
Method Article
Microinjection of Corn Planthopper, Peregrinus maidis, Embryos for CRISPR/Cas9 Genome Editing
In This Article
Summary
Herein are protocols for collecting and microinjecting precellular corn planthopper embryos for the purpose of modifying their genome via CRISPR/Cas9-based genome editing or for the addition of marked transposable elements through germline transformation.
Abstract
The corn planthopper, Peregrinus maidis, is a pest of maize and a vector of several maize viruses. Previously published methods describe the triggering of RNA interference (RNAi) in P. maidis through microinjection of double-stranded RNAs (dsRNAs) into nymphs and adults. Despite the power of RNAi, phenotypes generated via this technique are transient and lack long-term Mendelian inheritance. Therefore, the P. maidis toolbox needs to be expanded to include functional genomic tools that would enable the production of stable mutant strains, opening the door for researchers to bring new control methods to bear on this economically important pest. However, unlike the dsRNAs used for RNAi, the components used in CRISPR/Cas9-based genome editing and germline transformation do not easily cross cell membranes. As a result, plasmid DNAs, RNAs, and/or proteins must be microinjected into embryos before the embryo cellularizes, making the timing of injection a critical factor for success. To that end, an agarose-based egg-lay method was developed to allow embryos to be harvested from P. maidis females at relatively short intervals. Herein are provided detailed protocols for collecting and microinjecting precellular P. maidis embryos with CRISPR components (Cas9 nuclease that has been complexed with guide RNAs), and results of Cas9-based gene knockout of a P. maidis eye-color gene, white, are presented. Although these protocols describe CRISPR/Cas9-genome editing in P. maidis, they can also be used for producing transgenic P. maidis via germline transformation by simply changing the composition of the injection solution.
Introduction
The corn planthopper, Peregrinus maidis, is an economically important pest of maize1,2,3. They cause direct physical damage to the plant, both while feeding with their piercing-sucking mouthparts, and during reproduction when they lay their embryos directly into plant tissue2,4. Despite the multiple routes of direct damage to crops, the greatest impact these insects have on crop health is indirect, by acting as the vector of maize mosaic virus (MMV) and maize stripe virus5,6. MMV is capable of replicating in the body of its P. maidis vector, allowing the virus to persist in individual insects through the entirety of their lives, so they can continue to spread the virus to new host plants7,8. The most common methods for controlling P. maidis, and thus the viruses it vectors, are insecticides.
Unfortunately, mismanagement of these products has caused development of resistance in the target pest as well as pollution of the environment9. Therefore, new strategies are needed to reduce crop losses from this insect/virus-pest combination. Previous work demonstrated that RNA interference (RNAi) could be an effective control method for P. maidis because they are susceptible to downregulation in gene expression even when ingesting double-stranded RNA (dsRNA)10. However, the most effective way to administer dsRNA in the field would be through the plants the insects feed on; hence, crops could still be susceptible to any viruses the insects are already carrying. With the advent of CRISPR/Cas9 genome editing, new pest control strategies are possible, including Cas9-based gene drive11,12, which could be used to reduce the size of a pest population, or to replace said population with individuals resistant to the viruses they vector.
However, development and deployment of any type of gene-drive system will require the development of transgenic techniques. Such methods were not necessary for carrying out RNAi experiments in P. maidis because dsRNAs and/or siRNAs are presumed to be able to cross cell membranes due to the efficiency of RNAi in P. maidis10,13. This is not true for the DNAs and/or proteins employed in traditional transgenesis or in Cas9-based gene editing, either of which would be a precursor to creating insects carrying a gene drive. To accomplish gene editing or other forms of germline transformation, these DNAs and proteins are ideally microinjected into embryos during the syncytial blastoderm stage, prior to when the insect embryo cellularizes. Timing is critical, because the syncytial stage is the earliest part of development14,15. As P. maidis females preferentially lay their eggs in plant tissue, extracting sufficient quantities of precellular embryos for microinjections can be labor-intensive and time-consuming. Therefore, new techniques were developed to quickly collect and microinject P. maidis embryos prior to cellularization.
Protocol
1. Colony-level rearing of P. maidis adults
- Plant a minimum of four pots of corn per week per rearing cage, with 3-4 seeds per pot. Grow in an insect-free environment.
- When plants are ~5 weeks old, place inside a 30 cm x 30 cm x 60 cm cage.
- Obtain a sufficient quantity of P. maidis adults (~500) from a research laboratory or the wild, and place into an insect-proof cage with 9-12 corn plants (3-4 pots).
- Maintain the colony in an insect-rearing incubator at 25 °C (± 1 °C), with at least 70% humidity and a 14:10 light cycle.
- To generate an age-calibrated colony, remove all initial adults after four days of egg laying, and allow the embryos laid in the cage to hatch and age naturally.
- Move 5-week-old P. maidis insects (adults) to fresh corn plants for weekly subculture by collecting with an aspirator (Figure 1). Then, release the adults into a clean cage with fresh corn plants. To maintain a steady supply of young adults for experimental purposes, prepare fresh age-calibrated cages every week.
- Water the pots in the cages twice daily. Periodically clip stalks, remove decaying plant material, and replace with fresh corn pots as needed.
NOTE: With proper maintenance, a colony can last ~5 weeks (i.e., long enough for embryos laid in the cage to become adults).
2. Agarose-based egg-laying chamber
- Make egg collection dishes (oviposition medium) by pouring 1% w/v agarose in water into clean 100 mm x 15 mm Petri dishes. Store the oviposition medium at 4 °C after it solidifies.
- Prepare 10% w/v sucrose solution for feeding the adults. Store the sucrose solution at -20 °C for up to a month.
- Make a chamber to hold the adults by cutting a hole in the bottom of a 1 oz cup (see the Table of Materials) and gluing a screen over the hole for air exchange (Figure 2).
- Cut plastic paraffin wax film into 5 cm x 5 cm squares; set aside 2 squares for each cup.
- Collect ~15 1-week-old adult females from an age-calibrated P. maidis colony. To select females, examine the ventral side of the abdomen, and look for the ovipositor, which is typically darker than the rest of the abdomen (Figure 3). Hold adults for up to one hour in a 15 mL conical vial if setting up multiple egg-laying chambers. Chill the insects briefly on ice prior to sexing and transfer to the adult container.
NOTE: This examination can be done without a microscope. Adult females that have had time to feed and mate also typically have larger abdomens than adult males and are more docile; hence, they can more easily be selected from a cage population. - Transfer the females into an adult container, and seal the cup with 1 layer of plastic paraffin wax film by evenly stretching it 3-4 times its original size (Figure 4A,B).
- Apply 400 µL of 10% w/v sucrose solution to the top of the plastic paraffin wax film seal, and add a second layer of plastic paraffin wax film, stretching the plastic paraffin wax film exactly as above (Figure 4C,D).
NOTE: The sandwich of stretched plastic paraffin wax film pressurizes the sucrose solution, which is very important for adult feeding, but will not prevent females from piercing their ovipositors all the way through into the oviposition medium. - Place the adult chamber on an egg collection dish with the plastic paraffin wax film side directly on the oviposition medium, and wrap the entire egg-laying chamber with plastic wrap without covering the air holes, as these are required for air exchange (Figure 5).
- Incubate each egg-laying chamber at 25 °C with 70% humidity and a 14:10 light cycle.
- Change the sandwich of plastic paraffin wax film and 10% w/v sucrose solution daily, and remove any water that accumulates inside the cup.
3. Embryo collection and alignment in a high-humidity environment
- Set up a stereomicroscope-based microinjection system in a humidified space or hood (humidified hood; Figure 6) to ensure the working environment achieves at least 70% humidity throughout the microinjection process.
- Check the oviposition medium for eggs after the desired egg-lay period. Do this in a humidified hood or another humid environment.
NOTE: The egg-lay period typically used was overnight, from 6 PM to 10 AM, lasting ~16 h. - If any eggs are laid in the agarose, use fine forceps to carefully dig them out, and place them on the surface of the agarose to keep them moist (Figure 7A).
- Apply a strip of 1 mm x 15 mm double-sided tape on a 22 mm x 30 mm coverslip (Figure 7B). Place the coverslip tape-side up on the oviposition medium (Figure 7C).
- Pick up each individual egg from the agar surface, and move to the double-sided tape using a fine brush. Remove any eggs that are completely white or have black coloration on them. Healthy eggs will be the semi-transparent.
- Place the banana-shaped eggs on their side, with the larger end stuck on the double-sided tape (Figure 7D).
NOTE: Always keep the eggs in a high-humidity environment, such as a Petri dish cast with a layer of 1% agar on the bottom.
4. Preparation of CRISPR reagents and injection needles
- Pull quartz needles using a Flaming/Brown type micropipette puller.
- Bevel the quartz needles using a micropipette beveler.
- Use double-sided sticky tape to secure pulled needles in a clear container, such as a Petri dish, until ready to use.
- Prepare the injection solution by combining 0.5 µL of Cas9 protein (5 µg/µL stock solution) and 0.5 µL of sgRNA (4 µg/µL stock solution; see the Table of Materials) with 1 µL of phenol red buffer in a final volume of 5 µL. To precipitate particles that could clog the needle, vortex the solution briefly, and centrifuge for 3 min at maximum speed.
- Backfill the injection needle, taking care to leave the injection mix near the tapered end of the needle. Remove bubbles, if any, from the tip of the needle.
- Carefully place the backfilled needle into the needle holder, and tighten the stainless steel collar to hold the needle securely in place during microinjection.
- Generate a reliable flow of injection solution from the needle by gently stroking the beveled tip with a fine, dampened paintbrush, while delivering bursts of air pressure to the needle with the injection system.
NOTE: The needle is ready for injection when the injection mix can leave the tip in small amounts.
5. Microinjection and post-injection care
- Prepare a microinjection platform by filling a clean 100 mm x 15 mm Petri dish with 1% agar to form a level layer of agar that is flush with the top of the dish.
- Place a previously prepared coverslip with ~25 embryos upon the agar platform (Figure 8A).
NOTE: All injection steps must be performed inside a humidified hood (~70% humidity). - Check the injection pressure by placing the needle tip in a drop of water and initiating the injection cycle.
NOTE: A small amount of injection solution should disperse into the water if the pressure setting is correct (Figure 8B). - Insert the needle into the larger end of the embryo, approaching from the left side of the coverslip (Figure 8C). Deliver the injection solution into the egg, and pull out the needle quickly.
- After all eggs are injected, place the coverslip on the surface of a new 1% agar dish, and transfer the dish to a humidity chamber (Figure 9).
6. Incubating and hatching of embryos
- Put the hatching chamber in a 25 °C incubator for 6 days.
- Transfer any surviving embryos, using clean water and a fine brush, to a 35 mm x 10 mm Petri dish with water-moistened filter paper covering the bottom of the dish. Seal the Petri dish with plastic paraffin wax film, and hold at 25 °C to allow the embryos to hatch. Start checking the embryos 6 days post-injection for survival.
NOTE: First instar nymphs will start hatching around day 8. - Transfer nymphs, using a fine brush, to a Petri dish containing leaf clippings. Cover the dish, and seal with plastic paraffin wax film.
- Incubate the sealed dish of hatchlings on leaf cuttings for 48 h at 25 °C.
- Transfer all 2-day-old nymphs from a round of injections to a rearing cage with corn plants using a fine brush. If injectees with visible phenotype are recovered in sufficient numbers, rear them separately to maximize the recovery of the target trait in the next generation. Otherwise, perform mass-mating of all the injectees.
NOTE: Place hatchlings gently in the whorl of the corn plant to provide refuge and ensure proper humidity of their immediate environment. - Rear the insects in the conditions described above, ensuring proper temperature, humidity, and regular transfers to fresh corn plants.
- Screen progeny for expected phenotypes. Place individuals exhibiting the desired phenotype into their own cage to establish homozygous lines.
Results
The egg-lay chamber was specifically designed to enable P. maidis females to feed while ovipositing in a protective medium from which their eggs could easily be recovered. Using this method, sufficient quantities of precellular embryos were recovered for microinjection with DNA, RNA, and/or proteins. Adult P. maidis females usually lay eggs inside the leaf tissue of the corn plants, which makes getting enough eggs in a short amount of time a challenge because it requires a lot of leaf dissection. The artificial egg-laying environment provides a solution to overcome these problems. As shown in Table 1, 6,483 eggs were collected from a total of 645 females in 4 weeks. Females usually start laying eggs after day 2 and provide most eggs from day 4 to day 6. Oviposition activity slowed down by day 9. Each oviposition chamber was set up on Friday and checked for eggs from Sunday until the next Sunday. Following this schedule allowed most eggs to be collected for microinjections during the work week.
The first practical application of this egg-laying system was to test the efficacy of Cas9-mediated gene knockout, using the P. maidis ortholog of the eye-color gene, white (Pmw), as a target. Mutations in white are known to result in substantial loss of eye pigmentation in other insect species, and White is cell-autonomous, allowing mutations to be detected in injected individuals16,17. To increase the chance that even a small mutation might result in loss of function, guide RNAs were designed to cut within the region of the ATP-binding cassette, which is necessary for White function16. P. maidis embryos were injected with either 20% phenol red (injection buffer), injection buffer with Cas9 at a final concentration of 800 ng/µL (Cas9 control), or Cas9 in injection buffer along with three guide RNAs added at a concentration of 400 ng/µL each. The combination of three guides within one injection mix was intended to further maximize the chances of generating mutants, both by creating a large deletion, and by compensating for the possibility that any one guide might be ineffective for cutting. The developmental rates for each treatment were comparable (Table 2), with 50-60% of injected individuals showing signs of development. Hatch rates for the buffer and Cas9 controls were also comparable; however, the hatch rates of individuals receiving the three-guide mix was relatively lower. At this time, it is unclear if the reduced survival is the result of the loss of white function or the result of unintended consequences of the three-guide mix, such as off-target effects (see the discussion section). However, none of the individuals with complete loss of eye pigmentation (i.e., complete knockout) hatched, and none of the progeny of injected individuals had white eyes. The on-target efficacy of Cas9-based mutagenesis was verified two ways. First, injectees were screened for loss of eye pigmentation.
Of the 71 guide-injected individuals that developed, 23 showed some degree of pigment loss (Figure 10), and 9 of those individuals hatched, resulting in a knockout rate of ≥32%. No eye pigment loss was observed in either control treatment. Second, chromosomal mutations were confirmed via polymerase chain reaction (PCR)18 and sequencing19. Because a mutant line could not be recovered, genomic DNA was analyzed from pools of embryos injected with either the three-guide mix or buffer. The three-guide mix is expected to remove ~180 base pairs from the white locus. This can be seen in the PCR products amplified off of genomic DNA isolated from injected individuals, as well as the associated sequence data generated from those products (Figure 11). This combined evidence indicates that embryos were injected before cellularization occurred.

Figure 1: Aspirator. An effective aspirator can be assembled from attaching a vacuum pump at the intake, via plastic tubing, to a 15 mL plastic conical tube. Approximately 0.5 cm should be carefully removed from the bottom of the conical tube. A cotton ball should be placed in the conical tube, over the opening of the plastic tubing, to catch P. maidis adults as they are collected and keep them out of the vacuum pump. Please click here to view a larger version of this figure.
{kind=link}

Figure 2: Construction of adult containers. (A) The supplies needed (clockwise from top left): screen, hot glue gun, razor blade, 1 oz container. (B) A large hole should be cut in the bottom of the 1 oz container, and a square of screen is cut just large enough to cover this hole. (C) The screen is then glued over the hole using hot glue. (D) Once the glue is set, any excess mesh should be removed. Please click here to view a larger version of this figure.
{kind=link}

Figure 3: Sexing P. maidis adults. The ventral sides of male (left) and female (right) P. maidis adults are shown. The ovipositor, visible over the female abdomen, is the clearest indicator of the sex of an individual. Please click here to view a larger version of this figure.
{kind=link}

Figure 4: Sealing adults into containers. (A) A 5 cm x 5 cm square of plastic paraffin wax film. (B) The film should be stretched evenly to 3-4 times its original size. (C) Once adults have been put into the adult container, the stretched film should be placed over the opening to secure the adults. A 400 µL drop of 10% w/v sucrose solution should then be placed on top of the film. (D) To provide adequate feeding pressure for the adults, a second 5 cm x 5 cm square of plastic paraffin film should be similarly stretched and placed over the drop of sucrose. Please click here to view a larger version of this figure.
{kind=link}

Figure 5: Setting up an oviposition chamber. (A) The supplies needed (clockwise from top left): plastic wrap, a completed adult container (with adults), and a Petri dish with 1% agarose (oviposition medium). (B) The adult container should be placed onto the agarose with the plastic paraffin film/10% sucrose 'sandwich' placed directly on the oviposition medium. (C) Plastic wrap is used to secure the adult container to the oviposition medium. This keeps the medium from drying out too quickly. (D) Care should be taken to avoid covering the screen of the adult container, so that air exchange may still continue. (E) Diagram of the oviposition chamber. Please click here to view a larger version of this figure.
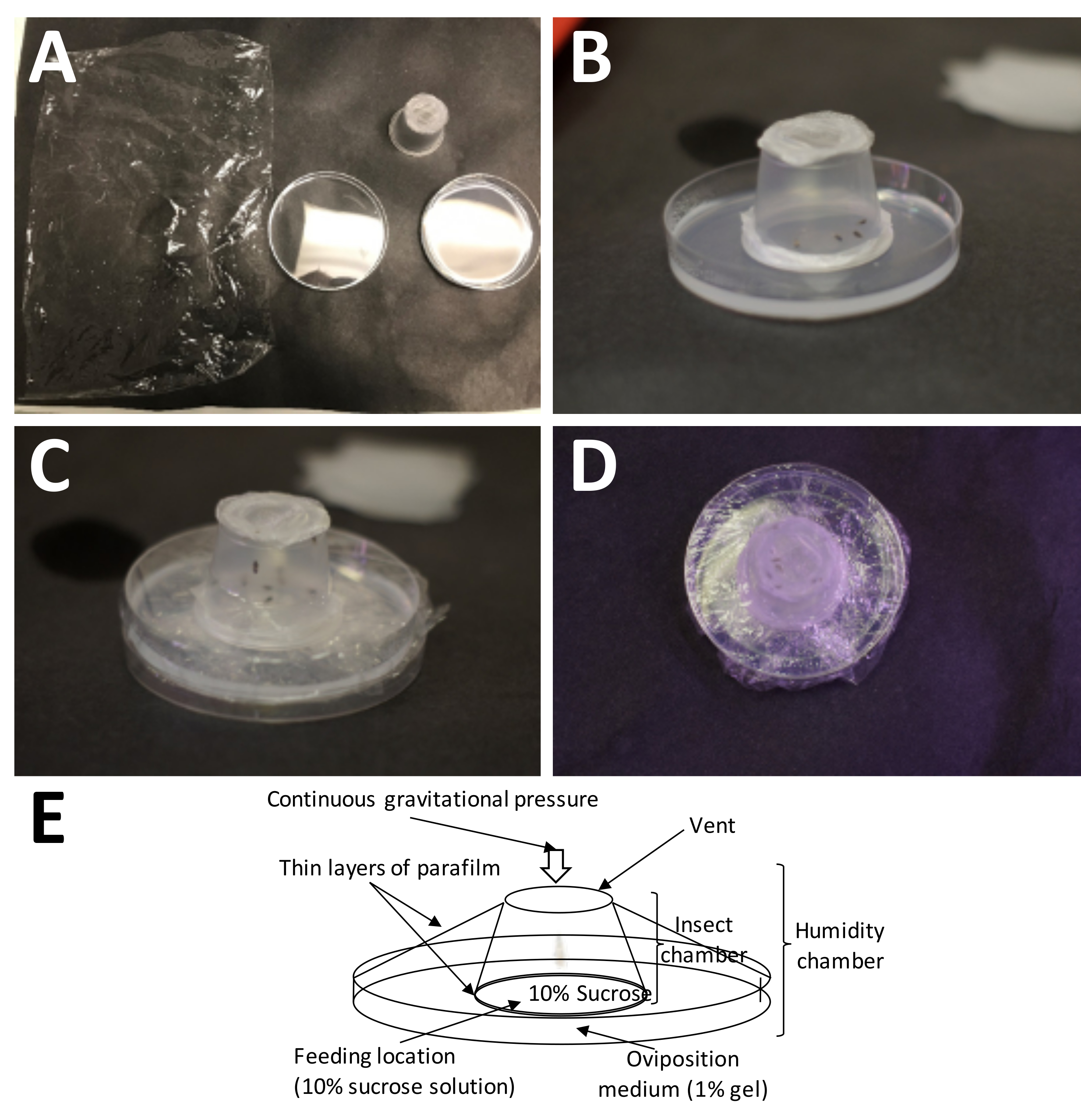{kind=link}

Figure 6: Humidified hood. A hood outfitted with a humidifier has been set up around the injection scope to minimize air drafts and maintain humidity while the embryos are being handled. Flaps can be folded over the entrance after the worker is in place, to aid in maintaining proper humidity levels. Please click here to view a larger version of this figure.
{kind=link}

Figure 7: Collecting embryos in preparation for injections. (A) Embryos that have been deposited in the oviposition medium. A pair of fine forceps are used to extract embryos from the medium and place them on its surface. (B) A narrow strip of 1 mm x 15 mm double-sided tape on a 22 mm x 30 mm coverslip. (C) The coverslip can be placed on the oviposition medium for ease of transferring embryos from the surface of the medium to the tape on the coverslip. (D) P. maidis embryos are banana shaped, with one end narrower than the other (narrow end indicated with red arrow head; wider end indicated with yellow arrow head in example embryo). The wide end of the embryo should be placed on the tape. Please click here to view a larger version of this figure.
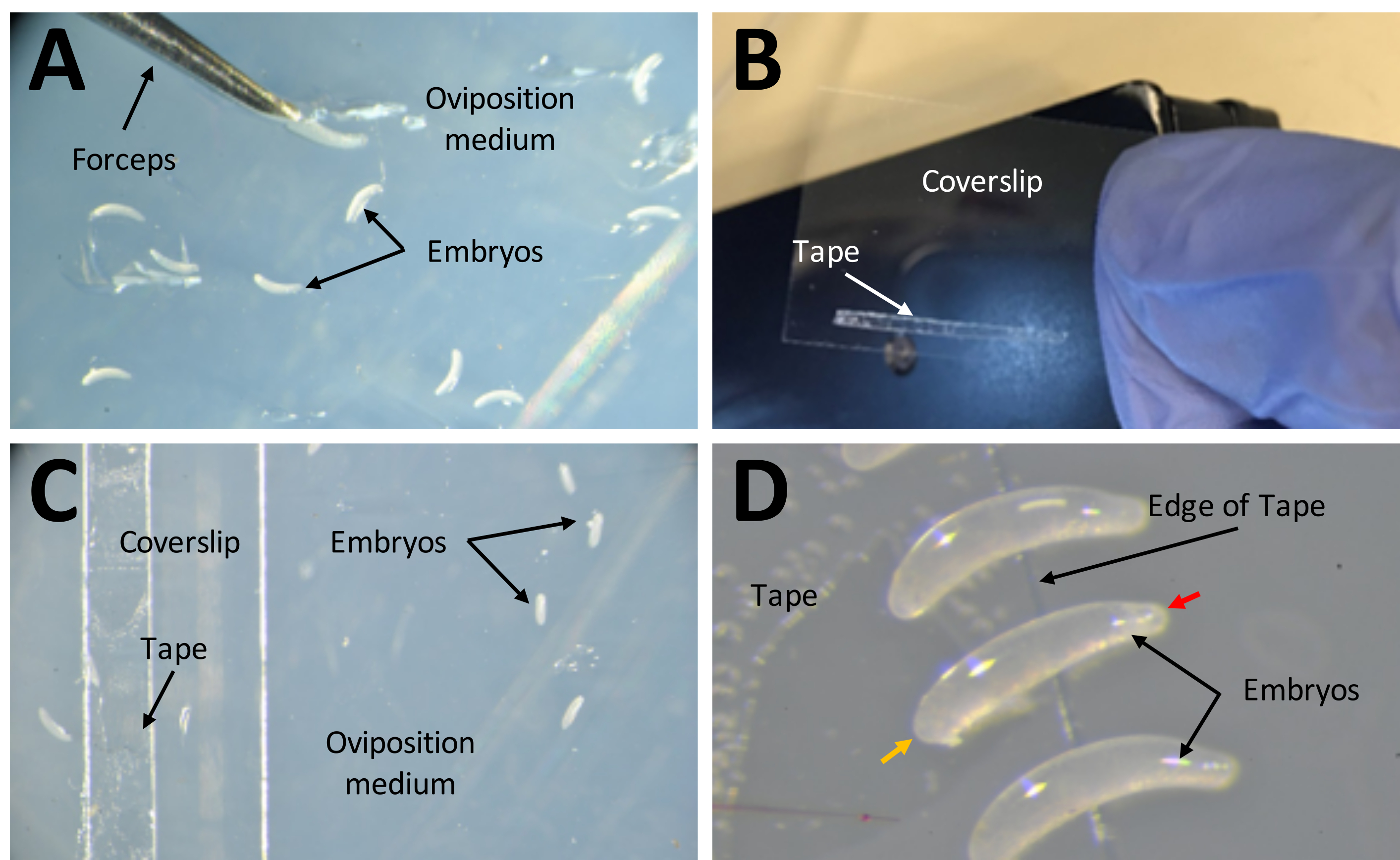{kind=link}

Figure 8: Injection. (A) The injection platform is a Petri dish filled to the brim with 1% agar. The coverslip with a strip of tape holding embryos should be placed directly on the surface of the injection platform. (B) The injection pressure should be tested before embryos are injected by 'injecting' a small amount of injection solution into a drop of water. This method can also be used at any time during the injection process to check the needle for clogs. (C) Embryos should be injected by inserting the needle into the larger end of the embryo. Injection solution should be visible if the injection was successful. Please click here to view a larger version of this figure.
{kind=link}

Figure 9: Post-injection care. (A) Once all the embryos on a coverslip have been injected, the coverslip should be placed in a fresh Petri dish containing 1% agarose. (B) The Petri dish with the coverslip can then be maintained in a humidity chamber (like the one shown) until embryos hatch. Please click here to view a larger version of this figure.
{kind=link}

Figure 10: Pmw knockout phenotype. (A) Age-matched control and (B) Pmw knockout embryos, with developing eyes indicated by black arrowheads. The embryo in B is mosaic, as a small stripe of pigmentation can be seen. (C) Age-matched control and (D) Pmw knockout hatchlings, with insets showing a different angle on the eyes. The hatchling in D is also mosaic. A white arrow points to an area in the main picture showing loss of pigmentation. Please click here to view a larger version of this figure.
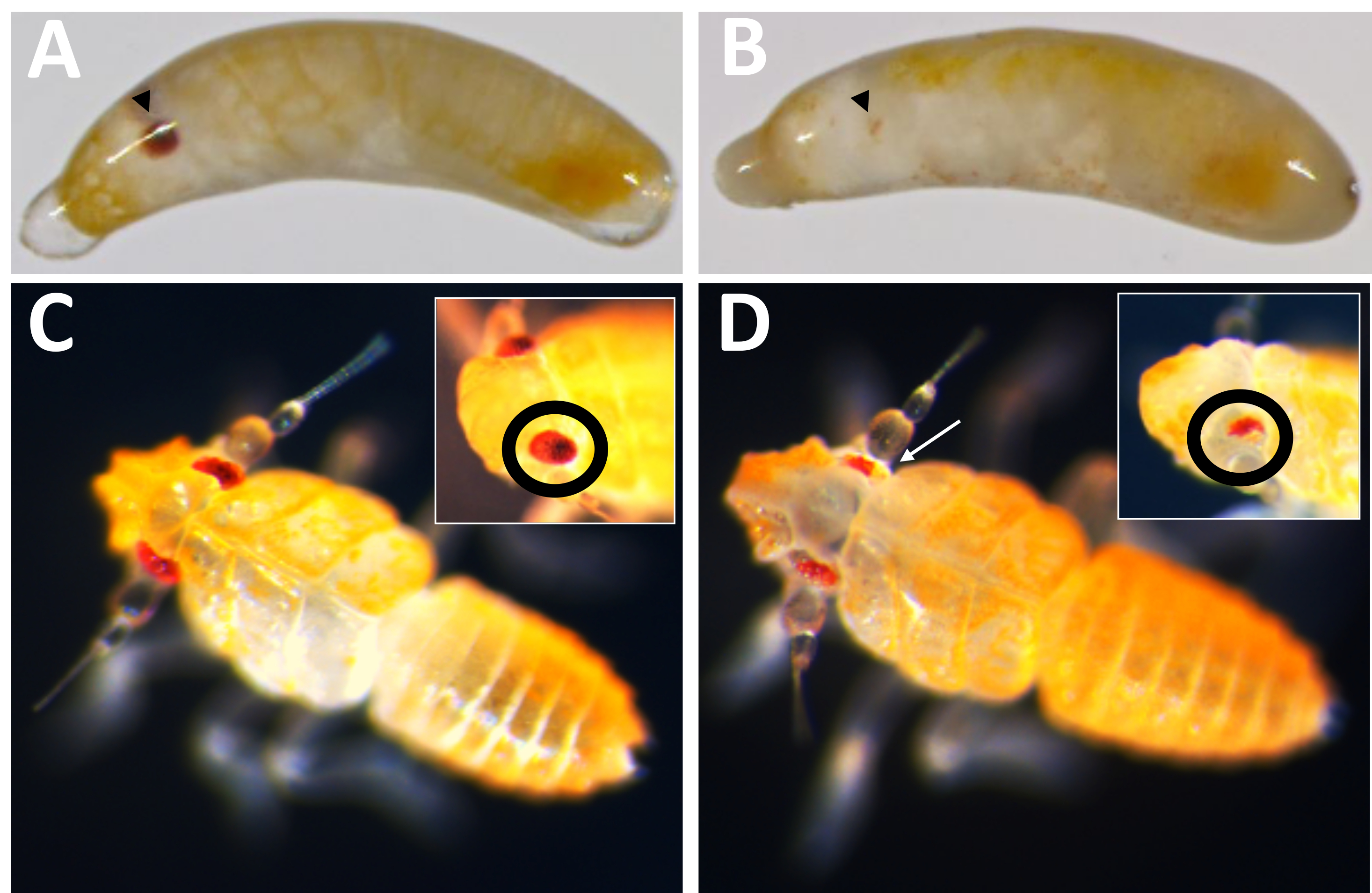{kind=link}

Figure 11: Pmw knockout sequence. (A) To-scale model of Pmw mRNA, marked in 500-bp increments, with locations of gRNA binding sites indicated: G1, blue; G2, yellow; G3, pink. Any frame-shift mutations generated at this point will disrupt the majority of the translation product. (B) Genomic context of gRNA sites, all in one exon (bold capitalized text). Guide binding sites are highlighted in the same colors as A, and PAMs are underlined. Span is ~300 bp. The in-frame translation of the exon is shown above, as single-letter abbreviations in capital text. Two motifs specific to eye pigment transporters are marked. The CDEPT motif of the Walker B functional domain is boxed in purple, and the IHQP motif of the H-loop domain is boxed in green. Both domains are critical to ATP-transporter function. (C) The Pmw target region was amplified using two rounds of PCR. The second-round product was examined on a gel for evidence of size-shift due to successful removal of the region between the guides. Lanes: L = 100 bp ladder; 1 = PCR water control; 2 = Buffer injected eggs; 3-4 = two separate sets of eggs injected with three-guide mix. Only embryos receiving the three-guide mix produced both the WT band (red arrow) and the band resulting from a complete excision (white arrow). (D) To confirm the identity of the lower (white arrow) band, this DNA was purified, cloned, and sequenced. The top line is the wild-type sequence. The other two lines are sequences from two clones. Three additional clones matched the bottom sequence. Blue highlight indicates the binding site of Guide 1, while pink highlight indicates the binding site of Guide 3. In both alleles, the entire region between these two guide sites has been deleted. Abbreviations: Pmw = Peregrinus maidis white gene; gRNA = guide RNA; PAM = protospacer adjacent motif; ATP = adenosine triphosphate; PCR = polymerase chain reaction; WT = wild-type; KO = knockout. Please click here to view a larger version of this figure.
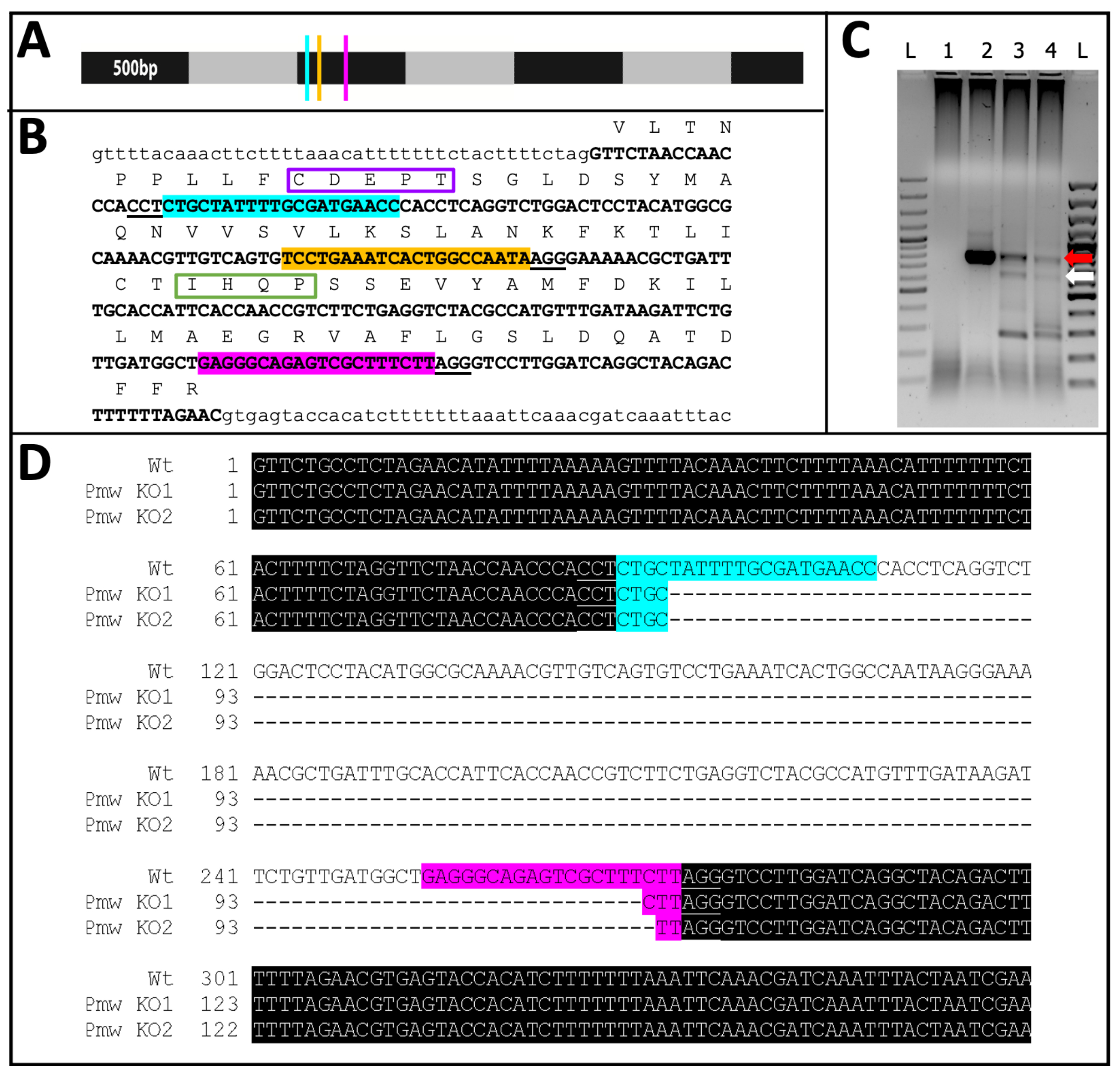{kind=link}
| Set | # of cups | # of females in each cup | # of eggs | Total # of eggs | |||||||
| Day 2 | Day 3 | Day 4 | Day 5 | Day 6 | Day 7 | Day 8 | Day 9 | ||||
| 1 | 10 | 15 | 0 | 26 | 166 | 355 | 530 | 193 | 91 | 37 | 1398 |
| 2 | 15 | 15 | 22 | 238 | 489 | 699 | 520 | 379 | 203 | 58 | 2608 |
| 3 | 8 | 15 | 0 | 57 | 230 | 190 | 116 | 80 | 34 | 1 | 708 |
| 4 | 10 | 15 | 0 | 226 | 446 | 519 | 301 | 179 | 24 | 15 | 1710 |
| Total | 43 | 15 | 23 | 547 | 1331 | 1763 | 1467 | 831 | 352 | 111 | 6483 |
Table 1: Representative egg collections from artificial oviposition environment. Data from four sets of egg-collection cups are shown, with egg tallies starting on the second day after setup and running through the ninth day.
| Injection Treatment | Total Injected | Total Developed | Total Hatched | Development Rate (%) | Hatch Rate (%) |
| Buffer | 39 | 20 | 12 | 51 | 31 |
| Cas9 | 39 | 24 | 14 | 61 | 36 |
| Cas9 +Pmw gRNAs | 121 | 71 | 28 | 59 | 28 |
Table2: Survival and knockout rates from injections of 3 different injection mixes.
Discussion
Egg-lay quality and nutrition
Recently, researchers working with a related species, Nilaparvata lugens, obtained the eggs they used for microinjections directly from the leaf, keeping the injected eggs in the leaf tissue until they hatched17. While this leaf-based method provided a more natural environment for embryonic development, it also increased the chances for infections and for egg damage during the removal process. The artificial oviposition system presented here provides a more uniform environment and reduces the chances of damage to the eggs from handling. By setting up the oviposition cups on Friday, the majority of the oviposited eggs were collected during a typical work week, to the benefit of those doing the microinjection work. One caveat to this method, however, is that the lack of nutrients in the 10% sucrose solution diet will eventually affect the health of the insects, and females in the cups usually start to die off after only 10 days. Egg quality also begins to drop off after 6 days, as evidenced by an increase in dead or unhealthy looking eggs. As a result, it is important to be selective of the eggs used for microinjections and to not keep the females after day 6.
Survival rate and humidity
Two factors seem to be critical for embryonic survival through the microinjection process. The most challenging aspect of handling P. maidis embryos is keeping them from desiccating after removal from the oviposition medium and throughout microinjection. As the eggs are typically laid inside plant tissue, they lack an adequate shell to prevent dehydration. Even in the humidified hood whole sets of eggs were lost due to desiccation. However, excessively high humidity could also affect the microinjections if water accumulated on the double-sided tape or on the scope. Unfortunately, egg dehydration was usually not easy to notice during the microinjection process, and they frequently appeared normal until 2 or 3 days later, when they turned completely transparent, showing no signs of development.
Needle quality also appears to play an important role in survival. The needle should be beveled to minimize unnecessary damage to the egg. When the needle is blocked, using the clearing function on the injector while gently stroking the tip of the needle with a dampened paintbrush (see step 4.7) typically returned the needle to a functional state. Regardless, putting only tiny amounts of injection solution (~0.25 µL) into each needle and switching to a new needle every few slides (~50-60 eggs) is recommended to ensure needle quality is maintained throughout the injection process.
Successful generation of a knockout phenotype
To successfully transform the germ cells, embryo microinjections usually have to be done as early as possible before cellularization. Depending on the insect species, the time window for completing the microinjections ranges from only a couple of hours to as long as a full day14,15,20. It is still unclear when P. maidis embryos undergo cellularization. Cas9-mediated knockout was tested on embryos as young as 4 h post-egg-lay (pel) to as many as 16 h pel, and the expected phenotypes were observed in all experiments, suggesting that all microinjections were performed within the precellurization window.
The P. maidis ortholog of the eye-color gene, white, was selected because the knockout phenotype was expected to be easy to screen in injectees due to its cell-autonomous nature. Indeed, as expected, both mosaic and total knockouts were clearly identifiable among embryos receiving the injection mixture containing Cas9 and guide RNAs. Unfortunately, no injectees with complete knockout hatched, and a mass mating of surviving injectees failed to generate white-eyed progeny. However, a mutant line was later successfully generated by targeting a different gene (Klobasa et al., in progress). This would suggest that the failure to establish a white mutant line is most likely due to either off-target effects (i.e., Cas9 cutting important regions elsewhere in the genome) generating a closely-linked lethal mutation, or to an unpredicted critical role for white in P. maidis.
Phenotypic and molecular data (Figure 8 and Figure 9) affirm that a significant knockout in the white locus was created in a sample of injected embryos, which would result in total loss of gene function. Moreover, while mutations in white are viable in some species, there is precedent for reduced white activity being detrimental21,22. That said, off-target effects cannot be completely ruled out. Predicting likely off-targets requires accurate genome sequence data23, which the current state of genomic resources in P. maidis makes impossible to do at this time. Regardless, with these new methods, testing other target genes can be done confidently, even moving towards more traditional transgenesis in an effort to bring new genetic tools to this pernicious pest.
Disclosures
The authors have nothing to disclose.
Acknowledgements
North Carolina State University, Department of Entomology and Plant Pathology, is part of a team supporting DARPA's Insect Allies Program. The views, opinions, and/or findings expressed are those of the authors and should not be interpreted as representing the official views or policies of the Department of Defense or the U.S. Government. The authors declare no competing interests. MDL, DR, and AEW conceived of the project and provided funding acquisition, project administration, and resources. FC, WK, NG and MDL conceived and designed the microinjection experiments; OH conceived and designed the egg-lay method. FC and WK performed the experiments; FC and WK analyzed the results; and FC, WK, NG, and MDL wrote the manuscript. The authors would like to offer special thanks to Kyle Sozanski and Victoria Barnett for their help in maintaining P. maidis colonies.
Materials
| Name | Company | Catalog Number | Comments |
| 1 oz Containers | Dart | P100N | Adult container for egg-laying setup |
| 15 mL Conical Tubes | Olympus | Genesee 28-103 | Serves as collection tube on vacuum aspirator setup |
| 15 mL Conical Tubes | Olympus | Genesee 28-106 | For making 10% sucorose solution and for holding adults when chilling before screening |
| Aspirator | Bioquip | 1135A | For handling planthoppers |
| Vacuum Aspirator | Fischer Technical | LAV-3 | Vacuum for aspirating larger numbers of insects |
| Blue Spectrum LED Lights | Home Depot | GLP24FS/19W/LED | Grow lights for potted corn plants hoppers are feeding on |
| Cas9 | TrueCut Cas9 Protein v2 | A36498 | Endonuclease for cutting planthopper genes |
| Clear Vinyl Tubing | Home Depot | 3/8 in. I.D. x 1/2 in. O.D. x 10 ft. | Connects collection tube to pump on vacuum aspirator setup |
| Corn planthoppers | North Carolina State University | N/A | Request from Dr. Anna Whitfield's lab |
| Cotton balls | Genessee | 51-101 | Serves as a filter/insect catcher in collection tube on vacuum aspirator setup |
| Double sided tape | Scotch Double Sided Tape | NA | Holding eggs for microinjection |
| Early Sunglow corn | Park Seed Company | 05093-PK-N | Corn for rearing planthoppers |
| epTIPS Microloader Tips | Eppendorf | C2554691 | Backfilling needle loading tips |
| Femtojet Microinjection System | Eppendorf | 5247 | Controls injection pressure (12-20 psi, depending on needle bore size) |
| Nutri-Fly Drosophila Agar | Genessee | 66-103 | Substrate for everything except egg-laying dish |
| Fine forceps | Bioquip | 4731 | Egg handling |
| General Purpose LE Agarose | Apex | 20-102 | Substrate inn egg-laying dish (oviposition medium) |
| Guide RNA 1 - GGUUCAUCGCAAAAUAGCAG | Synthego | CRISPRevolution sgRNA EZ Kit (1.5 nmol) | RNA guides for targeting planthopper white gene |
| Guide RNA 2 - UCUGAAAUCACUGGCCAAUA | Synthego | CRISPRevolution sgRNA EZ Kit (1.5 nmol) | RNA guides for targeting planthopper white gene |
| Guide RNA 3 - GAGGGCAGAGUCGCUUUCUU | Synthego | CRISPRevolution sgRNA EZ Kit (1.5 nmol) | RNA guides for targeting planthopper white gene |
| Humidifyer | Homedics | UHE-CM45 | For providing humidity in humidified hood |
| Humidity chamber | Billups-Rothenberg | MIC-101 | For holding injected embryos until hatching |
| Insect rearing cages | Bioquip (special order) | Close to 1450 L (has plastic front and mesh fabric sides) | Cage for planthoppers on corn |
| Laser-based Micropipiette Puller | Sutter Instruments | P-2000/G | For making injection needles / Heat = 700, FIL = 4, VEL = 40, DEL = 170, PUL = 160 |
| Leica M165 FC Fluorescence Stereomicroscope | Leica | M165 FC | Planthopper screening |
| Microinjection Scope | Leica | MZ12-5 | Microinjection scope outfited with an XY stage |
| Micromanipulator | Narishige | MN-151 | For positioning microinjection needle |
| Micropipette beveler | Sutter Instruments | FG-BV10-D | For beveling injection needles / Used 'fine' graded plate at 20° angle |
| Microscope Stage | AmScope | GT100 X-Y Gliding Table | For positioning and moving embryos under microscope |
| Miniature Paint Brush | Testor #2 8733 | Sold in 3 pack 281206 | Fine paintbrushes for embryo handling |
| Needle Holder | Narishige | HI-7 | For holding the microinjection needle |
| Percival Incubator | Percival | I41VLH3C8 | Rearing injectees until hatch |
| Petri Dishes (100 x 15 mm) | VWR | 89038-968 | Making agar dish for egg-lay |
| pGEM-T Easy Vector System I cloning kit | Promega | A1360 | Cloning Pm white target site |
| Phenol Red | Sigma | 143-78-8 | Microinjection buffer |
| Plain Microscope Slides or coverslip | Fisher Scientific | 12-549-3 | Hold eggs for microinjection |
| Plasmid DNA Midi Kit | Zymo | D4200 | Purification of injection-ready plasmid DNAs |
| Plastic paraffin film | Pechiney Plastic Packaging | PM-996 | Roll size 4 in. x 125 ft |
| Plastic wrap | Glad ClingWrap Plastic Wrap | NA | Wrap the entire egg-laying chamber |
| Primer - PmW CRISPR check F1 - AAGGAATTTCTGGAGGTGAAA | IDT | 25 nmole DNA Oligo | First-round Primer for amplifing across target site within the Pm white gene |
| Primer - PmW CRISPR check R1 - GATTCCTCGCTGTTGGGT | IDT | 25 nmole DNA Oligo | First-round Primer for amplifing across target site within the Pm white gene |
| Primer - PmW CRISPR check F3 - TCACAGACCCTGGTGCTAATC | IDT | 25 nmole DNA Oligo | Second-round Primer for amplifing across target site within the Pm white gene |
| Primer - PmW CRISPR check R3 - GTCCACAATCCACACTTCTGA | IDT | 25 nmole DNA Oligo | Second-round Primer for amplifing across target site within the Pm white gene |
| Quartz capillaries | Sutter Instruments | QF100-50-10 | For making microinjection needles / O.D. 1 mm, I.D. 0.7 mm, 10 cm length |
| Screen (White Organza Fabric) | Joann Fabrics | 16023889 | For covering the adult container |
| Sparkleen | Fisher Scientific | 04-320-4 | Wash dishes |
| Sucrose | Fisher Scientific | BP220-1 | To make 10% sucorose solution |
References
- Namba, R., Higa, S. Y. Host plant studies of the corn planthopper, Peregrinus maidis (Ashmead) in Hawaii. Proceedings of the Hawaiian Entomological Society. 21, 105-108 (1971).
- Singh, B. U., Seetharama, N. Host plant interactions of the corn planthopper, Peregrinus maidis Ashm.(Homoptera: Delphacidae) in maize and sorghum agroecosystems. Arthropod-Plant Interactions. 2 (3), 163-196 (2008).
- Tsai, J. Occurrence of a corn disease in Florida transmitted by Peregrinus maidis. Plant Disease Reporter. 59 (10), 830-833 (1975).
- Chelliah, S., Basheer, M. Biological studies of Peregrinus maidis (Ashmead) (Araeopidae: Homoptera) on sorghum. Indian Journal of Entomology. 27, 466-471 (1965).
- Lastra, J., Esparza, J. Multiplication of vesicular stomatitis virus in the leafhopper Peregrinus maidis (Ashm.), a vector of a plant rhabdovirus. Journal of General Virology. 32 (1), 139-142 (1976).
- Nault, L. R., Ammar, E. -. D. Leafhopper and planthopper transmission of plant viruses. Annual Review of Entomology. 34 (1), 503-529 (1989).
- Ammar, E. -. D., Tsai, C. -. W., Whitfield, A. E., Redinbaugh, M. G., Hogenhout, S. A. Cellular and molecular aspects of rhabdovirus interactions with insect and plant hosts. Annual Review of Entomology. 54, 447-468 (2009).
- Barandoc-Alviar, K., Ramirez, G. M., Rotenberg, D., Whitfield, A. E. Analysis of acquisition and titer of Maize mosaic rhabdovirus in its vector, Peregrinus maidis (Hemiptera: Delphacidae). Journal of Insect Science. 16 (1), 14 (2016).
- Tsai, J. H., Steinberg, B., Falk, B. W. Effectiveness and residual effects of seven insecticides on Dalbulus maidis (Homoptera: Cicadellidae) and Peregrinus maidis (Homoptera: Delphacidae). Journal of Entomological Science. 25 (1), 106-111 (1990).
- Yao, J., Rotenberg, D., Afsharifar, A., Barandoc-Alviar, K., Whitfield, A. E. Development of RNAi methods for Peregrinus maidis, the corn planthopper. PloS One. 8 (8), 70243 (2013).
- Esvelt, K. M., Smidler, A. L., Catteruccia, F., Church, G. M. Emerging technology: concerning RNA-guided gene drives for the alteration of wild populations. Elife. 3, 03401 (2014).
- Gantz, V. M., Bier, E. The mutagenic chain reaction: a method for converting heterozygous to homozygous mutations. Science. 348 (6233), 442-444 (2015).
- Yao, J., Rotenberg, D., Whitfield, A. E. Delivery of maize mosaic virus to planthopper vectors by microinjection increases infection efficiency and facilitates functional genomics experiments in the vector. Journal of Virological Methods. 270, 153-162 (2019).
- Kimelman, D., Martin, B. L. Anterior-posterior patterning in early development: three strategies. Wiley Interdisciplinary Reviews: Developmental Biology. 1 (2), 253-266 (2012).
- Mito, T., Nakamura, T., Noji, S. Evolution of insect development: to the hemimetabolous paradigm. Current Opinion in Genetics & Development. 20 (4), 355-361 (2010).
- Grubbs, N., Haas, S., Beeman, R. W., Lorenzen, M. D. The ABCs of eye color in Tribolium castaneum: orthologs of the Drosophila white, scarlet, and brown Genes. Genetics. 199 (3), 749-759 (2015).
- Xue, W. H., et al. CRISPR/Cas9-mediated knockout of two eye pigmentation genes in the brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Insect Biochemistry and Molecular Biology. 93, 19-26 (2018).
- Lorenz, T. C. Polymerase chain reaction: basic protocol plus troubleshooting and optimization strategies. Journal of Visualized Experiments: JoVE. (63), e3998 (2012).
- Sanger, F., Coulson, A. R. A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. Journal of Molecular Biology. 94 (3), 441-448 (1975).
- Chu, F. C., Wu, P. S., Pinzi, S., Grubbs, N., Lorenzen, M. D. Microinjection of Western Corn Rootworm, Diabrotica virgifera virgifera, embryos for germline transformation, or CRISPR/Cas9 genome editing. Journal of Visualized Experiments: JoVE. (134), e57497 (2018).
- Brent, C. S., Hull, J. J. RNA interference-mediated knockdown of eye coloration genes in the western tarnished plant bug (Lygus hesperus Knight). Archives of Insect Biochemistry and Physiology. 100 (2), 21527 (2019).
- Khan, S. A., Reichelt, M., Heckel, D. G. Functional analysis of the ABCs of eye color in Helicoverpa armigera with CRISPR/Cas9-induced mutations. Scientific Reports. 7 (1), 1-14 (2017).
- Manghwar, H., et al. CRISPR/Cas systems in genome editing: methodologies and tools for sgRNA design, off-target evaluation, and strategies to mitigate off-target effects. Advanced Science. 7 (6), 1902312 (2020).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved