Method Article
Флуоресценция РНК in situ гибридизация (FISH) для визуализации микробной колонизации и инфекции в кишечнике Caenorhabditis elegans
В этой статье
Резюме
Кишечные микробы, включая внеклеточные бактерии и внутриклеточные патогены, такие как вирус Орсе и микроспоридия (грибы), часто связаны с дикими нематодами Caenorhabditis . В этой статье представлен протокол для обнаружения и количественной оценки микробов, которые колонизируют и / или заражают нематод C. elegans , а также для измерения патогенной нагрузки после контролируемых инфекций в лаборатории.
Аннотация
Кишечник диких нематод Caenorhabditis населен различными микроорганизмами, включая бактерии кишечного микробиома и патогены, такие как микроспоридии и вирусы. Из-за сходства между Caenorhabditis elegans и клетками кишечника млекопитающих, а также из-за силы системы C. elegans , этот хозяин появился в качестве модельной системы для изучения взаимодействия кишечника и микроба хозяина in vivo. Хотя можно наблюдать некоторые аспекты этих взаимодействий с помощью микроскопии яркого поля, трудно точно классифицировать микробы и охарактеризовать степень колонизации или инфекции без более точных инструментов. Флуоресцентная гибридизация РНК in situ (FISH) может быть использована в качестве инструмента для идентификации и визуализации микробов у нематод из дикой природы или для экспериментальной характеристики и количественной оценки инфекции у нематод, инфицированных микробами в лаборатории. Зонды FISH, маркирующие высокообильную небольшую субъединицу рибосомальной РНК, производят яркий сигнал для бактерий и микроспоридиальных клеток. Зонды, предназначенные для нацеливания на сохраненные области рибосомной РНК, общие для многих видов, могут обнаруживать широкий спектр микробов, тогда как нацеливание на расходящиеся области рибосомальной РНК полезно для более узкого обнаружения. Точно так же зонды могут быть разработаны для маркировки вирусной РНК. Представлен протокол окрашивания РНК FISH либо параформальдегидом (PFA), либо фиксацией ацетона. Фиксация PFA идеально подходит для нематод, связанных с бактериями, микроспоридиями и вирусами, тогда как фиксация ацетона необходима для визуализации спор микроспорида. Животных сначала промывали и фиксировали в параформальдегиде или ацетоне. После фиксации зонды FISH инкубировали с образцами, чтобы обеспечить гибридизацию зондов с желаемой целью. Животных снова мыли, а затем исследовали на предметных стеклах микроскопа или с использованием автоматизированных подходов. В целом, этот протокол FISH позволяет обнаруживать, идентифицировать и количественно оценивать микробы, населяющие кишечник C. elegans , включая микробы, для которых нет доступных генетических инструментов.
Введение
Caenorhabditis elegans стал мощной модельной системой для изучения врожденного иммунитета и взаимодействия хозяина и микроба в эпителиальных клетках кишечника 1,2. Благодаря наличию прозрачного тела и всего 20 кишечных клеток, C. elegans представляет собой удобную систему мониторинга процессов микробной кишечной колонизации и инфицирования в условиях интактного организма. Клетки кишечника нематоды имеют множество морфологических и функциональных сходств с эпителиальными клетками кишечника млекопитающих, что делает их податливой моделью in vivo для рассечения процессов, которые управляют колонизацией микробиома и патогенной инфекцией 3,4,5,6.
Дикие C. elegans питаются различными микробами, которые колонизируют и заражают кишечник, и отбор проб этих нематод привел к открытию вирусов, эукариот (грибов, оомицетов) и бактерий, которые естественным образом связываются с этим хозяином 7,8,9,10. Вирус Орсе был обнаружен заражающим кишечник и в настоящее время является единственным известным природным вирусом C. elegans9. Микроспоридия - это связанные с грибами облигатные внутриклеточные патогены, которые являются наиболее часто встречающейся инфекцией при диком caenorhabditis, причем было обнаружено несколько видов, заражающих C. elegans и родственных нематод 8,11. Многие бактерии обычно обитают в кишечном просвете пойманных в дикой природе C. elegans, и несколько видов были установлены в качестве естественной модели для микробиома C. elegans (CeMbio)6,12,13,14. Обнаружение и характеристика микробов, которые естественным образом колонизируют и / или заражают C. elegans, имеет важное значение для понимания генетических механизмов, которые управляют этими взаимодействиями хозяина и микроба, а также для визуализации новых микробных процессов, которые происходят только в контексте интактного животного-хозяина.
После отбора проб дикие нематоды проверяются с помощью дифференциальной интерференционно-контрастной (ДВС-матрицы) для поиска фенотипов, которые указывают на инфекцию или колонизацию. Например, изменения стереотипного гранулированного внешнего вида клеток кишечника могут быть связаны с наличием внутриклеточной паразитарной инфекции8. В частности, потеря кишечных гранул и снижение цитозольной вязкости являются признаками вирусной инфекции, тогда как реорганизация кишечных гранул в «бороздки» может указывать на заражение микроспоридиями рода Nematocida 8,9. Поскольку существует большое разнообразие микробов, присутствующих в диких образцах C. elegans, может быть трудно различить микробы с помощью ДВС-микроскопии. Информацию о пространственном распределении микробов внутри хозяина также может быть трудно обнаружить из-за небольшого размера многих микробов15. Кроме того, культивирование каких-либо конкретных микробов, представляющих интерес in vitro, не всегда возможно, что приводит к трудностям в обнаружении и / или количественной оценке.
Флуоресцентная гибридизация РНК in situ (FISH) обеспечивает метод флуоресцентной маркировки микробов с использованием флуоресцентных зондов, которые связываются с РНК малой рибосомной субъединицы (SSU) в фиксированных клетках. Если анализ морфологических характеристик предполагает определенный класс микробов, могут быть использованы зонды FISH, нацеленные на конкретные или широкие классы таких микробов. Например, EUB338 считается универсальным зондом для бактериальной СБУ и обычно используется для обнаружения широкого спектра бактерий16. Протокол, описанный здесь, использует одноцепочечные ДНК-зонды, которые помечены флуорофором и специально разработаны, чтобы быть комплементарными к целевому SSU интересующего микроба, хотя есть ранее разработанные зонды, доступные16. Основным преимуществом нацеливания на SSU микробов является относительно большое содержание этой РНК, которая обычно составляет 80%-90% всей РНК в клетке, что приводит к окрашиванию с очень высоким отношением сигнал/шум17. Зонды также могут быть разработаны для нацеливания на РНК для обнаружения вирусов, таких как вирус Орсе 9,18, которые часто присутствуют в очень высоких копиях в инфицированных клетках, если вирус активно реплицируется.
В зависимости от результатов с известными зондами может потребоваться получение дополнительной информации о последовательности для разработки более конкретных зондов для подтверждения видов in situ. Общий подход заключается в использовании универсальных праймеров против законсервированных областей SSU (16S для бактерий и 18S для эукариот) для амплификации (с помощью ПЦР) областей, которые являются более расходящимися8. Используя эту информацию о последовательности, можно разработать зонды с большей видовой специфичностью. Эти зонды FISH могут затем обеспечить идентификацию микробов независимым от культуры способом8. Кроме того, РНК FISH может дать представление об уникальных морфологических характеристиках колонизации и инфекции, включая паттерны нити или локализациитканей 19,20. Одновременно могут использоваться различные цветные зонды FISH, что позволяет визуально различать микробы в образцах диких нематод, а также наблюдать динамику микроб-микроб внутри хозяина15,20. Кроме того, окрашивание РНК FISH может быть применено к исследованиям взаимодействия хозяина и патогена, где инфекция и колонизация известного вида могут быть легко количественно определены вручную или с помощью автоматизированных подходов для получения информации о патогенной нагрузке, например, при сравнении мутантов C. elegans, которые либо увеличили, либо уменьшили устойчивость к инфекции21.
протокол
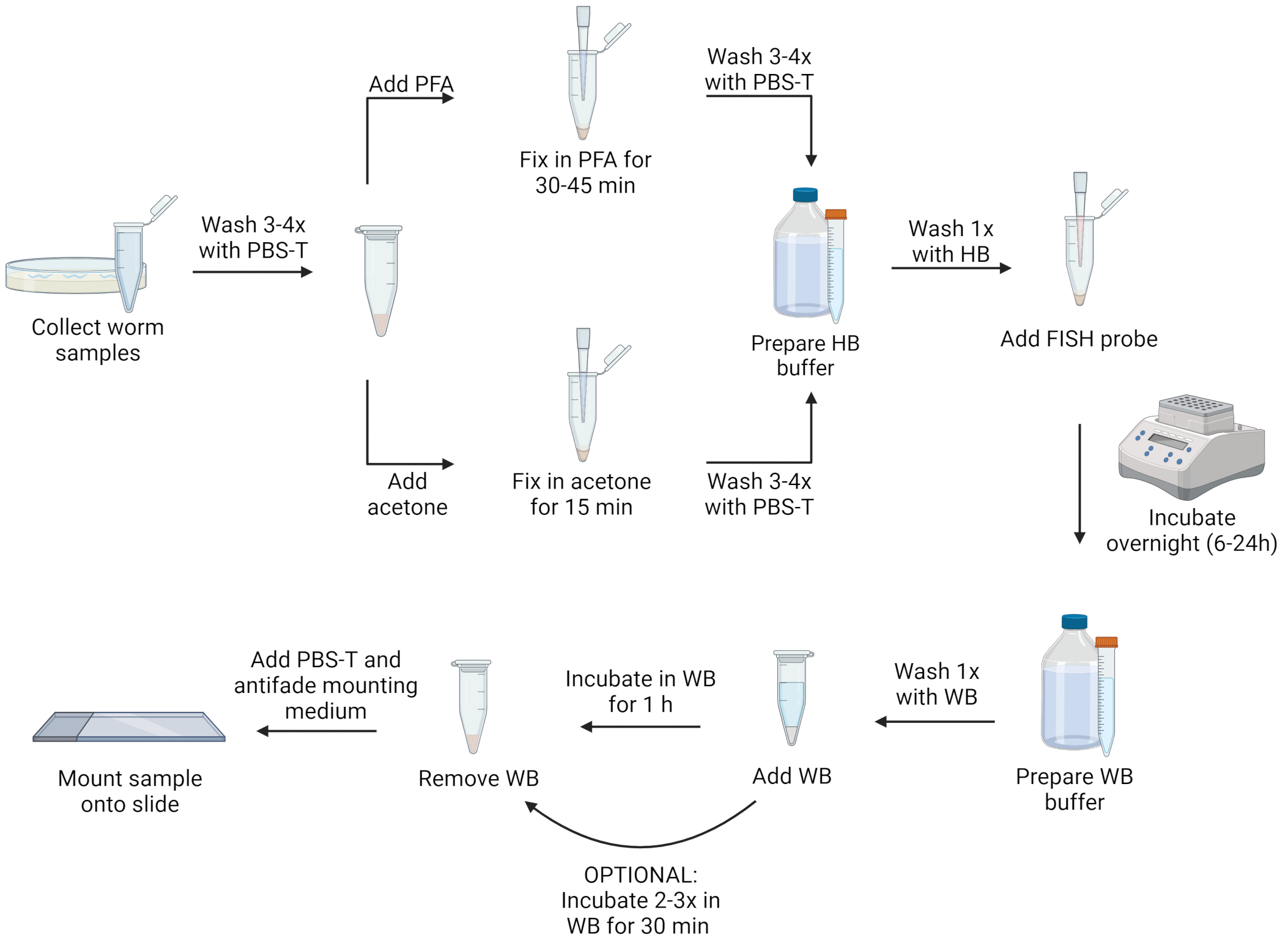
ПРИМЕЧАНИЕ: Нематоды могут быть зафиксированы либо раствором параформальдегида (PFA), либо ацетоном. PFA позволяет лучше визуализировать морфологию, чем ацетон, и может сохранять сигналы от трансгенного зеленого флуоресцентного белка (GFP), который разрушается ацетоном. Тем не менее, фиксация ацетона необходима для пермеабилизации микроспоридиевых спор, чтобы позволить маркировать эту стадию жизни. Кроме того, ацетон может быть более удобным, чем PFA, потому что он менее токсичен, и образцы могут храниться в течение нескольких дней в ацетоне в морозильной камере с температурой -20 ° C без необходимости удаления фиксатора. Ниже приведены два отдельных протокола, использующих либо раствор PFA, либо ацетон в качестве фиксатора. Визуализацию шагов протокола см. на рисунке 1.
1. Окрашивание FISH с фиксацией PFA
- Подготовка нематод с ассоциированными микробами
- Выращивайте нематод с желаемым микробом, представляющим интерес, на стандартных пластинах Nematode Growth Media (NGM), засеянных соответствующим источником пищи. Инкубируйте нематод при 20 °C до тех пор, пока не будет достигнута желаемая стадия жизни.
- Добавьте 2 мл среды минимальных солей M9 (42 мМ Na2HPO4, 22 мМ KH2PO4, 8,6 мМ NaCl, 19 мМ NH4Cl) + 0,1% Tween 20 к пластинам NGM, содержащим штамм Caenorhabditis , инфицированный или колонизированный желаемым микробом для визуализации.
ПРИМЕЧАНИЕ: Добавление моющего средства предназначено для предотвращения прилипания нематод к пипеткам и микрофьюж-трубкам, а также для помощи гранулированным нематодам стадии L1-L2. 0,1% Triton-X можно использовать вместо Tween 20. - Пипетки поднимают нематоды с пластин с помощью стеклянной пипетки и колбы Пастера и переносят их в маркированные микрофьюжные трубки объемом 1,5 мл.
ПРИМЕЧАНИЕ: Стеклянные пипетки предпочтительнее, потому что нематоды могут прилипать к пластиковым пипеткам, но добавление моющего средства (Tween 20 или Triton-X) может свести к минимуму эту проблему. - Используя микроцентрифугу, раскрутите образцы, содержащие колонизированных или инфицированных нематод, на 2000 х г в течение 60 с для животных L1 или 500 х г в течение 60 с для L4 или взрослых животных. Все последующие этапы центрифугирования будут выполняться на выбранной скорости.
- Извлеките супернатант из микрофрагмированных трубок с помощью пипетки. Избегайте нарушения гранулы нематоды, осторожно удаляя супернатант до 100 мкл над гранулой. Это можно оценить с помощью маркированных микрофьюжных трубок объемом 1,5 мл.
- Мойте нематоды для устранения внешних загрязнений
- Добавьте 1 мл 1x PBS (137 мМ NaCl, 2,7 мМ KCl, 10 мМ Na2HPO4, 1,8 мМ KH2PO4) + 0,1% Tween 20 (PBS-T) в микрофьюжные трубки.
- Вращайте образцы в микроцентрифуге с соответствующей скоростью (см. шаг 1.1.4). Используя пипетку, удалите все, кроме 100 мкл супернатанта. Это можно оценить с помощью маркированных микрофьюжных трубок объемом 1,5 мл.
- Повторите два вышеуказанных шага 2-3x.
ПРИМЕЧАНИЕ: Как правило, достаточно трех общих стирок; тем не менее, могут быть выполнены дополнительные промывки для удаления любых избыточных внешних загрязнений.
- Исправление нематод с помощью PFA
- В вытяжной капюшон добавляют 33 мкл 16% PFA в микрофьюжную трубку, содержащую 100 мкл супернатанта над гранулой нематоды, полученной на стадии 1.2.3, для конечной концентрации 4% PFA.
ВНИМАНИЕ: PFA является канцерогеном. Контакт с PFA может привести к сенсибилизации и раздражению кожи и повреждению глаз. PFA выделяет токсичные пары, которые могут привести к раздражению или сенсибилизации дыхательных путей. При использовании PFA работайте в вытяжном шкафу с надлежащими средствами индивидуальной защиты и обратитесь к соответствующим паспортам безопасности перед использованием. - Инкубируют образцы, содержащие нематод, колонизированных или зараженных интересующим микробом, в течение 30-45 мин при комнатной температуре. После инкубации храните образцы в 70% этаноле при 4 °C до тех пор, пока протокол не будет готов к продолжению.
ПРИМЕЧАНИЕ: Более короткий инкубационный период лучше подходит для поддержания сигнала GFP у трансгенных штаммов из-за деградации GFP PFA с течением времени. Более длительное время инкубации позволяет фиксатору лучше проникать в образцы. Лучше всего определять время инкубации эмпирически в зависимости от образца.
- В вытяжной капюшон добавляют 33 мкл 16% PFA в микрофьюжную трубку, содержащую 100 мкл супернатанта над гранулой нематоды, полученной на стадии 1.2.3, для конечной концентрации 4% PFA.
- Удаление решения PFA
- Вращайте образцы в микроцентрифуге с соответствующей скоростью (см. шаг 1.1.4). С помощью пипетки удалите как можно больше супернатанта, не нарушая гранулу.
ВНИМАНИЕ: Супернатант содержит PFA, который токсичен. Выбросьте супернатант и, по крайней мере, первые две промывки в качестве токсичных отходов в вытяжной вытяжке. - Добавьте 0,5 мл PBS-T к образцам в микрофьюжных пробирках.
- Выполните и повторите шаги 1.4.1 и 1.4.2 2-3x с PBS-T.
ПРИМЕЧАНИЕ: Выполнение большего количества стирок поможет уменьшить фоновый сигнал. В общей сложности рекомендуется не менее четырех стирок. - После последней промывки открутите образцы и удалите супернатант, оставив гранулу нетронутой.
- Вращайте образцы в микроцентрифуге с соответствующей скоростью (см. шаг 1.1.4). С помощью пипетки удалите как можно больше супернатанта, не нарушая гранулу.
- Подготовьте буфер гибридизации (HB) и промывайте нематод
- Приготовьте 1 мл HB (900 мМ NaCl, 20 мМ Tris pH 7,5, 0,01% SDS) на образец.
ПРИМЕЧАНИЕ: HB следует готовить свежим перед каждым использованием, чтобы избежать осадков. Однако общий буфер (900 мМ NaCl, 20 мМ Tris pH 7,5) может быть изготовлен заранее и храниться при комнатной температуре до тех пор, пока не понадобится HB. Перед применением готовят 1 мл на образец общего буфера и добавляют SDS к конечной концентрации 0,01%. - Добавьте 800 мкл HB в микрофьюжные трубки, содержащие гранулу нематоды. Гранулирование образцов в микроцентрифуге (см. этап 1.1.4). Удалите супернатант, не нарушая гранулу.
- Приготовьте 1 мл HB (900 мМ NaCl, 20 мМ Tris pH 7,5, 0,01% SDS) на образец.
- Гибридизация зонда FISH с требуемой целевой последовательностью
- Смешайте 100 мкл на образец подготовленного HB с желаемым зондом FISH до конечной концентрации 5-10 нг/мкл зонда.
ПРИМЕЧАНИЕ: Зонды FISH представляют собой 15-23-мер олиго, антисмысловые для SSU интересующего микроба и помечены цветным флуорофором, прикрепленным к 5' или 3' концу (см. Таблицу 1 для зондов, используемых здесь). Как правило, стоковые зонды FISH хранятся при 1 мг/мл. - Добавьте 100 мкл HB, содержащего зонд FISH, к каждому образцу. Перемешайте, осторожно щелкнув или перевернув трубки.
ПРИМЕЧАНИЕ: Различные цветные датчики FISH могут быть добавлены одновременно для визуализации нескольких флуоресцентных сигналов в одном образце (см. Рисунок 1B). - Инкубируют образцы в течение ночи (6-24 ч) в сухой ванне при 46-54 °C или термомешалке при 46-54 °C при 1 200 об/мин.
ПРИМЕЧАНИЕ: Инкубация при 46-48 °C обычно используется для гибридизации. Однако эту температуру, возможно, потребуется отрегулировать в зависимости от температуры плавления датчика FISH. В целом, температура гибридизации на 4 °C ниже температуры плавления.
- Смешайте 100 мкл на образец подготовленного HB с желаемым зондом FISH до конечной концентрации 5-10 нг/мкл зонда.
- Снимите зонд FISH и мойте нематод
- Подготовьте 3 мл промывочного буфера (WB) (900 мМ NaCl, 20 мМ Tris pH 7,5, 5 мМ ЭДТА, 0,01% SDS) на образец.
ПРИМЕЧАНИЕ: WB следует готовить свежим перед каждым использованием, чтобы избежать осадков. Однако общий буфер (900 мМ NaCl, 20 мМ Tris pH 7,5) может быть изготовлен заранее и храниться при комнатной температуре до тех пор, пока не понадобится буфер для стирки. Перед применением готовят 3 мл на образец общего буфера и добавляют ЭДТА к конечной концентрации 5 мМ и SDS к конечной концентрации 0,01%. - Центрифугирование образцов с соответствующей скоростью (см. этап 1.1.4). Удалите ГВ с помощью пипетки, при этом стараясь оставить гранулу нематоды нетронутой.
- Добавьте 1 мл подготовленного WB к каждому образцу.
- Центрифугирование образцов с соответствующей скоростью (см. этап 1.1.4). Снимите WB с помощью пипетки, при этом стараясь оставить гранулу нетронутой.
- Добавьте 1 мл подготовленного WB к каждому образцу.
- Инкубировать образцы в течение 1 ч при 48-56 °C в сухой ванне (или термальном смесителе при 48-56 °C при 1200 об/мин). При инкубации в сухой ванне осторожно переворачивайте трубки каждые 15-20 мин.
ПРИМЕЧАНИЕ: Как правило, 48 °C используется в качестве стандартной температуры стирки; однако эту температуру, возможно, потребуется отрегулировать при наличии высокого фонового сигнала. Температура стирки часто на 2 °C выше, чем температура гибридизации. Время инкубации в ВБ может быть сокращено до 30 мин для дальнейшего уменьшения фона, после чего следует повторить этапы 1.7.4 и 1.7.5. За этим следует один (для бактерий) или два (для микроспоридий спороплазм) 30-минутный инкубационный период при 48 °C. - Центрифугирование образцов с соответствующей скоростью (см. этап 1.1.4). Используя пипетку, снимите буфер для стирки, стараясь оставить гранулу нетронутой.
- Добавьте 100-500 мкл PBS-T к каждому из образцов. На этом этапе образцы могут храниться в PBS-T при 4 °C в течение недели, пока протокол не будет готов к продолжению.
- Подготовьте 3 мл промывочного буфера (WB) (900 мМ NaCl, 20 мМ Tris pH 7,5, 5 мМ ЭДТА, 0,01% SDS) на образец.
- Крепление нематод
- Центрифугирование образцов с соответствующей скоростью (см. этап 1.1.4). Удалите как можно больше PBS-T, не нарушая гранулы нематоды.
- (Необязательно) Добавьте к образцам 20 мкл антиотмерзающей монтажной среды с DAPI (Таблицей материалов).
- Загрузите пипеттор объемом 20 мкл с наконечником пипетки объемом 200 мкл и используйте ножницы, чтобы отрезать кончик пипетки, чтобы позволить пипетке более крупных нематод.
- С помощью разрезанного наконечника пипетки переложите 5-10 мкл гранулы на предметное стекло микроскопа. Крышка с чехлом 22 x 22. Для хранения слайдов запечатайте края обшивки лаком для ногтей и держите их в темной коробке при 4 °C до готовности к дальнейшему использованию.
2. Окрашивание FISH с фиксацией ацетона
- Приготовьте нематод с ассоциированными микробами, как описано в шаге 1.1.
- Мойте нематоды для устранения внешних загрязнений, как описано в шаге 1.2.
- Фиксация нематод ацетоном
- Удалите супернатант, не нарушая гранулу, и добавьте в образец 1 мл лабораторного ацетона.
ВНИМАНИЕ: Ацетон является легковоспламеняющейся жидкостью и парами. Хотя его можно приобрести без рецепта в качестве средства для снятия лака для ногтей, важно помнить, что ацетон вызывает серьезное раздражение глаз и может вызвать сонливость или головокружение. - Инкубируйте образцы, содержащие нематод, колонизированных или зараженных интересующим микробом, в течение 15 мин при комнатной температуре. После инкубации образцы могут храниться в ацетоне при -20 °C в течение 2 недель, пока протокол не будет готов к продолжению.
ПРИМЕЧАНИЕ: Не используйте этот протокол с трансгенными штаммами C. elegans , экспрессирующими GFP (или его аллельные мутированные формы), если это необходимо для поддержания флуоресценции, так как ацетон разрушает этот сигнал.
- Удалите супернатант, не нарушая гранулу, и добавьте в образец 1 мл лабораторного ацетона.
- Удалите ацетон
- Вращайте образцы в микроцентрифуге с соответствующей скоростью (см. шаг 1.1.4). С помощью пипетки удалите супернатант, не нарушая гранулу.
ВНИМАНИЕ: Супернатант содержит ацетон. Выбросьте супернатант и, по крайней мере, первые две промывки в качестве токсичных отходов в вытяжной вытяжке. - Добавьте 0,5 мл PBS-T к образцам в микрофьюжных пробирках.
- Следуйте и повторяйте шаги 2.4.1 и 2.4.2 2-4x с PBS-T.
ПРИМЕЧАНИЕ: Выполнение большего количества стирок поможет уменьшить фоновый сигнал. Всего рекомендуется выполнить четыре промывки. - После последней промывки открутите образцы и удалите супернатант, убедившись, что гранула не нарушена.
- Вращайте образцы в микроцентрифуге с соответствующей скоростью (см. шаг 1.1.4). С помощью пипетки удалите супернатант, не нарушая гранулу.
- Подготовьте буфер гибридизации (HB) и промывайте нематод, как описано в шаге 1.5.
- Гибридизируйте зонд FISH с желаемой целевой последовательностью, как описано в шаге 1.6.
- Снимите зонд FISH и помойте нематод
- Подготовьте 1,1 мл промывочного буфера (WB) (900 мМ NaCl, 20 мМ Tris pH 7,5, 5 мМ ЭДТА, 0,01% SDS) на образец.
ПРИМЕЧАНИЕ: WB следует готовить свежим перед каждым использованием, чтобы избежать осадков. Однако общий буфер (900 мМ NaCl, 20 мМ Tris pH 7,5) может быть изготовлен заранее и храниться при комнатной температуре до тех пор, пока не понадобится буфер для стирки. Перед применением готовят 1,1 мл на образец общего буфера и добавляют ЭДТА к конечной концентрации 5 мМ, а SDS до конечной концентрации 0,01%. - Центрифугирование образцов с соответствующей скоростью (см. пункт 1.1.4). Удалите ГВ с помощью пипетки, при этом стараясь оставить гранулу нематоды нетронутой.
- Добавьте 100 мкл подготовленного ВБ к каждому образцу.
- Центрифугирование образцов с соответствующей скоростью (см. пункт 1.1.4). Снимите WB с помощью пипетки, при этом стараясь оставить гранулу нетронутой.
- Добавьте 1 мл подготовленного WB к каждому образцу.
- Инкубировать образцы в течение 1 ч при 48-56 °C в сухой ванне (или термомешателе при 48-56 °C при 1 200 об/мин). При инкубации в сухой ванне осторожно переворачивайте трубки каждые 15-20 мин.
ПРИМЕЧАНИЕ: Как правило, 48 °C используется в качестве стандартной температуры стирки; однако эту температуру, возможно, потребуется отрегулировать при наличии высокого фонового сигнала. Температура стирки часто на 2 °C выше, чем температура гибридизации. - Центрифугирование образцов с соответствующей скоростью (см. этап 1.1.4). Используя пипетку, снимите буфер для стирки, стараясь оставить гранулу нетронутой.
- Добавьте 100-500 мкл PBS-T к каждому из образцов. На этом этапе образцы могут храниться в PBS-T при 4 °C в течение недели, пока протокол не будет готов к продолжению.
- Подготовьте 1,1 мл промывочного буфера (WB) (900 мМ NaCl, 20 мМ Tris pH 7,5, 5 мМ ЭДТА, 0,01% SDS) на образец.
- Установите нематод, как описано в шаге 1.8.
Результаты
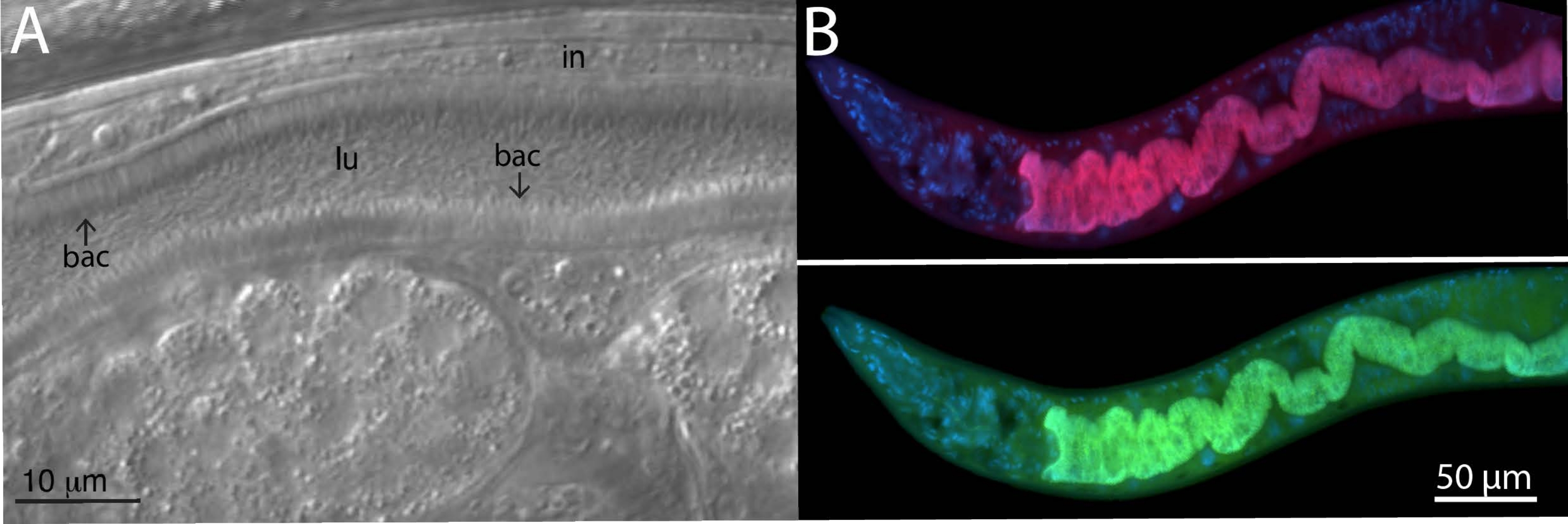
Для анализа бактерий микробиома на диких изолированных животных использовались специфические и универсальные зонды FISH для бактерий 16S. Дикий штамм Caenorhabditis tropicalis (JU1848) был взят из леса Нураг возле небольшой реки во Французской Гвиане из гниющих плодовпальмы 22. Под дифференциальным интерференционным контрастным микроскопом (ДВС-синдром) было обнаружено, что этот штамм нематоды колонизирован бактерией, которая, по-видимому, направленно прилипает к кишечному эпителию (рисунок 2A). Затем JU1848 был селективно очищен для устранения других микробных загрязнителей и обогащения для желаемой прилипшей бактерии23. С помощью универсального метода ПЦР бактерия была идентифицирована как новый вид в классе альфапротеобактерий. Зонд FISH, помеченный Cal Fluor Red 610, был затем разработан специально для последовательности 16S рРНК этой бактерии, чтобы обеспечить флуоресцентную визуализацию колонизации в пределах C. tropicalis (рисунок 2B). Универсальный 16S рРНК FISH зонд, способный связывать многие виды бактерий (EUB338), был помечен 6-карбоксифлуоресцином (FAM) и также был добавлен в этот образец. Зеленые и красные флуоресцентные сигналы полностью перекрываются, предполагая, что большинство бактерий, колонизирующих кишечник, являются прилипшими бактериями альфапротеобактерий. Эти животные были зафиксированы в PFA перед окрашиванием.
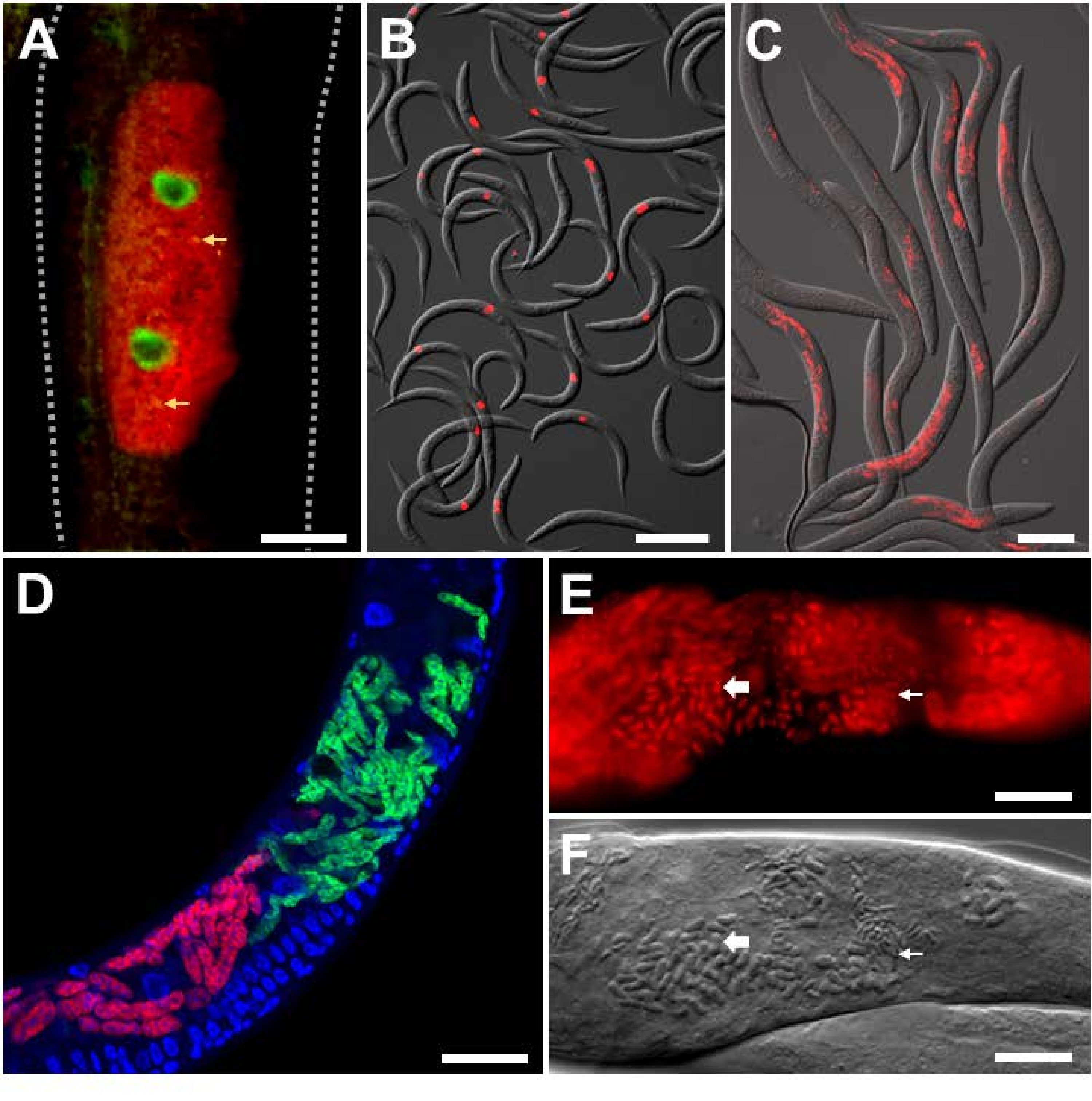
Для анализа экспериментальной инфекции в лаборатории с внутриклеточными патогенами известной идентичности на C. elegans с фоном дикого типа использовали вирус Орсе и микроспоридиан-специфические зонды FISH. Вирус Орсе является вирусом РНК с положительной цепью из семейства Nodaviridae и единственным природным вирусным патогеном, обнаруженным у C. elegans. Двудольный геном РНК вируса Орсе состоит из сегментов РНК1 и РНК2, и были разработаны зонды FISH, нацеленные на оба этих сегмента (рисунок 3A,B)9,18. В кишечнике вирусная РНК воспринимается гомологом RIG-I DRH-124, который необходим для активации транскрипционной защитной программы под названием Intracellular Pathogen Response (IPR)25,26,27. Транскрипция противовирусных генов IPR, по меньшей мере, частично контролируется фактором транскрипцииZIP-1 21. Здесь экспрессия ZIP-1::GFP локализована в кишечных ядрах клеток, которые показывают положительное окрашивание вируса Орсе FISH в цитоплазме (рисунок 3A)21. Показано, что несколько животных, окрашенных ORSAY-specific FISH, указывают силу этого сигнала для легкой количественной оценки (рисунок 3B). Животные, показанные на рисунке 3A,B, были зафиксированы в PFA.
Микроспоридный паразит под названием Nematocida parisii, что означает нематода-убийца из Парижа, является облигатным внутриклеточным патогеном кишечника. Было использовано несколько зондов FISH, которые маркируют 18S рРНК N. parisii, в том числе флуоресцентно помеченные зонды MicroA и MicroB. Показано, что несколько животных, окрашенных MicroB FISH, указывают силу этого сигнала для легкой количественной оценки (рисунок 3C). Кроме того, C. elegans инфицирован другими близкородственными микроспоридиями. Коинфекцию N2 с N. parisii и родственным N. ausubeli можно отличить с помощью этого протокола FISH путем разработки видоспецифичных зондов FISH, которые конкурируют друг с другом за связывание с дивергентной областью на 18S рРНК (рисунок 3D)28. В этом примере зонд N. parisii FISH имеет идеальное спаривание оснований с РРНК N. parisii 18S, но несоответствие 7 bp с N. ausubeli 18S рРНК. Обратное верно для зонда N. ausubeli. Таким образом, каждый видоспецифический зонд FISH будет конкурировать за привязку к родственным видам 18S над неродственными видами. Кроме того, использование DAPI для окрашивания ядер позволяет лучше локализовать инфекцию в контексте всего животного, особенно для кишечника, который имеет большие, легко идентифицируемые ядра. Рисунок 3C,D содержит животных, которые были зафиксированы в PFA. Более поздние инфекции N. parisii приводят к развитию меронтов в споры. Чтобы визуализировать споры N. parisii, животные должны быть зафиксированы в ацетоне, поскольку он проникает в споровую стенку лучше, чем PFA (рисунок 3E, F) 8. Полученное окрашивание FISH демонстрирует мелкие и крупные палочковидные структуры, которые, вероятно, соответствуют спорам N. parisii, которые окрашены специфическими для N. parisii зондами красного цвета.

Рисунок 1: Визуальное представление протокола FISH. Создано с помощью Biorender.com. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2: Окрашивание FISH дикого штамма C. tropicalis JU1848, колонизированного прилипшими бактериями в кишечнике. (A) Изображение Номарски, изображающее тысячи тонких бактерий бацилл (bac), направленно связывающихся с кишечником (in) JU1848, создавая волосоподобный фенотип в просвете (lu). Эта фигурная панель адаптирована из Morgan, E. et al. (2021)23. (B) Окрашивание FISH JU1848, зафиксированное в PFA, с использованием зонда с красной маркировкой (b002_16S_A-CF610), предназначенного для нацеливания на последовательность 16S рРНК прилипшей бактерии (сверху) и универсального зонда FISH с зеленой маркировкой (EUB338-FAM), предназначенного для нацеливания на 16S бактерий (снизу). Окрашивание Ядер хозяина по DAPI показано синим цветом. Последовательности зондов см. в таблице 1 . Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3: Окрашивание FISH C. elegans, инфицированных внутриклеточными патогенами. (A,B) ОКРАШИВАНИЕ FISH C. elegans , экспрессирующих ZIP-1::GFP и инфицированных вирусом Orsay, которые были зафиксированы PFA перед окрашиванием для сохранения сигнала GFP. Зонды Orsay 1 Red и Orsay 2 Red использовались для окрашивания патогенов. (A) Составное изображение состоит из объединенных красных и зеленых флуоресцентных каналов. Ядерная экспрессия ZIP-1::GFP индуцируется при вирусной инфекции Орсе и показана зеленым цветом. Автофлуоресценция из кишечных гранул показана желтым цветом и обозначена желтыми стрелками. Пунктирные линии очерчивают тело нематоды. Шкала = 25 мкм. (B) Составное изображение состоит из объединенных красных флуоресцентных и ОЭФ-каналов. Шкала bar = 200 мкм. (C,D) ОКРАШИВАНИЕ FISH диких C. elegans, инфицированных микроспоридиями, которые были зафиксированы в PFA. (C) Окрашивание рыбы диких C. elegans, инфицированных N. parisii и зафиксированных в PFA. Зонд MicroB-CF610 использовался для окрашивания патогенов. Составное изображение состоит из объединенных красных флуоресцентных и ОИСТ-каналов. Шкала bar = 100 мкм. (D) ОКРАШИВАНИЕ РЫБЫ диким типом C. elegans , коинфицированных N. parisii и N. ausubeli в кишечнике. Два патогена были совместно окрашены с использованием пары специфических зондов FISH, которые конкурируют за связывание с одной и той же областью 18S рРНК. N. parisii был окрашен с использованием MicroF-CF610 (красный), а N. ausubeli был окрашен с использованием MicroSp1A-FAM (зеленый). Окрашивание DAPI ядер хозяина видно синим цветом. Шкала бара = 25 мкм. (E) Окрашивание FISH ацетон-фиксированным диким типом C. elegans , инфицированным спорами N. parisii . MicroA-CF610 (красный) использовался для окрашивания (красный). Шкала бара = 15 мкм. (F) Изображение Номарского, изображающее споры N. parisii , видимые в (E). Шкала бара = 15 мкм. В (E) и (F) малые и большие стержневые структуры помечены маленькими и большими стрелками соответственно, которые соответствуют спорам N. parisii. Последовательности зондов см. в таблице 1 . Изображение, показанное в пункте (A), адаптировано из Lažetić, V. et al. (2022)21. Изображения, показанные в (B) и (C), адаптированы из Reddy, K. C. et al. (2019)26. Изображения, показанные в (E) и (F), адаптированы из Troemel, E. R. et al. (2008)8. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
{kind=link}
| Имя зонда | Специфичность зонда | Зондовый флуорофор | Последовательность зондов |
| EUB338-ФАМ | Бактериальный 16S (универсальный) | 5' 6-флуоресцеин (FAM) | GCTGCCTCCCGTAGGAGT |
| b002_16S_A-CF610 | Альфапротеобактерии 16S | Кал Флуор Красный 610 (CF610) | TGTACCGACCCTTAACGTTC |
| Орсе1 Красный | Вирус Орсе РНК1 | Кал Флуор Красный 610 (CF610) | GACATATGTGATGCCGAGAC |
| Орсе2 Красный | РНК вируса Орсе2 | Кал Флуор Красный 610 (CF610) | GTAGTGTCATTGTAGGCAGC |
| МикроА-CF610 | Nematocida parisii 18S | Кал Флуор Красный 610 (CF610) | CTCTGTCCATCCTCGGCAA |
| МикроБ-CF610 | Nematocida parisii 18S | Кал Флуор Красный 610 (CF610) | CTCTCGGCACTCCTTCCTG |
| МикроФ-CF610 | Nematocida parisii 18S | Кал Флуор Красный 610 (CF610) | AGACAAATCAGTCCACGAATT |
| МикроСп1А-ФАМ | Nematocida ausubeli 18S | 5' 6-флуоресцеин (FAM) | CAGGTCACCCCACGTGCT |
Таблица 1: Список последовательностей зондов FISH. Все зонды FISH были коммерчески приобретены с флуорофором, прикрепленным к 5'-концу (посредством пользовательского синтеза олигонуклеотидов; см. Таблицу материалов), а олигонуклеотиды были очищены обратнофазной ВЭЖХ.
Обсуждение
Дикие C. elegans естественным образом связаны с различными микробами. Исследователи могут использовать РНК FISH для обнаружения и идентификации этих микробов, а также для получения представления об их локализации в контексте целого животного. Микробы с желательными или интересными фенотипами могут быть идентифицированы с помощью этого метода, а затем выделены для дальнейшей характеристики и секвенирования. Обилие многочисленных бактериальных изолятов из диких C. elegans также может быть количественно определено с помощью РНК FISH29. Используя протокол, описанный здесь, также можно наблюдать за известными микроорганизмами внутри их хозяев и узнавать больше об их взаимодействиях. Важно отметить, что вирус Орсе и микроспоридия являются облигатными паразитами и не могут культивироваться независимо от хозяина, поэтому FISH является стандартным инструментом визуализации. Колонизация или инфекция также могут быть количественно определены с помощью РНК FISH с использованием нематод, выращенных на пластинах, засеянных желаемыми культивируемыми бактериями, представляющими интерес. В дополнение к окрашиванию микроорганизмов в кишечнике C. elegans, этот протокол может быть использован для других штаммов нематод, таких как C. tropicalis или Oscheius tipulae19,23.
Основным преимуществом протокола FISH является то, что он предлагает простой, быстрый и надежный метод окрашивания микробов, связанных с C. elegans. Изображения, полученные из окрашивания FISH, имеют высокое отношение сигнал/шум, которое достигается за счет использования зондов FISH, которые нацелены на обильную РНК SSU в образце. Поскольку обычно существует 30x или более высокие уровни рРНК, чем рДНК, большая часть сигнала от окрашивания FISH зондами, нацеленными на рРНК, обусловлена рРНК, а не рДНК30. Кроме того, РНК FISH позволяет увидеть инфекцию или колонизацию в контексте всего животного. Эта визуализация облегчается путем совместного окрашивания ядер хозяина с DAPI и / или использования флуоресцентных штаммов C. elegans, чтобы лучше выделить локализацию инфекции или колонизации в образце. Например, микроспоридиан-специфический FISH был использован для определения тканевого тропизма Nematocida displodere с использованием панели штаммов C. elegans с экспрессией GFP в разных тканях20. Кроме того, этот протокол поддается изменениям, которые позволяют исследователям определить идеальные условия, подходящие для их конкретных потребностей (например, регулировка периода фиксации, повышение температуры гибридизации).
Одним из важнейших шагов в протоколе FISH является фиксация образцов. Инкубационный период после добавления фиксатора необходим для того, чтобы дать агенту время для проницаемости образца. Более длительное время инкубации не идеально подходит для образцов, содержащих трансгенные флуоресцентные белки из-за деградации белка PFA с течением времени. Для образцов, содержащих GFP, крайне важно определить оптимальное время фиксации, чтобы обеспечить пермеабилизацию, сохраняя при этом сигнал GFP.
FISH может быть использован для окрашивания бактерий, вирусов или микроспоридий у C. elegans. Тем не менее, лучший тип фиксирующего агента, используемого для FISH, зависит от образца и последующих требований. Этот протокол представляет раствор PFA в качестве основного фиксирующего агента для окрашивания бактерий и вирусов. Однако PFA недостаточен для визуализации микроспоридиевых спор, поскольку он не может проникнуть в споровую стенку. Для визуализации спор следует использовать ацетон. Хотя фиксация ПФА эффективна для маркировки FISH других жизненных стадий микроспоридий, включая спороплазмы, меронты и споронты. Наблюдаются другие основные различия между фиксацией ацетона и фиксацией PFA; ацетон более удобен тем, что образцы можно быстро хранить в морозильной камере после добавления, без необходимости промывки. Однако ацетон быстро убивает любой существующий GFP в трансгенном хозяине. PFA является предпочтительным фиксатором, если важно сохранить некоторые физиологические структуры в организме хозяина, поскольку животные, фиксированные ацетоном, по-видимому, более деградированы, что затрудняет идентификацию некоторых тканей. Поскольку образцы фиксированы, этот протокол FISH не позволяет в реальном времени визуализировать взаимодействия хозяина и микроба in vivo. Тем не менее, импульсно-погоня за инфекцией, за которым следует окрашивание образцов FISH в различные моменты времени, может позволить увидеть некоторую динамику микробной инфекции 19,20,31.
Другим важным шагом на протяжении всего протокола является тщательная промывка образцов до и после гибридизации. Перед гибридизацией, при сборе червей в микрофрагменные трубки, избыточные бактерии или другие микробы из пластин NGM могут переноситься с образцом червя. Три мойки с PBS-T являются стандартными; однако для достаточного удаления внешних микроорганизмов может потребоваться больше промывок, особенно при использовании сильно загрязненных, изолированных в дикой природе C. elegans. При просмотре установленных образцов после FISH может быть какой-то остаточный зонд FISH, который производит большое количество сигнала на заднем плане образца. Температура стирки и количество стирок важны для удаления лишнего и неспецифического связанного зонда. Для уменьшения фоновой флуоресценции можно выполнять две-три промывки по 1 мл ВБ каждые 30 мин, вместо одной промывки 1 мл ВБ в течение часа. Для различных зондов FISH может потребоваться разная температура стирки. Как правило, температура стирки на 2 °C выше температуры гибридизации, но она может быть увеличена, если слишком много фоновой флуоресценции (высокий шум).
Протокол FISH использует флуоресцентные зонды, предназначенные для нацеливания на видоспецифическую микробную РНК, но зонды FISH могут быть разработаны для других транскриптов с высокой копией. Другие датчики FISH могут иметь различные температуры плавления, поэтому этапы инкубации, возможно, потребуется выполнить при более высокой или более низкой температуре, чем описано. Окрашивание FISH может идентифицировать пространственное распределение микробной колонизации или инфекции внутри хозяина, что позволяет характеризовать взаимодействия хозяин-микроб и микроб-микроб. Одним из ограничений является то, что одновременно можно использовать только несколько обычных флуорофоров, что уменьшает количество различных микроорганизмов, которые могут быть обнаружены с помощью FISH одновременно. Это ограничивает его использование для сложных исследований микробиома у C. elegans. Однако многоцветная рРНК-таргетированная FISH использует зонды, помеченные неканоническими флуорофорами, которые могут увеличить количество различных меток микробных групп15. Другим ограничением является то, что трудно различить близкородственные виды, особенно бактерии, которые имеют очень похожие последовательности SSU. Однако экстремальное расхождение последовательностей между видами микроспоридий помогает облегчить их дифференциацию с помощью этого протокола (рисунок 3)32,33.
В целом, этот протокол FISH описывает метод обнаружения микроорганизмов в C. elegans. Это позволяет исследователям использовать прозрачную и генетически поддающуюся обработке модельную систему для обнаружения и количественной оценки колонизации и инфекции в контексте интактного животного, а также для идентификации уникального микробного поведения или морфологии внутри хозяина. Препринтовая версия этой рукописи была опубликована во время рецензирования34.
Раскрытие информации
У авторов нет конфликта интересов.
Благодарности
Спасибо доктору Мари-Анн Феликс за предоставление нам штаммов диких нематод. Эта работа была поддержана NSF в рамках гранта CAREER Grant 2143718 и Калифорнийским государственным университетом в рамках премии CSUPERB New Investigator Award для RJL, NIH в рамках R01 AG052622 и R01 GM114139 для ERT, а также стипендией Американской кардиологической ассоциации для VL.
Материалы
| Name | Company | Catalog Number | Comments |
| 10% SDS | Invitrogen | AM9822 | |
| Acetone | Fisher Scientific | A-11-1 | |
| Antifade mounting serum with DAPI (Vectashield) | Vectalab | NC9524612 | |
| EDTA | Fisher Scientific | S311-500 | |
| FISH probes (see Table 1) | LGC Biosearch Technologies | FISH probes were commercially purchased via custom oligonucleotide synthesis | |
| KCl | Fisher Scientific | P217 | |
| KH2PO4 | Fisher Scientific | P-286 | |
| Na2HPO4 | Fisher Scientific | S375-500 | |
| NaCl | Fisher Scientific | S-671 | |
| NH4Cl | Fisher Scientific | A-661 | |
| Paraformaldehyde | Electron Microscopy Science | 50-980-487 | CAUTION: PFA is a carcinogen. Handle appropriately |
| Thermal mixer | Eppendorf | 5384000020 | |
| Tris base | Fisher Scientific | BP152 | |
| Triton X-100 | Fisher Scientific | BP-151 | |
| Tween-20 | Fisher Scientific | BP337-500 |
Ссылки
- Pukkila-Worley, R., Ausubel, F. M. Immune defense mechanisms in the Caenorhabditis elegans intestinal epithelium. Current Opinion in Immunology. 24 (1), 3-9 (2012).
- Balla, K. M., Troemel, E. R. Caenorhabditis elegans as a model for intracellular pathogen infection. Cellular Microbiology. 15 (8), 1313-1322 (2013).
- Dimov, I., Maduro, M. F. The C. elegans intestine: organogenesis, digestion, and physiology. Cell and Tissue Research. 377 (3), 383-396 (2019).
- Bossinger, O., Fukushige, T., Claeys, M., Borgonie, G., McGhee, J. D. The apical disposition of the Caenorhabditis elegans intestinal terminal web is maintained by LET-413. Developmental Biology. 268 (2), 448-456 (2004).
- Szumowski, S. C., Botts, M. R., Popovich, J. J., Smelkinson, M. G., Troemel, E. R. The small GTPase RAB-11 directs polarized exocytosis of the intracellular pathogen N. parisii for fecal-oral transmission from C. elegans. Proceedings of the National Academy of Sciences. 111 (22), 8215-8220 (2014).
- Samuel, B. S., Rowedder, H., Braendle, C., Félix, M. A., Ruvkun, G. Caenorhabditis elegans responses to bacteria from its natural habitats. Proceedings of the National Academy of Sciences. 113 (27), 3941-3949 (2016).
- Zhang, F., et al. Caenorhabditis elegans as a model for microbiome research. Frontiers in Microbiology. 8, 485 (2017).
- Troemel, E. R., Félix, M. -. A., Whiteman, N. K., Barrière, A., Ausubel, F. M. Microsporidia are natural intracellular parasites of the nematode Caenorhabditis elegans. PLoS Biology. 6 (12), 2736-2752 (2008).
- Felix, M. A., et al. Natural and experimental infection of Caenorhabditis nematodes by novel viruses related to nodaviruses. PLoS Biology. 9 (1), 1000586 (2011).
- Osman, G. A., et al. Natural infection of C. elegans by an oomycete reveals a new pathogen-specific immune response. Current Biology. 28 (4), 640-648 (2018).
- Zhang, G. A large collection of novel nematode-infecting microsporidia and their diverse interactions with Caenorhabditis elegans and other related nematodes. PLOS Pathogens. 12 (12), 1006093 (2016).
- Clark, L. C., Hodgkin, J. Commensals, probiotics and pathogens in the Caenorhabditis elegans model. Cell Microbiology. 16 (1), 27-38 (2014).
- Dirksen, P., et al. CeMbio - The Caenorhabditis elegans microbiome resource. G3. Genes, Genomes, Genetics. 10 (9), 3025-3039 (2020).
- Berg, M., et al. Assembly of the Caenorhabditis elegans gut microbiota from diverse soil microbial environments. The ISME Journal. 10 (8), 1998-2009 (2016).
- Michael, L., Markus, S., Petra, P., Holger, D. A multicolor fluorescence in situ hybridization approach using an extended set of fluorophores to visualize microorganisms. Frontiers in Microbiology. 10, 1383 (2019).
- Fuchs, B. M., et al. Flow cytometric analysis of the in situ accessibility of Escherichia coli 16S rRNA for fluorescently labeled oligonucleotide probes. Applied and Environmental Microbiology. 64 (12), 4973-4982 (1998).
- O'Neil, D., Glowatz, H., Schlumpberger, M. Ribosomal RNA depletion for efficient use of RNA-seq capacity. Current Protocols in Molecular Biology. 103 (1), 4-19 (2013).
- Franz, C. J., et al. Santeuil and Le Blanc viruses primarily infect intestinal cells in Caenorhabditis nematodes. Virology. 448, 255-264 (2014).
- Tran, T. D., Ali, M. A., Lee, D., Luallen, R. J. Bacterial filamentation as a mechanism for cell-to-cell spread within an animal host. Nature Communications. 13 (1), 1-11 (2022).
- Luallen, R. J., et al. Discovery of a natural microsporidian pathogen with broad tissue tropism in Caenorhabditis elegans. PLOS Pathogens. 12 (6), 1005724 (2016).
- Lažetić, V., et al. et al The transcription factor ZIP-1 promotes resistance to intracellular infection in Caenorhabditis elegans. Nature Communications. 13 (1), 1-16 (2022).
- Félix, M. -. A., et al. Species richness, distribution and genetic diversity of Caenorhabditis nematodes in a remote tropical rainforest. BMC Evolutionary Biology. 13 (1), 10 (2013).
- Morgan, E., Longares, J. F., Félix, M. A., Luallen, R. J. Selective cleaning of wild Caenorhabditis nematodes to enrich for intestinal microbiome bacteria. Journal of Visualized Experiments. (174), e62937 (2021).
- Sowa, J. N., et al. The Caenorhabditis elegans RIGI homolog DRH-1 mediates the intracellular pathogen response upon viral infection. Journal of Virology. 94 (2), 01173 (2020).
- Bakowski, M. A., et al. Ubiquitin-mediated response to microsporidia and virus infection in C. elegans. PLOS Pathogens. 10 (6), 1004200 (2014).
- Reddy, K. C., et al. Antagonistic paralogs control a switch between growth and pathogen resistance in C. elegans. PLoS Pathogens. 15 (1), 1007528 (2019).
- Reddy, K. C., et al. An intracellular pathogen response pathway promotes proteostasis in C. elegans. Current Biology. 27 (22), 3544-3553 (2017).
- Balla, K. M., Lažetić, V., Troemel, E. R. Natural variation in the roles of C. elegans autophagy components during microsporidia infection. PloS One. 14 (4), 0216011 (2019).
- Dirksen, P., et al. The native microbiome of the nematode Caenorhabditis elegans: gateway to a new host-microbiome model. BMC Biology. 14 (1), 38 (2016).
- Fu, R., Gong, J. Single cell analysis linking ribosomal (r)DNA and rRNA copy numbers to cell size and growth rate provides insights into molecular protistan ecology. The Journal of Eukaryotic Microbiology. 64 (6), 885-896 (2017).
- Willis, A. R., et al. A parental transcriptional response to microsporidia infection induces inherited immunity in offspring. Science Advances. 7 (19), (2021).
- Cuomo, C. A., et al. Microsporidian genome analysis reveals evolutionary strategies for obligate intracellular growth. Genome Research. 22 (12), 2478-2488 (2012).
- Reinke, A. W., Balla, K. M., Bennett, E. J., Troemel, E. R. Identification of microsporidia host-exposed proteins reveals a repertoire of rapidly evolving proteins. Nature Communications. 8 (1), 14023 (2017).
- Rivera, D. E., Lažetić, V., Troemel, E. R., Luallen, R. J. RNA fluorescence in situ hybridization (FISH) to visualize microbial colonization and infection in the Caenorhabditis elegans intestines. bioRxiv. , (2022).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены