
Bu içeriği görüntülemek için JoVE aboneliği gereklidir. Oturum açın veya ücretsiz deneme sürümünü başlatın.
Method Article
Bu Makalede
Özet
Kesik açık vazelin boşluk yaklaşım hızlı kanal kinetiği yüksek çözünürlüklü Xenopus oositlerde eksprese edilen voltaj-bağımlı iyon kanallarının iyonik ve yolluk akımlarının düşük ses kayıtları elde etmek için kullanılır. Küçük değişiklikler ile, voltaj kıskaç fluorometri kesme açık oosit protokolüne akuple edilebilir.
Özet
Kesik açık oosit Vaseline boşluk (COVG) voltaj kelepçe tekniği oositlerde heterolog iyon kanalları, elektrofizyolojik ve kinetik özelliklerinin analizi sağlar. Kesim açık kurulumdan Kayıtlar düşük büyüklüğü yolluk akımları, hızlı iyonik akım aktivasyonu ve deaktivasyonunu çözmek için özellikle yararlıdır. Iki elektrotlu voltaj kelepçe (TEVC) tekniği fazla temel faydaları artış kelepçe hızı, gelişmiş bir sinyal-gürültü oranı ve hücre içi ve hücre dışı ortamı ayarlama yeteneği de bulunmaktadır.
Burada, kesme açık setup ve protokol hem de voltaj kelepçe fluorometri özelliği eklemek için gerekli olan değişiklikleri göstermek için, (HNA V 1.5), Xenopus oositlerde eksprese edilen insan kardiyak sodyum kanalı kullanır.
Böyle HNA V 1.5 gibi hızlı aktive iyon kanalları, özellikleri, tam olarak hangi de, TEVC kullanılarak oda sıcaklığına yakın çözülemezoosit membran saat tamamı voltaj kontrolü zorlaştırır, kenetlenir. Bununla birlikte, kesme, açık teknik, hücre zarının sadece küçük bir kısmının izolasyon patch clamp teknikleriyle ilişkili kanal halsiz önlerken doğru hızlı kinetiği kaydetmek için gerekli olan hızlı bir sıkma sağlar.
COVG teknik, iyon kanalı kinetik ve elektrofizyolojik özellikleri ile bağlantılı olarak daha fazla protein, hücre dışı hareket uygulanır fluoroforlar, genetik olarak kodlanmış floresan proteinleri sokulması, ya da doğal olmayan amino asitlerin dahil sistein konjügasyonu ile izlenen voltaj kelepçe fluorometri kullanılarak deneye tabi tutulabilir ilgi 1'in bölgesine. Bu, ek veri flüoresan molekülü çevreleyen mikro-değişimlere ile proteinin voltaj bağımlı konformasyonel yeniden düzenlemeleri ile ilgili kinetik bilgi verir.
Giriş
Uzman voltaj kenetleme teknikleri, kontrol edilen membran potansiyelleri iyonik akımların kayıt izin verir. Yaygın iki elektrotlu voltaj kelepçe (TEVC) kullanılmış ve yama kelepçe teknikleri çok iyon kanallarının özellikleri hakkında güvenilir elektrofizyolojik bilgi sağlar. Ancak, bu yöntemlerin her ikisi de hızlı voltaj-kapılı sodyum kanalları ve bu tür Xenopus oosit gibi zarlarında diğer hızlı aktive kanallar için güvenilir verilerin ele geçirilmesini önlemek dezavantajları var. Bezanilla ve Stefani laboratuvarları dolayısıyla oosit 2 için kesim açık Vazelin boşluk gerilim kelepçe tekniği (COVG) geliştirdi. Bu teknik, Na +, K + ve Ca 2 + channels 3-8 kaydetmek için yaygın olarak uygulanmaktadır.
COVG kayıt sırasında, bir heterolog protein ifade oosit zarı üç bölgeye ayrılmıştır. Iyonik mevcut veriler olarak oositin üst bölgesinden kaydedilirüst bölgeyi çevreleyen banyo kolayca ve hızlı bir şekilde değiştirilebilir bir komut potansiyeli, kenetlenir. Orta bölge üst bölge 9 ile aynı potansiyele kenetlenir edilerek kaçak akımlara karşı koruma sağlar. Oosit açıklık (cut-açık) bir saponin çözeltisi ya da bir kanül aracılığıyla oluştuğu alt bölgesidir. Kimyasal ya da alt bölgesinde zarın manuel açma toprağa kenetlenir iç potansiyel kontrolünü sağlar ve alt bölme çözeltisi ile hücre iç sürekliliğini hale getirir. En iyi odasındaki çözelti değişimi dış çevresi değiştirir, oysa alt odasına çözümler perfüzyon, iç ortam özelliklerini ayarlayabilirsiniz. 
Şekil 1. Oosit Cut-Açık Gerilim-Kelepçe Banyo Kurulum Şeması. (A) Üstbirbirinden ayrılmış üç banyolarının görünümünde aşağı. COVG için bölmelerin boyutları şekil üzerinde görüntülenir. (B) testi pozisyonda hamam kurulum yan görünümü. resmi büyütmek için buraya tıklayın .
{kind=link}
COVG tekniğin avantajları, düşük akım gürültü (3 kHz 1 nA), dış ortamın iyonik kompozisyonu, iç ortam, hızlı zamanı çözünürlük (çürüme 20-100 mikro saniye olduğu zaman sabiti modüle yeteneği kontrolü, kapasite geçici), ve birkaç saat 9 stabil kayıtları. Dezavantajları özel ekipman gerektirir ve iki elektrotlu voltaj kelepçeleme (TEVC) 10 ile karşılaştırıldığında gerçekleştirmek için daha zor olmasıdır.
COVG yaklaşımı son derece özel ekipman ve karmaşık usul unsurları gerektirir iken, valu edinimi için izin verebilirmümkün elektrofizyolojik veriler. Böyle hızlı kinetik ve kuyruk akımları 4 ile akımlarını yolluk gibi bu veriler, kanal köhne dahil olmak üzere diğer voltaj sıkıştırma protokolleri ile ilgili bazı sorunları olmadan kaydedilebilir. COVG kurulum için küçük değişiklikler sıcaklık kontrolörleri ve gerilim kelepçe fluorometri (VKF) kullanımı için izin verebilirsiniz. COVG montaj içinde gerilim kelepçe fluorometri unsurların eklenmesi anda mevcut 11-13 kaydederken protein yapısal değişimleri izlemek için yeteneğini kazandıran veri çıkışını güçlendirir.
Protokol
1.. İlk Ekipman Kurulumu
- Elektrik ve mekanik gürültüyü önlemek için çevredeki Faraday kafesi olan bir titreşim izolasyon sistemi (örneğin bir hava tablo) sahne ve mikroelektrod manipülatör yerleştirin.
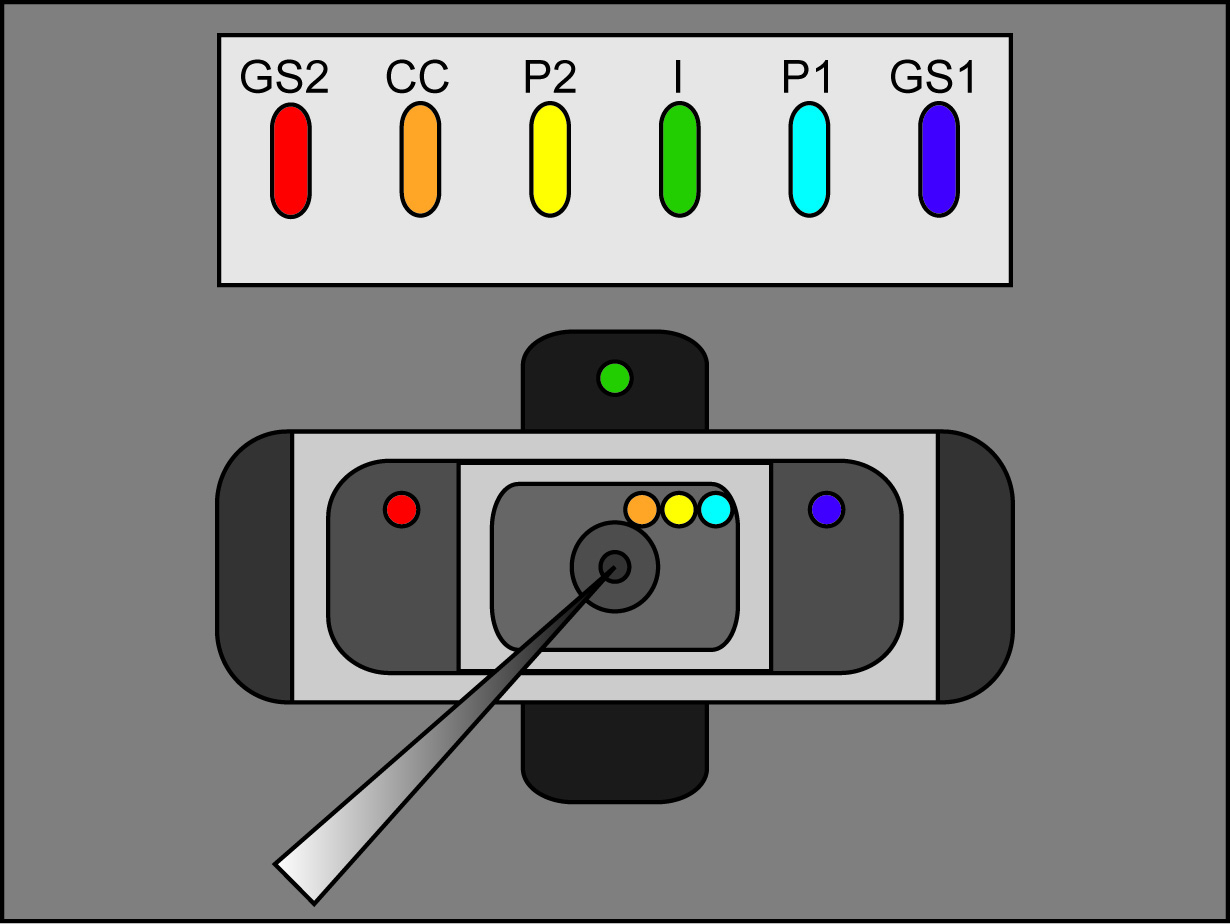
- 24 AWG tel altı-inç uzunluklarda altı Ag / AgCl pelet lehimleyin. Bu uzunlukları biri için (P1 bağlanabilir), bir ikinci tel birleşme yeri bir "Y" oluşturulur. Her kablonun ucunda amplifikatör ile birlikte bir altın BNC pimi, lehim.
- Banyo / Muhafız headstage (P1, P2, CC, GS1 ve GS2) 24 AWG tel lehimli beş Ag / AgCl pelet bağlayın. Bağlayın "Ben" "Ben" headstage ve V2 Headstage P1 çıkıyorsa ikinci tel Ag / AgCl pelet.
- Ekipman kılavuzlarının yönlere göre veri toplama ünitesine amplifikatörü bağlayın.
- Yerleştirin ve sıcaklık kontrolörü termosistörleri epoksi. Metal inşaat iskelelerinde bir delikten blok termistör KonuyuDoğrudan ısıtma / soğutma elemanının merkezi üzerinde d. Koyun ve çok yakın, ancak çözelti ile temas etmediğinden, sıcaklık iletken alt bölmeye gövdesinde açılan bir deliğe banyo termistör epoksi.
2. Yumurta ve Ön Hazırlık
- Böyle HNA V 1.5 gibi bir heterologously ifade kanalını kaydetmek için, (hSCN5a türetilmiş) mRNA sentezlemek ve çevresinde 4-5 gün Protokolü 4 gerçekleştirmeden önce bir Xenopus oosit içine enjekte edilir. HSCN5a için, en yüksek ifade 19 ° C sıcaklıkta 4-5 gün boyunca inkübe edildikten sonra elde edilen Ayrıntılı oosit ilgili talimatlar, mRNA hazırlanması, ve oosit enjeksiyon Richards ve Dempski 14 ve Cohen ve ark. 15'e bakınız.
- Protokol 4 başlamadan önce AgCl tel ve AgCl pelet klorürüne. Bunu yapmak için, en az 20 dakika boyunca çamaşır suyu içine bir tel ucunu ve granül yer ve sürece O / N olarak Peletler klorür edildikten sonraed, yapıştırıcı kullanılarak manifolduna yapıştırın.
Not: teller üzerinden akım sürüş de kablo ve pelet klorürüne kullanılabilir. Bu teknik chloridation hızını artırmak değil, aynı zamanda daha fazla ekipman gerektirir. Daha fazla talimat 16 gümüş teller klorid edilmesine Teknikleri bakın.
3. Agar Köprü Hazırlama
- Orta alev borosilikat kılcal borunun bir ucu kadar ısıtılarak en az altı agar köprüler olun. Kılcal boru ucu mavi alev üst kısmında olduğundan emin olun. Asıllarına hasar durumunda ekstra köprü yapmak.
- Kılcal tüp kadar ısıtılır sonra, tüp içinde 90 ° açılı bir bant için forseps kullanır. Pürüzsüz bir eğrilik ziyade ani bir köşesinde olan bir viraj hedefleyin ya da önemli ölçüde daha zor dolum yapar ve köprünün direncini arttırır cam, iç çapını azaltabilir.
- Ikinci bir 90 ° dirsek olunaynı yönde, birinci dirseğin kılcal boru 25 mm aşağı aynı adımları kullanarak.
Not: köprünün tam uzunluğu kadar uzun bir köprü büyüklüğü uyumlu olarak önemli değildir, ama sonuçta uzunlukları onlar üzerinde kullanılacak olan teçhizat için uygun olmalıdır. Köprü direnç uzunluğu ile orantılı olduğunu ve en aza indirgenmesi gerektiğini aklınızda tutun. - Kılcal tüpler soğutmalı sonra, yaklaşık 5 mm köprü "bacaklar" kesmek için elmas uçlu cam kesici kullanın.
- Agar 17 direncini azaltarak performansını artırmak için, üç "mevcut tedarik" köprü kılcal tüplere platin tel uzunlukları yerleştirin. Tüpün dışında maruz hiçbir tel olduğunu böylece herhangi bir aşırı platin tel kesti.
Not: platin nedeniyle yüksek maliyetine bağlı olarak, kırılmış köprüler almak ve ikinci bir yeniden tel. - Böyle bir mikropipet t olarak uygulanması ayrıca, ince uçlu ile kılcal boru içine platin tel itinip, tel, kılcal tüpün her iki ucunda da, camın daha kısa 1 mm olacak şekilde.
- HEPES ile tamponlanmış 1.2 g 1 M NMDG 100 ml olun. Bir pH metre kullanımı ve 7.4 'lük bir pH (~ 10 g) elde edilinceye kadar MES hidrat tozu eklenir. 7.4 'lük bir pH ulaşıldığında, pH elektrodu çıkarın. Bir depolama çözümü olarak tutmak için bir kenara çözeltisinden 40 ml ayarlayın.
- % 2-3 agar karışımı üretmek için granüle agar ekleyin. Ağar çözeltisi kadar karıştırın ve ısı çözülür ve net olduğunu. Aşırı ısınmasına veya aşırı viskoz ve köprüler zor olacak dolgu olacak gibi çözüm kaynatın etmeyin.
- Yeni bir behere agar çözeltisi hareket ve küçük bir karıştırma çubuğu ilave edin. Isıtma ve bir orta hızda karıştırmaya devam edin.
- Kılcal köprüleri yukarı bakacak bacakları ile bir seferde ekleyin. Zamanla köprüler agar ile dolacak. Alternatif olarak, küçük bir pipet ucuna bağlanmış bir şırınga aracılığıyla agar çözeltisi iterek köprüleri doldurun.
- Kabarcıklar bri var kezDGES, agar çözeltisinden köprüler almak ve kurutmak için bir kağıt havlu üzerinde köprü yerleştirin. Rezidüel kabarcıkları ile herhangi köprüler kabarcık çıkışını kolaylaştırmak için forseps ile ajite olabilir.
Not: kabarcıkları ile kurulmuş agar tamamen kaynar su içine batırılması ile köprüler kaldırılabilir. Agar çıkarıldıktan sonra, kalan artık suyun ayrılması için bir vakum hattı kullanır. Köprü sonra agar tedavisi için yeniden kullanılabilir. - Kuru köprüler aşırı agar çıkarın. 40 ml rezerv çözeltiye, 60 ml su eklenir ve çözelti içinde depolama köprüleri yerleştirin.
4. Kesim açık Rig Hazırlık
- Sıcaklık kontrol için su kaynağı ve sıcaklık kontrol ardından güç anahtarını açın. Banyo sıcaklığı belirtilen sıcaklığı (19 ° C) ulaşana kadar bekleyin.
- 0.2-0.5 MQ dirence bir Mikroelektrot çektirmesi ile borosilikat kılcal borudan Mikroelektronlar çekin.
Not: İndirme pipette direnç sıkma hızını artırır. Bununla birlikte, daha düşük direnç pipetler oositin zarar olasılığı daha yüksektir. Deney her uygulama için en iyi pipet direnç değerini belirlemek için gereklidir. - İç çözeltisi 50 ml kuru saponin 0.125 g karıştırılarak saponin çözeltisi hazırlayın. Bu,% 0.25 'lik çözeltinin yol açacaktır. Karıştırmak için hafifçe ters.
- Bir mikroskop altında, bir çok ince uçlu bir nesne ile orta bölmesi ve üst bölmenin alt tarafının üst tarafındaki deliğin kenarı etrafında vazelin küçük bir miktar uygulanır.
Not: vazelin "halka" delik üzerinde yerinde oosit tutmaya yardımcı olur ve contanın oluşumuna yardımcı olur. Ancak, çok fazla Vazelin tuzak kabarcıklar olacak ve oosit yüzeyine ulaşmasını önlemek çözümleri. - Orada yuvalarından hiçbir taşma ancak pelet ve köprü bacak uçları koy böylece Ag / AgCl pelet tutan manifoldu yuvalara 3 M KCl çözeltisi eklekırmızı. Yuvaları arasındaki istenmeyen elektrik bağlantılarını önlemek için ekstra KCl damlacıkları temizlemek.
- Alt ve orta banyo odalarına dış çözüm ekleyin.
- Köprünün başına bir bacak, her yuvada böylece AgCl pelet manifoldunda yuvalara ağar köprüleri yerleştirin. Köprülerin diğer bacak daha sonra ilgili odalarına yerleştirilir olacak (P1, P2, CC top; GS1, GS2 orta (bekçi), ben altta). Platin tel köprüler GS2, P2, ve ben yuvaları emin olun.
Not: emin köprüler odasının banyosuna takmadan önce distile su ile ve tamamen kuru yıkanır. - Veri toplama sistemi ve PC'de açın. Kayıt yazılımı başlatın.
5. Kesme açık Prosedürü
- Bir oosit olmadan üst ve orta oosit odaları yükleyin. İki mecliste delikler üst üste kalmamak off-center üst odasını kaydırın. Dış solüsyonu ile tüm odaları doldurun ve ilgili odalar tüm elektrotları yerleştirin.
Not: odalarını takarken tüm yol aşağı üst odasını itmeyin. Çok küçük bir boşluk, iki oda arasında olduğundan emin olun. Merkez dışı bir düzenleme daha iyi bir hücrenin varlığını taklit etmek için yanma odası direncini arttırır. "Köprüleri dengeleme" olarak adlandırılan bu süreç, ağar köprüleri arasında homojensizliğin doğabilecek ofset potansiyelleri dengeler.- Dış komut hem kelepçelerini kapatın. Amplifikatör geçerli değerini kontrol edin. Sıfır akım banyo / Guard baş-aşamasında arkasında ofset küçük bir tornavida P ile ayarlayın.
- Aktif için amplifikatör banyo / koruma anahtarını ve sıfır akım elde etmek için ofset GS ayarlayın.
- Hem (<100 nA) sıfıra yakın makul kadar "aktif" ve "pasif" arasında tekrarlayın.
- En iyi odası çıkarın ve bir pipet pompası kullanılarak orta banyo odası içine bir oosit aktarın. Emin olun oocyte banyonun ortasındaki deliğin üzerine konumlandırılır.
Not: VCF için bir oosit hazırlarken, yukarıya doğru bakacak şekilde hayvan kutup (koyu taraf) ile bölmenin içine etiketli hücre yerleştirin. VCF yapılırken değilse Cep oryantasyon önemli değildir. - Oosit ve banyo yüzeyi arasında bir yalıtım oluşturmak için bir aspiratör kullanılarak alt banyosundan fazla dış çözüm çıkarın.
- Odası içindeki delik oositin üstünde ortalanacak şekilde oosit üzerinde En iyi banyo odası yerleştirin. Delikten üst banyosuna zarın sadece küçük bir bölümünü açığa çıkartacak şekilde bu oosit sıkıca basılı kadar yavaşça bölmenin aşağı doğru baskı uygulanır, bir baş parmak ve orta parmak kullanılması.
Not: Oosit üst bölmeden basınç altında çıkıntı olabilir. Bu oosit yırtılmasına neden olacak gibi, üst bölmeye aşırı kuvvet uygulamayın. En iyi bölmesinin çapraz köşesine yerleştirilen cımbız uçlarına Appl için parmak için bir alternatif olarak kullanılabiliry aşağı doğru basınç. - Neredeyse tam kadar alt ve üst hamam dış çözüm ekleyin.
- Şekil 2 (köprü yerleştirme) görüldüğü gibi, her bir banyo dış çözelti içine agar köprülerin serbest bacaklar yerleştirin. Her köprü onun doğru banyo yerde dinlenme emin olun. (I alt banyosu, orta banyosunda GS1 ve GS2 köprüler ve P1, P2 ve üst banyosunda CC köprülerde köprü). Aktif için amplifikatör banyo / koruma anahtarı.
Not: emin köprüler, kuyulardan ve kayıt odaları arasında 3 M KCl bağlantıları vardır. Dahası, kayıt odası çözümleri üzerinde yükseltilmiş, böylece emin köprüler modası vardır olun.
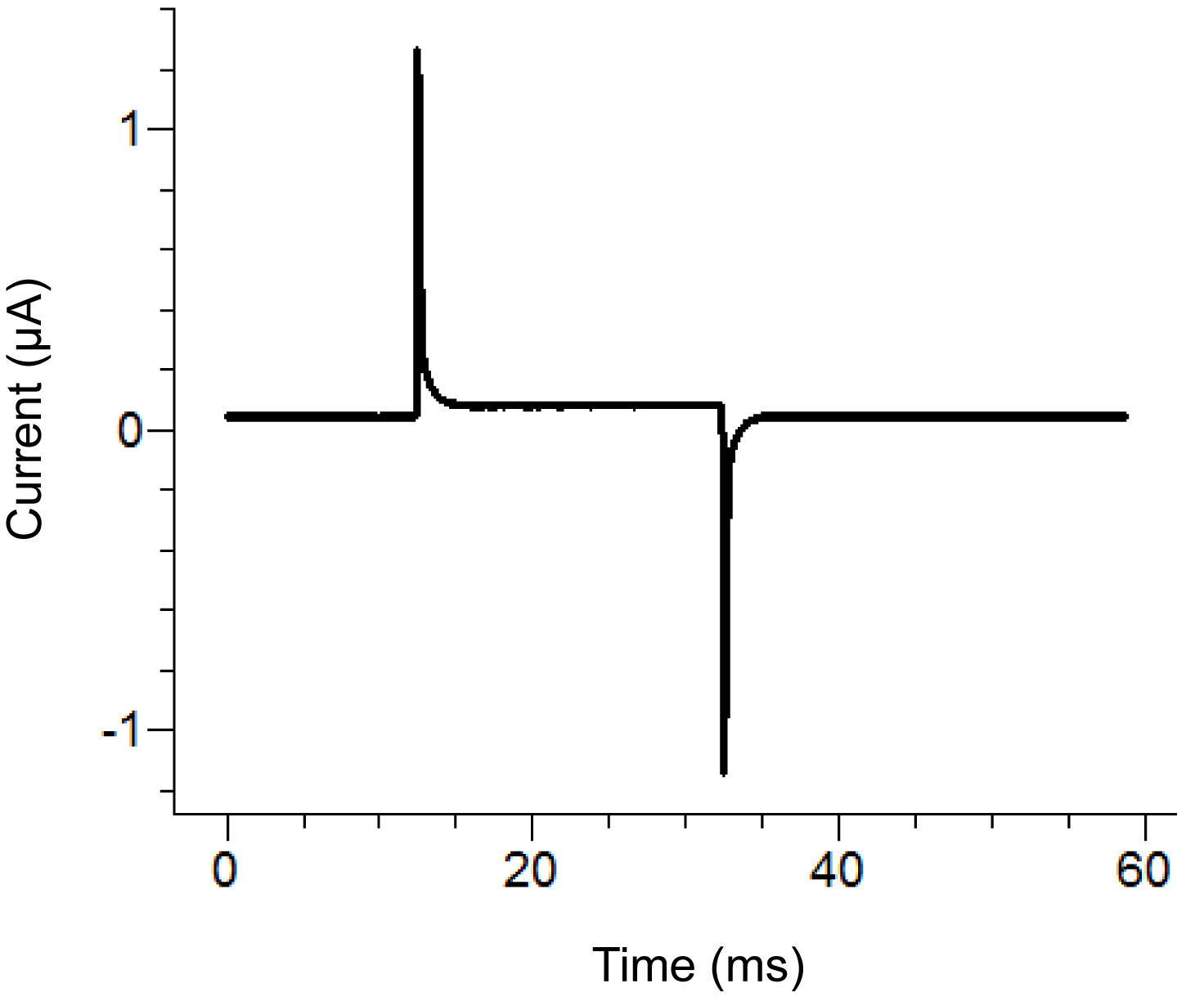
Şekil 2. Agar köprülerin serbest uçlarının Agar Bridge Kur konum diyagramı. Sıralama yerleriçeşitli banyolarında. resmi büyütmek için buraya tıklayın . - Kayıt yazılımı bir test protokolü başlatın. Nabız (pasif olarak banyo / koruma ile 0.3 MQ karşı gelen) daha büyük 100 nA bundan sonra banyo kapağının sıkılığı arttıracak olan bir 100 mV palsı uygulanarak üzerine, iki adet tepe arasındaki yatay bölümün dikey yer gösteriyorsa. İdeal test darbesinin bir örnek için Şekil 3'e bakın.
Not: Test protokolü banyo kapağı yeteri kadar sıkı ve tüm bileşenlerin doğru monte edilip edilmediğini görmek için bir gerilim darbe yayar. Alternatif olarak, amplifikatör test fonksiyonu kullanılabilir.
Şekil 3,. Yazılım Kayıt İdeal Testi Darbe. Testi darbeuygulanan protokole bağlı olarak yukarıdaki darbesine benzer görünmelidir. Tutma akımı (orta taban çizgisi) sıfıra yakın olmalıdır. resmi büyütmek için buraya tıklayın . - Alt banyosunda dış çözüm çıkarın ve saponin çözeltisi ile değiştirin. Saponin eklerken kabarcıkları oluşturarak önlemek için dikkatli olun. , En fazla değiştirilmesini sağlayan solüsyonu eklenirken, alt banyosunun ters ucunda emme uygulamak için.
- Saponin çözeltisi ilave edildikten sonra, yinelenen testi darbesi gözlemlemek. Test protokolü zirve azaltan veya kaybolursa sonra bu oosit altında bulunan bir baloncuk olduğu bir işaretidir. Bu durumda, tamamen saponin çözüm kaldırmak ve sonra değiştirin.
Not: Permeabilization genellikle taze saponin çözeltisi ile 30 saniye içinde tamamlanır. Tuzak kabarcıkları varsa çözümleri zorluk hücreye ulaşan olabilirveya foliküler katman bir kötü sindirilmiş oosit üzerinde kalırsa. Voltaj başak eğimi (çürüme zaman sabiti artış) azalır oosit (açılır) permeabilize edilmiştir.
Şekil 4. Testi Darbe Yumurta Permeabilization sırasında İzler. Test nabız protokolü seçilen izlerini 0.25% saponin çözüm alt oosit odasına tanıtıldı sonra. Izleri görülen çürüme zaman sabiti artış oosit geçirgenleştirmeden bir artış göstermektedir. resmi büyütmek için buraya tıklayın . - Hücre permeabilize sonra, saponin çözeltisi kaldırmak ve iç çözelti ile banyo doldurun. Test protokolünü durdurun.
Not: saponin erişim sağlar bilezarının nüfuz ile hücrenin iç, daha düşük banyo ve difüzyon ile oositin sitoplazması arasındaki iyon konsantrasyonlarının, dengeleme, çok yavaş bir süreçtir. Bu işlem, (Şekil 5) koşullarına bağlı olarak on dakika gerektirebilir. - Banyolarında çözümün herhangi bir yüksek düzeyleri kontrol edin ve bu kısa devreler ve düzensiz davranışlara neden olabilir gibi manifoldu kuyuları arasında KCI kristalize.
- Bir mikroelektrod içine 3 M KCl enjekte modifiye şırınga kullanın. Diğer parmaklar ile kuşaklama sırasında tek parmakla mikroelektrod birkaç kez hafifçe vurun.
Not: Bu adım mikroelektrot içinde herhangi hapsolmuş hava kabarcıklarını çıkarmak için gereklidir. - Açık mikroelektrot ucuna tel filamanın takarak mikromanipülatör koluna KCl dolu elektrot monte edin. Filament tutucu içine mikro elektrot ucunu itin ve elektrot gevşek olmadığından emin olun. Elektrot bağlayıcıyı sıkın.
Not: Matel normal çalışması için elektrot için daha AgCl kaplamaya sahip olduğundan emin olun ke. - Oosit hamam üzerindeki bir pozisyonda kolu salıncak ve daha fazla kol hareketini önlemek için kelepçeleri sıkın.
- Manipülatör düğmeleri kullanarak, banyo içine elektrod aşağı yürümek. Hiçbir test darbe uygulanır ve membran test özelliği bu noktada devreye olmadığından emin olun.
Not: elektrot ucu sıvı içine sokulur önce, V1-V2 voltaj kelepçesi üzerinde pozitif bir gerilim okuyacaktır. Elektrot ucu sıvı içine takıldığında, voltaj kelepçe üzerindeki gerilim metre sıfıra yakın bir değere değişmelidir. VCF kayıtları için, elektrot hedefi için odayı terk etmek oldukça sığ bir açıyla hücreyi yaklaşım gerekiyor. Merkez dışı elektrotla hücreyi Impaling, izole edilmiş zar yama kenarına yakın zamanda elektrot ile amacın çarpışmasını önlemek için yardımcı olur. - Elektrot yürürken durdurun. Ofset elektrot ayarlamaV1 butonuna basıp V1 ayarlayarak sıfıra V1 gerilimini azaltarak sıfıra potansiyel ofset. Buna ek olarak, V2 aynı ayar yapmak. Potansiyel farkı V1-V2 000 mV okumalısınız.
- Geri V1 geçin ve elektrot direnci ölçmek için Z-testi açın. Değeri yavaş yavaş düşer ve gerçek direnci yaklaşım olacaktır. 0.2-0.5 MQ bir direnç değeri için hedefliyoruz.
- Üst banyoda oosit görünür yama doğru elektrodu aşağı yürümeye devam. Mikroelektrot oosit çok yakın sonra, elektrot oosit girdiğinde V1-V2 okuma görmek için izlemek, mikroelektrot hücreye girdiğinde V1-V2 gerilimi negatif olur.
Not: bu noktada gösterilen değer hücrenin membran potansiyeli ve ifade edilen kanalları ve kullanılan çözeltilerin etkilenecektir. Çok mikroelektrot takma hücre zarına zarar verir. - Veri toplama protokolü açınkayıt yazılımı.
- Gerilim kelepçe üzerindeki kelepçe anahtarı çevirmek ve "ben" headstage bulunan düğmesini ayarlayarak komutu (örn. -100 mV) maç potansiyeli ayarlayın.
- Kapasitans ve direnç tazminat anahtarı çevirmek.
- "Testi" gerilim kelepçe "Komutlar" Bölgedeki açmak çevirin. Sinyali görselleştirmek için osiloskop kullanın. Osiloskopta kapasitif transientleri azaltmak için sinyal klima segmentinde Cm tazminat kolları ayarlayın. İlave ters zirveleri oluşursa veya tepe kayıt içine objeyi tanıtmak, hangi bir sigmoid eğrilik gelişmeye başlar noktasına zirveleri telafi fazla yapmayın.
- Kapasitans elle tatmin edici bir seviyeye düşürülmüş edildikten sonra, test şalteri kapatın.
- Kayıt yazılımı veri kayıt protokolü başlanılmalı.
{kind=link}
{kind=link}
{kind=link}
6. Temizleme
- Kayıtları arı varsan tamamlanmış, kelepçe ve banyo / koruma anahtarları da dahil olmak üzere gerilim kelepçe tüm çeşitli düğmelerini kapatın.
- Çeşitli banyoları agar köprüleri çıkarmak için forseps kullanın.
- En iyi banyo kaldırmak ve tüm hamam tüm çözümleri ve oosit aspire.
- Tüm banyoları yıkayın ve daha sonra bir vakum ile hamam aspire bir deiyonize su şişesi kullanın. Bu adım 3-5x tekrarlayın.
- Köprülerin kapalı kristalize KCI silin ve depolama çözümü köprüleri yerleştirin. Köprüler sürece doğru saklandığı gibi birçok hafta boyunca yeniden kullanılabilir.
- Birkaç kez manifold kuyulardan KCI çözeltisi aspire ve iyonu giderilmiş su ile durulama manifoldu.
- Sıcaklık kontrolü ve kayıt yazılımı dahil olmak üzere tüm çeşitli donanımları kapatın.
7. Gerilim Kelepçe fluorometri eklenmesi
- Önceden yayınlanmış bir vallahi protokol 16 E ile 6 Bölüm 1 adımları izleyinSite yönettiği Floresan Etiketleme ile yerinde Membran Proteinleri Konformasyonel Dinamikleri xamining: sayfayı görüntülemek için buraya tıklayın.
- 4X odak ayarlanmış bir VKF mikroskop kullanılarak yukarıda belirtilen COVG protokol 5,22 ile Bölüm 4'te adımları uygulayın.
Not: VCF kayıtları COVG ölçümlerde gerekli olandan daha büyük bir oosit banyo odaları gerektirir. (Özel VCF bölmelerin boyutları malzeme listesi yer almaktadır.) Daha büyük olan bu VCF bölmesi aynı zamanda objektif lens, mikroelektrot ve agar köprüler uyum sağlama kapasitesine sahiptir olmalıdır. Buna ek olarak, hayvan kutup (oositin karanlık tarafı) düşük arka plan VCF kayıtları için oda içinde yukarı doğru bakacak şekilde olması gerekmektedir. - Suda daldırma 40X objektif kullanarak odağa oositin üst kısmını getirin.
Not: 40X amacı için 4X geçiş belirli bir ge gerektirirkesim açık bileşenleri ve dikkatli dikkat ometry 40X objektif düşürücü elektrot, köprüler, ya da odaya vurmak şekilde değil. Üstelik nedeniyle üst bekçi artan hacmi, 40X objektif yerine ayarlandığında üst görevlisi o banyo hacmi orta bekçi ile bağlı olmadığından emin olun. - Oositin en üst hafif odak düzlemi üzerindedir, böylece, açıkta kalan oosit yüzeyinin çevresinde bir halka Focus.
Not: bakış alanı çok membran olup odası ile doldurulur, böylece xy düzleminde Ayar gerekli olabilir. XY için en kolay bir çeviri sahnede mikroskop yerleştirilmesi ile gerçekleştirilir. - Optik yoluna filtre küp taşı ve dedektör (diyot) için Mercekten ışık yolunu açın.
- VCF ışık kaynağı açın.
- Havai ışıklar, fiber ışık aydınlatıcı ve diğer ışık kaynaklarını kapatın.
Not: İdeal VKFkayıtları tamamen karanlık bir odada yapılmalıdır. - Kayıt yazılımı bir floresan protokol çalıştırın.
Sonuçlar
Şekil 4, görüntüler bir saponin çözeltisi olarak oositin geçirgenlikteki değişim oositin alt kısmına uygulanır. Nüfuziyet 5 saponin aşağıdaki difüzyon ile hücre içi çözelti değişim oranını göstermektedir Şekil. 20-40 dk bir kararlı duruma 2,18 gelmek için gereklidir.
Kayıt protokol oluşturulan Şekil 6A gösterisi izler. Şekil voltaj protokolü (ek) tepki olarak (P/-8 sızıntı ç?...
Tartışmalar
Kesim açık oosit Vazelin boşluk gerilim kelepçe tekniği hızlı veri çözünürlüğü sağlar, düşük gürültü, nispeten uzun protokolleri üzerinden 19 iç çözümü ve dış çözüm kompozisyon üzerinde kontrol ve istikrarlı kayıtları arttı. Bu avantajlar, ayrı Standart iki elektrotlu voltaj kelepçe ve yama kelepçe teknikleri bu teknik ayarlayın. Özel ekipman gerekli ve protokol nispeten zor olmasına rağmen, sistem optimize edildikten sonra, çok az sayıda sorunlar oluşur. Bu sody...
Açıklamalar
Yazarlar ifşa hiçbir şey yok.
Teşekkürler
Tüm St Louis Kardiyak Moleküler Mühendislik Lab Washington Üniversitesi'nden üyeleri. Bir Burroughs Bilimsel Arakesitte Fonu Kariyer Ödülü Hoşgeldiniz - 1010299 (JS).
Malzemeler
| Name | Company | Catalog Number | Comments |
| External Solution | Brand | Catalog Number | [Final], weight, or volume |
| N-methyl-D-glucamine (NMDG) | Sigma-Aldrich | M2004 | 25mM |
| MES Sodium Salt | Sigma-Aldrich | M5057 | 90mM |
| HEPES | Research Products International | H75030 | 20mM |
| Calcium hydroxide | Sigma-Aldrich | 239232 | 2mM |
| MES Hydrate | Sigma-Aldrich | M8250 | variable (pH to 7.4) |
| Internal Solution | |||
| N-methyl-D-glucamine (NMDG) | Sigma-Aldrich | M2004 | 105mM |
| MES Sodium Salt | Sigma-Aldrich | M5057 | 10mM |
| HEPES | Research Products International | H75030 | 20mM |
| Ethylene glycol-bis(2-aminoethylether)-N,N,N',N'-tetraacetic acid (EGTA) | Sigma-Aldrich | E4378 | 2mM |
| MES Hydrate | Sigma-Aldrich | M8250 | variable (pH to 7.4) |
| Depolarizing Solution | |||
| KCl | Sigma-Aldrich | 221473 | 110mM |
| Magnesium chloride | Sigma-Aldrich | M8266 | 1.5mM |
| Calcium Chloride | Caisson | C021 | 0.8mM |
| HEPES | Research Products International | H75030 | 10mM |
| Pipet Solution | |||
| KCl | Sigma-Aldrich | 221473 | 3M |
| Saponin Solution | |||
| Saponin | Sigma-Aldrich | 47036 | 0.125g |
| Internal Solution | See above | 50mL | |
| Agar Bridge Solution | |||
| N-methyl-D-glucamine (NMDG) | Sigma-Aldrich | M2004 | 100ml of 1M |
| HEPES | Research Products International | H75030 | 1.2g |
| MES Hydrate | Sigma-Aldrich | M8250 | variable (pH to 7.4) |
| Granulated Agar | Research Products International | A20250 | 3% |
| NMDG Storage Solution | |||
| NMDG, HEPES, MES Hydrate solution | see above | 40ml | |
| Water | 60ml | ||
| Name of Material/ Equipment | Company | Catalog Number | Comments/Description |
| High Performance Oocyte Clamp | Dagan | CA-1B | |
| Data Acquisition System | Axon CNS | Digidata 1440A | |
| Oscilloscope | Tektronix | TDS 210 | |
| Rack Power Filter | APC | G5 | |
| Heating/Cooling Bath Temperature Controller | Dagan | HCC-100A | |
| PC | Dell | Optiplex 990 | |
| pCLAMP 10.3 Voltage Clamp Software | Molecular Devices, LLC | pCLAMP10.3 | |
| TMC Vibration Control TableTop Platform | TMC | 64 SERIES | |
| TMC Vibration Control Air Table | TMC | 20 Series | |
| V1/I Electrode Data Collector | Dagan | part of CA-1B | |
| MX10L Micromanipulator | Siskiyou | MX10L | |
| Bath/Guard (I/V) Headstage (with appropriate connectors) | Dagan | part of CA-1B | |
| Microscope | Omano | OM2300S-JW11 | |
| Temperature Control Bath | Custom or Dagan | Custom or HE-204C | Custom chamber made from materials from Cool Polymers (D-series). Dagan also provides a prefeabricated stage (HE-204C). |
| Custom AgCl Pellet Container | Custom | Custom | Custom machined |
| Ag/AgCl electrode, pellet, 2.0 mm | Warner | E-206 | |
| External Oocyte Bath | Custom or Dagan | Custom or CC-1-T-LB | Custom machined or purchased from Dagan |
| Internal Oocyte Bath | Custom or Dagan | Custom or CC-TG-ND | Custom machined or purchased from Dagan |
| Capillaries for Agar Bridges and Pulled Electrodes | Warner | G150T-4 | |
| Rotatable Mounts for the Microscope, Micromanipulator, and Bath | Siskiyou | SD-1280P | |
| Fiber-Lite | Dolan-Jenner | LMI-600 | |
| Regular Bleach | Clorox | 470174-764 | |
| Xenopus laevis Oocytes | Nasco | LM535M (sexually mature females) | |
| 90 Na+ External Solution | See Solutions sheet | ||
| 10 Na+ Internal Solution | See Solutions sheet | ||
| 3 M KCL | See Solutions sheet | ||
| Saponin | Sigma-Aldrich | 47036 | |
| NMDG Storage Solution | See Solutions sheet | ||
| 5mL transfer pipets | SciMart | GS-52 | |
| Modified KCl electrode injector | BD | 309659 | Plastic syringe tip melted to allow for injection of solution into electrodes. Alternatively, a Microfil by WPI can be purchased. |
| Microvaccum | Custom | Custom | |
| Forceps | VWR | 63040-458 | |
| Oocyte Handling Tools (Pipette Pump) | VWR | 53502-222 | |
| Deionized Water Squirt Bottle | VWR | 16649-911 | |
| Vaseline Petroleum Jelly | Fisher Scientific | 19-086-291 | |
| Additional Materials Required for VCF Recordings: | |||
| VCF Microscope | Nikon | Eclipse FN1 | |
| Nikon CFI APO 40XW NIR Objective | Nikon | N40X-NIR | |
| X-Y Translator System for Fixed-Stage Upright Microscopes | Sutter Instruments | MT500-586 | |
| External VCF Oocyte Bath | Custom | Custom machined. The chamber dimensions are 2.7 x 1.9 x 0.4 cm. | |
| Internal VCF Oocyte Bath | Custom | Custom machined. The chamber dimensions are 1.6 x 1.6 x 0.4 cm. | |
| Modified Temperature Control Bath | Custom | Custom chamber made from materials from Cool Polymers (D-series). The chamber dimensions of the modified temperature controller bath are 2.7 x 1.9 x 0.3 cm for the horizontal chamber, and 1 x 2.5 x 0.5 cm for the vertical chamber. |
Referanslar
- Kalstrup, T., Blunck, R. Dynamics of internal pore opening in KV channels probed by a fluorescent unnatural amino acid. Proc. Natl. Acad. Sci. U.S.A. 110, 8272-8277 (2013).
- Stefani, E., Bezanilla, F. Cut-open oocyte voltage-clamp technique. Methods Enzymol. 293, 300-318 (1998).
- Muroi, Y., Chanda, B. Local anesthetics disrupt energetic coupling between the voltage-sensing segments of a sodium channel. J. Gen. Physiol. 133, 1-15 (2009).
- Stefani, E., Toro, L., Perozo, E., Bezanilla, F. Gating of Shaker K+ channels: I. Ionic and gating currents. Biophys. J. 66, 996-1010 (1994).
- Wang, S., Liu, S., Morales, M. J., Strauss, H. C., Rasmusson, R. L. A quantitative analysis of the activation and inactivation kinetics of HERG expressed in Xenopus oocytes. J. Physiolt. 502 (Pt 1), 45-60 (1997).
- Neely, A., Garcia-Olivares, J., Voswinkel, S., Horstkott, H., Hidalgo, P. Folding of active calcium channel beta(1b) -subunit by size-exclusion chromatography and its role on channel function. J. Biol. Chem. 279, 21689-21694 (2004).
- Silva, J. R., Goldstein, S. A. Voltage-sensor movements describe slow inactivation of voltage-gated sodium channels I: wild-type skeletal muscle. Na(V)1.4. J. Gen. Physiol. 141, 309-321 (2013).
- Silva, J. R., Goldstein, S. A. Voltage-sensor movements describe slow inactivation of voltage-gated sodium channels II: a periodic paralysis mutation in Na(V)1.4 (L689I). J. Gen. Physiol. 141, 323-334 (2013).
- Taglialatela, M., Toro, L., Stefani, E. Novel voltage clamp to record small, fast currents from ion channels expressed in Xenopus oocytes. Biophys. J. 61, 78-82 (1992).
- Clare, J. J., Trezise, D. J. . Expression and analysis of recombinant ion channels : from structural studies to pharmacological screening. , (2006).
- Cha, A., Zerangue, N., Kavanaugh, M., Bezanilla, F., Susan, G. A. . Methods in enzymology. 296, 566-578 (1998).
- Lakowicz, J. R. . Principles of fluorescence spectroscopy. 3rd edn. , (2006).
- Cha, A., Bezanilla, F. Characterizing voltage-dependent conformational changes in the Shaker K+ channel with fluorescence. Neuron. 19, 1127-1140 (1997).
- Richards, R., Dempski, R. E. Examining the conformational dynamics of membrane proteins in situ with site-directed fluorescence labeling. J. Vis. Exp. , (2011).
- Cohen, S., Au, S., Pante, N. Microinjection of Xenopus laevis oocytes. J. Vis. Exp. , (2009).
- Raynauld, J. P., Laviolette, J. R. The silver-silver chloride electrode: a possible generator of offset voltages and currents. J. Neurosci. Methods. 19, 249-255 (1987).
- Gagnon, D. G., Bissonnette, P., Lapointe, J. Y. Identification of a disulfide bridge linking the fourth and the seventh extracellular loops of the Na+/glucose cotransporter. J. Gen. Physiol. 127, 145-158 (2006).
- Pantazis, A., Olcese, R., Roberts, G. . Cut-Open Oocyte Voltage-Clamp Technique. In: Roberts G. (Ed.) Encyclopedia of Biophysics: SpringerReference. , (2013).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır