
Method Article
Микроинъекция кукурузного сеялки, Peregrinus maidis, эмбрионов для редактирования генома CRISPR/Cas9
В этой статье
Резюме
В настоящем документе представлены протоколы для сбора и микроинъекции преклеточных эмбрионов кукурузных кузнечиков с целью модификации их генома с помощью редактирования генома на основе CRISPR/Cas9 или для добавления маркированных переносимых элементов посредством трансформации зародышевой линии.
Аннотация
Кукурузный сеялка, Peregrinus maidis, является вредителем кукурузы и переносчиком нескольких вирусов кукурузы. Ранее опубликованные методы описывают запуск РНК-интерференции (РНКi) у P. maidis путем микроинъекции двухцепочечных РНК (дцРНК) нимфам и взрослым особям. Несмотря на силу РНКи, фенотипы, полученные с помощью этого метода, являются преходящими и не имеют долгосрочного менделевского наследования. Таким образом, набор инструментов P. maidis необходимо расширить, включив в него функциональные геномные инструменты, которые позволили бы производить стабильные мутантные штаммы, открывая двери для исследователей, чтобы применить новые методы борьбы с этим экономически важным вредителем. Однако, в отличие от дцРНК, используемых для РНКи, компоненты, используемые при редактировании генома на основе CRISPR/Cas9 и трансформации зародышевой линии, не могут легко пересекать клеточные мембраны. В результате плазмидные ДНК, РНК и/или белки должны быть микроинъецированы в эмбрионы до того, как эмбрион станет клеточным, что делает время инъекции критическим фактором успеха. С этой целью был разработан метод откладывания яиц на основе агарозы, позволяющий собирать эмбрионы у самок P. maidis через относительно короткие промежутки времени. Приведены подробные протоколы сбора и микроинъекции прецеллюлярных эмбрионов P. maidis с компонентами CRISPR (нуклеаза Cas9, комплексообразующая с направляющими РНК), а также представлены результаты нокаута гена P. maidis на основе Cas9 гена цвета глаз P. maidis, белого цвета. Хотя эти протоколы описывают редактирование генома CRISPR/Cas9 у P. maidis, они также могут быть использованы для получения трансгенных P. maidis путем трансформации зародышевой линии путем простого изменения состава раствора для инъекций.
Введение
Кукурузный сеялка, Peregrinus maidis, является экономически важным вредителем кукурузы 1,2,3. Они наносят прямой физический вред растению, как во время кормления своим колюще-сосущим ротовым аппаратом, так и во время размножения, когда откладывают свои зародыши непосредственно в растительную ткань 2,4. Несмотря на многочисленные пути прямого повреждения сельскохозяйственных культур, наибольшее влияние эти насекомые на здоровье сельскохозяйственных культур оказывают косвенное, поскольку выступают в качестве переносчиков вируса мозаики кукурузы (MMV) и вируса полосатости кукурузы 5,6. MMV способен размножаться в организме своего переносчика P. maidis, позволяя вирусу сохраняться в отдельных насекомых на протяжении всей их жизни, поэтому они могут продолжать распространять вирус на новые растения-хозяева 7,8. Наиболее распространенными методами борьбы с P. maidis и, следовательно, вирусами, которые он переносит, являются инсектициды.
К сожалению, неправильное обращение с этими продуктами привело к развитию резистентности у целевого вредителя, а также к загрязнению окружающей среды9. Поэтому необходимы новые стратегии для сокращения потерь урожая в результате этой комбинации насекомых/вирусов-вредителей. Предыдущая работа показала, что РНК-интерференция (РНКи) может быть эффективным методом контроля для P. maidis, поскольку они чувствительны к подавлению экспрессии генов даже при приеме двухцепочечной РНК (дцРНК)10. Однако наиболее эффективным способом введения дцРНК в полевых условиях будет введение через растения, которыми питаются насекомые; Следовательно, сельскохозяйственные культуры все еще могут быть восприимчивы к любым вирусам, которые уже переносят насекомые. С появлением редактирования генома CRISPR/Cas9 возможны новые стратегии борьбы с вредителями, в том числе генный драйв11,12 на основе Cas9, который может быть использован для уменьшения размера популяции вредителей или для замены указанной популяции особями, устойчивыми к вирусам, переносчиками которых они являются.
Однако разработка и внедрение любого типа системы генного драйва потребует разработки трансгенных методов. Такие методы не были необходимы для проведения экспериментов с РНК-интерференциями у P. maidis, поскольку предполагается, что дцРНК и/или миРНК способны пересекать клеточные мембраны из-за эффективности РНКи у P. maidis10,13. Это не относится к ДНК и/или белкам, используемым в традиционном трансгенезе или в редактировании генов на основе Cas9, любой из которых был бы предшественником создания насекомых, несущих генный драйв. Для выполнения редактирования генов или других форм трансформации зародышевой линии эти ДНК и белки в идеале микроинъекционно вводят эмбрионам во время стадии синцитиальной бластодермы, до того, как эмбрион насекомого процветет. Время имеет решающее значение, потому что синцитиальная стадия является самой ранней частью развития14,15. Поскольку самки P. maidis преимущественно откладывают яйца в растительную ткань, извлечение достаточного количества преклеточных эмбрионов для микроинъекций может быть трудоемким и трудоемким. Поэтому были разработаны новые методы для быстрого сбора и микроинъекции эмбрионов P. maidis до клеточности.
протокол
1. Выращивание взрослых особей P. maidis на уровне колоний
- Сажайте минимум четыре горшка кукурузы в неделю на клетку для выращивания, по 3-4 семени в горшке. Выращивайте в среде, свободной от насекомых.
- Когда растениям исполнится ~ 5 недель, поместите их в клетку размером 30 см x 30 см x 60 см.
- Приобретите достаточное количество взрослых особей P. maidis (~ 500) в исследовательской лаборатории или в дикой природе и поместите в защищенную от насекомых клетку с 9-12 растениями кукурузы (3-4 горшка).
- Поддерживайте колонию в инкубаторе для выращивания насекомых при температуре 25 ° C (± 1 ° C), влажности не менее 70% и световом цикле 14:10.
- Чтобы создать колонию, откалиброванную по возрасту, удалите всех первоначальных взрослых особей после четырех дней откладывания яиц и позвольте эмбрионам, отложенным в клетке, вылупиться и состариться естественным образом.
- Переместите 5-недельных насекомых P. maidis (взрослых особей) на свежие растения кукурузы для еженедельного субкультурирования, собирая с помощью аспиратора (рис. 1). Затем отпустите взрослых особей в чистую клетку со свежими растениями кукурузы. Чтобы поддерживать постоянный запас молодых особей для экспериментальных целей, каждую неделю готовьте свежие клетки, откалиброванные по возрасту.
- Поливайте горшки в клетках два раза в день. Периодически обрезайте стебли, удаляйте гниющий растительный материал и при необходимости заменяйте их свежими горшочками с кукурузой.
ПРИМЕЧАНИЕ: При надлежащем содержании колония может просуществовать ~ 5 недель (т.е. достаточно долго, чтобы эмбрионы, отложенные в клетке, стали взрослыми).
2. Камера яйцекладки на основе агарозы
- Приготовьте чашки для сбора яиц (среда для яйцекладки), насыпав 1% мас. арозы в воде в чистые чашки Петри размером 100 мм x 15 мм. Храните среду для откладки яиц при температуре 4 °C после ее затвердевания.
- Приготовьте 10% раствор сахарозы для кормления взрослых особей. Храните раствор сахарозы при температуре -20 °C до месяца.
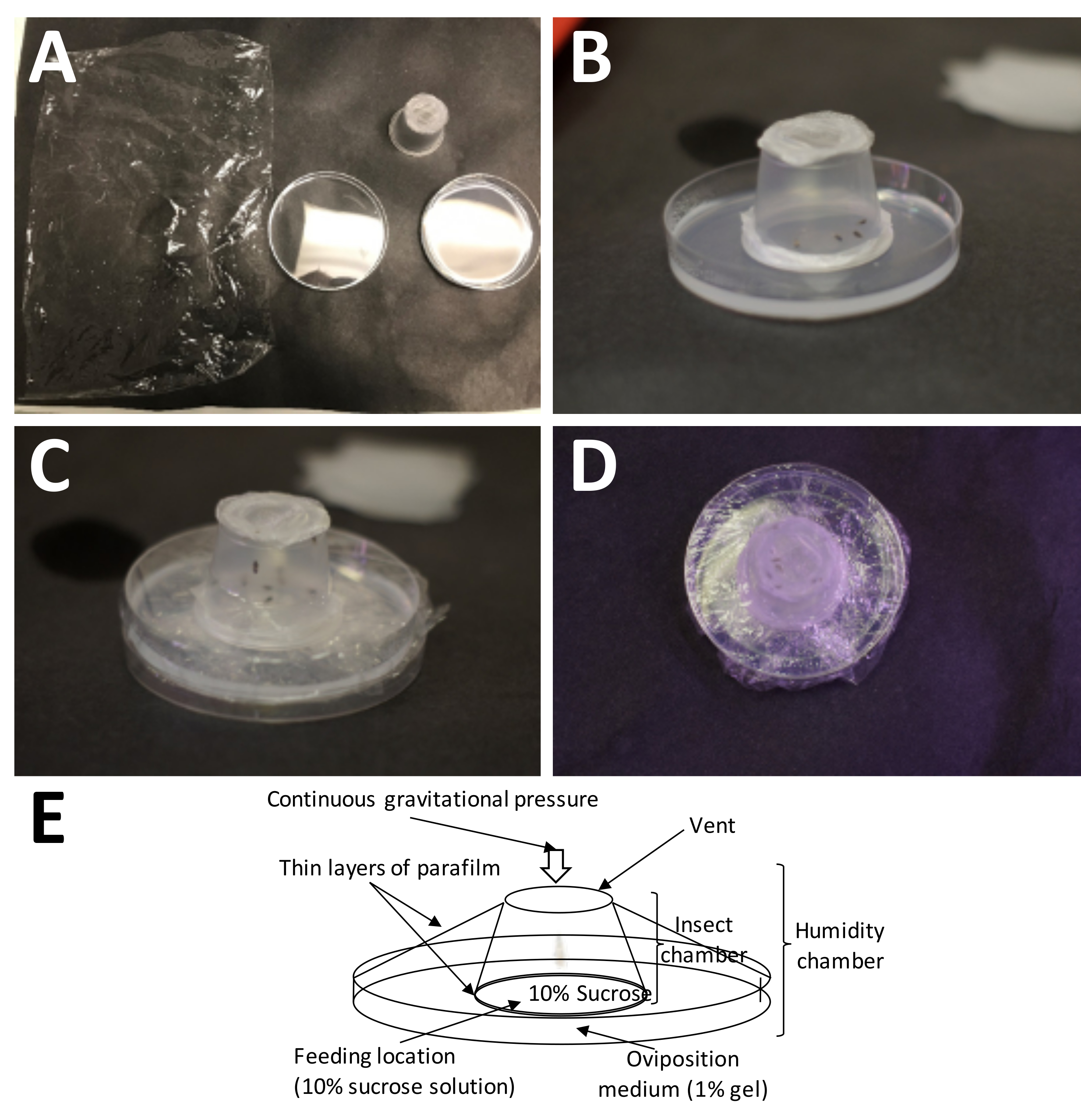
- Сделайте камеру для взрослых особей, вырезав отверстие в дне чашки объемом 1 унция (см. Таблицу материалов) и приклеив над отверстием экран для воздухообмена (рис. 2).
- Разрежьте пластиковую парафиновую восковую пленку на квадраты размером 5 см х 5 см; Отложите по 2 квадрата для каждой чашки.
- Соберите ~ 15 1-недельных взрослых самок из откалиброванной по возрасту колонии P. maidis. Чтобы выбрать самок, осмотрите вентральную сторону брюшка и найдите яйцеклад, который обычно темнее, чем остальная часть брюшка (рис. 3). Держите взрослых особей до одного часа в коническом флаконе объемом 15 мл, если вы устанавливаете несколько камер для яйцекладки. Ненадолго охладите насекомых на льду перед сексом и переложите во взрослый контейнер.
ПРИМЕЧАНИЕ: Это исследование можно проводить без микроскопа. Взрослые самки, у которых было время для кормления и спаривания, также обычно имеют больший живот, чем взрослые самцы, и более послушны; следовательно, их легче выбрать из популяции клеток. - Переложите самок во взрослый контейнер и запечатайте чашку 1 слоем пластиковой парафиновой пленки, равномерно растянув ее в 3-4 раза по сравнению с первоначальным размером (рис. 4A, B).
- Нанесите 400 мкл 10% раствора сахарозы по массе на верхнюю часть уплотнения пластиковой парафиновой пленки и добавьте второй слой пластиковой парафиновой восковой пленки, растягивая пластиковую парафиновую восковую пленку точно так же, как указано выше (рис. 4C, D).
ПРИМЕЧАНИЕ: Сэндвич из растянутой пластиковой парафиновой восковой пленки создает давление раствора сахарозы, что очень важно для кормления взрослых особей, но не мешает самкам прокалывать яйцеклады до упора в среду для яйцекладки. - Поместите взрослую камеру на чашку для сбора яиц пластиковой парафиновой пленкой непосредственно на среду для яйцекладки и оберните всю камеру для яйцекладки полиэтиленовой пленкой, не закрывая отверстия для воздуха, так как они необходимы для воздухообмена (рис. 5).
- Инкубируйте каждую камеру для яйцекладки при температуре 25 °C при влажности 70% и световом цикле 14:10.
- Ежедневно меняйте сэндвич из пластиковой парафиновой восковой пленки и 10% раствора сахарозы и удаляйте воду, которая скапливается внутри чашки.
3. Сбор и выравнивание эмбрионов в среде с высокой влажностью
- Установите систему микроинъекций на основе стереомикроскопа в увлажненном помещении или вытяжке (увлажненный колпак; Рисунок 6) для обеспечения влажности рабочей среды не менее 70% на протяжении всего процесса микроинъекции.
- Проверьте среду для яйцекладки на наличие яиц после желаемого периода яйцекладки. Делают это в увлажненной вытяжке или другой влажной среде.
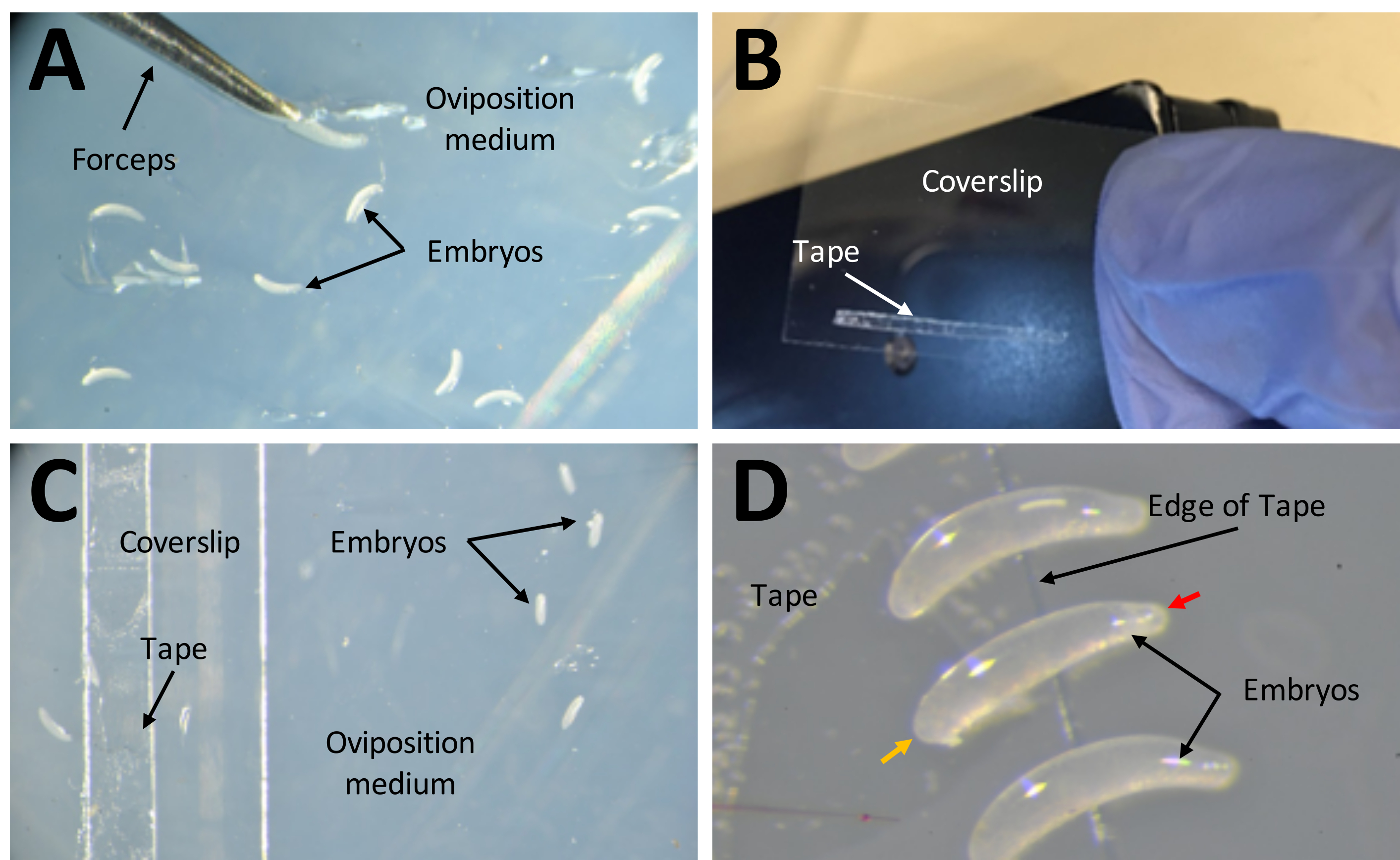
ПРИМЕЧАНИЕ: Обычно используемый период откладывания яиц был ночью, с 6 вечера до 10 утра, продолжительностью ~ 16 часов. - Если какие-либо яйца отложены в агарозе, используйте тонкие щипцы, чтобы осторожно выкопать их, и поместите их на поверхность агарозы, чтобы они оставались влажными (рис. 7A).
- Нанесите полоску двустороннего скотча размером 1 мм x 15 мм на покровное стекло размером 22 мм x 30 мм (рис. 7B). Положите покровную ленту стороной вверх на носитель яйцекладки (рис. 7C).
- Возьмите каждое отдельное яйцо с поверхности агара и переложите на двусторонний скотч с помощью тонкой щетки. Удалите все яйца, которые полностью белые или имеют черную окраску. Здоровые яйца будут полупрозрачными.
- Положите яйца в форме банана на бок так, чтобы больший конец был воткнут в двусторонний скотч (рис. 7D).
ПРИМЕЧАНИЕ: Всегда храните яйца в среде с высокой влажностью, например, в чашке Петри со слоем 1% агара на дне.
4. Приготовление реагентов CRISPR и инъекционных игл
- Вытащите кварцевые иглы с помощью съёмника микропипеток типа Flaming/Brown.
- Скосите кварцевые иглы с помощью микропипетки-фаски.
- Используйте двустороннюю липкую ленту, чтобы закрепить вытянутые иглы в прозрачном контейнере, например в чашке Петри, до тех пор, пока они не будут готовы к использованию.
- Приготовьте раствор для инъекций, объединив 0,5 мкл белка Cas9 (5 мкг/мкл исходного раствора) и 0,5 мкл сгРНК (4 мкг/мкл исходного раствора; см. Таблицу материалов) с 1 мкл фенольного красного буфера в конечном объеме 5 мкл. Чтобы осаждать частицы, которые могут засорить иглу, кратковременно встряхните раствор и центрифугу в течение 3 минут на максимальной скорости.
- Засыпьте инъекционную иглу, стараясь оставить инъекционную смесь рядом с коническим концом иглы. Удалите пузырьки, если таковые имеются, с кончика иглы.
- Осторожно поместите заполненную иглу в иглодержатель и затяните воротник из нержавеющей стали, чтобы надежно удерживать иглу на месте во время микроинъекции.
- Создайте надежный поток раствора для инъекций из иглы, осторожно поглаживая скошенный кончик тонкой влажной кистью, одновременно подавая всплески давления воздуха на иглу с помощью системы впрыска.
ПРИМЕЧАНИЕ: Игла готова к инъекции, когда инъекционная смесь может покинуть наконечник в небольших количествах.
5. Микроинъекционный и постинъекционный уход
- Подготовьте платформу для микроинъекций, заполнив чистую чашку Петри размером 100 мм x 15 мм 1% агара, чтобы сформировать ровный слой агара, который находится на одном уровне с верхней частью чашки.
- Поместите предварительно подготовленное покровное стекло с ~ 25 эмбрионами на агаровую платформу (рис. 8A).
ПРИМЕЧАНИЕ: Все этапы впрыска должны выполняться внутри увлажненного колпака (влажность ~ 70%). - Проверьте давление впрыска, поместив наконечник иглы в каплю воды и начав цикл впрыска.
ПРИМЕЧАНИЕ: Небольшое количество раствора для впрыска должно диспергироваться в воде, если настройка давления правильная (рис. 8B). - Вставьте иглу в больший конец эмбриона, приближаясь с левой стороны покровного стекла (рис. 8C). Доставьте раствор для инъекций в яйцеклетку и быстро вытащите иглу.
- После того, как все яйца будут введены, поместите покровное стекло на поверхность новой чашки с 1% агаром и перенесите блюдо в камеру влажности (рис. 9).
6. Инкубация и вылупление эмбрионов
- Поместите инкубационную камеру в инкубатор с температурой 25 °C на 6 дней.
- Перенесите все выжившие эмбрионы, используя чистую воду и тонкую щетку, в чашку Петри размером 35 мм x 10 мм с увлажненной водой фильтровальной бумагой, покрывающей дно чашки. Запечатайте чашку Петри пластиковой парафиновой пленкой и подержите при температуре 25 °C, чтобы эмбрионы вылупились. Начните проверять эмбрионы на выживаемость через 6 дней после инъекции.
ПРИМЕЧАНИЕ: Нимфы первого возраста начнут вылупляться примерно на 8-й день. - Переложите нимф с помощью тонкой кисти в чашку Петри, содержащую обрезки листьев. Накройте блюдо крышкой и запечатайте полиэтиленовой парафиновой пленкой.
- Инкубируйте запечатанную посуду с детенышами на листовых черенках в течение 48 ч при температуре 25 °C.
- Перенесите всех 2-дневных нимф из раунда инъекций в клетку для выращивания с растениями кукурузы с помощью тонкой щетки. Если инъекционные с видимым фенотипом восстанавливаются в достаточном количестве, выращивайте их отдельно, чтобы максимизировать восстановление целевого признака в следующем поколении. В противном случае проведите массовое спаривание всех инъекционных.
ПРИМЕЧАНИЕ: Аккуратно поместите детенышей в мутовку растения кукурузы, чтобы обеспечить убежище и обеспечить надлежащую влажность их ближайшего окружения. - Выращивайте насекомых в условиях, описанных выше, обеспечивая надлежащую температуру, влажность и регулярные переносы на свежие растения кукурузы.
- Скрининг потомства на ожидаемые фенотипы. Поместите особей, демонстрирующих желаемый фенотип, в их собственную клетку, чтобы установить гомозиготные линии.
Результаты
Камера для откладывания яиц была специально разработана для того, чтобы самки P. maidis могли питаться во время яйцекладки в защитной среде, из которой их яйца могли быть легко извлечены. Используя этот метод, достаточное количество преклеточных эмбрионов было извлечено для микроинъекций с ДНК, РНК и / или белками. Взрослые самки P. maidis обычно откладывают яйца внутри листовой ткани растений кукурузы, что затрудняет получение достаточного количества яиц за короткий промежуток времени, поскольку требует большого вскрытия листьев. Искусственная среда для яйцекладки обеспечивает решение этих проблем. Как показано в таблице 1, за 4 недели было собрано 6 483 яйца у 645 самок. Самки обычно начинают откладывать яйца после 2-го дня и дают большую часть яиц с 4-го по 6-й день. Активность яйцекладки замедлилась к 9 дню. Каждая камера для яйцекладки была установлена в пятницу и проверялась на наличие яиц с воскресенья до следующего воскресенья. Следование этому графику позволило собрать большинство яйцеклеток для микроинъекций в течение рабочей недели.
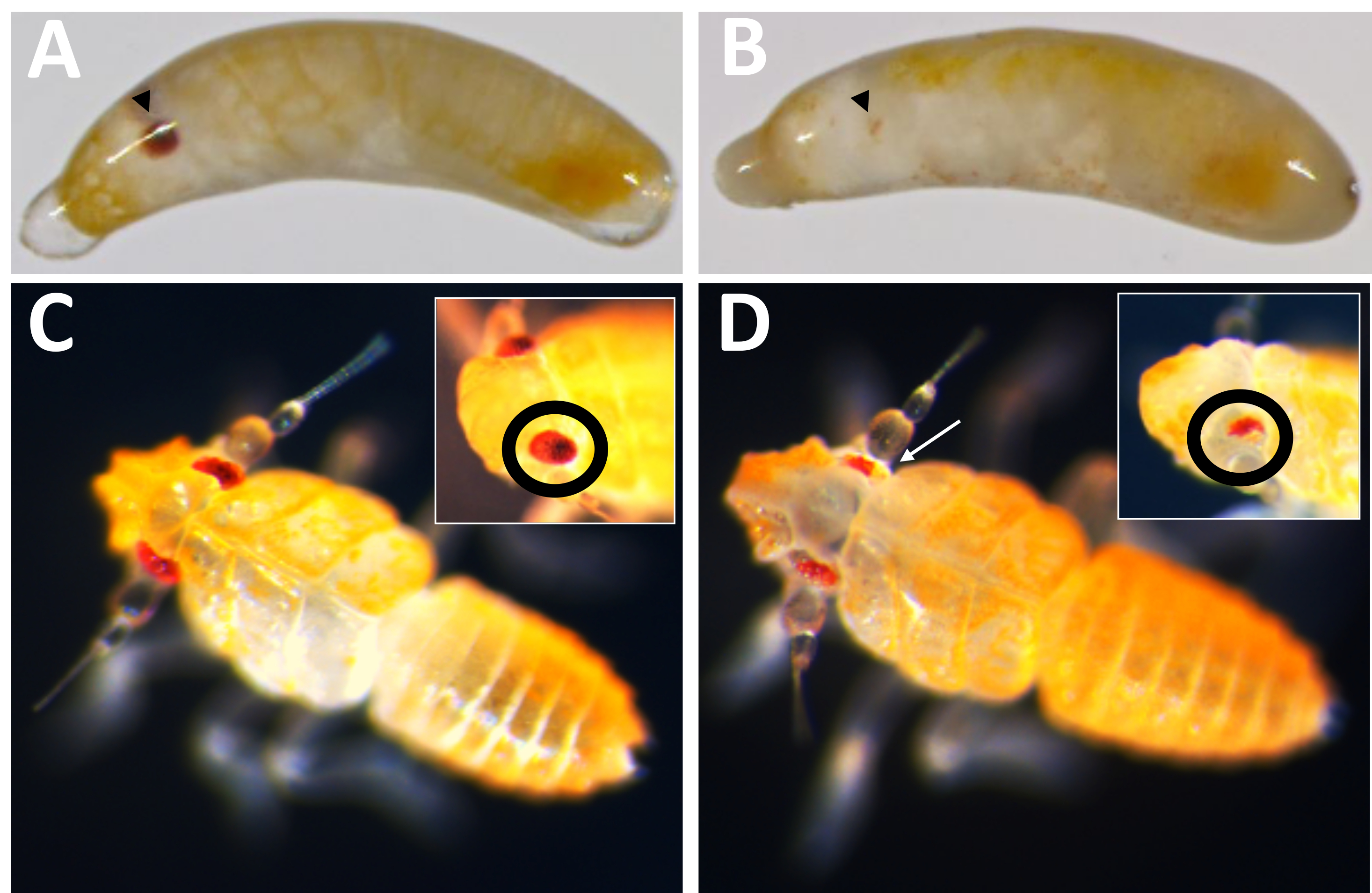
Первым практическим применением этой системы яйцекладки было тестирование эффективности нокаута гена, опосредованного Cas9, с использованием ортолога P. maidis гена цвета глаз, белого (Pmw) в качестве мишени. Известно, что мутации в белом цвете приводят к существенной потере пигментации глаз у других видов насекомых, а белый цвет является клеточно-автономным, что позволяет обнаруживать мутации у инъецированных особей16,17. Чтобы увеличить вероятность того, что даже небольшая мутация может привести к потере функции, направляющие РНК были разработаны для разрезания в области АТФ-связывающей кассеты, которая необходима для функцииУайта 16. Эмбрионам P. maidis вводили либо 20% феноловый красный (инъекционный буфер), либо инъекционный буфер с Cas9 в конечной концентрации 800 нг/мкл (контроль Cas9), либо Cas9 в инъекционном буфере вместе с тремя направляющими РНК, добавленными в концентрации 400 нг/мкл каждая. Комбинация трех направляющих в одной инъекционной смеси была предназначена для дальнейшего максимизации шансов на образование мутантов, как за счет создания большой делеции, так и за счет компенсации возможности того, что какой-либо один проводник может быть неэффективным для резки. Показатели развития для каждого лечения были сопоставимы (таблица 2), при этом у 50-60% инъецированных лиц наблюдались признаки развития. Коэффициенты вылупления для буфера и Cas9 также были сопоставимы; Тем не менее, частота вылупления особей, получавших трехнаправляющую смесь, была относительно ниже. В настоящее время неясно, является ли снижение выживаемости результатом потери функции белого или результатом непреднамеренных последствий трехнаправленной смеси, таких как нецелевые эффекты (см. раздел обсуждения). Тем не менее, ни один из людей с полной потерей пигментации глаз (т.е. полным нокаутом) не вылупился, и ни у одного из потомков инъецированных особей не было белых глаз. Целевая эффективность мутагенеза на основе Cas9 была проверена двумя способами. Во-первых, инъекционные были проверены на потерю пигментации глаз.
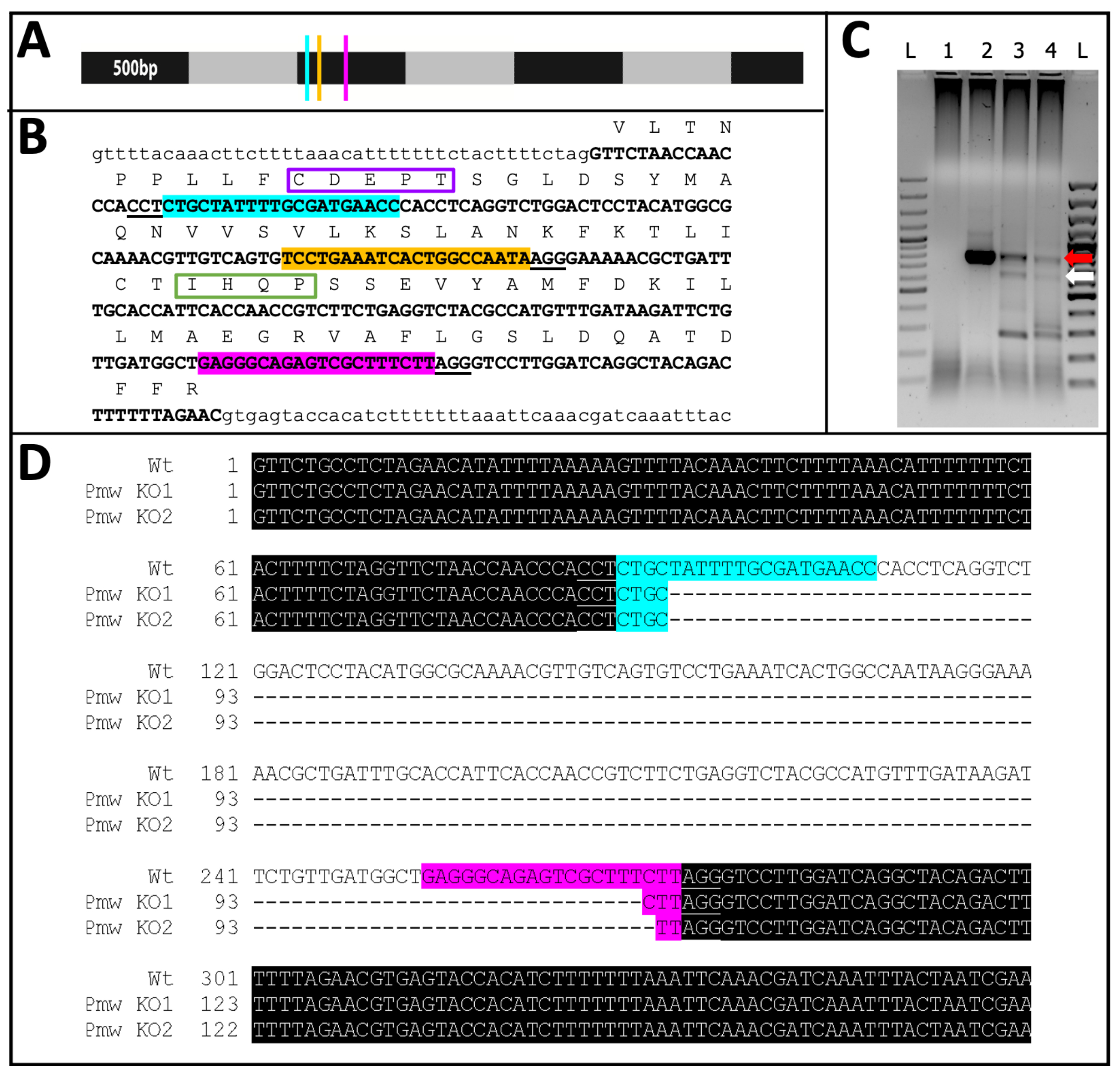
Из 71 человека, получившего инъекцию в гиде, 23 показали некоторую степень потери пигмента (рис. 10), и 9 из этих людей вылупились, что привело к частоте нокаутов ≥32%. Потеря пигмента глаз не наблюдалась ни в одном из контрольных методов лечения. Во-вторых, хромосомные мутации были подтверждены с помощью полимеразной цепной реакции (ПЦР)18 и секвенирования19. Поскольку мутантная линия не могла быть восстановлена, геномная ДНК была проанализирована из пулов эмбрионов, которым вводили либо трехнаправленную смесь, либо буфер. Ожидается, что трехнаправляющая смесь удалит ~ 180 пар оснований из белого локуса. Это можно увидеть в продуктах ПЦР, амплированных из геномной ДНК, выделенной у инъецированных людей, а также в связанных данных о последовательностях, полученных из этих продуктов (рис. 11). Эти комбинированные данные указывают на то, что эмбрионы были введены до того, как произошла клеточная структура.

Рисунок 1: Аспиратор. Эффективный аспиратор может быть собран путем присоединения вакуумного насоса на впуске через пластиковую трубку к пластиковой конической трубке объемом 15 мл. От дна конической трубки следует аккуратно удалить примерно 0,5 см. Ватный тампон следует поместить в коническую трубку над отверстием пластиковой трубки, чтобы поймать взрослых особей P. maidis , когда они собираются, и не допустить их попадания в вакуумный насос. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2: Конструкция контейнеров для взрослых. (A) Необходимые принадлежности (по часовой стрелке сверху слева): экран, пистолет для горячего клея, лезвие бритвы, контейнер на 1 унцию. (B) На дне контейнера емкостью 1 унция должно быть вырезано большое отверстие, и квадрат экрана должен быть достаточно большим, чтобы закрыть это отверстие. (C) Затем экран приклеивается к отверстию с помощью горячего клея. (D) После того, как клей застынет, все лишние сетки должны быть удалены. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3: Определение пола взрослых особей P. maidis. Показаны брюшные стороны взрослых особей P . maidis мужского (слева) и женского (справа). Яйцеклад, видимый над женским животом, является самым четким индикатором пола особи. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 4: Запечатывание взрослых в контейнеры . (A) Квадрат пластиковой парафиновой пленки размером 5 см х 5 см. (B) Пленка должна быть равномерно растянута в 3-4 раза по сравнению с первоначальным размером. (C) После того, как взрослые были помещены в контейнер для взрослых, растянутая пленка должна быть помещена над отверстием, чтобы закрепить взрослых. Затем поверх пленки следует поместить 400 мкл 10% раствора сахарозы по массе. (D) Чтобы обеспечить достаточное давление подачи для взрослых особей, второй квадрат пластиковой парафиновой пленки размером 5 см х 5 см следует аналогичным образом растянуть и поместить поверх капли сахарозы. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 5: Установка камеры для яйцекладки. (A) Необходимые принадлежности (по часовой стрелке сверху слева): полиэтиленовая пленка, готовый контейнер для взрослых (со взрослыми) и чашка Петри с 1% агарозы (среда для откладки яиц). (B) Контейнер для взрослых должен быть помещен на агарозу с пластиковой парафиновой пленкой/10% сахарозой, помещенной непосредственно на среду для яйцекладки. (C) Полиэтиленовая пленка используется для крепления взрослого контейнера к среде для яйцекладки. Это предотвращает слишком быстрое высыхание среды. (D) Следует соблюдать осторожность, чтобы не закрывать сетку контейнера для взрослых, с тем чтобы воздухообмен мог продолжаться. е) Схема камеры яйцекладки. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 6: Увлажненный колпак. Вокруг инжекционного прицела был установлен колпак, оснащенный увлажнителем, чтобы свести к минимуму сквозняки воздуха и поддерживать влажность во время обработки эмбрионов. Заслонки можно сложить над входом после того, как работник окажется на месте, чтобы помочь поддерживать надлежащий уровень влажности. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 7: Сбор эмбрионов при подготовке к инъекциям. (А) Эмбрионы, которые были депонированы в среде для яйцекладки. Пара тонких щипцов используется для извлечения эмбрионов из среды и размещения их на ее поверхности. (B) Узкая полоска двустороннего скотча размером 1 мм х 15 мм на покровном скотче размером 22 мм х 30 мм. (C) Покровное стекло может быть помещено на среду для яйцекладки для облегчения переноса эмбрионов с поверхности среды на ленту на покровном стекло. (D) Эмбрионы P. maidis имеют форму банана, с одним концом, более узким, чем другой (узкий конец обозначен красной головкой стрелки; более широкий конец обозначен желтой головкой стрелки в примере эмбриона). Широкий конец эмбриона следует поместить на ленту. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 8: Инъекция . (A) Инъекционная платформа представляет собой чашку Петри, заполненную до краев 1% агаром. Покровное стекло с полоской ленты, удерживающей эмбрионы, следует помещать непосредственно на поверхность инъекционной платформы. (B) Давление инъекции должно быть проверено до того, как эмбрионы будут введены, путем «введения» небольшого количества раствора для инъекций в каплю воды. Этот метод также можно использовать в любое время в процессе инъекции, чтобы проверить иглу на наличие засоров. (C) Эмбрионы следует вводить путем введения иглы в больший конец эмбриона. Раствор для инъекций должен быть виден, если инъекция прошла успешно. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 9: Послеинъекционный уход . (A) После того, как все эмбрионы на покровном стекло были введены, покровное стекло следует поместить в свежую чашку Петри, содержащую 1% агарозы. (B) Чашку Петри с покровным стеклом затем можно хранить во влажной камере (как показано на рисунке) до тех пор, пока не вылупятся эмбрионы. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 10: Фенотип нокаута PMW . (A) Контрольные эмбрионы соответствующего возраста и (B) нокаутированные эмбрионы Pmw с развивающимися глазами, обозначенными черными наконечниками стрелок. Зародыш в B мозаичный, так как можно увидеть небольшую полоску пигментации. (C) Контрольные по возрасту и (D) детеныши с нокаутом Pmw , со вставками, показывающими другой угол на глазах. Детеныш в D также мозаичный. Белая стрелка указывает на область на основном рисунке, показывающую потерю пигментации. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 11: Последовательность нокаута PMW. (A) Масштабная модель мРНК Pmw , отмеченная с шагом 500.н., с указанием местоположения сайтов связывания гРНК: G1, синий; G2, желтый; G3, розовый. Любые мутации со сдвигом кадров, сгенерированные на этом этапе, нарушат работу большей части продукта перевода. (B) Геномный контекст сайтов гРНК, все в одном экзоне (жирный шрифт с заглавной буквы). Направляющие привязки выделены теми же цветами, что и A, а PAM подчеркнуты. Размах ~300 б.. Внутрикадровый перевод экзона показан выше, в виде однобуквенных аббревиатур заглавным текстом. Отмечены два мотива, характерные для переносчиков пигмента глаза. Мотив CDEPT функционального домена Walker B окрашен в фиолетовый цвет, а мотив IHQP домена H-loop — в зеленый. Оба домена имеют решающее значение для функции АТФ-транспортера. (C) Целевая область Pmw была амплифицирована с помощью двух раундов ПЦР. Продукт второго раунда был исследован на геле на наличие признаков изменения размера из-за успешного удаления области между направляющими. Полосы движения: L = лестница 100.н.; 1 = контроль воды ПЦР; 2 = буферные инъецированные яйца; 3-4 = два отдельных набора яиц, вводимых трехнаправляющей смесью. Только эмбрионы, получавшие трехнаправляющую смесь, продуцировали как полосу WT (красная стрелка), так и полосу, полученную в результате полного иссечения (белая стрелка). (D) Чтобы подтвердить идентичность нижней полосы (белая стрелка), эта ДНК была очищена, клонирована и секвенирована. Верхняя строка — это последовательность дикого типа. Две другие строки являются последовательностями из двух клонов. Три дополнительных клона соответствовали нижней последовательности. Синим цветом обозначено место привязки Guide 1, а розовым выделено место привязки Guide 3. В обоих аллелях вся область между этими двумя направляющими сайтами была удалена. Сокращения: Pmw = белый ген Peregrinus maidis ; гРНК = направляющая РНК; PAM = смежный мотив протоспейсера; АТФ = аденозинтрифосфат; ПЦР = полимеразная цепная реакция; WT = дикий тип; KO = нокаут. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
| Набор | # чашек | # самок в каждой чашке | # яиц | Всего # яиц | |||||||
| День 2 | День 3 | День 4 | День 5 | День 6 | День 7 | День 8 | День 9 | ||||
| 1 | 10 | 15 | 0 | 26 | 166 | 355 | 530 | 193 | 91 | 37 | 1398 |
| 2 | 15 | 15 | 22 | 238 | 489 | 699 | 520 | 379 | 203 | 58 | 2608 |
| 3 | 8 | 15 | 0 | 57 | 230 | 190 | 116 | 80 | 34 | 1 | 708 |
| 4 | 10 | 15 | 0 | 226 | 446 | 519 | 301 | 179 | 24 | 15 | 1710 |
| Итог | 43 | 15 | 23 | 547 | 1331 | 1763 | 1467 | 831 | 352 | 111 | 6483 |
Таблица 1: Репрезентативные коллекции яйцеклеток из искусственной среды яйцекладки. Показаны данные из четырех наборов чашек для сбора яиц, при этом подсчет яиц начинается на второй день после установки и продолжается до девятого дня.
| Инъекционное лечение | Общее количество инъекций | Всего разработано | Всего вылупившихся | Темп развития (%) | Коэффициент вылупления (%) |
| Буфер | 39 | 20 | 12 | 51 | 31 |
| Cas9 | 39 | 24 | 14 | 61 | 36 |
| ГРНК Cas9 + Pmw | 121 | 71 | 28 | 59 | 28 |
Таблица 2: Выживаемость и частота нокаутов при инъекциях 3 различных инъекционных смесей.
Обсуждение
Яйценоскость и питательность
Недавно исследователи, работающие с родственным видом, Nilaparvata lugens, получили яйца, которые они использовали для микроинъекций, непосредственно из листа, сохраняя введенные яйца в ткани листа до тех пор, пока они не вылупятся17. Хотя этот метод на основе листьев обеспечил более естественную среду для эмбрионального развития, он также увеличил вероятность инфекций и повреждения яиц в процессе удаления. Представленная здесь система искусственной яйцекладки обеспечивает более однородную среду и снижает вероятность повреждения яиц при обращении. Установив чашки для яйцекладки в пятницу, большинство яйцекладок были собраны в течение типичной рабочей недели в пользу тех, кто выполнял работу по микроинъекциям. Однако одно предостережение к этому методу заключается в том, что недостаток питательных веществ в рационе с 10% раствором сахарозы в конечном итоге повлияет на здоровье насекомых, и самки в чашках обычно начинают отмирать уже через 10 дней. Качество яиц также начинает падать через 6 дней, о чем свидетельствует увеличение количества мертвых или нездоровых яиц. В результате важно быть избирательным в отношении яиц, используемых для микроинъекций, и не удерживать самок после 6-го дня.
Приживаемость и влажность воздуха
Два фактора, по-видимому, имеют решающее значение для эмбрионального выживания в процессе микроинъекций. Наиболее сложным аспектом работы с эмбрионами P. maidis является предотвращение их высыхания после удаления из среды для яйцекладки и во время микроинъекций. Поскольку яйца обычно откладываются внутри растительной ткани, им не хватает адекватной скорлупы, чтобы предотвратить обезвоживание. Даже в увлажненном колпаке из-за высыхания терялись целые наборы яиц. Однако чрезмерно высокая влажность также может повлиять на микроинъекции, если вода скопится на двустороннем скотче или на прицеле. К сожалению, обезвоживание яйцеклеток обычно было нелегко заметить в процессе микроинъекций, и они часто казались нормальными до 2 или 3 дней спустя, когда они становились полностью прозрачными, не проявляя никаких признаков развития.
Качество иглы также, по-видимому, играет важную роль в выживании. Игла должна быть скошена, чтобы свести к минимуму ненужное повреждение яйца. Когда игла заблокирована, использование функции очистки на инжекторе, осторожно поглаживая кончик иглы влажной кистью (см. шаг 4.7), обычно возвращает иглу в функциональное состояние. Несмотря на это, рекомендуется вводить только небольшое количество инъекционного раствора (~ 0,25 мкл) в каждую иглу и переключаться на новую иглу каждые несколько слайдов (~ 50-60 яиц), чтобы обеспечить поддержание качества иглы на протяжении всего процесса инъекции.
Успешная генерация нокаутного фенотипа
Чтобы успешно трансформировать половые клетки, микроинъекции эмбриона обычно должны быть сделаны как можно раньше до клеточности. В зависимости от вида насекомых временное окно для завершения микроинъекций колеблется от пары часов доцелого дня 14,15,20. До сих пор неясно, когда эмбрионы P. maidis подвергаются клеточному образованию. Cas9-опосредованный нокаут был протестирован на эмбрионах в возрасте от 4 часов после яйцекладки (pel) до 16 часов, и ожидаемые фенотипы наблюдались во всех экспериментах, предполагая, что все микроинъекции были выполнены в течение окна прецеллюризации.
Ортолог P. maidis гена цвета глаз, белый, был выбран потому, что ожидалось, что нокаутный фенотип будет легко скринингом у инъекционных из-за его клеточной автономности. Действительно, как и ожидалось, как мозаичные, так и полные нокауты были четко идентифицированы среди эмбрионов, получавших инъекционную смесь, содержащую Cas9 и направляющие РНК. К сожалению, ни один инъецированный с полным нокаутом не вылупился, и массовое спаривание выживших инъецированных не привело к появлению белоглазого потомства. Тем не менее, мутантная линия была позже успешно сгенерирована путем нацеливания на другой ген (Klobasa et al., в процессе). Это говорит о том, что неспособность установить белую мутантную линию, скорее всего, связана либо с нецелевыми эффектами (т.е. Cas9, разрезающий важные области в других частях генома), порождающими тесно связанную летальную мутацию, либо с непредсказуемой критической ролью белых у P. maidis.
Фенотипические и молекулярные данные (рис. 8 и рис. 9) подтверждают, что в образце инъецированных эмбрионов был создан значительный нокаут в белом локусе, что привело бы к полной потере функции гена. Более того, в то время как мутации в белом цвете жизнеспособны у некоторых видов, есть прецедент, когда снижение активности белых было вредным21,22. Тем не менее, нельзя полностью исключать нецелевые эффекты. Для прогнозирования вероятных нецелевых объектов требуются точные данныепоследовательности генома 23, что в настоящее время невозможно сделать из-за текущего состояния геномных ресурсов P. maidis. Несмотря на это, с помощью этих новых методов можно уверенно тестировать другие гены-мишени, даже переходя к более традиционному трансгенезу в попытке привнести новые генетические инструменты в этот пагубный вредитель.
Раскрытие информации
Авторам раскрывать нечего.
Благодарности
Университет штата Северная Каролина, факультет энтомологии и патологии растений, является частью команды, поддерживающей программу DARPA «Союзники насекомых». Высказанные взгляды, мнения и/или выводы принадлежат авторам и не должны интерпретироваться как представляющие официальную точку зрения или политику Министерства обороны или правительства США. Авторы заявляют об отсутствии конкурирующих интересов. MDL, DR и AEW задумали проект и обеспечили привлечение финансирования, администрирование проекта и ресурсы. FC, WK, NG и MDL задумали и разработали эксперименты по микроинъекциям; О.Х. придумал и разработал метод откладывания яиц. FC и WK провели эксперименты; FC и WK проанализировали результаты; и FC, WK, NG и MDL написали рукопись. Авторы хотели бы выразить особую благодарность Кайлу Созански и Виктории Барнетт за их помощь в поддержании колоний P. maidis.
Материалы
| Name | Company | Catalog Number | Comments |
| 1 oz Containers | Dart | P100N | Adult container for egg-laying setup |
| 15 mL Conical Tubes | Olympus | Genesee 28-103 | Serves as collection tube on vacuum aspirator setup |
| 15 mL Conical Tubes | Olympus | Genesee 28-106 | For making 10% sucorose solution and for holding adults when chilling before screening |
| Aspirator | Bioquip | 1135A | For handling planthoppers |
| Vacuum Aspirator | Fischer Technical | LAV-3 | Vacuum for aspirating larger numbers of insects |
| Blue Spectrum LED Lights | Home Depot | GLP24FS/19W/LED | Grow lights for potted corn plants hoppers are feeding on |
| Cas9 | TrueCut Cas9 Protein v2 | A36498 | Endonuclease for cutting planthopper genes |
| Clear Vinyl Tubing | Home Depot | 3/8 in. I.D. x 1/2 in. O.D. x 10 ft. | Connects collection tube to pump on vacuum aspirator setup |
| Corn planthoppers | North Carolina State University | N/A | Request from Dr. Anna Whitfield's lab |
| Cotton balls | Genessee | 51-101 | Serves as a filter/insect catcher in collection tube on vacuum aspirator setup |
| Double sided tape | Scotch Double Sided Tape | NA | Holding eggs for microinjection |
| Early Sunglow corn | Park Seed Company | 05093-PK-N | Corn for rearing planthoppers |
| epTIPS Microloader Tips | Eppendorf | C2554691 | Backfilling needle loading tips |
| Femtojet Microinjection System | Eppendorf | 5247 | Controls injection pressure (12-20 psi, depending on needle bore size) |
| Nutri-Fly Drosophila Agar | Genessee | 66-103 | Substrate for everything except egg-laying dish |
| Fine forceps | Bioquip | 4731 | Egg handling |
| General Purpose LE Agarose | Apex | 20-102 | Substrate inn egg-laying dish (oviposition medium) |
| Guide RNA 1 - GGUUCAUCGCAAAAUAGCAG | Synthego | CRISPRevolution sgRNA EZ Kit (1.5 nmol) | RNA guides for targeting planthopper white gene |
| Guide RNA 2 - UCUGAAAUCACUGGCCAAUA | Synthego | CRISPRevolution sgRNA EZ Kit (1.5 nmol) | RNA guides for targeting planthopper white gene |
| Guide RNA 3 - GAGGGCAGAGUCGCUUUCUU | Synthego | CRISPRevolution sgRNA EZ Kit (1.5 nmol) | RNA guides for targeting planthopper white gene |
| Humidifyer | Homedics | UHE-CM45 | For providing humidity in humidified hood |
| Humidity chamber | Billups-Rothenberg | MIC-101 | For holding injected embryos until hatching |
| Insect rearing cages | Bioquip (special order) | Close to 1450 L (has plastic front and mesh fabric sides) | Cage for planthoppers on corn |
| Laser-based Micropipiette Puller | Sutter Instruments | P-2000/G | For making injection needles / Heat = 700, FIL = 4, VEL = 40, DEL = 170, PUL = 160 |
| Leica M165 FC Fluorescence Stereomicroscope | Leica | M165 FC | Planthopper screening |
| Microinjection Scope | Leica | MZ12-5 | Microinjection scope outfited with an XY stage |
| Micromanipulator | Narishige | MN-151 | For positioning microinjection needle |
| Micropipette beveler | Sutter Instruments | FG-BV10-D | For beveling injection needles / Used 'fine' graded plate at 20° angle |
| Microscope Stage | AmScope | GT100 X-Y Gliding Table | For positioning and moving embryos under microscope |
| Miniature Paint Brush | Testor #2 8733 | Sold in 3 pack 281206 | Fine paintbrushes for embryo handling |
| Needle Holder | Narishige | HI-7 | For holding the microinjection needle |
| Percival Incubator | Percival | I41VLH3C8 | Rearing injectees until hatch |
| Petri Dishes (100 x 15 mm) | VWR | 89038-968 | Making agar dish for egg-lay |
| pGEM-T Easy Vector System I cloning kit | Promega | A1360 | Cloning Pm white target site |
| Phenol Red | Sigma | 143-78-8 | Microinjection buffer |
| Plain Microscope Slides or coverslip | Fisher Scientific | 12-549-3 | Hold eggs for microinjection |
| Plasmid DNA Midi Kit | Zymo | D4200 | Purification of injection-ready plasmid DNAs |
| Plastic paraffin film | Pechiney Plastic Packaging | PM-996 | Roll size 4 in. x 125 ft |
| Plastic wrap | Glad ClingWrap Plastic Wrap | NA | Wrap the entire egg-laying chamber |
| Primer - PmW CRISPR check F1 - AAGGAATTTCTGGAGGTGAAA | IDT | 25 nmole DNA Oligo | First-round Primer for amplifing across target site within the Pm white gene |
| Primer - PmW CRISPR check R1 - GATTCCTCGCTGTTGGGT | IDT | 25 nmole DNA Oligo | First-round Primer for amplifing across target site within the Pm white gene |
| Primer - PmW CRISPR check F3 - TCACAGACCCTGGTGCTAATC | IDT | 25 nmole DNA Oligo | Second-round Primer for amplifing across target site within the Pm white gene |
| Primer - PmW CRISPR check R3 - GTCCACAATCCACACTTCTGA | IDT | 25 nmole DNA Oligo | Second-round Primer for amplifing across target site within the Pm white gene |
| Quartz capillaries | Sutter Instruments | QF100-50-10 | For making microinjection needles / O.D. 1 mm, I.D. 0.7 mm, 10 cm length |
| Screen (White Organza Fabric) | Joann Fabrics | 16023889 | For covering the adult container |
| Sparkleen | Fisher Scientific | 04-320-4 | Wash dishes |
| Sucrose | Fisher Scientific | BP220-1 | To make 10% sucorose solution |
Ссылки
- Namba, R., Higa, S. Y. Host plant studies of the corn planthopper, Peregrinus maidis (Ashmead) in Hawaii. Proceedings of the Hawaiian Entomological Society. 21, 105-108 (1971).
- Singh, B. U., Seetharama, N. Host plant interactions of the corn planthopper, Peregrinus maidis Ashm.(Homoptera: Delphacidae) in maize and sorghum agroecosystems. Arthropod-Plant Interactions. 2 (3), 163-196 (2008).
- Tsai, J. Occurrence of a corn disease in Florida transmitted by Peregrinus maidis. Plant Disease Reporter. 59 (10), 830-833 (1975).
- Chelliah, S., Basheer, M. Biological studies of Peregrinus maidis (Ashmead) (Araeopidae: Homoptera) on sorghum. Indian Journal of Entomology. 27, 466-471 (1965).
- Lastra, J., Esparza, J. Multiplication of vesicular stomatitis virus in the leafhopper Peregrinus maidis (Ashm.), a vector of a plant rhabdovirus. Journal of General Virology. 32 (1), 139-142 (1976).
- Nault, L. R., Ammar, E. -. D. Leafhopper and planthopper transmission of plant viruses. Annual Review of Entomology. 34 (1), 503-529 (1989).
- Ammar, E. -. D., Tsai, C. -. W., Whitfield, A. E., Redinbaugh, M. G., Hogenhout, S. A. Cellular and molecular aspects of rhabdovirus interactions with insect and plant hosts. Annual Review of Entomology. 54, 447-468 (2009).
- Barandoc-Alviar, K., Ramirez, G. M., Rotenberg, D., Whitfield, A. E. Analysis of acquisition and titer of Maize mosaic rhabdovirus in its vector, Peregrinus maidis (Hemiptera: Delphacidae). Journal of Insect Science. 16 (1), 14 (2016).
- Tsai, J. H., Steinberg, B., Falk, B. W. Effectiveness and residual effects of seven insecticides on Dalbulus maidis (Homoptera: Cicadellidae) and Peregrinus maidis (Homoptera: Delphacidae). Journal of Entomological Science. 25 (1), 106-111 (1990).
- Yao, J., Rotenberg, D., Afsharifar, A., Barandoc-Alviar, K., Whitfield, A. E. Development of RNAi methods for Peregrinus maidis, the corn planthopper. PloS One. 8 (8), 70243 (2013).
- Esvelt, K. M., Smidler, A. L., Catteruccia, F., Church, G. M. Emerging technology: concerning RNA-guided gene drives for the alteration of wild populations. Elife. 3, 03401 (2014).
- Gantz, V. M., Bier, E. The mutagenic chain reaction: a method for converting heterozygous to homozygous mutations. Science. 348 (6233), 442-444 (2015).
- Yao, J., Rotenberg, D., Whitfield, A. E. Delivery of maize mosaic virus to planthopper vectors by microinjection increases infection efficiency and facilitates functional genomics experiments in the vector. Journal of Virological Methods. 270, 153-162 (2019).
- Kimelman, D., Martin, B. L. Anterior-posterior patterning in early development: three strategies. Wiley Interdisciplinary Reviews: Developmental Biology. 1 (2), 253-266 (2012).
- Mito, T., Nakamura, T., Noji, S. Evolution of insect development: to the hemimetabolous paradigm. Current Opinion in Genetics & Development. 20 (4), 355-361 (2010).
- Grubbs, N., Haas, S., Beeman, R. W., Lorenzen, M. D. The ABCs of eye color in Tribolium castaneum: orthologs of the Drosophila white, scarlet, and brown Genes. Genetics. 199 (3), 749-759 (2015).
- Xue, W. H., et al. CRISPR/Cas9-mediated knockout of two eye pigmentation genes in the brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Insect Biochemistry and Molecular Biology. 93, 19-26 (2018).
- Lorenz, T. C. Polymerase chain reaction: basic protocol plus troubleshooting and optimization strategies. Journal of Visualized Experiments: JoVE. (63), e3998 (2012).
- Sanger, F., Coulson, A. R. A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. Journal of Molecular Biology. 94 (3), 441-448 (1975).
- Chu, F. C., Wu, P. S., Pinzi, S., Grubbs, N., Lorenzen, M. D. Microinjection of Western Corn Rootworm, Diabrotica virgifera virgifera, embryos for germline transformation, or CRISPR/Cas9 genome editing. Journal of Visualized Experiments: JoVE. (134), e57497 (2018).
- Brent, C. S., Hull, J. J. RNA interference-mediated knockdown of eye coloration genes in the western tarnished plant bug (Lygus hesperus Knight). Archives of Insect Biochemistry and Physiology. 100 (2), 21527 (2019).
- Khan, S. A., Reichelt, M., Heckel, D. G. Functional analysis of the ABCs of eye color in Helicoverpa armigera with CRISPR/Cas9-induced mutations. Scientific Reports. 7 (1), 1-14 (2017).
- Manghwar, H., et al. CRISPR/Cas systems in genome editing: methodologies and tools for sgRNA design, off-target evaluation, and strategies to mitigate off-target effects. Advanced Science. 7 (6), 1902312 (2020).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены