
Method Article
Feeding and Quantifying Animal-Derived Blood and Artificial Meals in Aedes aegypti Mosquitoes
In This Article
Summary
The goal of this protocol is to deliver animal-derived and artificial blood meals to Aedes aegypti mosquitoes through an artificial membrane feeder and precisely quantify the volume of meal ingested.
Abstract
Females of certain mosquito species can spread diseases while biting vertebrate hosts to obtain protein-rich blood meals required for egg development. In the laboratory, researchers can deliver animal-derived and artificial blood meals to mosquitoes via membrane feeders, which allow for manipulation of meal composition. Here, we present methods for feeding blood and artificial blood meals to Aedes aegypti mosquitoes and quantifying the volume consumed by individual females.
Targeted feeding and quantification of artificial/blood meals have broad uses, including testing the effects of meal components on mosquito behavior and physiology, delivering pharmacological compounds without injection, and infecting mosquitoes with specific pathogens. Adding fluorescein dye to the meal prior to feeding allows for subsequent meal size quantification. The meal volume consumed by mosquitoes can be measured either by weight, if the females are to be used later for behavioral experiments, or by homogenizing individual females in 96-well plates and measuring fluorescence levels using a plate reader as an endpoint assay. Meal size quantification can be used to determine whether changing the meal components alters the meal volume ingested or if meal consumption differs between mosquito strains. Precise meal size quantification is also critical for downstream assays, such as those measuring effects on host attraction or fecundity. The methods presented here can be further adapted to track meal digestion over the course of days or to include multiple distinguishable markers added to different meals (like nectar and blood) to quantify the consumption of each meal by a single mosquito.
These methods allow researchers to singlehandedly perform high-throughput measurements to compare the meal volume consumed by hundreds of individual mosquitoes. These tools will therefore be broadly useful to the community of mosquito researchers for answering diverse biological questions.
Introduction
We present a protocol for feeding modified blood meals to Aedes aegypti mosquitoes using an artificial membrane feeder and precisely measuring the meal volume consumed by each individual mosquito. This protocol can be flexibly adapted to alter the content of the meal or to compare the meal volume consumed by different experimental groups of mosquitoes.
The Ae. aegypti mosquito threatens global health by spreading pathogens that cause diseases including yellow fever, dengue fever, chikungunya, and Zika1,2,3,4,5. Ae. aegypti females are obligate blood-feeders; they must consume vertebrate blood to obtain the necessary protein for egg development, and each clutch of eggs requires a full blood meal from at least one host6,7,8. The female mosquito first bites her host by piercing the skin with her stylet and injecting saliva, which contains compounds that trigger the host’s immune response9. She then feeds by pumping blood through her stylet into her midgut. While consuming a blood meal from an infected host, she may ingest blood-borne pathogens6,8, which then migrate from the mosquito’s midgut to her salivary glands10. Female mosquitoes infected in this manner can spread disease by injecting pathogens along with saliva when biting subsequent hosts11,12. Understanding and quantifying the mechanisms of blood-feeding behavior are crucial steps in controlling the transmission of mosquito-borne diseases.
Many laboratory protocols for mosquito rearing and experimentation use live animals including mice, guinea pigs, or humans as a blood source13,14,15,16. The use of live animals imposes ethical concerns as well as complex requirements for personnel training, animal housing and care, and compliance with Institutional Animal Care and Use Committee (IACUC) policies. It also limits the types of compounds that can be delivered to mosquitoes, which constrains the studies that can be carried out17.
Artificial blood-feeding apparatuses, which typically use a membrane system to simulate host skin, are useful tools for studying blood-feeding behaviors that circumvent the need for the maintenance of live hosts. Whole blood can be purchased from a number of vendors and fed to mosquitoes using heated, water-jacketed artificial membrane feeders or similar devices18,19. In this protocol, we demonstrate the use of small, disposable membrane feeders termed “Glytubes”. This membrane feeder, previously published by Costa-da-Silva et al. (2013)20, can be easily assembled from standard laboratory equipment, making it ideal for delivering blood meals to moderate numbers of mosquitoes and straightforward to scale up for testing larger groups or multiple meal formulations. The Glytube is an inexpensive and efficient alternative to other commercial artificial feeders, which may require larger meal volumes and are more suitable for batch feeding large groups of mosquitoes on a single meal formulation21.
This protocol includes two sections: preparing/delivering artificial meals and quantifying consumption. In the first section, Glytubes are used as an efficient means to deliver manipulated diets. Whole blood may be substituted with an entirely artificial meal to compare the effects of blood substitutes in lieu of a blood meal. A recipe adapted from Kogan (1990)22 is presented here, although multiple artificial meal formulations have been developed23,24. Furthermore, feeding is a less invasive and less laborious method to introduce pharmacological compounds than injection. Due to the low total volume required for each meal (1–2 mL), this protocol provides an attractive delivery method to reduce the amounts of expensive reagents. Ae. aegypti females readily consume protein-free meals of saline solution with adenosine 5′-triphosphate (ATP)25,26, which provides a baseline for measuring the effects of single meal components. For example, Neuropeptide Y-like receptor 7 (NPYLR7) in Ae. aegypti is known to mediate host-seeking suppression after a protein-rich blood meal, and when NPYLR7 agonists are added to a protein-free saline meal, female mosquitoes exhibit host-seeking suppression similar to those that have consumed whole blood7.
In the second section, steps for quantifying the volume of each meal consumed by an individual female mosquito are presented. This assay is fluorescence-based and captures feeding status in higher resolution than methods in which females are classified as “fed” or “unfed” based on visual assessment of abdominal distension alone. By adding fluorescein to the meal prior to feeding, meal volumes ingested by individuals can be quantified by homogenizing each mosquito in a 96-well plate and measuring fluorescence intensity as a readout. This assay can measure differences in feeding vigor in response to variables such as meal composition or the mosquitoes’ genetic background. Precise quantification is critical for intermediate meal sizes, for example when females are offered suboptimal meals containing feeding deterrents or when they consume sucrose meals of variable sizes27. If fed mosquitoes are required for subsequent behavioral assays after meal size quantification, meal size can instead be calculated by weighing anesthetized females in groups and estimating the average increased mass per individual. Although less precise than fluorescein marking, weighing still provides an aggregated estimate of meal volume and allows examination of the meal’s effect on downstream processes, such as fecundity or subsequent host attraction. While blood meal size is variable and can be influenced by a myriad of factors11,28,29, ingested meal sizes measured with the methods described here are consistent with previous quantifications7,30,31.
Protocol
Blood-feeding procedures were not performed using live animals or human hosts and complied with the guidelines set by The Rockefeller University Institutional Animal Care and Use Committee (IACUC) and Institutional Review Board (IRB).
1. Meal preparation
- Preparation of the phagostimulant, adenosine 5′-triphosphate
- Prepare a 25 mM solution of aqueous NaHCO3 (molecular weight, MW = 84.006 g/mol). For 100 mL of 25 mM NaHCO3, add 210 mg of NaHCO3 to a volumetric flask and fill with double-distilled water (ddH2O) to a total volume of 100 mL. Using a magnetic stir bar, thoroughly mix the solution until all the NaHCO3 is dissolved.
- Reconstitute ATP disodium salt hydrate (MW = 551.14 g/mol) in the aqueous 25 mM NaHCO3 to a final concentration of 200 mM ATP. For a total volume of 10 mL of 200 mM ATP in 25 mM NaHCO3 buffer, add 1.1 g of ATP disodium salt hydrate to a volumetric flask and fill with 25 mM NaHCO3 buffer to a total volume of 10 mL. Using a magnetic stir bar, thoroughly mix the solution until all the ATP is dissolved.
NOTE: To minimize hydrolysis of ATP, it must be buffered by a salt solution such as NaHCO3. - Aliquot the ATP solution and store at -20 °C.
NOTE: This stock solution of ATP is typically made fresh every six months and is used for all meals described below. To prevent degradation, ATP aliquots should not undergo multiple freeze-thaw cycles or be heated along with other meal components.
- Preparation of the fluorescent tracer solution, fluorescein
- Prepare a 2% (w/v) stock solution of aqueous fluorescein. For a total stock solution volume of 10 mL, mix 0.2 g of fluorescein disodium salt with 10 mL of ddH2O in a 15 mL conical tube wrapped in aluminum foil at room temperature. This stock solution of fluorescein can be used for dilution in all meals described below.
NOTE: As fluorescein is light-sensitive, avoid exposure to light by wrapping containers in aluminum foil.
- Prepare a 2% (w/v) stock solution of aqueous fluorescein. For a total stock solution volume of 10 mL, mix 0.2 g of fluorescein disodium salt with 10 mL of ddH2O in a 15 mL conical tube wrapped in aluminum foil at room temperature. This stock solution of fluorescein can be used for dilution in all meals described below.
- Preparation of animal-derived blood meals
- Calculate the number of meals needed to feed all mosquitoes; each Glytube holds a 2 mL meal and feeds approximately 25 mosquitoes. Prepare one additional meal to calibrate the standard curve for fluorescence readings. Unless stated otherwise, all steps in this section describe reagent amounts required to prepare one meal with a final volume of 2 mL.
- For animal-derived blood meals, transfer 1.98–2 mL defibrinated sheep blood into a 15 mL conical tube (see step 3.3 for desired volume of blood).
NOTE: Commercially defibrinated sources of vertebrate blood, including from sheep, guinea pigs, and humans, may be used13. Prior to use, ensure that the purchased blood has not passed its expiry date and mix it well by inverting the bottle, especially if there is visible separation of blood components. - For optimal feeding, add ATP to a final concentration of 1–2 mM after the sheep blood has been warmed to 45 °C in a water bath. For a final concentration of 1 mM ATP, add 10 μL of the 200 mM ATP stock solution to 1.99 mL of pre-warmed blood and mix. For a final concentration of 2 mM ATP, add 20 μL of the 200 mM ATP stock to 1.98 mL of pre-warmed blood and mix. If ATP is not to be added, warm 2 mL of defibrinated sheep blood.
- If fluorescence-based quantification of meal size is to be subsequently carried out, add fluorescein solution to a final concentration of 0.002% (2 μL of 2% fluorescein stock in 2 mL total meal volume). Reduce the volume of blood by the same amount as the fluorescein added. Retain 1 mL of the final meal formulation containing 0.002% fluorescein to generate the reference standard curve. Treat the retained volume identically to the meal that is being delivered to mosquitoes; expose to the same light and temperature conditions throughout the duration of the experiment, and subsequently freeze this along with the delivered meal.
- Preparation of artificial blood meals
- Calculate the number of meals needed to feed all mosquitoes; each Glytube holds a 2 mL meal and feeds approximately 25 mosquitoes. Prepare one additional meal to calibrate the standard curve for fluorescence readings. Unless stated otherwise, all steps in this section describe reagent amounts required to prepare one 2 mL meal.
- To prepare artificial blood (adapted from Kogan (1990)22), as in Table 1, first make a stock solution of 400 mM NaHCO3. For a total volume of 10 mL of 400 mM NaHCO3 (MW = 84.006 g/mol), add 336 mg of NaHCO3 to a volumetric flask and fill with double-distilled water (ddH2O) to a total volume of 10 mL. Using a magnetic stir bar, thoroughly mix the solution until all the NaHCO3 is dissolved.
- For the protein components of artificial blood, prepare stock solutions of 50 mg/mL of γ-globulins in 400 mM NaHCO3, 35 mg/mL of hemoglobin in ddH2O, and 300 mg/mL of albumin in ddH2O. Protein stock solutions can be stored at 4 °C for up to 2 months. The final concentration of total human proteins in artificial blood is 125 mg/mL. This includes final concentrations of 15 mg/mL γ-globulins, 8 mg/mL hemoglobin, and 102 mg/mL albumin.
- For each 2 mL meal, combine 600 µL of γ-globulins, 460 µL of hemoglobin, 680 µL of albumin, and 250 µL of ddH2O from stock solutions listed in Table 1. Wait to add 10 µL of 200 mM ATP stock solution until after the meal has been warmed to 45 °C, immediately before presenting the meal.
- If fluorescence-based quantification of meal size is to be subsequently carried out, add fluorescein solution to a final concentration of 0.002% (2 μL of 2% fluorescein stock in 2 mL total meal volume). Reduce the volume of ddH2O in step 4.4 by the same amount as the fluorescein added. Retain at least 1 mL of the final meal formulation containing 0.002% fluorescein to generate the reference standard curve. Treat the retained volume identically to the meal that is being delivered to mosquitoes; expose to the same light and temperature conditions throughout the duration of the experiment, and subsequently freeze this along with the delivered meal.
- Preparation of example protein-free saline meals (adapted from Duvall et al. (2019)7)
NOTE: Protein-free saline meals can be prepared in multiple ways7,27,32. The saline meal presented here is a protein-free version of the artificial blood recipe described above.- Calculate the number of meals needed to feed all mosquitoes; each Glytube holds a 2 mL meal and feeds approximately 25 mosquitoes Prepare one additional meal to calibrate the standard curve for fluorescence measurements. Unless stated otherwise, all steps in this section describe reagent amounts required to prepare one 2 mL meal.
- To prepare the saline meal, make a stock solution of 400 mM NaHCO3. For a total volume of 10 mL of 400 mM NaHCO3 (MW = 84.006 g/mol), add 336 mg of NaHCO3 to a volumetric flask and fill with ddH2O to a total volume of 10 mL. Using a magnetic stir bar, thoroughly mix the solution until all the NaHCO3 is dissolved.
- For each 2 mL meal, combine in a 15 mL conical tube 600 µL of 400 mM NaHCO3 with 1.39 mL of ddH2O. Wait to add 10 µL of the 200 mM ATP stock solution until after the meal has been warmed to 45 °C in a water bath.
- If fluorescence-based quantification of meal size is to be subsequently carried out, add fluorescein solution to a final concentration of 0.002% (2 μL of 2% fluorescein stock in 2 mL of total meal volume). Reduce the volume of ddH2O in step 5.3 by the same amount as the fluorescein added. Retain at least 1 mL of the final meal formulation containing 0.002% fluorescein to generate the reference standard curve. Treat the retained volume identically to the meal that is being delivered to mosquitoes; expose to the same light and temperature conditions throughout the duration of the experiment, and subsequently freeze this along with the delivered meal.
2. Meal delivery to mosquitoes
- Setting up mosquito containers for feeding
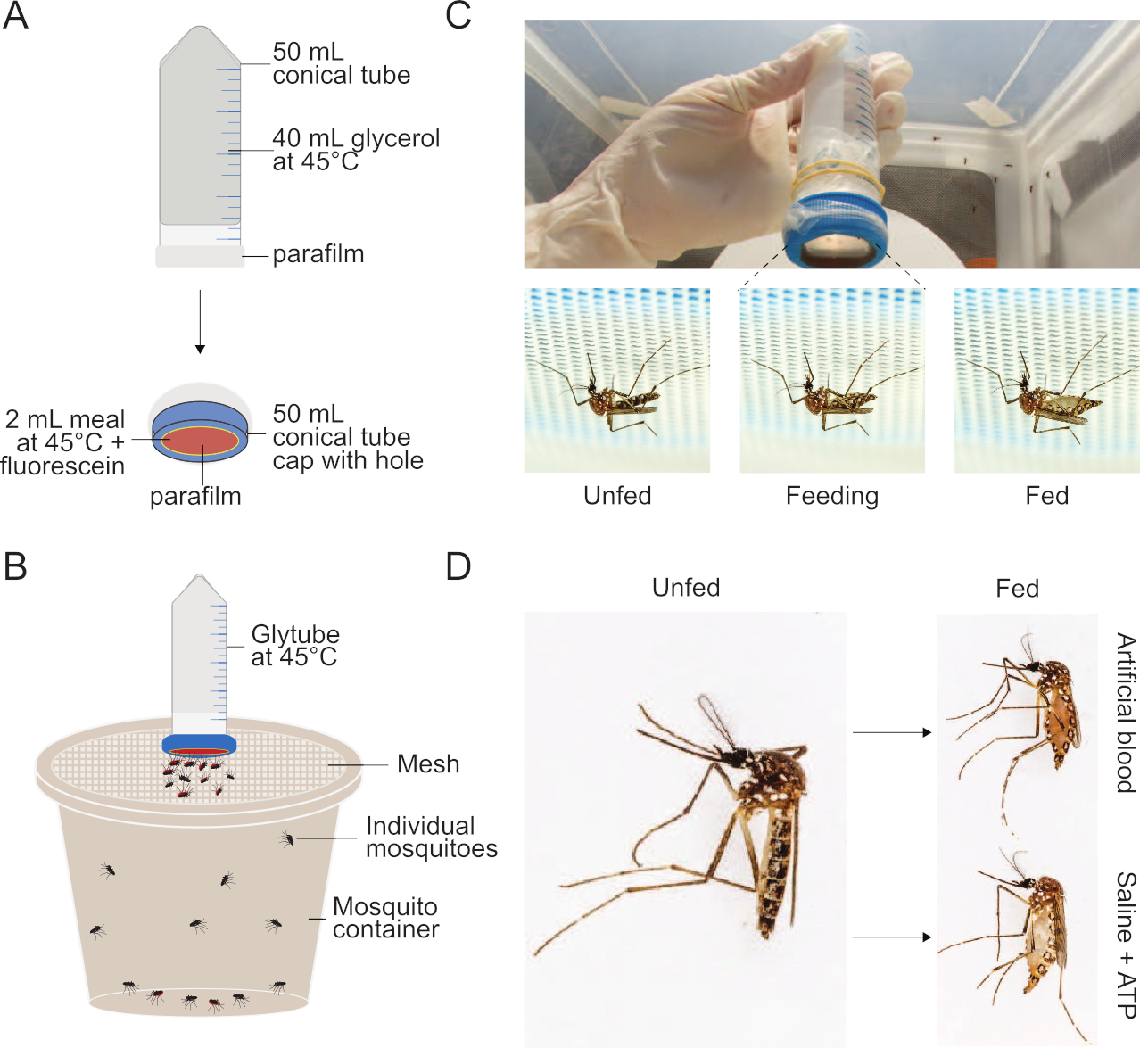
NOTE: Mosquitoes can be fed in a variety of containers as long as the following criteria are met. Ensure that the container is large enough for mosquitoes to fly around in, but not so large that it will be difficult for the mosquitoes to locate the mesh surface and begin feeding. The mesh used to cover the container can vary in material and hole size. The holes must be large enough for the female mosquito’s stylet to pierce through, but not so large that the mosquito can escape. Secure the mesh firmly so that it is taut, and the Glytube can rest stably on its surface throughout the feeding period.- An example container (Figure 1) is a modified 946 mL (32 oz) high density polyethylene (HDPE) plastic bucket. To replicate this setup, use a razor blade to cut a central hole of ~10 cm diameter in the bucket lid. To assemble the container for occupation by mosquitoes, secure a ~400 cm2 square piece of white 0.8 mm polyester mosquito netting on top of the bucket, securely pushing the perforated lid down over it to snap tightly.
- Collect female mosquitoes that are at least 3 days post-eclosion to ensure that they are mature enough to blood feed. Optimal feeding rates are observed after 7 days33.
- Place female mosquitoes into the container and cover with mesh. If the container is densely populated with mosquitoes, increase the number of Glytubes used. Optimal feeding is achieved with ~25 mosquitoes/Glytube. This reduces competition for access to the feeding membrane.
- Set aside a control group of unfed mosquitoes that will not be offered a meal. In the weight measurement protocol, weigh the unfed group separately and use this weight to estimate weight gain in the experimental group that fed on a meal. In the fluorescence-based quantification protocol, add the unfed group of mosquitoes to the wells for the standard curve calculations and for negative controls. To match baseline mosquito tissue autofluorescence in the experimental group, ensure that the standard curve and negative control wells contain an unfed mosquito.
- Constructing and setting up the Glytube (adapted from Costa-da-Silva et al. (2013)20)
- As depicted in Figure 1, to generate a heat source, fill a 50 mL conical tube with 40 mL of 100% glycerol. Seal the open conical tube with a 5 cm × 5 cm piece of parafilm and repeat with an additional piece of 5 cm × 5 cm parafilm to minimize the chance of leakage. Optionally, the parafilm can be held in place using rubber bands. Invert the tube to ensure that there are no holes or gaps.
- To create the meal delivery device, cut a centered hole of 2.5 cm diameter in the screw cap of the conical tube using a sharp razor blade or, for better consistency, a lathe. Stretch a 5 cm × 5 cm piece of parafilm evenly so that it roughly doubles in size. The parafilm should be thin enough that mosquitoes can easily pierce through it, but there should be no leaks. Seal over the outer surface of the screw cap to fully cover the hole and set the cap aside.
NOTE: To increase attraction to the Glytube, prior to stretching the parafilm, perfume it with human odor by gently rubbing it on a patch of human skin with no cosmetics applied, taking care that no holes are created. This is recommended if the experiment is not aimed at investigating the sensory cues required for mosquitoes to approach the meal. - Heat both the sealed tube of glycerol and the meal (with all components except ATP) in a 42–45 °C water bath for at least 15 min. Do not pre-heat ATP; add it immediately before starting the experiment.
- Add ATP to the warmed meal and vortex thoroughly. Pipette 2 mL of the warmed meal into the inner chamber of the screw cap and gently place the inverted, warmed, glycerol-filled 50 mL conical tube in it. Partially screw the cap with the meal onto the glycerol-filled tube—just enough to prevent leakage of the meal or the glycerol.
NOTE: The meal volume used can range between 1 mL and 2.5 mL. Lower volumes may be especially useful when meals are used to deliver compounds that are scarce or expensive. It is important to work quickly at this step so that the meal does not cool down to ambient temperature and reduce the likelihood of maximal feeding. The rate of cooling will depend on the ambient temperature of the room where these steps are conducted, but they should typically be completed within 5 min at 25 °C. - Place the assembled Glytube on top of the mosquito container and allow the mosquitoes access to feed for at least 15 min to achieve maximal feeding rates.
- For optimal feeding, place mosquito containers inside a chamber equipped with a CO2 pad, and allow at least 15 min of acclimation at 25–28 °C and 70–80% humidity prior to delivering the meal. The assay chamber used here is a simple and low cost modification of a previously published setup16. It uses a translucent polypropylene storage box of size 36 cm L × 31 cm W × 32 cm H with a removable lid. A 1.5 cm diameter hole made in the chamber wall allows CO2 delivery through silicone tubing. The CO2 diffusion pad is affixed to the inner center of the lid for delivery of purified air and CO2 to condition the chamber atmosphere during the trial.
NOTE: Make sure host cues (heat and CO2, with optional host odor16) are present so that the mosquitoes are attracted to the membrane feeder. If mosquitoes are not crowding underneath the Glytube, check that CO2 is properly delivered and that the meal and Glytube are sufficiently warm. If an external CO2 source is not available, CO2 can be delivered via puffs of exhaled human breath. - After feeding, the Glytube cap can be discarded as biohazard waste or reused after soaking in a low percentage bleach solution and thoroughly rinsing in water.
3. Quantification of consumed meals
- Weighing mosquitoes to be used for further experiments
NOTE: Weighing mosquitoes to quantify meal size allows them to be used for further live experimentation, but this method requires taking weight measurements from a group of 5 mosquitoes. Since weights of individual mosquitoes are difficult to precisely measure using most laboratory balances, variability in individual meal size cannot be easily quantified by measuring weights. Weighing is only recommended for situations in which females visibly engorge on the meal.- Cold anesthetize mosquitoes by moving their container to a 4 °C cold room or placing it on ice.
- Weigh groups of 5 females from the unfed cohort (i.e., mosquitoes that were never offered a meal) and calculate their average weight as the estimate of the “pre-feeding” weight. The average weight of an unfed mosquito depends on genotype, sex, and rearing conditions. Unfed female Ae. aegypti mosquitoes reared with ad libitum access to sucrose typically weigh approximately 2 mg each.
- From the experimental cohort (i.e., mosquitoes that were offered a meal), sort females into “fed” and “not fed” piles based on the abdominal distension observable by eye7. Divide each of the “fed” and “not fed” piles, respectively, into groups of 5 mosquitoes for weighing. Mosquitoes within each group of 5 should be derived from the same experimental cohort for taking group weight measurements. Calculate the average weight per female from each of the “fed” and “not fed” piles of the experimental group.
- Fluorescence measurement for end-point analysis7,27,34
NOTE: To obtain precise meal size measurements from individual mosquitoes that are no longer required for further live experimentation, store the mosquitoes and the remaining 1 mL of meal containing 0.002% fluorescein at -20 °C immediately after feeding. The experiment can be paused here. This method is outlined in Figure 2.- To generate a reference standard curve, prepare a serial dilution of the same meal containing 0.002% fluorescein that was offered to the experimental group of mosquitoes. There will be a total of 8 standard curve solutions. In each of these solutions, the final volume of meal containing 0.002% fluorescein will be 5, 2.5, 1.25, 0.625, 0.3125, 0.15625, 0.078125, or 0 µL, and each will be in 1x phosphate-buffered saline (PBS) for a total volume of 100 µL (e.g., 5 µL of meal containing 0.002% fluorescein in 95 µL of 1x PBS).
- To make the first solution of the standard curve, add 50 µL of meal containing 0.002% fluorescein to 950 µL of 1x PBS and vortex thoroughly (final volume: 5 µL of meal containing 0.002% fluorescein in 95 µL of 1x PBS). To make the rest of the standard curve solutions, perform a 2-fold dilution for each step by taking 500 µL from the previous tube and adding it to a new tube containing 500 µL of 1x PBS. Vortex well before preparing the next 2-fold dilution.
- To prepare wells to be used for generating a reference standard curve, pipette 100 µL of each of the standard curve solutions into each of the 8 wells in the first column of a 96-well PCR plate. Add 1 unfed control mosquito to each of the same 8 wells in the first column of the plate. Repeat in the second column of the plate for a replicate measurement.
NOTE: If experimental groups are offered different meal types, a separate reference standard curve must be prepared for each meal type. - Add 100 µL of 1x PBS in each remaining well for the unfed control and experimental groups. If tissue is to be disrupted in subsequent steps using a bead mill homogenizer or vortex, add one 3 mm borosilicate solid-glass bead to each well.
- As a negative control, add 1 unfed mosquito to each well in the next 2 columns of the plate. The fluorescence measured in this group sets a baseline cutoff to account for tissue autofluorescence and will be used to determine whether a mosquito in the experimental group fed on the meal.
- Add 1 mosquito per well to the remaining wells from the experimental groups that were offered a meal.
- Seal the plate carefully and disrupt the tissue by manual grinding. The abdomen should be thoroughly homogenized to release the meal. Methods to disrupt tissue include using a bead mill homogenizer with 3 mm borosilicate solid-glass beads (30 Hz for 30 seconds), vortex mixer with 3 mm borosilicate solid-glass beads, or a pestle grinder without beads.
- Centrifuge the plate at 2000 rpm for 1–2 min to collect the lysate.
- Prepare a black 96-well plate with 180 µL of 1x PBS in each well.
- Transfer 20 µL of lysate to each well with 180 µL of 1x PBS and mix. If available, use a multi-channel pipette at this step for increased speed and better consistency.
- Measure fluorescence intensity of each well using a plate reader on the 485/520 excitation/emission channel. Generate the reference standard curve by plotting the known volume of meal against the corresponding fluorescence intensity measurement.
- Using the reference standard curve generated, extrapolate the meal volume ingested by each of the experimental group mosquitoes. Subtract the average fluorescence intensity reading of the negative control group of unfed mosquitoes from the fluorescence intensity reading of each experimental group individual to correct for baseline tissue autofluorescence.
Results
Figure 1 presents a schematic for assembling the Glytube, whereas Figure 2 shows an overview of the experimental design to measure meal size using the fluorescence-based assay described here. Figure 3 provides representative fluorescein meal size measurements from a blood-feeding experiment. Figure 4, Figure 5, and Figure 6 illustrate a sampling of biological questions that can be addressed using this protocol. Applications of the protocol are wide-ranging and include altering blood meal composition, feeding pharmacological compounds, precisely quantifying sub-optimal blood meals or smaller nectar meals, and comparing feeding behavior across mosquito genotypes.
To generate a standard curve for meal volume calculations, fluorescence readings are plotted from the designated reference wells each containing an unfed mosquito and a known volume of the meal with 0.002% fluorescein (Figure 3A). Fluorescence readings from the remaining wells, which contain mosquitoes from either the negative control group of unfed mosquitoes or the experimental group of mosquitoes offered a meal, are compared to this standard curve to quantify the meal volume (µL) consumed by each mosquito (Figure 3B). To validate the baseline readings in this assay, it should be confirmed that mosquitoes from the unfed negative control group are not assigned a positive value of µL consumed (Figure 3B, left). Although all females in the experimental group were offered the blood meal, some mosquitoes fed (Figure 3B, middle) and some did not (Figure 3B, right). This result demonstrates that two types of data can be obtained from this protocol: 1) the percentage of total females that feed on a given meal, and 2) the volume ingested by the females that feed on a given meal.
This protocol can be used to deliver and quantify meals with various protein compositions. Figure 4A,B show data collected using meals with added fluorescein. The proportion of mosquitoes that fed and the meal volume they ingested, respectively, were calculated from the fluorescence readings. These readings are highly sensitive and allow for precise quantification of µL, but have the limitation that mosquitoes cannot be used for future live experiments. Figure 4C,D show data collected from an independent experiment with mosquitoes that were scored as fed or unfed by eye after they were offered meals without fluorescein. Meal size was calculated as average weight/female from groups of 5 mosquitoes. Although these weight measurements are less sensitive than fluorescence measurements, they allow the females to be recovered and used for further live experimentation. The proportion of mosquitoes that feed can vary across different experimental days, as reflected in Figure 4A and Figure 4C.
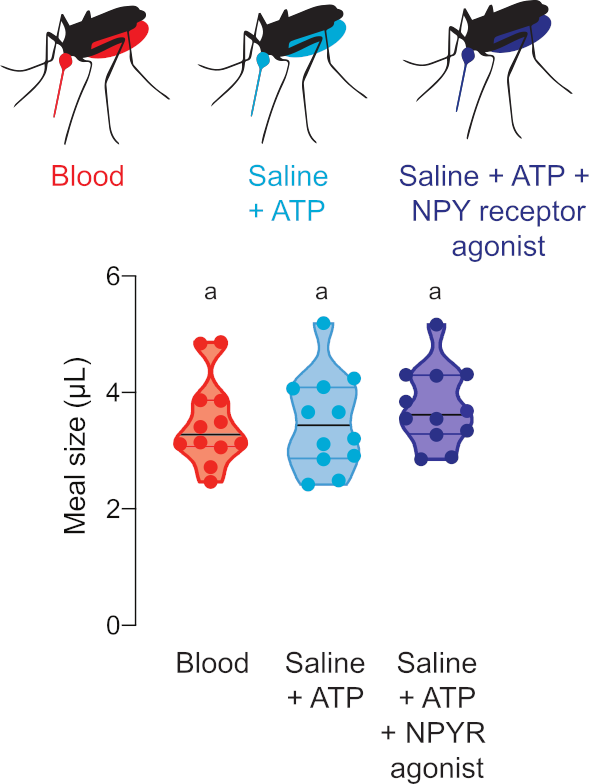
Figure 5 shows the volume consumed of meals containing drugs that regulate mosquito host-seeking behavior. In these experiments, females were offered blood, saline + ATP, or saline + ATP meals with 100 μM of the human NPY Y2 receptor agonist, TM30338. This drug alters host-seeking behavior through activation of Ae. aegypti NPY-like receptor 7. Measuring meal sizes is critical for the interpretation of experiments to assess the effect of this drug on post-blood-feeding behavior because it allows the researcher to calculate the dose consumed by each female.
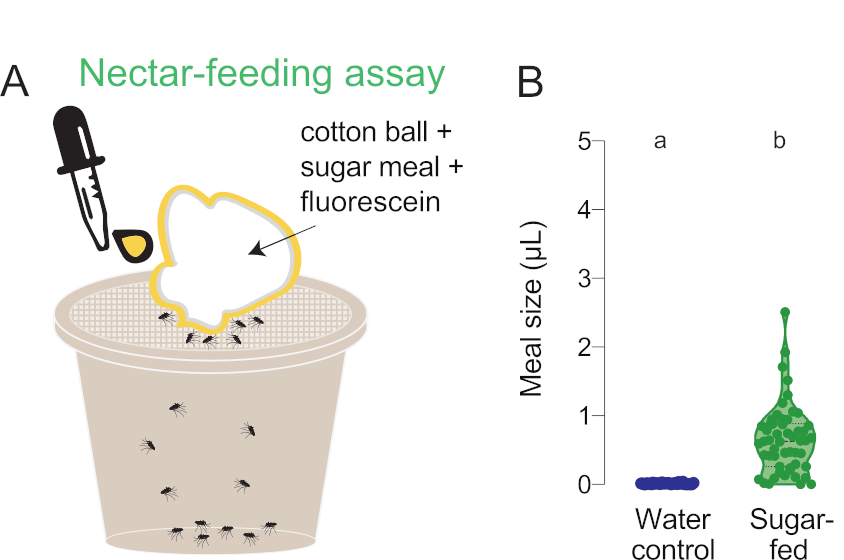
In the previous examples, females were fed either blood or substitute blood meals, all of which resulted in 3–5 µL meals (Figure 3, Figure 4, Figure 5). This fluorescence-based assay can also be used to measure smaller and/or more variable meal sizes that cannot be accurately discerned from average group weight measurements. In Figure 6, the same fluorescence quantification protocol was used to measure nectar-feeding behavior by exchanging the Glytube for a cotton ball saturated with 10% sucrose containing 0.002% fluorescein. Nectar sugars cannot be presented in the Glytube assay because females cannot detect the presence of nectar sugars with the stylet and do not initiate feeding27. These data allow the researcher to determine that sugar meals are consistently smaller than blood meals, in agreement with previous work34 (Figure 6).

Figure 1: Setup of Glytube method used to feed meals to mosquitoes. (A) Schematic of a deconstructed Glytube used to feed blood and other meals to mosquitoes. (B) Schematic of a Glytube presented atop a container of mosquitoes with a mesh lid. Female mosquitoes can pierce through the mesh lid to feed. (C) Photographs of the Glytube (top), and female Aedes aegypti mosquitoes before, during, and after feeding (bottom, from left to right) on a Glytube-delivered meal. Mosquitoes are shown piercing through the mesh covering their container to access the membrane feeder. (D) Photographs showing the appearance of female Ae. aegypti mosquitoes that are unfed (left) and that have engorged on either an artificial blood meal (right, top) or a saline + ATP meal (right, bottom). The Glytube method was previously published in Costa-da-Silva et al. (2013)20. Photographs in (C) and (D) are courtesy of Alex Wild. Please click here to view a larger version of this figure.
{kind=link}

Figure 2: Schematic of how to quantify meal size after Glytube blood-feeding protocol. (A) Mosquitoes are offered a meal with fluorescein (top, experimental group) or no meal (bottom, unfed negative control group). (B) Individual mosquitoes are added to a 96-well plate after terminating the feeding experiment. (C) Standard curve is generated using known amounts of meal containing 0.002% fluorescein. (D) Mosquitoes are homogenized to release any consumed fluorescein, and fluorescence levels in each well are quantified using a plate reader. This fluorescence quantification method is modified from Liesch et al. (2013)34. Please click here to view a larger version of this figure.
{kind=link}

Figure 3: Glytube blood-feeding experiment with fluorescein-based quantification. (A) Standard curve measurements obtained from the wells where a mosquito from the unfed control group was added to a known quantity of meal containing 0.002% fluorescein (y-axis scale = arbitrary units). (B) Meal volume calculated using fluorescence readings for females in the unfed control group (left, black, n = 40), the experimental group that fed on blood (middle, red, n = 37), and the experimental group that did not feed on blood (right, red, n = 23). Each point represents a measurement from an individual female. Data are shown as median with range. Letters indicate statistically distinct groups, Kruskal-Wallis test with Dunn’s multiple comparison, p<0.01. These data were published in Jové et al. (2020)27. Please click here to view a larger version of this figure.
{kind=link}

Figure 4: Quantification of meals with differing protein composition. Females were offered meals of either sheep blood (red), artificial blood with human blood proteins (Kogan (1990)22) (orange), or protein-free saline + ATP meal (aqua)7. (A) Percentage of females fed scored using fluorescence readings. Each point represents a group of 12–16 females. Data are shown as medians with ranges, n = 12. (B) Meal volume calculated using fluorescence readings. Each point represents a measurement from an individual female in a single trial from Figure 4A. Data are shown as medians with ranges, n = 12. (C) Percentage of females fully engorged after artificial membrane feeding, scored by eye. Each point represents the percent of females engorged from groups of 20–30 females. Data are shown as medians with ranges, n = 23. (D) Meal sizes scored as weight/female after feeding status was scored by eye. Weights were calculated as the average of groups of 5 mosquitoes. Data are shown as medians with ranges, n = 23. A–D: Letters indicate statistically distinct groups, Kruskal-Wallis test with Dunn’s multiple comparison, p<0.05. Please click here to view a larger version of this figure.
{kind=link}

Figure 5: Quantification of meals with pharmacological compounds. Females consume meals of the same size of sheep blood (red), saline + ATP (aqua), and saline + ATP + 100 µM dose of human NPY Y2 receptor agonist TM30338 (dark blue). Meal volume calculated using fluorescence readings. Each point represents a measurement from an individual female. Data are shown as medians with ranges, n = 12. Letters indicate statistically distinct groups, Kruskal-Wallis test with Dunn’s multiple comparison, p<0.05. Please click here to view a larger version of this figure.
{kind=link}

Figure 6: Quantification of smaller nectar meals. (A) Schematic of nectar-feeding assay. (B) Meal volume calculated using fluorescence readings for wild-type females offered meals of either water (blue, n = 36) or 10% sucrose (green, n = 53), each with 0.002% fluorescein, in the nectar-feeding assay. Each point represents a measurement from an individual female. Data are shown as medians with ranges. Letters indicate statistically distinct groups, Mann-Whitney test, p<0.05. These data were published in Jové et al. (2020)27. Please click here to view a larger version of this figure.
{kind=link}
| Artificial Blood Meal | |||
| Concentration of Stock Solution (mg/mL) | Volume of Stock Solution in Meal (μL/mL) | Final Meal Concentration (mg/mL) | |
| Protein Components* | |||
| γ-Globulins | 50 | 300 | 15 |
| Hemoglobin | 35 | 230 | 8 |
| Albumin | 300 | 340 | 102 |
| Total Protein | - | - | 125 |
| Non-Protein Components | |||
| Concentration of Stock Solution (mM) | Volume of Stock Solution in Meal (μL/mL) | Final Meal Concentration (mM) | |
| NaCl | In γ-globulin stock | - | 5-10 |
| NaHCO3 | In γ-globulin stock | - | 120 |
| ATP | 200 | 5 | 1 |
| Water | - | 125 | - |
| *Protein components are prepared in stock solution of double-distilled water, except for γ-Globulins, which are dissolved in 400 mM NaHCO3 and include a variable amount of NaCl (2-4%) in the product. | |||
Table 1: Recipe for preparing artificial blood meals (adapted from Kogan (1990)22). Artificial blood consists of protein and non-protein components regularly found in human blood and provides the option to vary the ratios of these components. Mosquitoes can produce eggs after feeding on artificial blood7,22.
| Saline Meal | |||
| Component | Concentration of Stock Solution (mM) | Volume of Stock Solution in Meal (μL/mL) | Final Meal Concentration (mM) |
| NaCl | - | - | - |
| NaHCO3 | 400 | 300 | 120 |
| ATP | 200 | 5 | 1 |
| Water | - | 695 | - |
Table 2: Recipe for saline meal with ATP (adapted from Duvall et al. (2019)7). Protein-free saline meals can be used to deliver compounds of interest to mosquitoes while still mimicking the abdominal distension that occurs after blood-feeding, but without triggering the egg development that occurs when proteins are ingested.
Discussion
For many laboratory applications, artificial membrane feeders offer distinct benefits compared to live hosts by allowing researchers the ability to directly manipulate the contents of the meal. Although multiple methods are available for artificial membrane feeding, the method described here offers advantages in flexibility, cost, and throughput. In comparison to other commercial membrane feeders, the Glytube assay requires a small meal volume, making it an efficient delivery mechanism for costly reagents, including drugs or pathogens, by minimizing the total volume required7,35. As both the protein-free saline and artificial blood meals promote engorgement, compounds or pathogens can be added to either meal as a high-throughput and non-invasive alternative to injections. Additionally, each component of the Glytube can easily be washed, replaced, or scaled up to deliver and quantify multiple meal types without cross-contamination of the feeding apparatus.
To quantify meal volumes consumed by mosquitoes, the fluorescence-based method enables more precise meal size quantification than weighing the mosquitoes before and after feeding. It should be noted that this method is an end-point assay. In contrast, weighing allows the mosquitoes to be kept alive for further experimentation. By using a plate reader, the fluorescence-based method can be easily scaled up for high-throughput quantification of meals consumed by hundreds of individual females.
To achieve high feeding rates, a combination of sufficient host cues must be present to activate female host-seeking behavior and attract females to the feeder. If mosquitoes are not crowding underneath the Glytube, the meal may not be properly warmed, or CO2 delivery may not be sufficient. Addition of human odor to the membrane surface reliably increases attractiveness of the artificial membrane. If mosquitoes are observed underneath the Glytube but fail to feed, the meal composition may be at fault. Females may not feed if the meal itself is not warm, the blood is too old, or if the additives to the meal are intrinsically aversive or cause an undesirable chemical reaction36. Additional ATP also reliably increases feeding rates and can be scaled up to a final concentration of 2 mM in each of the recipes provided. Females may not feed if the parafilm is not pulled taut across the Glytube cap; the parafilm should be uniformly transparent and should not buckle, as this prevents the female from being able to effectively pierce the parafilm with her stylet. If the meal leaks through the Glytube onto the mesh, the parafilm may have torn during the stretching process and should be replaced.
Changing the meal composition can also allow researchers to manipulate the length of time needed to clear the meal from the midgut as well as the subsequent host-seeking behavior. The meals presented here require 24–36 h for digestion7 similar to animal-derived blood. After feeding on any of these meals, females will suppress host-seeking during the digestion time window. Since the saline meal lacks protein, females return to host-seeking after the meal is cleared. If a faster return is desirable, researchers can choose alternate “quick clearing” saline meals that are excreted in approximately 6 h27. While the composition of the saline meal presented here is matched to directly compare results with the artificial blood meal, the “quick clearing” meal more closely matches physiological salt levels found in vertebrate blood.
The methods described here have limitations that should be considered before selecting the assay that is most suited to the researcher’s experimental goals. The fluorescein measurements described do not allow mosquitoes to be used again for additional experimentation. However, weight measurements can be taken prior to meal size quantification using the fluorescein assay. If weight and meal size are consistent across multiple trials for a given meal, weight can be used as a proxy in future experiments. Moreover, this protocol does not distinguish between deficits in host-seeking versus blood-feeding behavior; mosquitoes that show impairments in finding the membrane feeder will have a reduction in feeding rates and/or meal size. By adding a camera to record behavior throughout the assay, researchers can determine whether the females cannot find the Glytube, or whether they find the Glytube, but do not feed.
The assay described here can be adapted to explore many outstanding questions related to feeding behavior in mosquitoes. For example, the contribution of specific blood proteins can be explored by altering the ratio of constituent proteins or total protein concentration in the artificial blood meal. To evaluate meal sizes from multiple feeding events, dyes with distinct fluorescence spectra can be added to differentiate meals from unique sources37. This protocol can also be modified to separately stimulate the internal mouthparts that detect blood and that are used for ingestion (i.e., stylet), and the chemosensory appendages that contact skin (i.e., labium, legs) as the mosquito lands to begin blood feeding36. For example, if ligands are added directly to the meal, they do not contact the labium and legs, since the membrane is pierced only by the stylet. If ligands are added to the outer surface of the parafilm instead, they remain separated from the meal and may be contacted by the labium and legs36. Finally, the detailed kinetics of blood-feeding behavior are not well understood and the method presented here could be modified to combine high-resolution tracking with machine learning tools to extract behavioral readouts of locomotion, posture, and feeding dynamics38.
This protocol is aimed at being user-friendly and cost-effective, with the ability to serve researchers employing pharmacological and genetic manipulations to study mosquito blood-feeding and post-blood-feeding behavior.
Disclosures
The authors have nothing to disclose.
Acknowledgements
We thank Nipun Basrur, Adriana K. Rosas Villegas, Nadav Shai, and Trevor Sorrells for comments on the manuscripts, and Zhongyan Gong and Kyrollos Barsoum for technical assistance. We thank Alex Wild for photographs used in Figure 1. K.V. was supported by the Boehringer Ingelheim Fonds PhD fellowship. V.J. was supported in part by NIH T32-MH095246. This work was supported in part by a grant to The Rockefeller University from the Howard Hughes Medical Institute through the James H. Gilliam Fellowships for Advanced Study program to V.J. This material is based upon work supported by the National Science Foundation Graduate Research Fellowship Program under Grant No. NSF DGE-1325261 to V.J. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Materials
| Name | Company | Catalog Number | Comments |
| 15 mL conical tubes | Fisher Scientific | 14-959-70C | |
| 3 mm diameter borosilicate solid-glass bead | MilliporeSigma | Z143928 | For use for bead mill homogenizer; not required if using pellet pestle grinder |
| 32 oz. high-density polyethylene (HDPE) plastic cup | VWR | 89009-668 | Example mosquito container used for feeding assays shown; alternate options can be used |
| 50 mL conical tubes | Fisher Scientific | 14-959-49A | |
| 96-well black polystyrene plate | ThermoFisher | 12-566-09 | |
| 96-well PCR plate sealing film | Bio-Rad | MSB1001 | Alternate options can be used |
| 96-well PCR plates | Bio-Rad | HSP9621 | Alternate options can be used |
| Adenosine 5′-triphosphate (ATP) disodium salt hydrate | MilliporeSigma | A6419 | |
| Albumin (human serum) | MilliporeSigma | A9511 | |
| Aluminum foil | Fisher Scientific | 01-213 | Alternate options can be used to block light entering fluorescein container |
| Balance | Fisher Scientific | 01-911 | Alternate options can be used |
| Bead mill homogenizer | Qiagen | 85300 | Not required if using pellet pestle grinder |
| Cotton ball | Fisher Scientific | 22456880 | For nectar-feeding; alternate options can be used |
| Defibrinated sheep blood | Hemostat Laboratories | DSB100 | Alternate options can be used |
| Drosophila CO2 fly pad | Tritech Research | MINJ-DROS-FP | Alternate options can be used |
| Fluorescein | MilliporeSigma | F6377 | |
| Fluorescence plate-reader | ThermoFisher | VL0000D0 | Alternate options can be used |
| Gamma-globulin (human blood) | MilliporeSigma | H7379 | |
| Glycerol | MilliporeSigma | G7893 | |
| Hemoglobin (human) | MilliporeSigma | G4386 | |
| Laboratory wrapping film - parafilm | Fisher Scientific | 13-374 | |
| Magnetic stirrer | Fisher Scientific | 90-691 | Alternate magnetic stirrers can be used |
| Microcentrifuge for 96-well plate | VWR | 80094-180 | Alternate options can be used |
| Microcentrifuge Tubes | MilliporeSigma | 2236412 | Alternate options can be used |
| Pellet pestle grinder | VWR | KT749521-1500 | Not required if using bead mill homogenizer |
| Phosphate buffered solution (PBS) | Fisher Scientific | BW17-516F | Optional |
| Razor blades | Fisher Scientific | 12-640 | Alternate options can be used, such as a lathe for better consistency of cutting |
| Rubber bands | |||
| Silicone tubing | McMaster Carr | Needed if using a fly pad for CO2 delivery | |
| Sodium bicarbonate (NaHCO3) | Fisher Scientific | S233 | |
| Sodium chloride (NaCl) | MilliporeSigma | S9888 | |
| Stir bars | Fisher Scientific | 14-512 | Alternate magnetic stir bars can be used |
| Translucent polypropylene storage box with removable lid | Example box used for feeding assays shown | ||
| Vortex mixer | |||
| Water bath | Alternate heating device may be used | ||
| White 0.8 mm polyester mosquito netting | American Home & Habit Inc. | F03A-PONO-MOSQ-M008-WT | Alternate options can be used |
References
- Bhatt, S., et al. The global distribution and burden of dengue. Nature. 496 (7446), 504-507 (2014).
- Rogers, D. J., Wilson, A. J., Hay, S. I., Graham, A. J. The global distribution of yellow fever and dengue. Advances in Parasitology. 62 (05), 181-220 (2006).
- Chouin-Carneiro, T., et al. Differential susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika virus. PLoS Neglected Tropical Diseases. 10 (3), (2016).
- Guerbois, M., et al. Outbreak of Zika virus infection, Chiapas State, Mexico, 2015, and first confirmed transmission by Aedes aegypti mosquitoes in the Amercias. Journal of Infectious Diseases. 214 (9), 1349-1356 (2016).
- Weaver, S. C., et al. Zika virus: history, emergence, biology, and prospects for control. Antiviral Research. 130, 69-80 (2016).
- Attardo, G. M., Hansen, I. A., Raikhel, A. S. Nutritional regulation of vitellogenesis in mosquitoes: implications for anautogeny. Insect Biochemistry and Molecular Biology. 35 (7), 661-675 (2005).
- Duvall, L. B., Ramos-Espiritu, L., Barsoum, K. E., Glickman, J. F., Vosshall, L. B. Small-molecule agonists of Ae. aegypti neuropeptide Y receptor block mosquito biting. Cell. 176 (4), 687-701 (2019).
- Dimond, J. B., Lea, A. O., Hahnert, W. F., DeLong, D. M. The amino acids required for egg production in Aedes aegypti. The Canadian Entomologist. 88 (2), 57-62 (1956).
- Guerrero, D., Cantaert, T., Missé, D. Aedes mosquito salivary components and their effect on the immune response to arboviruses. Frontiers in Cellular and Infection Microbiology. 10, 1-11 (2020).
- Raquin, V., Lambrechts, L. Dengue virus replicates and accumulates in Aedes aegypti salivary glands. Virology. 507, 75-81 (2017).
- Farjana, T., Tuno, N. Multiple blood feeding and host-seeking behavior in Aedes aegypti and Aedes albopictus (diptera: Culicidae). Journal of Medical Entomology. 50 (4), 838-846 (2013).
- Scott, T. W., Takken, W. Feeding strategies of anthropophilic mosquitoes result in increased risk of pathogen transmission. Trends in Parasitology. 28 (3), 114-121 (2012).
- Ross, P. A., Lau, M. J., Hoffmann, A. A. Does membrane feeding compromise the quality of Aedes aegypti mosquitoes. PLoS ONE. 14 (11), 1-19 (2019).
- Ross, P. A., Axford, J. K., Richardson, K. M., Endersby-Harshman, N. M., Hoffmann, A. A. Maintaining Aedes aegypti mosquitoes infected with wolbachia. Journal of Visualized Experiments. 2017 (126), 1-8 (2017).
- Briegel, H., Hefti, M., DiMarco, E. Lipid metabolism during sequential gonotrophic cycles in large and small female Aedes aegypti. Journal of Insect Physiology. 48 (5), 547-554 (2002).
- McMeniman, C. J., Corfas, R. A., Matthews, B. J., Ritchie, S. A. S., Vosshall, L. B. Multimodal integration of carbon dioxide and other sensory cues drives mosquito attraction to humans. Cell. 156 (5), 1060-1071 (2014).
- Pakes, S. P., et al. Guide for the Care and Use of Laboratory Animals. , National Academic Press. Washington, DC, USA. (2011).
- Deng, L., Koou, S. Y., Png, A. B., Ng, L. C., Lam-Phua, S. G. A novel mosquito feeding system for routine blood-feeding of Aedes aegypti and Aedes albopictus. Tropical Biomedicine. 29 (1), 169-174 (2012).
- Gunathilaka, N., Ranathunge, T., Udayanga, L., Abeyewickreme, W. Efficacy of blood sources and artificial blood feeding methods in rearing of Aedes aegypti (Diptera: Culicidae) for sterile insect technique and incompatible insect technique approaches in Sri Lanka. BioMed Research International. 2017, 3196924(2017).
- Costa-da-Silva, A. L., et al. Glytube: a conical tube and parafilm M-based method as a simplified device to artificially blood-feed the Dengue vector mosquito, Aedes aegypti. PLoS ONE. 8 (1), 53816(2013).
- Carvalho, D. O., et al. Mass production of genetically modified Aedes aegypti for field releases in Brazil. Journal of Visualized Experiments. 83 (83), 1-10 (2014).
- Kogan, P. H. H. Substitute blood meal for investigating and maintaining Aedes aegypti (Diptera: Culicidae). Journal of Medical Entomology. 27 (4), 1-4 (1990).
- Gonzales, K. K., Hansen, I. A. Artificial diets for mosquitoes. International Journal of Environmental Research and Public Health. 13 (12), (2016).
- Baughman, T., et al. A highly stable blood meal alternative for rearing Aedes and Anopheles mosquitoes. PLoS Neglected Tropical Diseases. 11 (12), 0006142(2017).
- Galun, R. Feeding stimuli and artificial feeding. Bulletin of the World Health Organization. 36, 590-593 (1967).
- Galun, R. Feeding response in Aedes aegypti: stimulation by adenosine triphosphate. Science. 142, 1674-1675 (1963).
- Jové, V., et al. Sensory Discrimination of Blood and Floral Nectar by Aedes aegypti Mosquitoes. Neuron. 108, 1-18 (2020).
- Petersen, M. T., et al. The impact of the age of first blood meal and Zika virus infection on Aedes aegypti egg production and longevity. PLoS ONE. 13 (7), 1-15 (2018).
- Sissoko, F., et al. Frequent sugar feeding behavior by Aedes aegypti in Bamako, Mali makes them ideal candidates for control with Attractive Toxic Sugar Baits (ATSB). PLoS ONE. 14 (6), 0214170(2019).
- Houseman, J. G., Downe, A. E. R. Methods of measuring blood meal size and proteinase activity for determining the effects of mated state of digestive processes of female Aedes aegypti (L.) (Diperta: Culicidae). The Canadian Entomologist. 18, 241-248 (1986).
- Redington, B. C., Hockmeyer, W. T. A method for estimating blood meal volume in Aedes aegypti using a radioisotope. Journal of Insect Physiology. 22 (7), 961-966 (1976).
- Gonzales, K. K., et al. The effect of SkitoSnack, an artificial blood meal replacement, on Aedes aegypti life history traits and gut microbiota. Scientific Reports. 8 (1), 1-14 (2018).
- Klowden, M. J. The endogenous regulation of mosquito reproductive behavior. Experientia. 46 (7), 660-670 (1990).
- Liesch, J., Bellani, L. L., Vosshall, L. B. Functional and genetic characterization of neuropeptide Y-like receptors in Aedes aegypti. PLoS Neglected Tropical Diseases. 7 (10), 22486(2013).
- Frances, S. P., Sithiprasasna, R., Linthicum, K. J. Laboratory evaluation of the response of Aedes aegypti and Aedes albopictus uninfected and infected with Dengue virus to Deet. Journal of Medical Entomology. 48 (2), (2011).
- Dennis, E. J., Goldman, O. V., Vosshall, L. B. Aedes aegypti mosquitoes use their legs to sense DEET on contact. Current Biology. 29 (9), 1551-1556 (2019).
- Harrington, L. C., et al. Heterogeneous feeding patterns of the Dengue vector, Aedes aegypti, on individual human hosts in rural Thailand. PLoS Neglected Tropical Diseases. 8 (8), 3048(2014).
- Hol, F. J., Lambrechts, L., Prakash, M. BiteOscope, an open platform to study mosquito biting behavior. eLife. 9, 1-24 (2020).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved