
Method Article
Субклеточное фракционирование для выделения синаптических компонентов из мышиного мозга
В этой статье
Резюме
Этот протокол представляет собой надежный, подробный метод получения высокочистых синаптосом, синаптических везикул и других синаптических фракций из мозга мыши. Этот метод позволяет оценить синаптические процессы, включая биохимический анализ локализации и функции белка с компартментным разрешением.
Аннотация
Синаптические терминалы являются первичными сайтами нейронной коммуникации. Синаптическая дисфункция является отличительной чертой многих нервно-психических и неврологических расстройств. Таким образом, характеристика синаптических субкомпонентов биохимической изоляцией является мощным методом выяснения молекулярных основ синаптических процессов, как в здоровье, так и в болезни. Этот протокол описывает выделение синаптических терминалов и синаптических субкомпонентов от мозга мыши путем субклеточного фракционирования. Во-первых, герметичные синаптические терминальные структуры, известные как синаптосомы, выделяются после гомогенизации мозговой ткани. Синаптосомы представляют собой нейрональные пре- и постсинаптические компартменты с защемленными и герметичными мембранами. Эти структуры сохраняют метаболически активное состояние и ценны для изучения синаптической структуры и функции. Затем синаптосомы подвергают гипотоническому лизису и ультрацентрифугированию для получения синаптических субкомпонентов, обогащенных синаптическими везикулами, синаптическим цитозолом и синаптической плазматической мембраной. Чистота фракции подтверждается электронной микроскопией и анализом биохимического обогащения белков, специфичных для субсинаптических компартментов. Представленный метод является простым и ценным инструментом для изучения структурно-функциональных характеристик синапса и молекулярной этиологии различных заболеваний головного мозга.
Введение
Синапсы являются основными вычислительными единицами мозга, через которые нейроны общаются и выполняют разнообразные и изысканно сложные функции. Синапсы, таким образом, имеют основополагающее значение для здоровья мозга1; синаптическая дисфункция подразумевается как источник или результат многих расстройств2. Синапсы состоят из пре- и постсинаптических терминалов, расширений двух разных нейронов, которые тесно аппонированы и разделены синаптической щелью, пересекаемой молекулами синаптической адгезии. Информационные потоки из пре- и постсинаптического компартмента в виде химических мессенджеров, называемых нейротрансмиттерами1. Молекулярные процессы, участвующие в нейротрансмиссии, являются активными направлениями исследований 3,4,5. Понимание патогенных процессов в синаптических терминалях и реакции синапсов на патологию в других нейронных подкомпонах являются важнейшими шагами к решению проблем головного мозга 1,2. Несколько методологических достижений, преимущественно применяемых к мышиным моделям, продвинули это стремление6. Выделение синаптических фракций дифференциальным центрифугированием является одним из таких методов смены парадигмы, который позволил детально оценить синаптические процессы в здоровье и болезни.
Мозг взрослого человека состоит из 80-90 миллиардов нейронов 7,8. Среди видов мышей мозг крысы содержит примерно ~ 200 миллионов нейронов, в то время как мыши имеют ~ 70 миллионов 9,10. Каждый нейрон образует тысячи специфических синаптических связей с сетью высокополяризованных нейронов, смешанных с глиальными клетками и плотной сосудистой системой. В такой сложной и гетерогенной ткани когда-то было немыслимо выделять и изучать синапсы как самостоятельную систему. В 1960-х годах Виктор Уиттакер, Кэтрин Хебб и другие сделали это возможным, изолировав интактные синаптические терминалы с использованием субклеточного фракционирования 11,12,13,14. В попытке выделить синаптические везикулы (SV) они гомогенизировали мозг с помощью жидкой силы сдвига в изоосмотической (0,32 М) сахарозе с последующей ультрацентрифугированием. Они получили защемленные, закрытые плазматической мембраной, неповрежденные нервные окончания или варикозное расширение вен, которые они назвали частицами нервных окончаний (NEPs)11,13. Поскольку структурные и функциональные характеристики синапса были сохранены в этих структурах, НЭП позже были названы «синаптосомами» для конгруэнтности с другими субклеточными органеллами13,15. Стоит отметить, что работы Эдуардо де Робертиса и его коллег, придумавших термин «синаптический везикул», пересекались с работами Уиттакера и его коллег и способствовали валидации выделения и характеристики «синаптосомы» 16,17,18.
Синаптосомы представляют собой физиологически активные структуры, которые содержат все клеточные и молекулярные свойства, необходимые для хранения, высвобождения и обратного захвата нейротрансмиттеров13,18. Сохранение ключевых синаптических характеристик in vitro и свобода от несинаптических компонентов также способствуют полезности этого метода изоляции. Синаптосомы внесли огромный вклад в понимание химических и физиологических свойств нейротрансмиссии и в настоящее время используются для изучения синаптических молекулярных процессов и их изменений при заболеваниях 19,20,21,22,23. Синаптосомы также являются исходным исходным материалом для выделения синаптических компонентов, таких как SV, везикулы с клатриновым покрытием (CCV), синаптический цитозоль, синаптическая плазматическая мембрана, синаптические митохондрии, молекулы синаптической адгезии и другие компоненты, представляющие интерес, которые могут облегчить понимание молекулярных механизмов синаптической функции 18,19,20,24,25,26, 27,28. Эти субсинаптические компоненты могут быть получены осмотическим лизисом синаптосом и ультрацентрифугированием ультрацентрифугирования градиента плотности сахарозы15,29. Хотя оригинальный метод субклеточной фракционирования исследовательской группы Уиттакера, как известно, эффективен в выделении качественных синаптосом и SV13,30, недавние оптимизации повышают чистоту субклеточных фракций 22,23,31,32. В этой статье представлена очень подробная и доступная версия классического протокола субклеточного фракционирования мышиной ткани мозга для выделения синаптосом, SV и других субсинаптических компонентов.
протокол
Все эксперименты с мышами были одобрены Институциональным комитетом по уходу за животными и их использованию (IACUC) в Йельском университете (Протокол 2021-11117) и выполнены в учреждении, аккредитованном Ассоциацией по оценке и аккредитации Laboratory Animal Care International (AAALAC). Уход за животными и их содержание соответствовали Руководству по уходу за лабораторными животными и их использованию33 и были предоставлены Йельским центром ресурсов для животных (YARC). Животные содержались в 12-часовом светло-темном цикле с доступом ad libitum к пище и воде. Пять-восемь мышей или от двух до четырех крыс на генотип или состояние требуются для следующего протокола. Меньше крыс необходимо из-за их больших объемов мозга. Аналогичным образом, возраст подопытных животных может влиять на выход фракции; дополнительные мыши могут потребоваться в возрасте до 2 месяцев. В остальном изложенные процедуры распространяются как на мышиные виды, так и на здоровых взрослых животных любого возраста. Репрезентативные данные, представленные в этом исследовании, использовали мышей дикого типа (C57BL/6J) (возраст = 2 месяца; четыре самца и четыре самки на реплику), полученных из коммерческого источника (см. Таблицу материалов).
1. Экспериментальная подготовка
ПРИМЕЧАНИЕ: Этот протокол требует ~ 11 часов для завершения работы одного исследователя. Настоятельно рекомендуется завершить настольную установку (рисунок 1), подготовку буфера (таблица 1), предварительное охлаждение центрифуг и роторов до 4 °C, а также сбор и маркировку необходимых материалов и оборудования (см. Таблицу материалов) за день до выполнения протокола, где это применимо.

Рисунок 1: Настольная установка. До вскрытия мозга (А) гомогенизаторы стекла Dounce и (Б) все буферы охлаждались на льду. (C) Растворы ингибиторов протеазы размораживали на льду. Получен второй контейнер с влажным льдом для центрифужных трубок, Дьюар жидкого азота (не показан) и (D) контейнер сухого льда для кратковременного хранения образцов, мгновенно замороженных в жидком азоте. (E) Пробирки микроцентрифуги были предварительно маркированы для всех образцов, так как во время этой процедуры были собраны четыре аликвоты каждого образца субклеточной фракции на генотип или состояние (экономящий время совет: тщательно маркировать все пробирки за день до проведения эксперимента). (F) Соответствующий контейнер для отходов биологической опасности, (G) 70% этанола, (H) хирургические инструменты и (I) абсорбирующая поверхностная прокладка. Необходимые центрифужные трубки и одноразовые материалы были отведены для эффективного доступа во время реализации протокола (не показаны). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
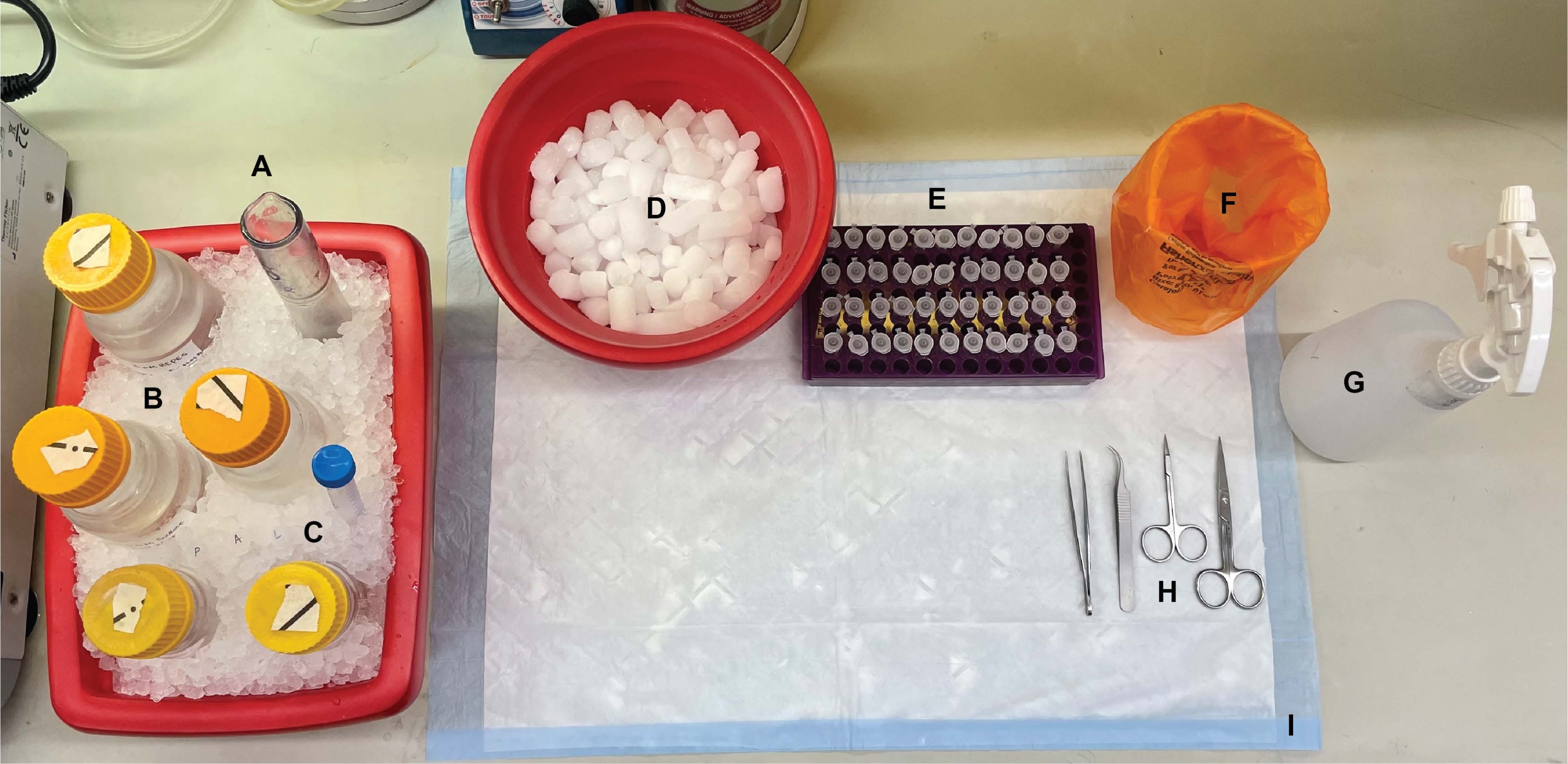{kind=link}
- Подготовьте столешницу к операции и соберите ножницы и щипцы, необходимые для иссечения мозга (см. Таблицу материалов). Предварительно маркированные микроцентрифужные трубки объемом 1,5 мл для биопсии хвоста мыши и четыре трубки на собранную фракцию, как показано на рисунке 2.
- Получите два контейнера с влажным льдом, один контейнер сухого льда и настольную колбу с жидким азотом Дьюара.
- Размораживание фенилметилсульфонилфторида (PMSF), пепстатина А, апротинина и растворов лейпептина на льду (см. Таблицу материалов). Подготовьте необходимые буферы (таблица 1).
ПРИМЕЧАНИЕ: Растворы сахарозы могут быть приготовлены заранее и сохранены при температуре 4 °C. Однако ингибиторы протеазы (размороженные запасы и таблетки) необходимо добавлять свежими во все буферы в начале эксперимента из-за нестабильности этих реагентов в водных растворах. Кроме того, все буферы должны быть подготовлены стеклянной посудой без моющих средств и водой без моющих средств, чтобы обеспечить сбор неповрежденных синаптосом. - Охладите все буферы и стеклянные гомогенизаторы Dounce (см. Таблицу материалов) на льду. Установите центрифуги на 4 °C и охладите роторы до 4 °C.
- Добавьте 14 мл буфера А (таблица 1) к гомогенизатору Доунса на льду.
Таблица 1: Состав субклеточных фракционных буферов. Пожалуйста, нажмите здесь, чтобы загрузить эту таблицу.

Рисунок 2: Обзор протокола субклеточного фракционирования. Сводная схема стадий субклеточного фракционирования и собранных образцов. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
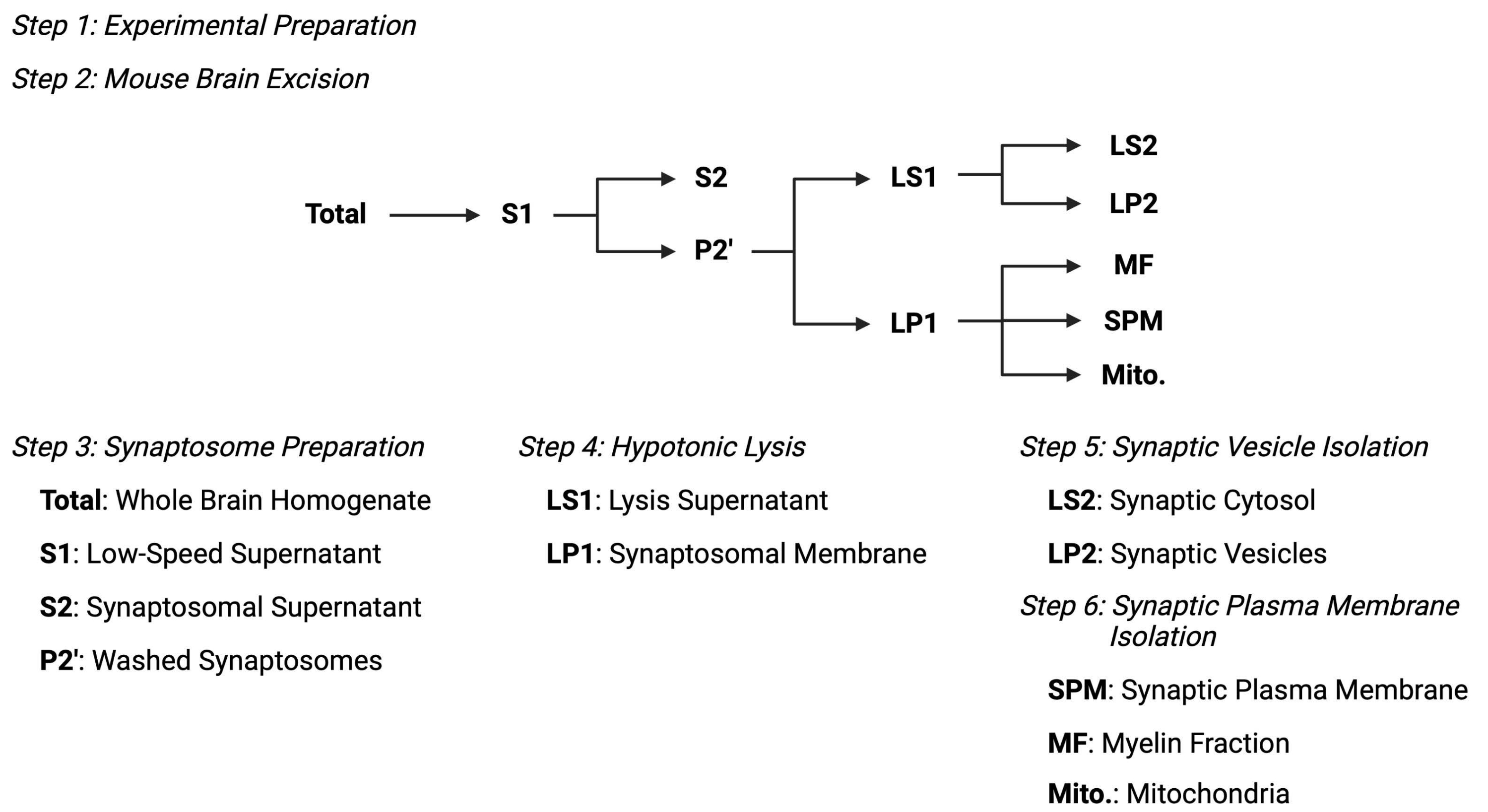{kind=link}
2. Иссечение мозга мыши

Рисунок 3: Черепно-лицевая анатомия. (А) Дорсальный вид черепа мыши с соответствующими черепно-мозговыми структурами. (B) Левый боковой вид черепа и мозга мыши с указанием соответствующих черепных структур и анатомических направлений. Пунктирные линии обозначают места, где должны быть сделаны разрезы. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
- Глубоко обезболивайте каждую мышь 100% изофтораном в анестезиологической камере, расположенной в вытяжном шкафу или шкафу биобезопасности, используя открытый капельный метод34. Жертвуйте каждой мышью вывихом шейного отдела позвоночника с последующим быстрым обезглавливанием. Чередуйте генотипы или экспериментальные группы для каждого жертвоприношения и вскрытия12. Получить биопсию хвоста после эвтаназии путем иссечения тонкими ножницами 2 мм дистального кончика хвоста. Храните ткань для генотипирования.
- Опрыскивайте обезглавленную голову 70% этанолом, чтобы предотвратить прилипание волос к тканям и хирургическим инструментам во время рассечения.
- Вставьте тонкие ножницы под кожу при обезглавливающем разрезе на перикраниальную глубину и сделайте средний сагиттальный разрез до межнационального шва (рисунок 3А), чтобы втянуть кожу головы из черепа.
- Работая от затылочной области к каждому височному аспекту, обрежьте фасцию и мышцу, чтобы обнажить внешнюю поверхность черепа за пределами каждого внешнего акустического мяса (рисунок 3B).
- Закрепите кожу головы и ростральный аспект черепа недоминирующей рукой. С другой вставьте тонкие ножницы 2 мм в каудальную сторону большого отверстия, где спинной мозг виден выходящим. Сделайте разрез средней линии до тех пор, пока ножницы не достигнут внутренней поверхности внутритеменной кости (рисунок 3; пунктирные линии).
ПРИМЕЧАНИЕ: Во время первоначального разреза ножницы должны быть параллельны спинному мозгу с давлением, приложенным к внутренней поверхности черепа, чтобы предотвратить повреждение ствола мозга и мозжечка. - Измените угол наклона ножниц так, чтобы лезвия шли параллельно спинной поверхности черепа. Продолжайте продвижение среднего сагиттального разреза рострально через теменную и лобную кости, используя сагиттальные и межфронтальные швы в качестве ориентира. Используйте постоянное восходящее давление, чтобы избежать повреждения коры головного мозга. Прекратите разрез сразу за межназальным швом (рисунок 3А).
- Сделайте небольшой перпендикулярный разрез (~3 мм) к носовой кости, ростральный к межназальному шву, положив ножницы перпендикулярно черепу с каждым лезвием, расположенным на носово-предчелюстном шве и сделав один ровный разрез (рисунок 3; пунктирные линии).
ПРИМЕЧАНИЕ: Этот шаг увеличит легкость втягивания черепа и будет иметь решающее значение для сбора обонятельной луковицы, если эта область представляет интерес. - Укрепляя ростральный аспект, используйте одну сторону пары текстурированных щипцов, чтобы осторожно поднять череп вверх от мозга, а затем латерально и вентрально. Повторяйте вдоль средней линии по мере необходимости, затем для другого полушария, пока вся поверхность мозга не будет обнажена.
- Используя изогнутые щипцы или тонкий шпатель, осторожно приподнимите ростральную сторону мозга. Перережьте зрительный и черепно-мозговой нервы, чтобы завершить иссечение из черепа.
- Для каждого состояния соберите от пяти до восьми мозгов мыши вместе в охлажденный стеклянный гомогенизатор Dounce, содержащий 14 мл буфера А (таблица 1).
3. Препарат синаптосомы
ПРИМЕЧАНИЕ: Схемы этой процедуры показаны на рисунке 4.

Рисунок 4: Препарат синаптосомы. Схема шага 3, генерация синаптосом (P2'). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
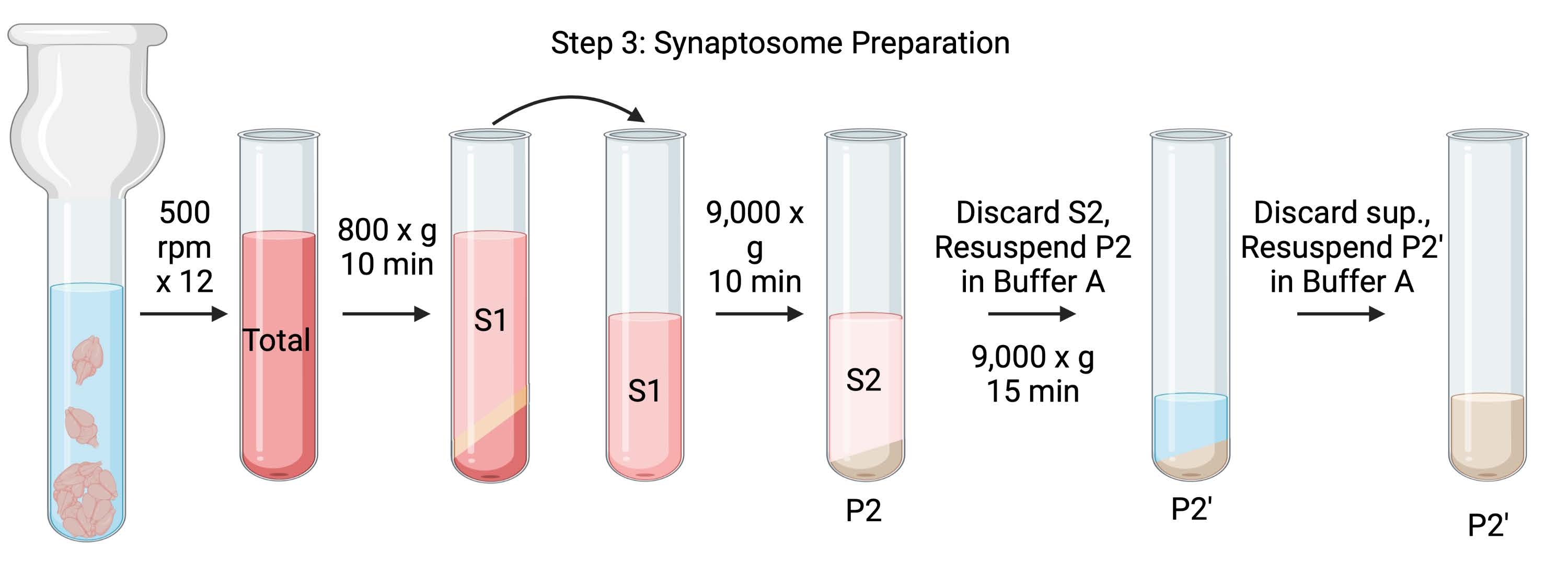{kind=link}
- Гомогенизируйте мозг с помощью стеклянного гомогенизатора Dounce в 12 проходах вверх-вниз при 500 оборотах в минуту (всего). Делайте небольшую паузу при каждом нисходящем ударе, чтобы обеспечить тщательную гомогенизацию ткани. Гомогенизируйте предпочтительно в ледяной ванне, чтобы избежать согревания и денатурации белка. Возьмите 5 мкл аликвот для определения концентрации белка методом анализа бицинхониновой кислоты (BCA, см. Таблицу материалов). Возьмите 100 мкл аликвот лизата всего мозга для вестерн-блотта (WB). Для этого и всех последующих образцов (рисунок 2) возьмем две аликвоты для BCA и две аликвоты для WB. Все собранные аликвоты заморозить во флеш-аликвоте и хранить при −80 °С.
- Раскрутите общий гомогенат мозга в высокоскоростной центрифуге с круглым дном (14 мл) (см. Таблицу материалов) при 800 х г в течение 10 мин при 4 °C для получения супернатанта (S1). Переместите S1 в новую центрифужную трубку, оставив гранулу (P1), которая содержит неповрежденные клетки и ядра. Избегайте пипетки пушистых, белых, рыхлых, поверхностных гранул. Возьмите 2 x 5 мкл S1 для BCA и 2 x 100 мкл S1 для WB.
- Отжимать S1 при 9000 х г в течение 15 мин при 4 °C с получением синаптосомального супернатанта (S2) и сырой гранулы синаптосомы (P2). Возьмите 2 x 10 мкл S2 для BCA и 2 x 500 мкл S2 для WB. Выбросьте супернатант после получения аликвот и переходите к следующему шагу с гранулой.
- Повторное суспендирование Р2 в 3 мл ледяного буфера А с ингибиторами протеазы и центрифугой при 9000 х г в течение 15 мин при 4 °С с получением супернатанта (S2') и промытых синаптосом (Р2'). Выбросьте супернатант и сохраните гранулу.
- Повторное суспендирование P2' в 3 мл буфера A. Избегайте повторного использования темно-красной части на дне гранулы, которая в основном содержит митохондрии. Возьмите 2 x 20 мкл P2' для BCA и 2 x 100 мкл P2' для WB.
ПРИМЕЧАНИЕ: Это может быть достигнуто путем осторожного смешивания краев и поверхности гранулы для повторного суспендирования белых синаптосом, направляя кончик пипетки от красного центра гранулы.
4. Гипотонический лизис
ПРИМЕЧАНИЕ: Схемы этой процедуры показаны на рисунке 5.

Рисунок 5: Гипотонический лизис. Схема шага 4, гипотонический лизис синаптосом для генерации супернатанта лизиса (LS1) и фракций синаптосомной мембраны (LP1). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
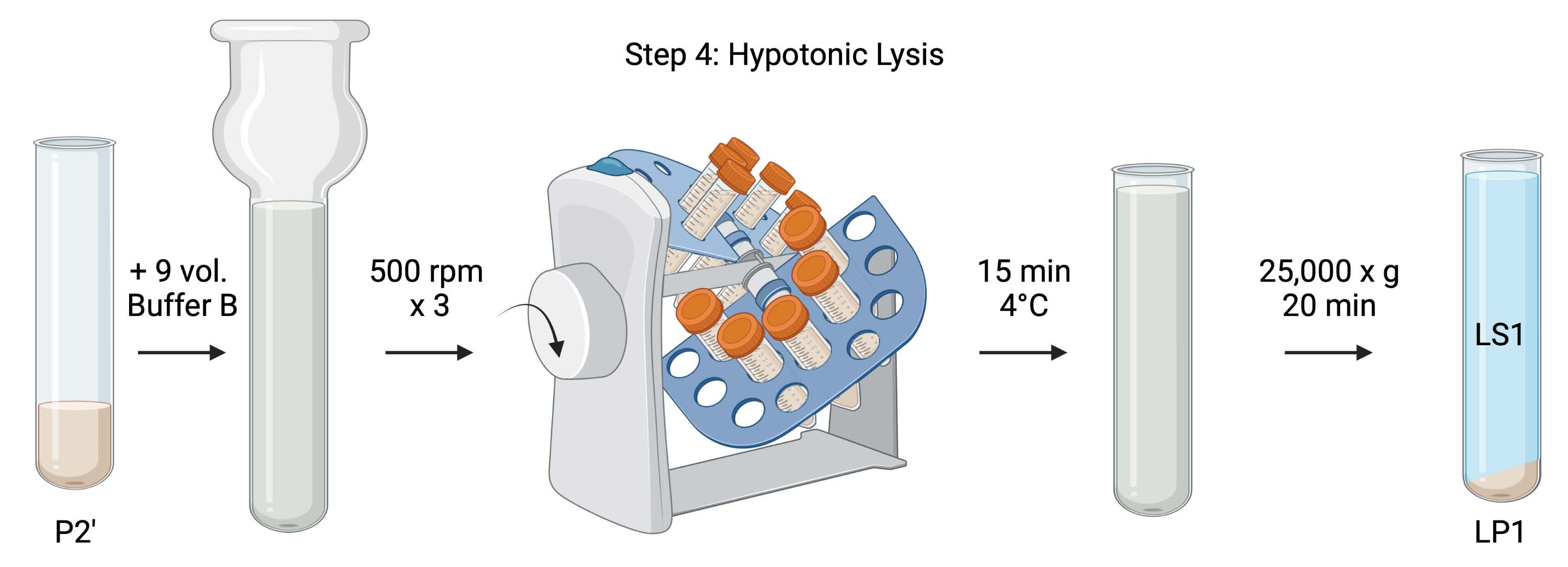{kind=link}
- Для гипотонического лизиса промытых синаптосом добавляют 9 объемов охлажденного буфера B (таблица 1) к повторно суспендированному P2' (~27 мл). Гомогенизируйте синаптосомы в стеклянном гомогенизаторе Dounce (три прохода вверх-вниз при 500 об/мин).
- Перенесите образцы в колпачковые конические центрифужные трубки объемом 50 мл. Поверните их на трубчатом револьвере в холодном помещении 4 °C в течение 15 минут.
- Центрифугу лизировали P2' при 25 000 х г в течение 20 мин при 4 °C с получением лизисного супернатанта (LS1) и лизисной гранулы, содержащей синаптосомные мембраны (LP1). Возьмите 2 x 50 мкл LS1 для BCA и 2 x 400 мкл LS1 для WB. Поместите LS1 в колпачковую центрифужную трубку для ультрацентрифугирования (см. Таблицу материалов).
5. Изоляция синаптических пузырьков
ПРИМЕЧАНИЕ: Схемы этой процедуры показаны на рисунке 6.

Рисунок 6: Выделение синаптических пузырьков и изоляция синаптической плазматической мембраны. (A) Схема стадии 5, выделение фракций синаптического цитозола (LS2) и синаптического везикулы (LP2) и (B) стадии 6, генерации миелина (MF), синаптической плазматической мембраны (SPM) и митохондриальной (Mito.) фракций после ультрацентрифугирования градиентов сахарозы. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
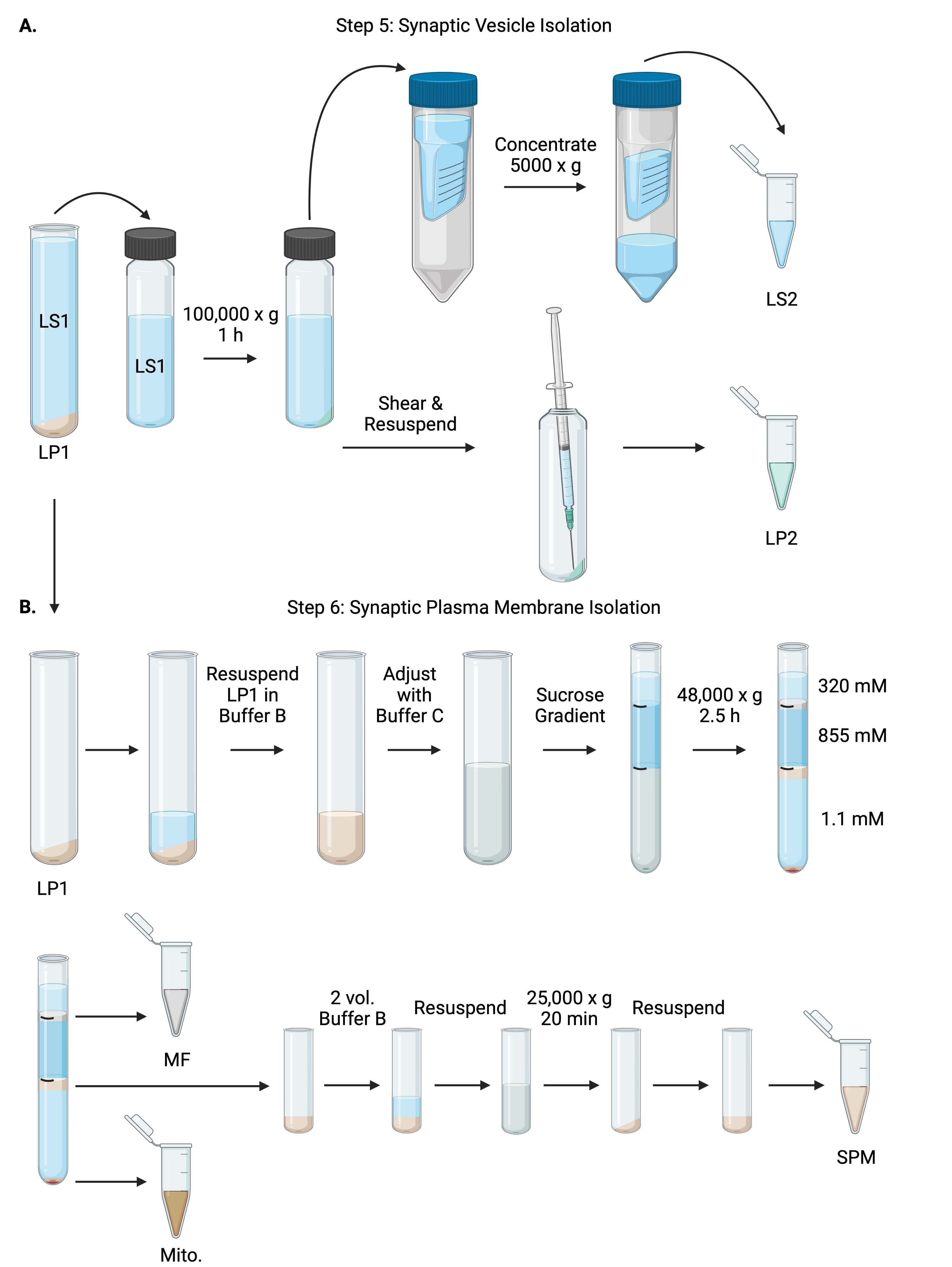{kind=link}
- Центрифуга LS1 с фиксированным углом ультрацентрифуги ротора (см. Таблицу материалов) при 100 000 х г в течение 60 мин при 4 °C с получением синаптического цитозольного супернатанта (LS2) и синаптической пузырьковой гранулы (LP2). LP2 будет маленьким, полупрозрачным и прочно прилипшим к боковой стороне трубки центрифуги.
- Повторное суспендирование LP2 в 500 мкл буфера A. Используя иглу 23 G и шприц 1 мл, сдвиг LP2 с мягким тритурацией. Возьмите 2 x 10 мкл LP2 для BCA и 2 x 250 мкл LP2 для WB.
- Перенос LS2 (~30 мл) на центробежные фильтрующие блоки с отсечкой 10 кДа (см. Таблицу материалов).
ПРИМЕЧАНИЕ: Если белки размером менее 10 кДа представляют интерес, доступны центробежные фильтрующие блоки отсечки 4 кДа, но это приведет к увеличению времени вращения. - Сконцентрировать LS2 примерно до 0,5 мл путем прядения при 5000 х г в течение 1 ч при 4 °C. Возьмите 2 x 10 мкл концентрированного LS2 для BCA и 2 x 250 мкл концентрированного LS2 для WB. После запуска спина перейдите непосредственно к шагу 6.1.
6. Синаптическая изоляция плазматической мембраны
- Повторное суспендирование LP1 (шаг 4.3) в 1 мл буфера B (таблица 1). Возьмите 2 x 10 мкл LP1 для BCA и 2 x 50 мкл LP1 для WB. Отрегулируйте оставшийся LP1 до конечного объема 7,5 мл и конечной концентрации сахарозы 1,1 М с буфером B и буфером C (таблица 1).
- Перенесите 7,5 мл повторного суспендированного LP1 в ультрацентрифужную трубку объемом 14 мл (см. Таблицу материалов). Осторожно наложите LP1 с 3,75 мл буфера D (таблица 1), а затем наложите 1,25 мл буфера A (или большего объема, чтобы заполнить чуть ниже верхней части трубки центрифуги). Избегайте пипетки вниз по боковой стороне трубки, которая нарушит границы градиента сахарозы. После наложения каждой фракции сахарозы пометьте верхнюю часть раствора ручкой. Балансируйте пробирки для ультрацентрифугирования по весу, а не по объему, с добавлением буфера А по каплям в пределах 10 мг. Центрифуга при 48 000 х г в течение 2,5 ч при 4 °C в качающемся роторе ультрацентрифуги (см. Таблицу материалов).
- Получите изображения интактных градиентов после ультрацентрифугирования, чтобы задокументировать отчетливость каждого интерфейса сахарозы и успех фракционирования.
- Аккуратно удаляют поверхностный слой сахарозы 320 мМ (буфер А). Восстановите фракцию миелина (MF) на границе раздела сахарозы 320 мМ/855 мМ в объеме 800 мкл. Восстановление фракции синаптической плазматической мембраны (SPM) на границе раздела сахарозы 855 мМ/1,1 М в объеме 1000 мкл. Пипетка каждой фракции поднимается от стенки трубки по кругу, чтобы обеспечить сбор полной фракции. Осторожно аспирируйте оставшуюся сахарозу и восстановите митохондриальную гранулу (Mito.) путем повторного использования в 200 мкл буфера B. Возьмите 2 x 100 мкл MF для BCA и 2 x 10 μL Mito. для BCA; разделите оставшуюся часть MF и Mito. выборки пополам для ВБ.
- Разбавляют фракцию СЗМ 2 объемами буфера В (~2 мл), а затем центрифугу в роторе с фиксированным углом в центрифужной трубке объемом 3,5 мл (см. Таблицу материалов) при 25 000 х г в течение 20 мин при 4 °C. Выбросьте супернатант и повторно суспендируйте гранулу SPM в буфере А для окончательного объема 250 мкл. Возьмите 2 x 5 мкл SPM для BCA и разделите оставшийся SPM пополам для WB.
- Выполните BCA для определения концентрации белка в каждом образце, учитывая переменный объем аликвоты.
ПРИМЕЧАНИЕ: Для анализа WB рекомендуемая концентрация рабочего белка для всех субклеточных фракций составляет 2 мкг/мкл (или настолько высокая, насколько это достижимо для LS1 и MF).
Результаты
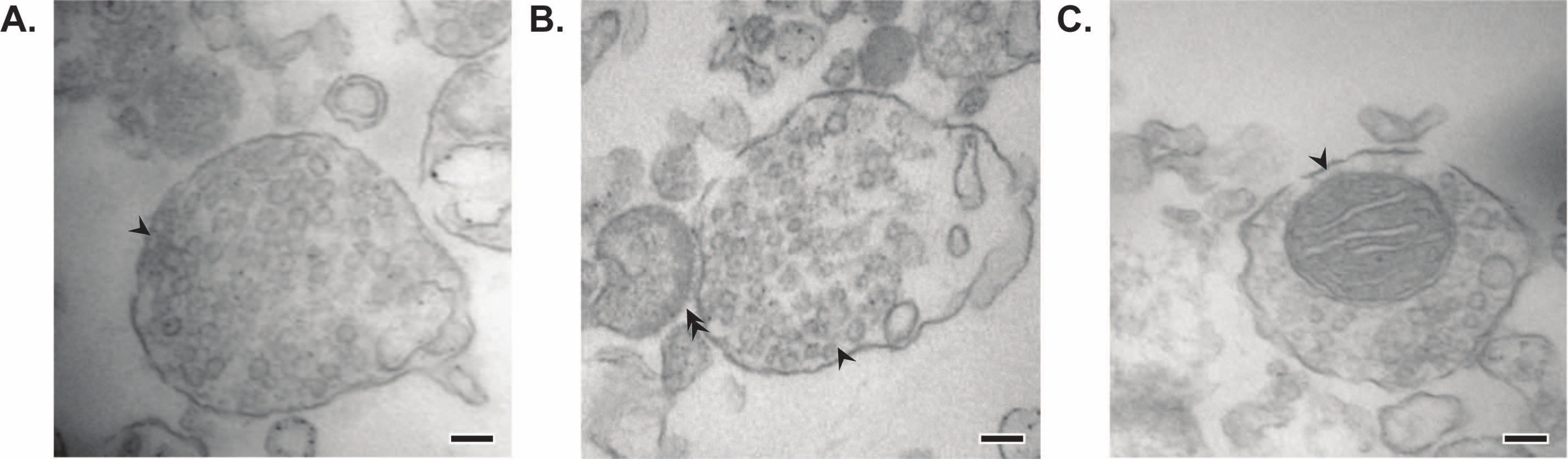
Представленный метод приводит к 11 субклеточным фракциям мозга, которые могут быть подвергнуты дальнейшей очистке и различным формам последующего анализа35,36. Золотым стандартом метода оценки качества синаптосом, SV23 и других компонентов является электронная микроскопия (ЭМ) (рисунок 7). Количественное иммуноблоттинг для белков, присутствующих в специфических субклеточных фракциях, также может быть выполнено для оценки маркеров чистоты фракций (рисунок 8). Например, иммуноблотический анализ фракций выявляет обогащение N-кадгерина (CDH2, название UniProt) в фракции синаптической плазматической мембраны (SPM), α-синуклеина (SYUA) в синаптическом цитозоле (LS2), синаптофизина (SYPH) в фракции синаптических везикул (LP2) и основного белка миелина (MBP) в фракции миелина (MF) по сравнению с уровнями белка в исходном гомогенате всего мозга (Total) (Рисунок 8) ). После установления чистоты фракции (например, обратите внимание на отсутствие CDH2 во фракции LS2 или многократное увеличение SYPH во фракции LP2), количественное иммуноблоттинг может быть использовано для определения локализации интересующих белков или запроса различий в распределении белка между генотипами или методами лечения. Понимание субклеточной локализации синаптических белков может позволить рассечение ранее не описанных функций белка. Кроме того, этот метод может прояснить дефекты трафика или синаптическую дисфункцию в болезненных состояниях, особенно в сочетании с функциональными анализами. Например, наша команда использовала этот метод для идентификации пула ферментативно активного пальмитоилового белка тиоэстеразы 1, который обогащен синаптическим цитозолем19.

Рисунок 7: Электронная микроскопия (ЭМ) синаптосом. (А) Репрезентативное ЭМ-изображение синаптосомы, содержащей синаптические везикулы (стрелка). (B) Репрезентативное ЭМ-изображение синаптосомы с пре- (стрелка) и постсинаптическим компонентами (двойная стрелка). (C) Репрезентативное ЭМ-изображение синаптосомы, содержащей синаптические везикулы и митохондрию (стрелка) (шкала = 100 нм). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 8: Иммуноблотический анализ субклеточных фракций. (А) Маркеры чистоты субклеточной фракции (обозначены номенклатурой UniProt) соответствующим образом локализованы по сравнению со всем гомогенатом мозга (тотальным): N-кадгерин (CDH2) в фракции синаптической плазматической мембраны (SPM), синаптофизин 1 (SYPH) и синаптоянин 1 (SYNJ1) в синаптической фракции, обогащенной везикулами (LP2), α-синуклеин (SYUA) в синаптом цитозоле (LS2), и основной белок миелина (MBP) в миелиновой фракции (MF). (B) Анализ количественной оценки иммуноблота выявляет обогащение (кратное изменение от общего) маркеров чистоты фракций. Данные представляются в виде среднего ± стандартного отклонения на шкале log10. Пунктирная линия указывает на изменение в 1,5 раза (y = 0,176) (n = 3 эксперимента с репликацией с 8 мышами дикого типа; возраст = 2 месяца; n = 4-5 пятен для SYPH, SYUA, MBP, с n = 3 построенными значениями, ранее опубликованными Gorenberg et al.19; n = 5 для SYNJ1; n = 1 для CDH2). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
Обсуждение
В своих основополагающих исследованиях Уиттакер и его коллеги использовали четыре морфологических критерия для идентификации синаптосом: (1) структуры имеют герметичную плазматическую мембрану; (2) структуры содержат SV, напоминающие по размеру и количеству в нервных окончаниях и варикозном расширении вен in situ; (3) структуры обладают одной или несколькими небольшими митохондриями; и (4) пресинаптическая мембрана часто прилипает к постсинаптическому компоненту 11,12,13. Хотя первые два критерия обычно применяются к каждому методу изоляции, в самых последних протоколах, описанных в этой статье, не все результирующие синаптосомы будут иметь митохондрии и прикрепленные постсинаптические терминали. Приблизительно 60% синаптосом будут иметь митохондрии, и только до 15%, по оценкам, имеют прикрепленные постсинаптические терминалы37. Если постсинаптические компоненты представляют особый интерес, то использование изотонического буфера гомогенизации, подобного Кребсу, и фильтрации под давлением для обогащения, как известно, дает высокие концентрации синаптосом с постсинаптическими терминалями (также называемыми синаптоневросомами)22,38.
Метод жертвоприношения животного может влиять на качество синаптосом и синаптических субфракций. Взрослые животные, принесенные в жертву с использованием метода эвтаназии, который не требует анестезии, приведут к наилучшему качеству фракции. Кроме того, мозг должен быть свежерассечен, а не заморожен и гомогенизирован с использованием соотношения буфера гомогенизации (вес/объем) 1:10 для наиболее жизнеспособных синаптических фракций22. Мозг имеет гетерогенную популяцию синапсов, которые можно дифференцировать по типу нейротрансмиттеров, которые они несут. Образование синаптосом, как правило, не зависит от типа синапсов или содержания нейротрансмиттеров13. Исключение составляют мшистые волокна в мозжечке, которые, как известно, разрушаются в оптимальных условиях для получения синаптосом из остальной части мозга39,40. Таким образом, удаление мозжечка перед гомогенизацией мозга рекомендуется, если исключение этой области не влияет на экспериментальную цель. Если вы заинтересованы в выделении синаптосом определенного нейротрансмиттерного характера, области мозга, которые обогащаются нейронами, содержащими интересующий нейротрансмиттер, могут быть сначала выделены. Однако такой подход наложит ограничения на выход конечной фракции в зависимости от размера интересующего региона (возраст животных также является соображением). Существуют иммунохимические методы выделения нейромедиатор-специфических синаптосом, но жизнеспособность и выход будут значительно скомпрометированы22. Если оценка метаболической жизнеспособности синаптосом важна, можно использовать измерение высвобождения нейротрансмиттера 41,42 или некоторые ферментативные анализы43.
Общие загрязняющие вещества в препаратах синаптосом включают микросомы, свободные митохондрии, SV, а также нейрональные и глиальные мембраны. Загрязнение может быть уменьшено путем увеличения количества промывок на фракциях P1 и P222 и предотвращения повторного суспендирования красной митохондриальной гранулы на последующих этапах. В экспериментах, где метаболическая жизнеспособность и время имеют решающее значение, уменьшение количества стирок и использование градиентов Фиколла или Перколла над градиентами сахарозы будет полезным 44,45,46. Эти методы также значительно снижают загрязнение. Оригинальный протокол Уиттакера дал высококачественные SV. Дальнейшая оптимизация Nagy et al.23, включенная в этот метод, производит SV с замечательной однородностью и чистотой без существенного ущерба для выхода36. Если представляют интерес специфические подтипы SV, такие как глутаматергические (VGLUT-1-содержащие) или GABAergic (VGAT-1-содержащие) SV, иммуноизолирование с использованием специфических антител может быть выполнено47,48. Альтернативные способы также доступны для выделения CCV из синаптосом, которые из-за дифференциальной плотности могут не присутствовать на том же интерфейсе, что и SV, полученные с помощью этого метода 20,49,50.
В целом, настоящий протокол выделения синаптических компонентов может быть дополнительно оптимизирован для получения фракций с улучшенной однородностью и жизнеспособностью на основе качества и количества исходной ткани мозга и экспериментальных целей. Для получения дополнительной информации об устранении неполадок следует обратиться к главам книг Данкли и Робинсона22 и Ганцеллы и др.36.
Раскрытие информации
Авторам нечего раскрывать.
Благодарности
Мы хотели бы поблагодарить P. Colosi за подготовку EM изображения. Эта работа была поддержана Национальными институтами здравоохранения (R01 NS064963, SSC; R01 NS110354, SSC; R01 NS083846, SSC; R21 NS094971, SSC; Т32 НС007224, СМТ; T32 NS041228, SMT), Министерство обороны США (W81XWH-17-1-0564, SSC; W81XWH-19-1-0264, VDJ), Aligning Science Across Parkinson's (ASAP) Collaborative Research Network (SSC) и Michael J. Fox Foundation Target Advancement Program (MJFF-020160, SSC & VDJ). Мы создавали графические иллюстрации, используя BioRender.com.
Материалы
| Name | Company | Catalog Number | Comments |
| 1 mL TB Syringe | BD | 309649 | |
| 1.5 mL Eppendorf Tubes | USA Scientific | 1415-2500 | |
| 14 mL, Open-Top Thinwall Ultra-Clear Tube | Beckman Coulter | 344060 | Compatible with SW 40 Ti |
| 23 Gauge Precision Glide Hypodermic Needle | BD | 305145 | |
| 26.3 mL, Polycarbonate Bottle with Cap Assembly | Beckman Coulter | 355618 | Compatible with Ti70 |
| 3.5 mL, Open-Top Thickwall Polypropylene Tube | Beckman Coulter | 349623 | Compatible with TLA-100.3 |
| 50 mL Falcon Tubes | Fisher Scientific | 14-432-22 | |
| Amicon Ultra-15 Centrifugal Filter Unit | Millipore Sigma | UFC901024 | |
| Aprotinin | Sigma-Aldrich | A6279 | 1 mg/mL in diH2O |
| Avanti J-26 XP Centrifuge | Beckman Coulter | B22984 | <26,000 rpm |
| Benchtop HDPE Dewar Flask | Thermo Scientific | 5028U19 | |
| C57BL/6J Mice | The Jackson Labs | 000664 | |
| Centrifuge 5810R | Eppendorf | EP022628168 | <14,000 rpm |
| complete, Mini, EDTA-free Protease Inhibitor Cocktail Tablets | Roche | 11873580001 | Add 1 tablet per 50 mL of solution |
| Curved Forceps | Fine Science Tools | 11273-20 | |
| Fine Surgical Scissors | Fine Science Tools | 8r | |
| Glas-Col Tissue Homogenizing System | Cole-Parmer | UX-04369-15 | |
| Graefe Forceps | Fine Science Tools | 11650-10 | |
| High-Speed Polycarbonate Round Bottom Centrifuge Tubes | ThermoFisher | 3117-0500 | Compatible with JA20 |
| Isofluorane | Henry Schein Animal Health | NDC 11695-6776-2 | |
| JA-20 Rotor | Beckman Coulter | 334831 | |
| Leupeptin | American Bio | AB01108 | 1 mg/mL in diH2O |
| N-[2-Hydroxyethyl] piperazine-N’-[2-ethanesulfonic acid] (HEPES) | American Bio | AB00892 | |
| Optima L-80 XP Ultracentrifuge | Beckman Coulter | <100,000 rpm | |
| Optima TLX Ultracentrifuge | Beckman Coulter | <120,000 rpm | |
| Pepstatin A | Thermo Scientific | 78436 | 1 mg/mL in DMSO |
| Phenylmethylsulfonyl fluoride (PMSF) | American Bio | AB01620 | |
| Pierce BCA Protein Assay Kit | Thermo Scientific | 23335 | For determination of protein concentration |
| Pipette Tips | |||
| Serological Pipettes | |||
| Sucrose | Sigma-Aldrich | S0389 | |
| Surgical Scissors | Fine Science Tools | 14002-12 | |
| SW 40 Ti Swinging-Bucket Rotor | Beckman Coulter | 331301 | |
| Teflon-Coated Pestle and Mortar Tissue Grinder | Thomas Scientific | 3431D94 | |
| Ti70 Rotor | Beckman Coulter | 337922 | |
| TLA-100.3 Rotor | Beckman Coulter | 349490 | |
| Tube Revolver | Dot Scientific | DTR-02VS |
Ссылки
- Synaptic Transmission. Principles of Neural Science, Fifth Edition. Kandel, E. R., Schwartz, J. H., Jessell, T. M., Siegelbaum, S. A., Hudspeth, A. J., Education, A. J. , McGraw-Hill Education. New York, NY. (2014).
- Lepeta, K., et al. Synaptopathies: synaptic dysfunction in neurological disorders - A review from students to students. Journal of Neurochemistry. 138 (6), 785-805 (2016).
- Südhof, T. C., Malenka, R. C. Understanding synapses: Past, present, and future. Neuron. 60 (3), 469-476 (2008).
- Südhof, T. C. The molecular machinery of neurotransmitter release (Nobel lecture). Angewandte Chemie International Edition. 53 (47), 12696-12717 (2014).
- Jahn, R., Boyken, J. Molecular Regulation of Synaptic Release. Neuroscience in the 21st Century: From Basic to Clinical. Pfaff, D. W. , Springer. New York. New York, NY. 351-401 (2013).
- Xiong, H., Gendelman, H. E. Current Laboratory Methods in Neuroscience Research. , Springer. New York. New York, NY. (2014).
- Azevedo, F. A., et al. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. Journal of Comparative Neurology. 513 (5), 532-541 (2009).
- Herculano-Houzel, S. The remarkable, yet not extraordinary, human brain as a scaled-up primate brain and its associated cost. Proceedings of the National Academy of Sciences of the United States of America. 109, 10661-10668 (2012).
- Herculano-Houzel, S., Lent, R. Isotropic fractionator: A simple, rapid method for the quantification of total cell and neuron numbers in the brain. Journal of Neuroscience. 25 (10), 2518-2521 (2005).
- Herculano-Houzel, S., Mota, B., Lent, R. Cellular scaling rules for rodent brains. Proceedings of the National Academy of Sciences of the United States of America. 103 (32), 12138-12143 (2006).
- Gray, E. G., Whittaker, V. P. The isolation of nerve endings from brain: An electron-microscopic study of cell fragments derived by homogenization and centrifugation. Journal of Anatomy. 96, 79-88 (1962).
- Gray, E. G., Whittaker, V. P. The isolation of synaptic vesicles from the central nervous system. Journal of Physiology. 153, 35-37 (1960).
- Whittaker, V. P. Thirty years of synaptosome research. Journal of Neurocytology. 22 (9), 735-742 (1993).
- Jahn, R., Fasshauer, D. Molecular machines governing exocytosis of synaptic vesicles. Nature. 490, 201-207 (2012).
- Whittaker, V. P., Michaelson, I. A., Kirkland, R. J. The separation of synaptic vesicles from nerve-ending particles ('synaptosomes). Biochemical Journal. 90 (2), 293-303 (1964).
- De Robertis, E., Rodriguez De Lores Arnaiz, G., Pellegrino De Iraldi, A. Isolation of synaptic vesicles from nerve endings of the rat brain. Nature. 194, 794-795 (1962).
- De Robertis, E., Pellegrino De Iraldi, A., Rodriguez, G., Gomez, C. J. On the isolation of nerve endings and synaptic vesicles. The Journal of Biophysical and Biochemical Cytology. 9 (1), 229-235 (1961).
- Zimmermann, H., Whittaker, V. P. The Discovery of the Synaptosome and Its Implications. Synaptosomes. Murphy, K. M. , Springer. New York. New York, NY. 9-26 (2018).
- Gorenberg, E. L., et al. Identification of substrates of palmitoyl protein thioesterase 1 highlights roles of depalmitoylation in disulfide bond formation and synaptic function. PLoS Biology. 20 (3), 3001590(2022).
- Vidyadhara, D. J., et al. Dopamine transporter and synaptic vesicle sorting defects initiate auxilin-linked Parkinson's disease. bioRxiv. , (2022).
- Schrimpf, S. P., et al. Proteomic analysis of synaptosomes using isotope-coded affinity tags and mass spectrometry. Proteomics. 5 (10), 2531-2541 (2005).
- Dunkley, P. R., Robinson, P. J. Synaptosome Preparations: Which Procedure Should I Use. Synaptosomes. Murphy, K. M. , Springer. New York. New York, NY. 27-53 (2018).
- Nagy, A., Baker, R. R., Morris, S. J., Whittaker, V. P. The preparation and characterization of synaptic vesicles of high purity. Brain Research. 109 (2), 285-309 (1976).
- Takamori, S., et al. Molecular anatomy of a trafficking organelle. Cell. 127 (4), 831-846 (2006).
- Wagner, J. A., Kelly, R. B. Topological organization of proteins in an intracellular secretory organelle: the synaptic vesicle. Proceedings of the National Academy of Sciences of the United States of America. 76 (8), 4126-4130 (1979).
- Jahn, R., Schiebler, W., Ouimet, C., Greengard, P. A 38,000-dalton membrane protein (p38) present in synaptic vesicles. Proceedings of the National Academy of Sciences of the United States of America. 82 (12), 4137-4141 (1985).
- Binotti, B., Jahn, R., Pérez-Lara, Á An overview of the synaptic vesicle lipid composition. Archives of Biochemistry and Biophysics. 709, 108966(2021).
- Siegel, D. P., Ware, B. R. Electrokinetic properties of synaptic vesicles and synaptosomal membranes. Biophysical Journal. 30 (1), 159-172 (1980).
- Whittaker, V. P., Michaelson, I. A., Kirkland, R. J. The separation of synaptic vesicles from disrupted nervending particles. Biochemical Pharmacology. 12 (3), 300-302 (1963).
- Clementi, F., Whittaker, V. P., Sheridan, M. N. The yield of synaptosomes from the cerebral cortex of guinea pigs estimated by a polystyrene bead "tagging" procedure. Zeitschrift für Zellforschung und Mikroskopische Anatomie. 72, 126-138 (1966).
- Carlson, S. S., Wagner, J. A., Kelly, R. B. Purification of synaptic vesicles from elasmobranch electric organ and the use of biophysical criteria to demonstrate purity. Biochemistry. 17 (7), 1188-1199 (1978).
- Huttner, W. B., Schiebler, W., Greengard, P., De Camilli, P. Synapsin I (protein I), a nerve terminal-specific phosphoprotein. III. Its association with synaptic vesicles studied in a highly purified synaptic vesicle preparation. Journal of Cell Biology. 96 (5), 1374-1388 (1983).
- Hawkins, P., et al. A guide to defining and implementing protocols for the welfare assessment of laboratory animals: eleventh report of the BVAAWF/FRAME/RSPCA/UFAW Joint Working Group on Refinement. Laboratory Animals. 45 (1), 1-13 (2011).
- Risling, T. E., Caulkett, N. A., Florence, D. Open-drop anesthesia for small laboratory animals. Canadian Veterinary Journal. 53 (3), 299-302 (2012).
- Deutsch, C., Drown, C., Rafalowska, U., Silver, I. A. Synaptosomes from rat brain: Morphology, compartmentation, and transmembrane pH and electrical gradients. Journal of Neurochemistry. 36 (6), 2063-2072 (1981).
- Ganzella, M., Ninov, M., Riedel, D., Jahn, R. Isolation of synaptic vesicles from mammalian brain. Methods in Molecular Biology. 2417, 131-145 (2022).
- Dunkley, P. R., et al. A rapid Percoll gradient procedure for isolation of synaptosomes directly from an S1 fraction: homogeneity and morphology of subcellular fractions. Brain Research. 441 (1-2), 59-71 (1988).
- Schwartz, R. D., Skolnick, P., Hollingsworth, E. B., Paul, S. M. Barbiturate and picrotoxin-sensitive chloride efflux in rat cerebral cortical synaptoneurosomes. FEBS Letters. 175 (1), 193-196 (1984).
- Pittaluga, A., Thellung, S., Maura, G., Raiteri, M. Characterization of two central AMPA-preferring receptors having distinct location, function and pharmacology. Naunyn-Schmiedeberg's Archives of Pharmacology. 349 (6), 555-558 (1994).
- Israël, M., Whittaker, V. P. The isolation of mossy fibre endings from the granular layer of the cerebellar cortex. Experientia. 21 (6), 325-326 (1965).
- Khvotchev, M., Lonart, G., Südhof, T. C. Role of calcium in neurotransmitter release evoked by alpha-latrotoxin or hypertonic sucrose. Neuroscience. 101 (3), 793-802 (2000).
- Lonart, G., Janz, R., Johnson, K. M., Südhof, T. C. Mechanism of action of rab3A in mossy fiber LTP. Neuron. 21 (5), 1141-1150 (1998).
- Nicholls, D. G., Sihra, T. S. Synaptosomes possess an exocytotic pool of glutamate. Nature. 321 (6072), 772-773 (1986).
- Dunkley, P. R., Jarvie, P. E., Robinson, P. J. A rapid Percoll gradient procedure for preparation of synaptosomes. Nature Protocols. 3 (11), 1718-1728 (2008).
- Cotman, C. W., Matthews, D. A. Synaptic plasma membranes from rat brain synaptosomes: Isolation and partial characterization. Biochimica et Biophysica Acta. 249 (2), 380-394 (1971).
- Booth, R. F., Clark, J. B. A rapid method for the preparation of relatively pure metabolically competent synaptosomes from rat brain. Biochemical Journal. 176 (2), 365-370 (1978).
- Takamori, S., Riedel, D., Jahn, R. Immunoisolation of GABA-specific synaptic vesicles defines a functionally distinct subset of synaptic vesicles. Journal of Neuroscience. 20 (3), 4904-4911 (2000).
- Burger, P. M., et al. Synaptic vesicles immunoisolated from rat cerebral cortex contain high levels of glutamate. Neuron. 3 (6), 715-720 (1989).
- Blondeau, F., et al. Tandem MS analysis of brain clathrin-coated vesicles reveals their critical involvement in synaptic vesicle recycling. Proceedings of the National Academy of Sciences of the United States of America. 101 (11), 3833-3838 (2004).
- Maycox, P. R., Link, E., Reetz, A., Morris, S. A., Jahn, R. Clathrin-coated vesicles in nervous tissue are involved primarily in synaptic vesicle recycling. Journal of Cell Biology. 118 (6), 1379-1388 (1992).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены