативного эксперимента замедленного периода. 2 x 105 Раджи-клетки и 2 х 105 юркатных клеток были добавлены в 1 см2 колодца. Верхняя панель показывает канал передачи, средняя панель состоит из CMAC (синий) канал (клетки Раджи), а нижняя панель показывает как передачу плюс CMAC объединены каналы. В качестве эталона желтые стрелки маркируют некоторые синаптические конъюгаты, в то время как зеленые стрелки указывают на синаптические конъюгированные, сделанные одной клеткой Jurkat и несколькими клетками Раджи (сложные конъюги). Снижение концентрации клеток (105 или менее клеток в 1 см2 хорошо) будет обойти образование сложных клеточных конъюгированных, но не может быть достаточно клеточных конъюгирований для последующего анализа поляризованного трафика (см. ниже), что, в свою очередь, уменьшит также шансы найти и изображение возникающих синаптических конъюги.
Рисунок 2 представляет собой скриншоты, соответствующие параметрам изображения, используемым для одновременного захвата двух различных флюорохромов (CMAC и GFP-CD63) с помощью соответствующего программного обеспечения (т.е. NIKON NIS_AR) в эксперименте по промежуток времени, соответствующему видео 1.
Видео 1 (Иммунологические синапсы, сырые) представляет собой лимфоцит Jurkat T, выражающий GFP-CD63 (маркер многовезикулярных тел-MVB, зеленые пузырьки) и образование двойного синапса между 1 ячейкой Jurkat и 2 клетками Раджи, помеченными CMAC, синим цветом (один из них подвергается поглощению). Было зафиксировано одновременное движение MVB внутри ячейки Jurkat в сторону областей синапса (частота захвата кадров - 1 кадр каждые 20 секунд для GFP-CD63; скорость воспроизведения видео - 2 кадра в секунду). Синаптические конъюги были получены и изображены после описанного выше протокола (Шаг 6.2), который позволил изображение возникающих синапсов(видео 1). Одновременное захват каналов флуоресценции GFP-CD63 и CMAC осуществлялось с помощью соответствующего программного обеспечения (т.е. NIS-AR, Таблица материалов). Видео представляет собой необработанные данные из репрезентативного эксперимента замедленного заключения. Автоматическая система фокусировки была определена с соответствующим смещением по отношению к клеткам Раджи, привязанным к стекольу дну, чтобы обеспечить стабильный фокус вдоль эксперимента.
Видео 2 (иммунологические синапсы, после деконволюции) соответствует видео 1,но деконволюция изображений канала флуоресценции GFP-CD63 была выполнена с использованием соответствующего программного обеспечения деконволюции (т.е. Гюйгенов), используя оптический вариант «широкого поля» и надлежащие оптические параметры (шаг 8). Этот деконволютный канал был впоследствии объединен с CMAC, сырьеканалом. Налицо улучшение как соотношения сигнала к шуму, так и резкости изображения. Деконволюция была выполнена после приобретения, как описано выше. Для получения более подробной информации о деконволюции программного обеспечения, пожалуйста, обратитесь к4.
Видео 3 (иммунологический синапс, после фиксации и окрашивания иммунофлюоресценции) представляет собой q-stack (z-шаг размером 0,8 мкм, 5 кадров) фиксированного конъюгирования ИС после шага 6 протокола (фиксация ацетона). После фиксации, иммунофлуоресценция была выполнена в соответствии со стандартными протоколами25 с помощью Phalloidin для визуализации F-актина (зеленый), анти-CD63 для визуализации MVB (магента), анти--тубулин для визуализации MTOC (красный). CMAC (синий) маркирует ячейку Raji. Впоследствии конъюгация была изображена эпифлюоресценцией и несколько каналов deconvoluted с использованием "широкого поля" оптический вариант и надлежащие оптические параметры (шаг 7). Впоследствии каналы, связанные с дековолюцией, были объединены в CMAC, сырой канал, как указано в различных панелях (CMAC plus Transmittance-TRANS, CMAC plus Phallodin, CMAC plus anti-CD63 и CMAC plus anti-q-tubulin, соответственно). Белая стрелка маркирует синапс, в то время как зеленая стрелка маркирует MVB, а желтая стрелка этикетки MTOC. Подробная количественная оценка поляризации MVB и MTOC была объяснена в другом месте25.
Представленный здесь подход предполагает формирование конъюгов от клетки к клетке и, одновременно, замедленное приобретение широкоугольной флуоресценционной микроскопией (WFFM), за которым следует обработка изображений (после приобретения деконволюции). Эта тактика улучшила соотношение сигнала к шуму (SNR) изображений, улучшила их временное разрешение и позволила синхронизировать приобретение нескольких фторхромов в новых синапических конъюгированных4. Кроме того, протокол хорошо сочетается с последующими методами фиксации конечных клеток (параформальдегид, ацетон или метанол), что позволит дополнительно оформить иммунофлуоресцентное окрашивание и анализировать25 (видео 3). Этот протокол также совместим с лазерной сканирующей конфокальной микроскопией и другими самыми современными методами микроскопии.

Рисунок 1: Представитель двухразмерного микроскопического поля синаптических конъюгированных. Изображение представляет собой первый кадр из репрезентативного эксперимента замедленного выполнения в соответствии с протоколом. Верхняя панель показывает канал передачи, канал CMAC средней панели (клетки Raji) и нижнюю панель как слитые каналы. В качестве эталона желтые стрелки маркируют некоторые синаптические конъюги. Зеленые стрелки указывают на сложные синаптические конъюги (т.е. одна ячейка Джурката, устанавливающая синапсы с более чем одной ячейкой Раджи). Захваченные с 40x EWD (0.6 NA) цели. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 2: Настройки микроскопа по времени. Изображение соответствует нескольким скриншотам, соответствующим параметрам изображения, используемым для одновременного захвата двух различных флюорохромов (CMAC и GFP-CD63) с использованием соответствующего программного обеспечения (т.е. NIKON NIS_AR) в эксперименте по промежуток времени, соответствующему видео 1. Каждый кадр для канала GFP-CD63 был захвачен каждые 20 с. Только один кадр для УФ-канала из восьми кадров для канала GFP был захвачен для поддержания жизнеспособности клеток в ходе эксперимента. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Видео 1: Иммунологические синапсы, сделанные клеткой Jurkat, выражающей GFP-CD63, необработанные данные. Клетки Raji B, помеченные синим сливным трекером (CMAC, синий), были импульсивными с SEE в течение 30 минут, и были сформированы синапсы с клетками Jurkat, выражающими GFP-CD638. Были зафиксированы покадровые сроки, соответствующие каналам GFP-CD63 и CMAC (20 с на кадр; скорость воспроизведения видео - 2 кадра на с) и показан репрезентативный пример. Захваченные с 60x PLAN APO (1.4 NA) цели. Пожалуйста, нажмите здесь, чтобы посмотреть это видео (Право нажмите, чтобы скачать).

Видео 2: Иммунологические синапсы, сделанные клеткой Jurkat, выражающей GFP-CD63, после деконволюции. То же самое, что видео 1, но изображения были deconvoluted с помощью программного обеспечения deconvoltion. Налицо улучшение соотношения сигнала к шуму и повышенная резкость, обусловленная устранением загрязняющей из фокуса флуоресценции. Пожалуйста, нажмите здесь, чтобы посмотреть это видео (Право нажмите, чтобы скачать).

Видео 3: Фиксированный и иммуноокрашенный иммунологический синапс. На рисунке показана репрезентативная фиксированная синаптика после шага 7 Протокола и последующего иммунофлуоресценции. CMAC (синий) этикетки raji клетки, передатчик (TRANS), чтобы показать синаптический конъюгированный (белая стрелка), Phalloidin этикетки F-актин (зеленый), анти-CD63 этикетки MVB (magenta, зеленая стрелка) и анти--тбулина этикетки MTOC (красный, желтая стрелка), соответственно. Эти каналы были изображены эпифлюоресценции, deconvoluted и объединены, как указано. Видео включает в себя стек (z-шаг размером 0,8 мкм, 5 кадров). Белая стрелка маркирует синаптической область контакта, в то время как зеленая стрелка маркирует MVB, а желтая стрелка этикетки MTOC. Пожалуйста, нажмите здесь, чтобы посмотреть это видео (Право нажмите, чтобы скачать).
Обсуждение
Ограничение этого протокола заключается в том, что не все синапсы будут идеально ориентированы на оптическую ось. Используя этот метод, нет никакого способа предсказать и / или влиять на идеальную ориентацию для иммунной визуализации синапса. Чтобы решить эту проблему, мы исключаем из последующих анализов все случайно захваченные синапсы, которые, наконец, не соответствуют идеальным критериям. Эти синапсы, достаточно благоприятны, не очень часто. Тем не менее, можно обойти это ограничение, используя несколько экспериментальных подходов4.
Поляризованный релиз CD63 (дегрануляция) может быть количественно с помощью других дополнительных методов, таких как окрашивание поверхности клеток CD63 (CD63 для переселения на поверхность клетки) в живых клетках (не фиксированных и не пронизанных), после шага 6, и последующей стирки и фиксации, как ранее показано8. Кроме того, CD63 релиз на экзосомы8,9 и экзосомы количественнонатжение наночастиц отслеживания анализов9,25 может быть выполнена. Эти подходы, безусловно, совместимы с нашим протоколом, при условии, что фиксация выполняется после иммунофлуоресценции поверхности клетки живых клеток.
Мы обнаружили, что идеальное количество клеток (для 1 см2 хорошо в 8-ну хорошо камеры слайд) составляет 4 х 105 клеток (2 х 105 Raji клеток и 2 х 105 Jurkat клетки), так как они эффективно придерживаться нижней части колодца. Связывание эффективности к пластику (с использованием фибронектина) или стеклянного дна (с использованием поли-L-лизин) микрооползней, как правило, не является проблемой. Более высокие числа клеток могут не производить разрывов между присоединенными клетками и, впоследствии, сложными синаптические конъюгированными(рисунок 1); обе ситуации нежелательны, когда одноклеточные конъюгаты должны быть изображены, например, в экспериментах по поляризации MTOC или секреторных гранул. Меньшее количество клеток может уменьшить шансы найти конъюги, особенно когда трансинфицированные клетки Jurkat оспариваются СБ для генерации синапсов. Пожалуйста, обратите внимание, что мы заметили, что температурно-стабилизированный инкубатор стадии на месте на микроскопx X/ Y этапе до начала протокола (т.е., 1-2 ч заранее, чтобы стабилизировать этап / микроскоп установки) поддерживает стабильные X, Y, - параметры, что имеет решающее значение для надлежащего изображения. Автоматическая система фокусировки в конечном итоге компенсирует небольшие вариации.
Когда некоторые методы трансдукции генов, такие как электропорация используются для экспресса флуоресцентных химерных белков (т.е. GFP-CD63) в клонах Jurkat (Видео 1), значительная часть клеток может умереть после электропорации. Это может быть проблемой, поскольку, хотя мертвые клетки не образуют синапсы, когда они находятся в избытке над живыми, транс-инфицированных клеток, они могут вмешиваться в формирование конъюгированных, сделанных живыми Т клеток. Мы обнаружили, что тщательное устранение мертвых клеток из трансзараженных культур с помощью среды градиента плотности следующие стандартные протоколы до шага конъюгированного формирования действительно может увеличить шансы правильно спряжения изображения. Кроме того, низкая эффективность трансфекции (0,20%) может быть важным предостережением, так как это, в сочетании с умеренной конъюгированной эффективности формирования (около 60%)25, уменьшит вероятность найти конъюги, сделанные трансинфицированных клеток. Это не является проблемой, когда нетранспередаваемые клетки используются для получения конъюги в конечных экспериментах и последующей фиксации. 8 камерных слайдов микроколодца совместимы с обычными протоколами иммунофлуоресценции. Это повышает гибкость вышеуказанного протокола с различными целями. Фиксация с ацетоном может быть проблемой для рассмотрения при использовании камерных слайдов с пластиковыми дном колодцев. Тем не менее, есть коммерчески доступные 8 микроколодца микроскоп камеры слайды, содержащие стеклянные днища, которые совместимы с ацетоном фиксации. Удалите пластиковые крышки, когда ацетон используется для фиксации клеток, культивированных в 8 microwell стеклянных нижних слайдов камеры.
Рекомендуется, чтобы микроскоп оснащался моторизованной стадией XY, моторизованной репрелепрелитируемой репрелесолисткой и автоматической системой фокусировки (например, Perfect Focus System) или эквивалентными добавками. Когда мульти-хорошо приобретение требуется25, автоматическая система фокусировки обеспечит стабильный фокус на всем протяжении эксперимента. Предыдущий опыт показывает, что, установив соответствующий фокус компенсируется на клетки Раджи, как движение Т-клеток(видео 1) и микроскоп этап / камеры слайд движений в XY многоточечных экспериментов, могут быть компенсированы. Это действительно удобно для многократного захвата замедленного времени.
Черно-белая, панхроматическая и охлажденная заряженная соединенная камера (CDD) была использована, но более высокой чувствительности, флуоресценции научной дополнительной металлооксидной полупроводниковой (sCMOS) камера желательна, так как это уменьшит время экспозиции камеры и повысит временное разрешение. Короткое время экспозиции камеры мы использовали (рейтинг форме 100 мс до 500 мс) в сочетании с автоматической флуоресценции затвор позволяет длительный промежуток времени захвата (до 24 ч) с адекватным разрешением времени (1 кадр в минуту или меньше, до 16 Позиций XY) без значительного флуоресценции отбеливания и / или потери в жизнеспособности клеток. Моторизованный этап позволяет многоточечный (XY) захвата и увеличивает шанс найти и изображение возникающих и развивающихся синапсов в идеальной ориентации, но и позволяет приобретение изображения в многофункциональных камерных слайдов, когда различные клоны Jurkat должны быть одновременно сопряжены25. Высокая численная диафрагма цели (т.е. 60x, 1.4) необходима для того, чтобы получить наилучшие результаты при анализе трафика секреторных гранул.
RAJI-SEE-Jurkat представляет собой устоявшуюся иммунологическую синапсовую модель, которая была использована множеством исследователей, так как она была первоначально описана11. Мы адаптировали наш протокол к этой модели, чтобы правильно изобразить ранние этапы формирования ИГ. Наша цель состояла в том, чтобы улучшить ранние подходы20 ранее последовали для изучения поляризации MTOC и секреторного механизма по отношению к ИГ. Примечательно, что конъюги, сделанные с этим протоколом производят F-актин реорганизации на синапсе, настройка канонического SMAC, сопутствуя MVB поляризованного трафика25. Эти важные события были также проанализированы и проверены конфокальной микроскопии25.
Кинетические различия в поляризованном трафике существуют между различными типами ИС. Например, поляризованная транспортировка литические гранулы из CTLs происходит в секундах или очень мало минут, в то время как несколько цитокиносодержащих пузырьков из Th лимфоцитов занять от нескольких минут до нескольких часов до конца. Эти временные различия должны быть приняты во внимание заранее, для того, чтобы разработать наилучшую стратегию и выбрать наиболее подходящий экспериментальный и визуальный подход, так как для некоторых визуальных утех (т.е. лазерное сканирование конфокальной микроскопии (LSCM)), время может быть ограничивающим фактором, так как время захвата гораздо выше, чем соответствующее разрешение времени (1 минута или меньше)4. Это не является ограничением, когда широкополевой флуоресценции микроскопии (WFFM) используется, как описано в протоколе выше. Так как в CTLs, поляризация MTOC к синапсу длится всего несколько минут3,6,17, различные конкретные состояния современной микроскопии подходы отличаются от описанных здесь (но укрывательство более высокое пространственное и временное разрешение) необходимы для того, чтобы правильно изображение этих синапсов26,27, в основном, когда несколько микроскопов (многоточечные захват) изображения. Эти высокие разрешения, новые подходы могут быть также использованы для визуализации синапсов, сделанных Th лимфоцитов, хотя экономичные и / или логистических причин (т.е., основное оборудование, необходимое для некоторых из этих методов визуализации стоит 6-7 раз больше, чем описано здесь), безусловно, может представлять собой ограничение для этих современных методов визуализации4. Тот факт, что IS сделаны Th лимфоцитов являются долгоживущими, и то обстоятельство, что в Th лимфоцитов MTOC, лимфокин-содержащих секреторных пузырьков и MVB занять от нескольких минут до часов для транспортировки и док-станции IS22, делает этот протокол идеальным, доступным подходом для визуализации TH-APC IS.
WFFM в сочетании с деконволюцией изображений после приобретения представляет собой интересный подход, и не только экономические причины поддерживают эту стратегию. Внутреннее плохое разрешение в оси q (наиболее важное предостережение техники) может быть улучшено с помощью пост-приобретения изображения deconvoltion4 (сравните видео 1 с видео 2). Deconvolution использует расчетный подход к обработке изображений, который может улучшить соотношение сигнала к шуму и разрешение изображения и контрастироватьот 27 до 2 раз, вплоть до 150-100 нм в оси XY и 500 нм в оси4.
Использование высокой чувствительности, высокой скорости считывания и широкого динамического диапазона, новых флуоресценции sCMOS камеры улучшат качество изображения и уменьшит отбеливание флуоресценции. Гибкость, предлагаемая описанным здесь протоколом спряжения от клеток к клетке, позволяет сочетать описанный клеточный подход с несколькими методами современной микроскопии, как в живых клетках, так и в стационарных клетках, и ожидаемый результат действительно улучшит наши знания иммунологического синапса.
Хотя мы внедрили и проверили протокол с помощью простых в обращении, хорошо установленных клеточных линий, потенциал подход может позволить визуализации более физиологических взаимодействий, когда первичные Т-клетки и различные типы антиген-представления клеток (таких как дендритные клетки макрофагов) используются5. В этом контексте, этот протокол был также расширен и проверен с помощью суперантигена (SEB) импульсной мыши EL-4 клеточной линии, используемой в качестве APC, чтобы бросить вызов первичной мыши T лимфобластов9. Действительно, первичные T лимфоциты, CTLs, в частности, оказали более недолговечные и динамические синаптические контакты (см. Дополнительное видео 8 в ссылке9) для тех, кто видел с моделью SEE-Raji и Jurkat. Вариабельность синапических контактных режимов лучше всего просмотреть для первичных взаимодействий Т-клеток с дендритными клетками или В-клетками в двухмерных эквивалентах ткани in vitro, которые также могут быть записаны и проанализированы с помощью этого протокола. Кроме того, помимо суперантигенов, этот метод можно использовать для изображения других типов синапсов. Например, он может быть использован в TCR трансгенных, антиген-специфических Т-клеток модели, например, с использованием овальбумин конкретных морин OT1/OT2 системы или трансфекции Т-клеток с антиген-специфических Т-клеточных рецепторов. Это открывает множество экспериментальных возможностей в ближайшем будущем.
Раскрытие информации
Авторы не заявляют о конфликте интересов.
Благодарности
Мы признаем все прошлые и нынешние члены лаборатории за их щедрый вклад. Эта работа была поддержана грантами от испанского министра и Компетивидада (MINECO), Plan Nacional de Investigacion Cientefica (SAF2016-77561-R to M.I., которая была частично предоставлена за счет финансирования FEDER-EC). Мы признаем, факультет медицины (UAM) и Departamento де аудиовизуальных факультетов Медицины за их поддержку и средства, предоставляемые для производства видео. Мы признаем NIKON-Europe за непрерывную и превосходную техническую и теоретическую поддержку. Бесплатный доступ к этой статье спонсируется Nikon.
Материалы
| Name | Company | Catalog Number | Comments |
| Camera Nikon DS-QI1MC | Nikon | MQA11550 | Cooled Camera Head |
| CMAC | ThermoFisher Scientific | C2110 | Cell tracker blue |
| JURKAT cells | ATCC | ATCC TIB-152 | Effector T lymphocytes |
| μ-Slide 8 well ibiTreat, μ-Slide 8 well Glass-Bottom | IBIDI | Cat.No: 80826, 80827 | Cell culture and cell imaging supports |
| Microscope NIKON Eclipse Ti-E | Nikon | NIKON Eclipse Ti-E | Wide-field fluorescence, fully-motorized microscope equipped with Perfect Focus System (PFS) option |
| Microscope Stage Incubator with 3-channel manual gas mixer and gas bubbler/ humidity module | OKOLAB | H201-NIKON-TI-S-ER | Cell culture atmosphere |
| Raji Cells | ATCC | ATCC CCL-86 | APC |
| RPMI medium GIBCO | ThermoFisher Scientific | 21875034 | Culture medium |
| Streptococcus Enterotoxin E (SEE) | Toxin Technology, Inc | EP404 | Bacterial Toxin |
| Software Huygens Essential | SVI | Huygens Essential | Image Deconvolution software |
| Software ImageJ | NIH | Image J | Image software |
| Software Nikon NIS-AR | Nikon | NIS-Elements AR | Image capture and analysis software |
Ссылки
- Fooksman, D. R., et al. Functional anatomy of T cell activation and synapse formation. Annual Review of Immunology. 28, 79-105 (2010).
- de la Roche, M., Asano, Y., Griffiths, G. M. Origins of the cytolytic synapse. Nature Reviews. Immunology. 16, 421-432 (2016).
- Griffiths, G. M., Tsun, A., Stinchcombe, J. C. The immunological synapse: a focal point for endocytosis and exocytosis. The Journal of Cell Biology. 189, 399-406 (2010).
- Calvo, V., Izquierdo, M. Imaging Polarized Secretory Traffic at the Immune Synapse in Living T Lymphocytes. Frontiers in Immunology. 9, 684 (2018).
- Friedl, P., den Boer, A. T., Gunzer, M. Tuning immune responses: diversity and adaptation of the immunological synapse. Nature Reviews. Immunology. 5, 532-545 (2005).
- Xie, J., Tato, C. M., Davis, M. M. How the immune system talks to itself: the varied role of synapses. Immunological Reviews. 251, 65-79 (2013).
- Colombo, M., Raposo, G., Théry, C. Biogenesis, Secretion, and Intercellular Interactions of Exosomes and Other Extracellular Vesicles. Annual Review of Cell and Developmental Biology. 30, 255-289 (2014).
- Alonso, R., et al. Diacylglycerol kinase alpha regulates the formation and polarisation of mature multivesicular bodies involved in the secretion of Fas ligand-containing exosomes in T lymphocytes. Cell Death & Differentiation. 18, 1161-1173 (2011).
- Mazzeo, C., Calvo, V., Alonso, R., Merida, I., Izquierdo, M. Protein kinase D1/2 is involved in the maturation of multivesicular bodies and secretion of exosomes in T and B lymphocytes. Cell Death & Differentiation. 23, 99-109 (2016).
- Mittelbrunn, M., et al. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nature Communications. 2, 282 (2011).
- Montoya, M. C., et al. Role of ICAM-3 in the initial interaction of T lymphocytes and APCs. Nature Immunology. 3, 159-168 (2002).
- Abraham, R. T., Weiss, A. Jurkat T cells and development of the T-cell receptor signalling paradigm. Nature Reviews. Immunology. 4, 301-308 (2004).
- Huse, M., Quann, E. J., Davis, M. M. Shouts, whispers and the kiss of death: directional secretion in T cells. Nature Immunology. 9, 1105-1111 (2008).
- Peters, P. J., et al. Cytotoxic T lymphocyte granules are secretory lysosomes, containing both perforin and granzymes. The Journal of Experimental Medicine. 173, 1099-1109 (1991).
- Vignaux, F., et al. TCR/CD3 coupling to Fas-based cytotoxicity. The Journal of Experimental Medicine. 181, 781-786 (1995).
- de Saint Basile, G., Menasche, G., Fischer, A. Molecular mechanisms of biogenesis and exocytosis of cytotoxic granules. Nature Reviews. Immunology. 10, 568-579 (2010).
- Huse, M. Microtubule-organizing center polarity and the immunological synapse: protein kinase C and beyond. Frontiers in Immunology. 3, 235 (2012).
- Yi, J., et al. Centrosome repositioning in T cells is biphasic and driven by microtubule end-on capture-shrinkage. The Journal of Cell Biology. 202, 779-792 (2013).
- Jang, J. H., et al. Imaging of Cell-Cell Communication in a Vertical Orientation Reveals High-Resolution Structure of Immunological Synapse and Novel PD-1 Dynamics. Journal of Immunology. 195, 1320-1330 (2015).
- Kupfer, A., Singer, S. J. Cell biology of cytotoxic and helper T cell functions: immunofluorescence microscopic studies of single cells and cell couples. Annual Review of Immunology. 7, 309-337 (1989).
- Dustin, M. L., Depoil, D. New insights into the T cell synapse from single molecule techniques. Nature Reviews. Immunology. 11, 672-684 (2011).
- Dustin, M. L. Supported bilayers at the vanguard of immune cell activation studies. Journal of Structural Biology. 168, 152-160 (2009).
- Jambrina, E., et al. Calcium influx through receptor-operated channel induces mitochondria-triggered paraptotic cell death. The Journal of Biological Chemistry. 278, 14134-14145 (2003).
- Fuss, I. J., Kanof, M. E., Smith, P. D., Zola, H. Isolation of whole mononuclear cells from peripheral blood and cord blood. Current Protocols in Immunology. , 1 (2009).
- Herranz, G., et al. Protein Kinase C delta Regulates the Depletion of Actin at the Immunological Synapse Required for Polarized Exosome Secretion by T Cells. Frontiers in Immunology. 10, 851 (2019).
- Ritter, A. T., et al. Actin depletion initiates events leading to granule secretion at the immunological synapse. Immunity. 42, 864-876 (2015).
- Combs, C. A., Shroff, H. Fluorescence Microscopy: A Concise Guide to Current Imaging Methods. Current Protocols in Neuroscience. 79, (2017).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены