
Method Article
Recording Temperature-induced Neuronal Activity through Monitoring Calcium Changes in the Olfactory Bulb of Xenopus laevis
* These authors contributed equally
In This Article
Summary
Here we describe a protocol for measuring and analyzing temperature responses in the olfactory bulb of Xenopus laevis. Olfactory receptor neurons and mitral cells are differentially stained, after which calcium changes are recorded, reflecting a sensitivity of some neural networks in the bulb to temperature drops induced at the nose.
Abstract
The olfactory system, specialized in the detection, integration and processing of chemical molecules is likely the most thoroughly studied sensory system. However, there is piling evidence that olfaction is not solely limited to chemical sensitivity, but also includes temperature sensitivity. Premetamorphic Xenopus laevis are translucent animals, with protruding nasal cavities deprived of the cribriform plate separating the nose and the olfactory bulb. These characteristics make them well suited for studying olfaction, and particularly thermosensitivity. The present article describes the complete procedure for measuring temperature responses in the olfactory bulb of X. laevis larvae. Firstly, the electroporation of olfactory receptor neurons (ORNs) is performed with spectrally distinct dyes loaded into the nasal cavities in order to stain their axon terminals in the bulbar neuropil. The differential staining between left and right receptor neurons serves to identify the γ-glomerulus as the only structure innervated by contralateral presynaptic afferents. Secondly, the electroporation is combined with focal bolus loading in the olfactory bulb in order to stain mitral cells and their dendrites. The 3D brain volume is then scanned under line-illumination microscopy for the acquisition of fast calcium imaging data while small temperature drops are induced at the olfactory epithelium. Lastly, the post-acquisition analysis allows the morphological reconstruction of the thermosensitive network comprising the γ-glomerulus and its innervating mitral cells, based on specific temperature-induced Ca2+ traces. Using chemical odorants as stimuli in addition to temperature jumps enables the comparison between thermosensitive and chemosensitive networks in the olfactory bulb.
Introduction
Over the last years, temperature sensitivity has no longer been described as a somesthetic sense only, but also as a physiological function relevant for the olfactory system. In rodents, the main olfactory bulb receives input from the Grueneberg ganglion (GG), an organ in the nasal cavity, consisting of thermosensitive neurons. GG neurons respond to cool temperatures1 as well as to chemical stimuli, and their chemosensitivity is modulated by temperature fluctuations2. These observations suggest that the olfactory bulb may integrate chemical and temperature information collected at the nose. In order to explore this hypothesis, we present here a set of experiments enabling the detection of temperature responses in the olfactory bulb of non-transgenic animals, using the Xenopus laevis larva as a model. The organization of the olfactory system in these animals closely resembles that of mammals. The olfactory receptor neurons of premetamorphic X. laevis terminate in tufts, and make synaptic contacts with the dendrites of second-order neurons, the mitral cells. Pre- and postsynaptic fibers intermingle and form skein-like neuropil structures called glomeruli3. The abundant synapses of the glomerular layer represent the first processing center of olfactory information. Mitral cells further integrate the sensory input and convey it to higher olfactory areas.
We have developed a protocol combining electroporation of olfactory receptor neurons (ORNs) with calcium-sensitive and non-sensitive dyes followed by bolus loading of the postsynaptic network of glomeruli and mitral cells. The staining by electroporation of two spectrally distinct dyes loaded in the nasal cavities serves to single out the γ-glomerulus3 through its bilateral innervation by ORNs from both olfactory epithelia. Thus, the location of the γ-glomerulus is identified prior to further measurements. Subsequently, bolus loading4 with Fluo-8 acetoxymethyl (Fluo-8 AM) is carried out in a volume comprising the γ-glomerulus. Imaging calcium changes with fast confocal microscopy allows the visualization of temperature responses in the 3D neuropil surrounding the γ-glomerulus, a unique temperature-sensitive glomerulus in this system5. Mitral cells innervating this specific structure can also be identified by their Ca2+ signals responsive to induced temperature drops. Next, activity correlation imaging6 uses the specific Ca2+ traces of these cells to reveal the dendritic morphology of thermosensitive mitral cells. Alternating repeated applications of cold Ringer solution and chemical odorants in one measurement can be used to visualize the mitral cell networks for odor and temperature processing surrounding the γ-glomerulus and identify potential overlaps. To unambiguously assign the responses to either the chemical or the temperature stimulus, we constantly monitor temperature at the olfactory epithelium.
Protocol
All experiments with Xenopus laevis tadpoles were performed according to the guidelines approved by the Göttingen University Committee of Ethics in Animal Experimentation.
1. Electroporation
- Choose animals of stages 49-54 according to Nieuwkoop and Faber7.
- Make sure that the recording setup consists of a stereomicroscope with a large working distance and an electroporation device which can apply voltage pulses of 20 V for 20 msec at a frequency of at least 2 Hz. Apply the voltage pulses via two platinum electrodes with a rough diameter of 200 µm so that they can be inserted into the tadpoles' nostrils without causing damage.
- Prepare crystals of a dextran conjugated dye (e.g., Calcium Green 10 kDa Dextran, or Alexa Fluor 647 10 kDa Dextran) by dissolving the typically delivered amount of 5 mg in about 100 µl of distilled water and letting small droplets of 2 µl dry on a sheet of Parafilm.
Note: The crystals dry in less than a day and can be stored afterwards at -18 °C in the freezer for a period of over a year. - Anesthetize the animals by placing them in tap water containing 3% Tricaine methane sulfonate for 1-2 min until it reaches the surgical anesthesia state characterized by decreased heart rate, loss of all movements and lack of responses to mechanical stimuli.
- Immerse the anesthetized animal for 10 sec in pure tap water.
- Place the animal on a gel cushion and fix it by placing needles around it without harming it.
Note: Do not fix the animal by poking needles through its skin. - Gently dry the area surrounding the nostrils with a tissue.
- Place the animal under the stereomicroscope and focus on the nostrils.
- Use forceps to pick a dye crystal of size matching the nostril hole, place it in one of the nasal cavities and wait until it dissolves completely, which takes less than 1 min. If the crystals are smaller, add 2 or 3 into each cavity until they produce a non-translucent high-concentrated solution.
- Place the cathode on the skin of the tadpole and the anode into one of the nostrils.
Note: This particular setting applies to the dyes cited in step 1.3. For dyes with a different polarity, the staining results may be improved by placing the cathode into the nostrils. If unsure about the polarity of the dye, both electrodes can be placed simultaneously in the nostrils (one per nostril), and the polarity alternated between single voltage pulses. - Apply six pulses of 20 V and 20 msec with approximately 0.5 sec of stimulus interval.
Note: Small bubbles should appear around the electrode in the nostril during the electroporation provided that the electrodes are in contact with the skin and the solution in the nostril. If no bubbles are visible check the connecting cables and make sure the electroporating device is delivering the desired voltage pulse. - Repeat the procedure (1.9-1.11) for the second nostril.
- Transfer the electroporated tadpole into a beaker filled with tap water at room temperature. Wait 5 to 10 min till the animal regains consciousness and resumes its swimming. Feed it and let it recover for at least 1 day during which the dye is transported along the olfactory nerve to the olfactory bulb.
- Use the electroporated animals within 1-7 days following the electroporation for the best imaging results. Give at least 24 hr time for dye transport and recovery before imaging.
2. Whole Mount Preparation
- Anesthetize the animal in tap water containing 3% Tricaine methane sulfonate until all movements have stopped and it no longer responds to mechanical stimulations.
- Transfer the tadpole to a gel cushion under a stereomicroscope and fix it tightly by poking needles through the skin on each side of the forebrain.
- Sacrifice the tadpole by severing its spinal cord.
- Use a scalpel to dissect out a block of tissue containing both nasal cavities, the olfactory nerves and olfactory bulbs (Figure 1A). Make the first incision close to left nostril-without touching it-and move the blade forward cutting alongside the left olfactory nerve and bulb until the telencephalic-diencephalic border. Do likewise on the right side. Isolate the preparation from the rest of the nervous system by making a final cut posterior to the telencephalon.
- Dispose of the body of the tadpole and carefully flip the tissue block upside down so that the ventral side of the brain preparation faces upwards.
- Pin the tissue block again by inserting two needles between the olfactory nerves.
- Put a drop of Frog Ringer solution (98 mM NaCl, 2 mM KCl, 1 mM CaCl2, 2 mM MgCl2, 5 mM glucose, 5 mM Na-pyruvate, 10 mM HEPES, adjusted to pH 7.8, osmolarity of 230 mOsmol/L) on the tissue.
- Remove the meninges covering the axon sorting zone and the olfactory bulbs by making three incisions using fine scissors. Starting from the posterior edge of the tissue block, cut caudorostrally closely along the left olfactory bulb until the entrance point of the olfactory nerves into the bulbs (the axon sorting zone).
- Repeat step 2.8 for the right hemisphere.
- Raise the meninges with forceps and make the third cut, perpendicular to the previous ones at the axon sorting zone.
Note: The ventral side of the olfactory bulb is now accessible for imaging and bolus loading. - To improve the image quality in the transmitted light image, repeat the procedure for the dorsal side. These extra steps can facilitate the navigation of the micropipette for bolus loading, but are not strictly necessary.
- Transfer the sample to a recording chamber filled with Frog Ringer for further processing and imaging. Make sure that the ventral side is facing upwards and secure the sample with a net of nylon fibers spanned over a small platinum frame.
- For an easier application of stimuli place the nasal cavities on top of one of the nylon fibers.
3. Bolus Loading
- Dissolve 50 µg of the calcium AM dye (e.g., Fluo-8 AM) in 20 µl of dimethyl sulfoxide (DMSO) containing 20% of Pluronic F-127 (w/v) to prepare a stock solution of AM dye. Freeze the stock solutions in small aliquots of 1-2 µl volume. The stock solution is stable for at least half a year but avoid freeze-thaw cycles.
- Use a micropipette puller and pull pipettes with a resistance of 5-8 MΩ and a tip diameter of 1-2 µm.
- Dissolve the prepared stock solution of the dye in Frog Ringer's solution at a concentration of 250-500 µM.
- Add MK571 to reach a concentration of 500 µM to block multidrug resistance transporters.
- Fill the micropipette with 10 µl of the solution using an elongated pipette tip and remove all air bubbles by flicking the micropipette.
- Mount the micropipette in the pipette holder and make sure that pressure can be applied either manually with a syringe or a pneumatic drug ejection device and monitor the applied pressure with a gauge.
- If possible, use a microscope setup with the capability to excite the injected dye so that the outflow from the micropipette tip can be visualized and adjusted. Ideally, use a confocal imaging setup. Use an upright microscope with an immersion objective so that the tissue can be reached with the micropipette.
- Place the whole mount preparation under the microscope, follow the olfactory nerve until the bulb and focus onto the area of interest in the olfactory bulb.
- Perfuse fresh Ringer solution through the recording chamber in order to increase the viability of the preparation and wash out the dye leaking from the micropipette tip.
- Lower the pipette onto the surface of the olfactory bulb, with the tip pointing into the rostral direction of the preparation.
- Apply a small and constant positive pressure (~25 hPa) to the micropipette to prevent clogging of the pipette and gently insert it into the tissue.
- Once the pipette has breached the outer tissue layers, move it in a rostro-dorsal direction into the mitral cell layer. (Note that a counter staining of olfactory receptor neurons by electroporation is helpful to adjust the micropipette's position.) Ideally, place the pipette tip at a distance of 50-100 µm from the desired recording site.
- For staining the temperature-sensitive γ-glomerulus, target an area about 50 µm rostrally from the position of the presynaptic neuropil of the γ-glomerulus.
- Apply a positive pressure in the range of 100-200 hPa. Adjust the pressure strength depending on the size of the micropipette tip. Confirm the outflow from the pipette by watching for slight tissue movements visible under brightlight illumination when pressure is applied for the first time.
- Maintain a constant pressure for about 10 min while the micropipette remains in the tissue. If possible, check the neuropil in the meantime for cell loading under fluorescent illumination as successful loading will result in visible cell somata staining with increasing intensity over time.
Note: Often the reason for unsuccessful loading is pipette clogging due to tissue entering the tip or aggregated dye clusters. It is sometimes possible to rescue the pipette by gently breaking its tip against the lower surface of the recording chamber. However, the pipette tip should not exceed a few micrometers. Compensate larger pipette openings by applying lower pressure during bolus loading. - After the 10 min of loading, reduce the applied pressure to zero and check the staining.
Note: The size of the stained area varies significantly and depends on several parameters like amount of injected dye and location of ejection site. A good staining covers an area of about 100 µm x 100 µm. - If the stained area is too small or does not cover the desired recording site, repeat steps 3.10-3.13. Use the same micropipette for the next injection if it has not yet clogged.
- Wait at least 30 min after the last injection before starting any experiments to allow dye uptake and deesterification. Continue to perfuse the recording chamber with fresh Ringer solution at all times.
4. Measurement Settings
- Make sure that the measurement setup consists of a confocal microscope with sufficient speed to record three dimensional volumes. Choose an acquisition rate of at least 1 Hz per stack.
Note: Suitable setups include for example line-illumination microscopes scanning the sample line-wise instead of point by point. A simple realization of such a setup has been previously described6. Other options are spinning disk microscopes. If no such setup is available, it is still possible to measure smaller areas with a normal point-scanning setup. - Set the measurement parameters so that a volume large enough to fit the size of a glomerulus can be covered.
Note: The typical thickness of a recording volume is in the range of 20 µm which should be covered by at least 5 layers. - Check all presynaptic measurements for bleaching. Adjust the laser power until the average fluorescence intensity of the recorded images does not drop over the time course of the recording.
- Limit the measurement time and area for recordings on the postsynaptic side to avoid bleaching as much as possible, although for measurements exceeding 20-30 sec bleaching is likely to occur.
5. Odor Application and Temperature Experiments
- Pour 25 ml of fresh Ringer solution (previously stored at 4 °C) in a 50 ml tube. Place the tube in an ice bucket.
- Monitor the temperature of the cooled Ringer by inserting the clean probe of a thermometer into the tube. Wait until the temperature drops below 1 °C before starting the experiment.
- Prepare 50 ml of L-histidine dissolved in Ringer solution at a concentration of 10 µM.
- Use a funnel applicator8 or similar application systems allowing stimulus delivery concomitantly to the perfusing Ringer so that the water flow in the chamber remains constant and uninterrupted during the release of the stimulus solution. Position the funnel in such a way that the distal outlet is less than 1 mm away from the olfactory epithelium.
- Place a NiCr-Ni temperature sensor connected to a digital thermometer close to the epithelium and the outlet of the funnel applicator. Wire the thermometer output port to a computer to record and visually display voltage changes reflecting small temperature fluctuations.
- Before starting the experiment, connect another thermosensor to a standard thermometer to establish the voltage-to-temperature scale factor.
- Survey the bath temperature in the recording chamber and ensure that it does not exceed 22 °C.
- Start the image acquisition and sequentially apply 200-400 µl of cold Ringer, L-histidine and room-temperature Ringer (20-22 °C) via an electronic pipette with an interstimulus interval of 20-30 sec. For a better control of the stimulus application, release the stimulus with a trigger signal sent by the chosen imaging setup to the pipette if possible. Repeat the application protocol for reproducible results.
- Take sets of longer recordings with several rounds of stimulation since they are preferable for post-imaging analysis with activity correlation imaging.
Note: A compromise between the recording time and the slice viability has to be found. Measurements of ca. 2 min covering 6 stimulus applications provide sufficient activity for a good reconstruction of the network.
6. Image Processing Using Activity Correlation Imaging (ACI)
- Image Processing of the Pre-synaptic Recordings
- In recordings with stimulation by cold Ringer solution and histidine, distinguish the temperature-sensitive γ-neuropil from the neighboring histidine-sensitive glomerulus by their different response profiles (see Kludt et al.5).
- Create a maximum intensity projection in the axial direction from recordings of the brain preparation in which ORNs have been electroporated with two different dyes to visualize the bilateral innervation of the γ-glomerulus.
- Image Processing of the Post-synaptic Recordings Using Activity Correlation Imaging (ACI)
- Select recordings with sufficient time points (more than 100 frames) and a decent amount of activity (at least two events).
Note: The activity can be either spontaneous to reveal the processes of single mitral cells or induced by several applications of stimuli during the same recording to reveal the cellular networks activated by the olfactory stimuli. - Choose recordings where the measured structures move no more than 2-3 pixels over the time course of the recording.
Note: Recordings with too much movement can be rescued by applying a shift correction procedure described in Kludt et al.5 - Check whether the recordings suffer from bleaching. If the average intensity of the whole image drops over the time course of the recording, bleach correction is necessary, otherwise skip the next two steps.
- Calculate the linear trend for the time trace of each pixel by performing a linear regression.
- Subtract the result from each pixel individually to eliminate the linear component of the bleaching.
Note: As an alternative Legendre fitting can be used as described by Bao and Schild9. - Download the ready-to-use MATLAB script together with a step-by-step guide for activity correlation imaging (ACI) as described by Junek et al.6
Note: Follow the steps described in Junek et al.6 as an alternative to using the provided MATLAB script. - Move the files 'aci.m', 'matVis.m' and 'aci_roiSelector.m' from the downloaded container to the MATLAB path of the system used for data evaluation.
- Load the raw data acquired in step 5.8 as a variable to MATLAB's user workspace organized as a [x,y,z,t]-matrix with x and y referring to the lateral dimensions, z, the axial direction and t, the time course.
- Call 'aci' from the MATLAB command line.
- In the user interface (UI) which appears, select 'Prepare Data', then select the variable containing the data and a directory in which the results will be saved.
Note: For a full description of the UI see the manual accompanying the aci script. - Scroll through the measured z-layers by moving the corresponding slider in the UI to get an overview of the variance map displayed.
- Enter the size of the region of interest (ROI) into the UI. For a mitral cell soma adjust the ROI to span approximately 10 µm in the lateral and 5 µm in the axial direction. For a glomerulus, the slightly higher values of 20 µm laterally and 10 µm axially are appropriate.
Note: The ROI size has to be entered as a number of pixels, which depends on the pixel scaling chosen for the recording. - Select a ROI containing the γ-glomerulus and additional regions for each visible soma of the surrounding mitral cells by clicking with the middle mouse button onto the center of the cell/glomerulus.
- Close the main UI which triggers the calculation of correlation maps for all reference traces. The result is automatically saved and displayed.
- Select recordings with sufficient time points (more than 100 frames) and a decent amount of activity (at least two events).
Results
The electroporation of olfactory receptor neurons (ORNs) was achieved with Alexa Fluor dyes or calcium indicators conjugated with Dextran molecules for anterograde labelling via active axonal transport. While the former dyes provide bright staining of the sensory neurons and their axon terminals branching in the glomerular layer of the bulb, the latter allow the measurement of neuronal activity in these cells (Figures 1 and 2). First, the position of the thermosensitive γ-glomerulus and its innervation pattern was visualized by electroporating Alexa Fluor 647 Dextran and Alexa Fluor 546 Dextran in the left and right olfactory epithelia, respectively (Figure 1A). Twenty-four hr after the procedure, the ORNs in the nostrils, the two olfactory nerves and the glomeruli in both hemispheres were visible under fluorescent microscopy. The different glomerular clusters were identifiable by their respective positions, notably the small cluster comprising the γ-glomerulus (Figure 1B). A small number of contralateral olfactory fibers ran through the contralateral olfactory bulb, crossed the anterior commissure and terminated in the ipsilateral γ-glomerulus (Figure 2A).
In order to record calcium responses of the presynaptic fibers of the γ-glomerulus, Calcium Green Dextran was electroporated in ORNs, according to the same procedure. Negative temperature jumps were induced at the nostril via the controlled release of ice-cold Ringer solution (0-1 °C). A 3D volume comprising the γ-glomerulus was imaged under a fast line-illumination confocal microscope. ΔTs of -1 °C were sufficient to trigger cold responses in the γ-glomerulus and its afferents, recognizable as reversible peaks in the ΔF/F Ca2+ traces (Figure 2B, C).
Further experimental steps were carried out to measure cold-induced activity in the mitral cells connected to the γ-glomerulus via their ramifying dendritic endings. These postsynaptic fibers and the surrounding neuropil were effectively stained by bolus loading of the calcium-sensitive dye Fluo-8 AM, performed a few days after Alexa 647 Dextran had been electroporated into ORNs (Figure 3A, B). Mitral cells were filled with Fluo-8 AM, and the olfactory epithelium was stimulated twice according to the following paradigm: cold Ringer, histidine (10 µM) and room-temperature Ringer, applied subsequently. Two reference traces were taken from regions of interest in the recorded volume, one responding exclusively to temperature drops, the other one, to histidine only. Activity correlation imaging (ACI)6 was computed based on the selected reference traces to visualize with high contrast the dendritic morphology of the postsynaptic networks corresponding either to the temperature or the histidine-responsive Ca2+ signal (Figure 3C, D). Finally, thermosensitive and chemosensitive maps were color-coded and overlaid on top of each other, showing how temperature and chemical information is conveyed from individual glomeruli to the olfactory second-order neurons (Figure 3E). For a description of the integration and processing of both types of information in shared olfactory networks, see Kludt et al.5

Figure 1: Overview of the ORN electroporation and bolus loading. (A) Olfactory receptor neurons in both nasal cavities of X. laevis larvae were electroporated with Alexa dyes or calcium-sensitive dyes coupled to Dextran molecules. The fluorescent indicators were transported anterogradely until the terminal axonal arborization. 24 hr after electroporation, the glomerular layer in both hemispheres showed fluorescent staining. (B) Schematic of the cellular organization of the olfactory bulb in one hemisphere. The glomerular layer spans the bulb in clusters: the medial, small, intermediate and lateral clusters. Olfactory information is transferred from the receptor neurons to mitral cells via excitatory synapses in glomeruli. Periglomerular cells and granule cells are inhibitory neurons modulating olfactory processing and encoding. The γ-glomerulus (cyan) was easily identified as the small neuropil where ipsilateral (red) and contralateral (orange) olfactory fibers merged. (C) Bolus loading was achieved in the vicinity of the γ-glomerulus to stain the post-synaptic neuropil consisting primarily of mitral cells and their dendritic trees branching considerably in the glomerular layer. Please click here to view a larger version of this figure.
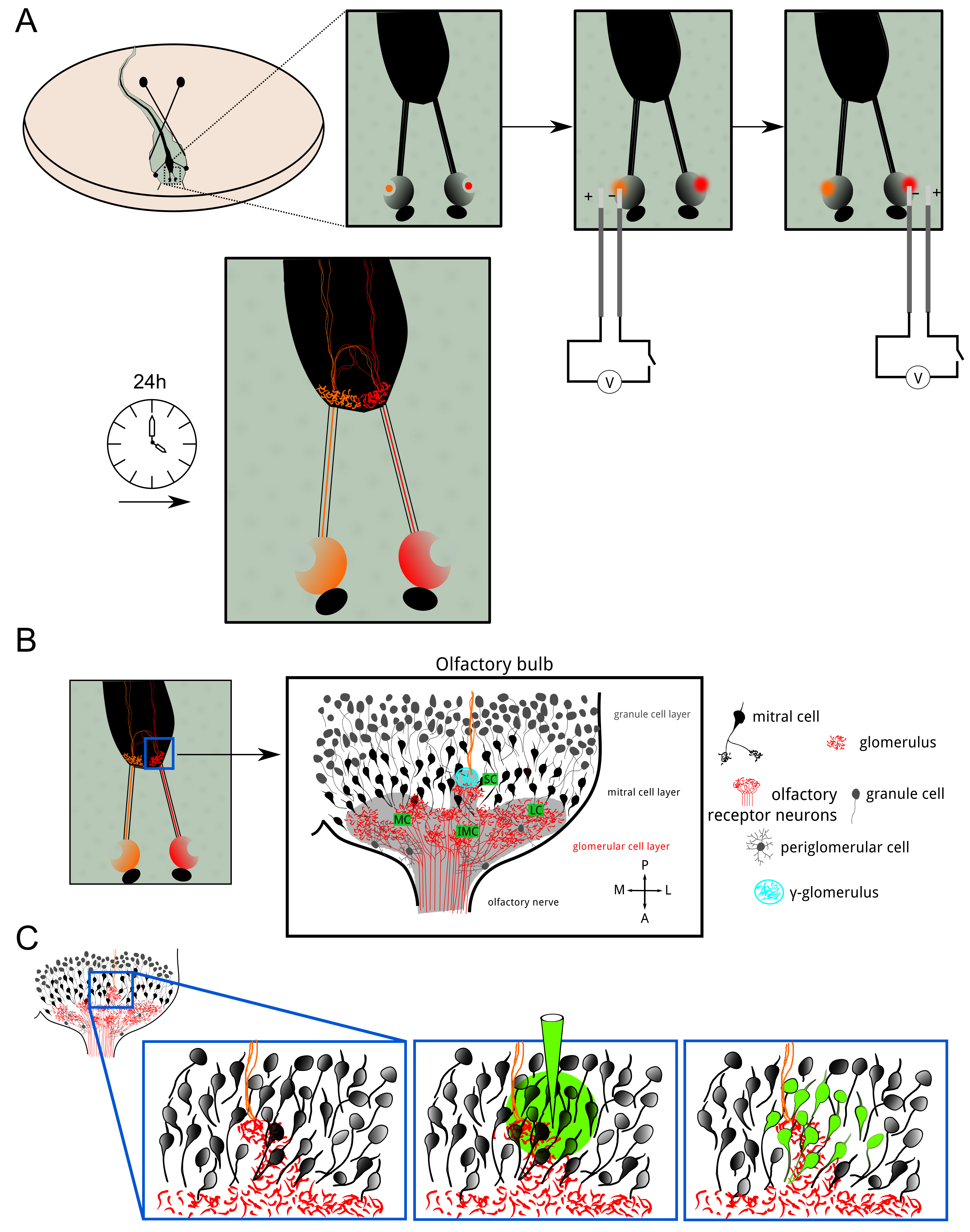{kind=link}

Figure 2: Electroporation of ORNs reveals structural and functional connectivity. (A) Bilateral innervation of the γ-glomerulus by ipsilateral (green) and contralateral (red) olfactory receptor neurons (ORNs). Scale bar = 50 µm. (B) The olfactory bulb after electroporation with the calcium-sensitive dye Calcium Green Dextran. The intermedial (IMC), medial (MC) and the small cluster (SC) are visible. The image is a maximum projection of 100 µm-thick measurement volume. Scale bar = 50 µm. (C) Close-up view of the small cluster. The recorded volume (12 µm) is represented in a maximal projection. The image depicts the basal fluorescent level in gray and the color-coded ΔF/F map as an overlay. The maximum response to stimulation with cold Ringer solution is plotted. The γ-glomerulus reacted strongly while the two neighboring glomeruli remain silent. The inset shows the ΔF/F trace for the γ-glomerulus corresponding to the indicated region of interest. The blue bar represents the stimulus application. Scale bar = 20 µm. Please click here to view a larger version of this figure.
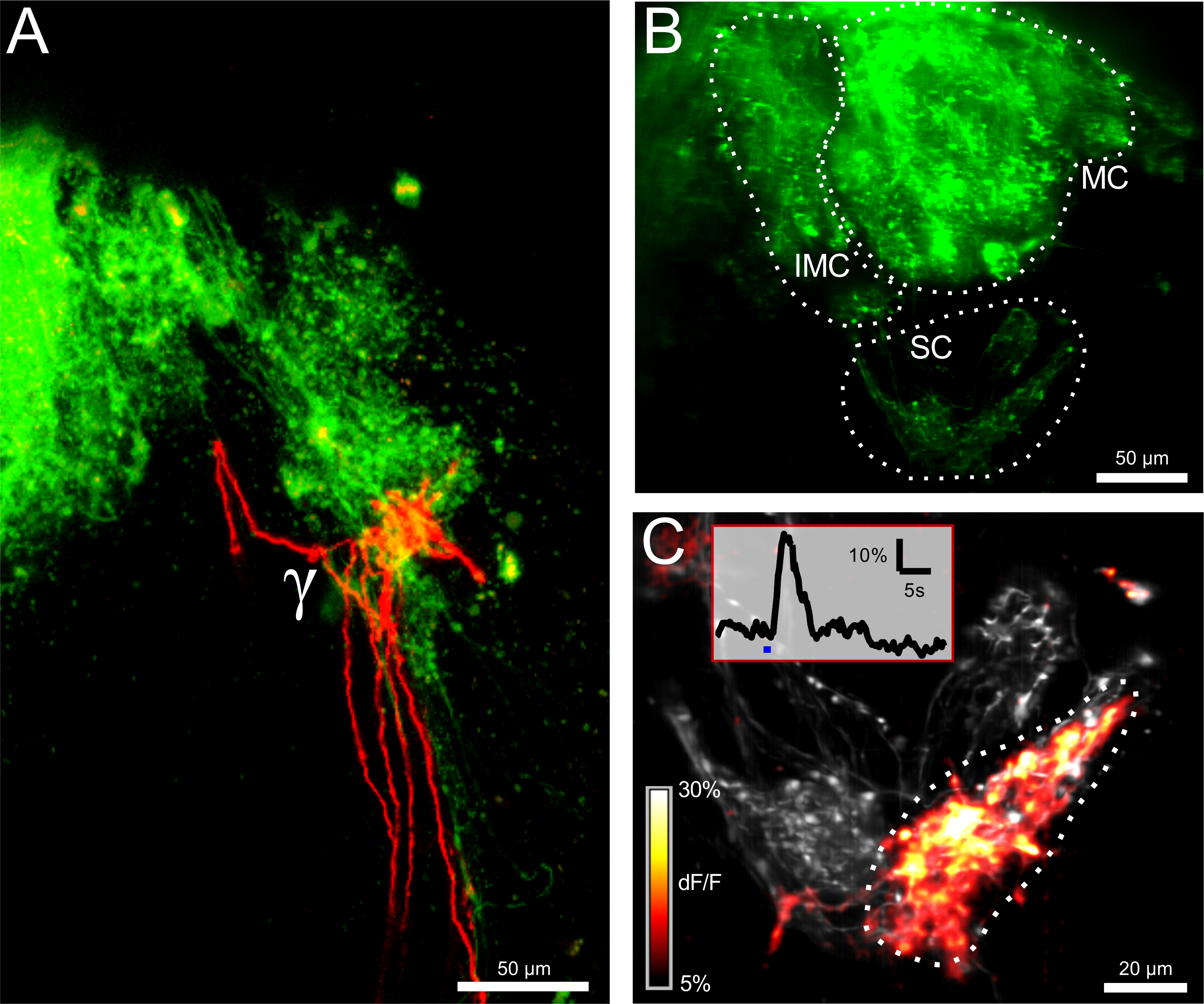{kind=link}

Figure 3: Bolus loading and ACI dissociate thermosensitive and chemosensitive networks. (A) Axons of ORNs terminating in the small cluster were stained by electroporation with the non-calcium sensitive dye Alexa 647 Dextran. The dotted line outlines the γ-glomerulus. (B) Image of the same region as in (A) in the second measurement channel after bolus loading with the calcium-sensitive dye Fluo-8 AM. Some mitral cell somata were visible but the contrast was limited. Following the arrows, two response traces were plotted, which were used for activity correlation imaging (ACI). Blue, red and black bars below the traces depict the onset of application of cold Ringer, histidine (10 µM) and room-temperature Ringer as negative control, respectively. The two Ca2+ traces were taken from different regions of interest of the measured volume. (C) The ACI result of the trace in (B) highlighting areas responding predominantly to cold Ringer. (D) The ACI result of the trace in (B) highlighting areas responding predominantly to histidine. (E) Overlay of the two ACI maps. Mitral cells responding to histidine and the innervated glomerulus (red) were easily distinguishable from thermosensitive mitral cells and the γ-glomerulus (cyan). All images of this figure are maximum intensity projections of a 28 µm-thick volume. Scale bar = 20 µm. Please click here to view a larger version of this figure.
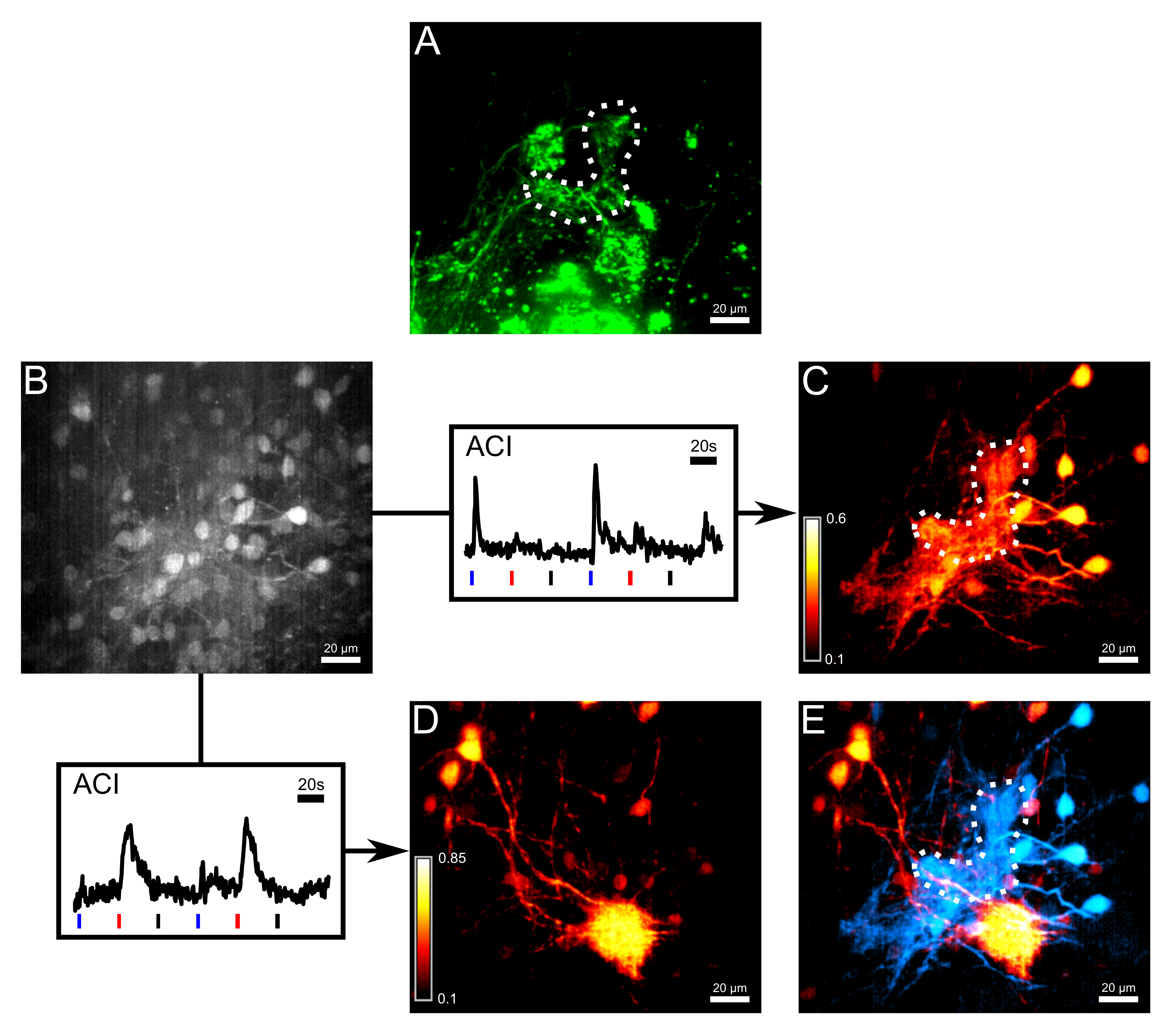{kind=link}
Discussion
The methods presented herein aim at recording temperature processing in the olfactory bulb of Xenopus laevis tadpoles. The protocol stains first and second order neurons in the olfactory bulb and provides a sample preparation in which the olfactory system remains mainly intact. Thus, the activation of the temperature-sensitive γ-glomerulus can be monitored and compared with its chemo-sensitive neighboring glomeruli. The unique bilateral innervation of this glomerulus is visualized by cell electroporation with spectrally different dyes. Furthermore, bolus loading allows for the staining of mitral cells spanning a large volume within the olfactory bulb. The neuronal network processing temperature-induced signals is revealed by taking calcium measurements with repeated stimulus applications and subsequently analyzing the data with activity correlation imaging.
The protocol highlights two sophisticated staining procedures, both of which require cautious manipulation and practice in order to achieve satisfying and reproducible results. During the electroporation any injury of the animals has to be avoided, especially when positioning the electrodes into the nostrils. Optimally, no contact with the olfactory epithelium should occur. Note that the animals are still living after the electroporation procedure and their recovery time must be taken into account. If the staining remains too weak after one round of electroporation, which can happen depending on the types of dyes used, its intensity can be enhanced by increasing dye concentration in the nostrils. Since dextran-coupled molecules are transported via several mechanisms including slow axonal transport (at speed of 1-2 mm/day10) and passive diffusion, another alternative is to wait 48 hours after electroporation before sacrificing the animals. Alternatively, the electroporation may be repeated after one day of recovery.
Bolus loading is a critical step since the amount of dye entering the mitral cells is difficult to regulate and depends on various parameters like pipette tip size and location of the application. Monitoring the procedure under a confocal fluorescence microscope proves to be useful for adjusting the duration of dye application and thus generating similar staining results across preparations. Furthermore, previously electroporated tadpoles should be used to determine the best position for the dye application by identifying the position of the small cluster (comprising the γ-glomerulus). The most critical step during measurements is to avoid both shift and bleaching of the sample. The shift can be avoided by carefully positioning the Ringer flow under the microscope. As to limit the bleaching of the area of interest, the measurement time should be reduced to the essential.
Bolus loading staining with calcium-sensitive dyes only provides very limited contrast since healthy cells generally have low calcium levels and thus show weak basal fluorescence. Applying activity correlation imaging circumvents this limitation by generating contrast based on activity and highlights structures with similar calcium signals. This post-acquisition analysis method computes the correlation factor between the calcium signal of a selected region of interest (reference trace) and that of each individual pixel in the 3D volume. Therefore, the results obtained strongly depend on the activity pattern selected as reference trace. If the main focus is to visualize mitral cell innervation patterns, a reference signal derived from spontaneous neuronal activity is preferred, and choosing the most active mitral cells will produce the best results. For revealing the chemo- or thermo-sensitive networks of mitral cells, reference traces only containing responses to either histidine or cold Ringer should be selected. The selection of an entire glomerulus or mitral cell soma as region of interest may not always provide a clear reference trace, especially if the structures responding to the two different stimuli are lying on top of each other. In such a case, it is often useful to select a smaller area of the glomerulus or cell body as region of interest.
In the last decades, electroporation has been described as an efficient method to stain single or multiple cells11,12. Here it is used to specifically label olfactory receptor neurons. Dextran-conjugated molecules give the highest efficiency, and for non-calcium sensitive dyes, the range of selection is broad and covers the complete spectrum typically used in fluorescence microscopy13. However, calcium-sensitive dyes which are successfully electroporated into olfactory receptor neurons are at the moment limited to calcium-green dextran, and Fluo-4 dextran if still commercially available. Besides, the recordings primarily target superficial layers on the ventral surface of the olfactory bulb only, since the penetration depth of fast measurement techniques is limited. Two-photon imaging can partly overcome this limitation but often lacks speed and restricts the amount of selectable calcium-sensitive dyes further.
We described here a protocol for measuring temperature-induced activity in the olfactory bulb. The brain neuropil is scanned as a three dimensional volume to visualize the complex cellular networks involved in the olfactory processing of temperature. Measuring temperature-induced activity in the olfactory bulb has been very recently reported5 and requires a specifically customized procedure combining different techniques. A major asset of the techniques presented above is that hundreds of cells are imaged in three dimensions in a preparation where most of the olfactory system remains intact. These advantages put high demands on the staining techniques as well as the brain preparation and imaging. For example, cell electroporation and bolus loading hit large amounts of cells in the olfactory epithelium and bulb, and thus enable the visualization of complete cellular networks. Furthermore, the delivery of chemical indicators via bolus loading instead of genetically encoded fluorophores enables measurements in a potentially larger set of species. Other alternatives like bath incubation with AM dyes primarily work in slices which damage the olfactory bulb severely, leaving only a few hundred micrometers of intact tissue. In comparison, the whole mount preparation used in our protocol ensures for example that the bilateral innervation of the γ-glomerulus remains intact and the recordings are thus taken in a still operative system. Finally, the imaging itself is done by line-illumination microscopy allowing the acquisition of 3D volumes. Line-illumination microscopy is one of the confocal techniques providing the highest possible acquisition rates6 which are necessary to cover a large fraction of the olfactory bulb. Slower acquisition systems may be used but have the disadvantage that the size of the recorded volume must be reduced. In recent years, other methods for fast image acquisition have been developed and can be used as alternatives14,15. Nevertheless, line-illumination microscopy remains one of the easiest methods to gain both sufficient speed and resolution. Here follows some information as guidelines for selecting suitable imaging setups. Since calcium imaging is done within from thick brain preparations, the setup should provide decent confocality and the objectives should have numerical apertures of 1.0 or higher. For a reference point, the recordings taken with the line illumination microscope correspond to images taken with a standard laser scanning microscope with a pinhole size of 0.5-1 airy units. Fast acquisition speed is desirable. A volume with a thickness of 20 µm covered by at least 5 layers, a lateral field of view of 100 µm x 100 µm and a pixel size of 0.5 µm or smaller should be scanned at a minimum speed of 1 Hz per stack. Reducing the confocality can increase the amount of photons counted and thus allows for faster acquisitions if necessary, but has the drawback of recording more out-of-focus light. However, since such an approach increases the thickness of the optical slices, it can actually facilitate the tracing of dendrites through different z-planes after application of ACI6.
The necessary tools to extensively study temperature processing in olfactory bulb networks are presented herein. Temperature-induced activity is recorded in the first and second order neurons via calcium-sensitive dyes and signals both arriving and departing from the γ-glomerulus. Furthermore, the extent to which individual mitral cells process both chemical and temperature information can be assessed. Since the preparation leaves the olfactory bulb intact, the role of the bilateral innervation in the olfactory processing can be further studied. The procedure is also useful for revealing whether and how thermo- and chemoinformation is encoded in overlapping olfactory networks5. Finally, the techniques mentioned above are not limited to the study of the temperature responses in the olfactory bulb but can be applied for a more general evaluation of the olfactory system, especially the cellular processing networks in large three dimensional volumes. Bolus loading and activity correlation imaging are powerful tools to observe and compare the activity of dozens of neurons, making them applicable to different brain networks16.
Disclosures
The authors have nothing to disclose.
Acknowledgements
This project was funded by the DFG Excellence Cluster 171, the Center for Nanoscale Microscopy and Molecular Physiology of the Brain, the Bernstein Center for Computational Neuroscience and the ENC-Network, an Erasmus Mundus Joint Doctoral Program. The authors thank Stephan Junek, Mihai Alevra and Guobin Bao for providing MATLAB codes and custom-written programs for image evaluation and data analysis.
Materials
| Name | Company | Catalog Number | Comments |
| Reagents | |||
| Sodium chloride | Merck Millipore | 1064040500 | |
| Potassium chloride | Merck Millipore | 1049360250 | |
| Calcium chloride dihydrate | Merck Millipore | 1023820250 | |
| Magnesium chloride hexahydrate | Merck Millipore | 1058330250 | |
| D(+)-Glucose | Merck Millipore | 1083371000 | |
| Sodium pyruvate | Sigma-Aldrich | P2256 | |
| HEPES | Merck Millipore | 1101100250 | |
| Calcium Green 10 kDa Dextran | Thermo Fisher Scientific | C-3713 | |
| Dextran, Alexa Fluor 647 | Thermo Fisher Scientific | D-22914 | |
| Dextran, Alexa Fluor 546 | Thermo Fisher Scientific | D-22911 | |
| Fluo-8 AM | TEFlabs | 203 | |
| MK571 | Alexis Biochemicals | 340-021-M005 | |
| MS-222 | Sigma-Aldrich | E10521 | |
| Pluronic acid F-127 | Sigma-Aldrich | P2443 | powder |
| L-Histidine monohydrochloride monohydrate | Sigma-Aldrich | 53370 | |
| DMSO | Merck Millipore | 1029522500 | |
| Equipment | |||
| Electronic pipette | BrandTech | HandyStep Electronic Repeating Pipette | |
| NiCr-Ni thermocouple | Greisinger Elektronik | GTF 300 | |
| Micropipette puller | Narishige | Model PC-10 | two-step puller |
| Funnel applicator | (Custom-made) | ||
| Line-illumination microscope | (Custom-made) | otherwise, a commercially available spinning disk microscope | |
| Objective W Plan-Apochromat 63X/1.0 | Zeiss | 441470-9900-000 | |
| Objective W Plan-Apochromat 40X/1.0 DIC | Zeiss | 441452-9900-000 | |
| Software | |||
| MATLAB | The MathWorks | from R2010b upwards |
References
- Mamasuew, K., Breer, H., Fleischer, J. Grueneberg ganglion neurons respond to cool ambient temperatures. Eur J Neurosci. 28 (9), 1775-1785 (2008).
- Brechbühl, J., Moine, F., Broillet, M. -C. Mouse Grueneberg ganglion neurons share molecular and functional features with C. elegans amphid neurons. Front. Behav. Neurosci. 7 (193), (2013).
- Gaudin, A., Gascuel, J. 3D atlas describing the ontogenic evolution of the primary olfactory projections in the olfactory bulb of Xenopus laevis. J Comp Neurol. 489 (4), 403-424 (2005).
- Stosiek, C., Garaschuk, O., Holthoff, K., Konnerth, A. In vivo two-photon calcium imaging of neuronal networks. Proc Natl Acad Sci U S A. 100 (12), 7319-7324 (2003).
- Kludt, E., Okom, C., Brinkmann, A., Schild, D. Integrating temperature with odor processing in the olfactory bulb. J Neurosci. 35 (20), 7892-7902 (2015).
- Junek, S., Chen, T. W., Alevra, M., Schild, D. Activity correlation imaging: visualizing function and structure of neuronal populations. Biophys J. 96 (9), 3801-3809 (2009).
- Nieuwkoop, P. D., Faber, J. Normal Table of Xenopus laevis. , Garland Publishing Inc. New York. (1994).
- Schild, D. A computer-controlled device for the application of odors to aquatic animals. J Electrophysiol Tec. 12 (2), 71-79 (1985).
- Bao, G., Schild, D. Fast and Accurate Fitting and Filtering of Noisy Exponentials in Legendre Space. PLoSONE. 9 (3), (2014).
- Terasaki, M., Schmidek, A., Galbraith, J. A., Gallant, P. E., Reese, T. S. Transport of cytoskeletal elements in the squid giant axon. Proc Natl Acad Sci U S A. 92 (25), 11500-11503 (1995).
- Haas, K., Sin, W. C., Javaherian, A., Li, Z., Cline, H. T. Single-Cell Electroporationfor Gene Transfer In Vivo. Neuron. 29 (3), 583-591 (2001).
- Haas, K., Jensen, K., Sin, W. C., Foa, L., Cline, H. T. Targeted electroporation in Xenopus tadpoles in vivo-from single cells to the entire brain. Differentiation. 70 (4-5), 148-154 (2002).
- Paredes, R. M., Etzler, J. C., Watts, L. T., Lechleiter, J. D. Chemical Calcium Indicators. Methods. 46 (3), 143-151 (2008).
- Salome, R., et al. Ultrafast random-access scanning in two-photon microscopy using acousto-optic deflectors. J Neurosci Methods. 154 (1), 161-174 (2006).
- Keller, P. J., Ahrens, M. B., Freeman, J. Light-sheet imaging for systems neuroscience. Nature Methods. 12 (1), 27-29 (2015).
- Hjorth, J. J., et al. Detection of silent cells, synchronization and modulatory activity in developing cellular networks. Dev Neurobiol. , (2015).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved