
Для просмотра этого контента требуется подписка на Jove Войдите в систему или начните бесплатную пробную версию.
Method Article
В этой статье
Резюме
Подход Вазелин разрыв отключения открытым используется для получения низких шумовых записи ионных и стробирования токов от потенциал-зависимых ионных каналов, выраженных в Xenopus ооциты с высоким разрешением быстрой кинетики канала. С небольшим изменением, зажим напряжение флюометрия может быть соединен с протоколом ооцитов вырезом открыт.
Аннотация
Ооцит Вазелин разрыв отключение открытой (CoVG) техника зажим напряжения позволяет для анализа электрофизиологических и кинетических свойств гетерологичных ионных каналов в ооцитов. Записи от установки бортового открытым особенно полезны для решения малых токов величина литниковые, быстрого ионной активации текущий и дезактивацию. Основные преимущества над двухэлектродной зажима напряжения (TEVC) техники включают повышенную скорость зажим, улучшение соотношения сигнал-шум, и способность модулировать внутриклеточной и внеклеточной среде.
Здесь мы используем человеческий канал сердечная натрия (HNA V 1.5), выраженную в Xenopus ооцитах, чтобы продемонстрировать настройку бортового открытым и протокол, а также изменения, которые необходимы, чтобы добавить зажим напряжение возможность флуорометрии.
Свойства быстрых активирующих ионных каналов, таких как HNA V 1,5, не могут быть полностью решены около комнатной температуре с использованием TEVC, в whicч полнота ооцита мембраны зажимается, что делает контроль напряжения трудно. Однако, в технике вырезом открыт, выделение только небольшой части клеточной мембраны позволяет быстро зажима, необходимого, чтобы точно записать быструю кинетику, предотвращая канала краткое изложение, связанное с методами патч зажима.
В сочетании с техникой CoVG, ионных каналов кинетики и электрофизиологические свойства могут быть дополнительно анализировали с помощью напряжения зажим флуорометрии, где движение белок отслеживается с помощью цистеина конъюгации внеклеточно, применяемых флуорофоров, вставка генетически кодируемых флуоресцирующих белков, или включения неприродных аминокислот в интересующей области 1. Это дополнительные данные дает кинетическую информацию о потенциал-зависимых конформационных перестроек белка через изменения в микросреды, окружающей флуоресцентные молекулы.
Введение
Специализированный методы напряжение зажимные разрешить запись ионных токов при контролируемых мембранных потенциалов. Широко используется двухэлектродной зажим напряжения (TEVC) и методы патч зажим обеспечивают надежную электрофизиологические информацию о свойствах многих ионных каналов. Тем не менее, оба этих метода имеют свои недостатки, которые препятствуют приобретению надежных данных для быстрых напряжения натриевых каналов и других каналов быстро активирующих в мембранах, таких как те, из Xenopus ооциты. В Безания и Стефани лаборатории следовательно разработана методика Вазелин разрыв зажим напряжения бортового открытым (CoVG) для ооцитов 2. Методика была широко применяться для записи, Na +, K + и Ca 2 + каналов 3-8.
Во время записи CoVG, гетерологичный белок, экспрессирующих ооцитов мембрана состоит из трех областей. Ионный ток данные записываются с верхней области ооцита какванна, окружающий верхнюю область крепится к командной потенциалом, который может быть легко и быстро изменить. Средней области защищает от токов утечки, будучи прикрепленными к тем же потенциалом, верхней области 9. В нижней области, где отверстие ооцитов (вырезать открыт) происходит с использованием раствора сапонина или канюли. Химические или ручное открывание мембраны в донной области позволяет управлять внутреннего потенциала, который крепится к земле, и оказывает внутрь клетки прилегает к нижней камере раствора. Перфузии решений в нижнюю камеру можно настроить свойства внутренней среды, в то время как обмен решение в верхней камере изменяет внешний антураж. 
Рисунок 1. Ооцитов Cut-Открыть Напряжение-зажим Ванна Схема установки. () ТопВвиду вниз из трех ванн, разделенных друг от друга. Размеры камер для CoVG отображаются на рисунке. (B) Вид сбоку установки ванны в положении тестирования. Нажмите здесь, чтобы увеличить изображение .
{kind=link}
Преимущества метода CoVG включают низкую текущую шум (1 нА при 3 кГц), контроль ионного состава внешней среды, в способности модулировать внутренний СМИ, разрешение быстро времени (20-100 мкс постоянная времени распада Емкость переходные), и стабильные записи в течение нескольких часов 9. К недостаткам можно отнести, что он требует специального оборудования и труднее выполнять по сравнению с двумя напряжения на электродах зажима (TEVC) 10.
В то время как подход CoVG требует высокой специализированное оборудование и сложные процедурные элементы, он может позволить для приобретения Valuспособные электрофизиологические данные. Эти данные, такие как стробирования токов с быстрой кинетики и хвостовые токи 4, могут быть записаны без некоторых проблем, связанных с другими протоколами напряжение зажима включая канала кратком изложении. Незначительные изменения в настройки CoVG может позволить для использования регуляторов температуры и зажим напряжение флуорометрии (VCF). Включение напряжения зажим элементов флуорометрии в сборке CoVG может увеличить выход данных присвоением способность контролировать белка конформационные изменения, одновременно записывая текущее 11-13.
протокол
1. Начальная настройка оборудования
- Поместите сцену и микроэлектродной манипулятор в системе вибрации изоляции (например таблицы воздуха) с окружающей клетки Фарадея для предотвращения электрического и механического шума.
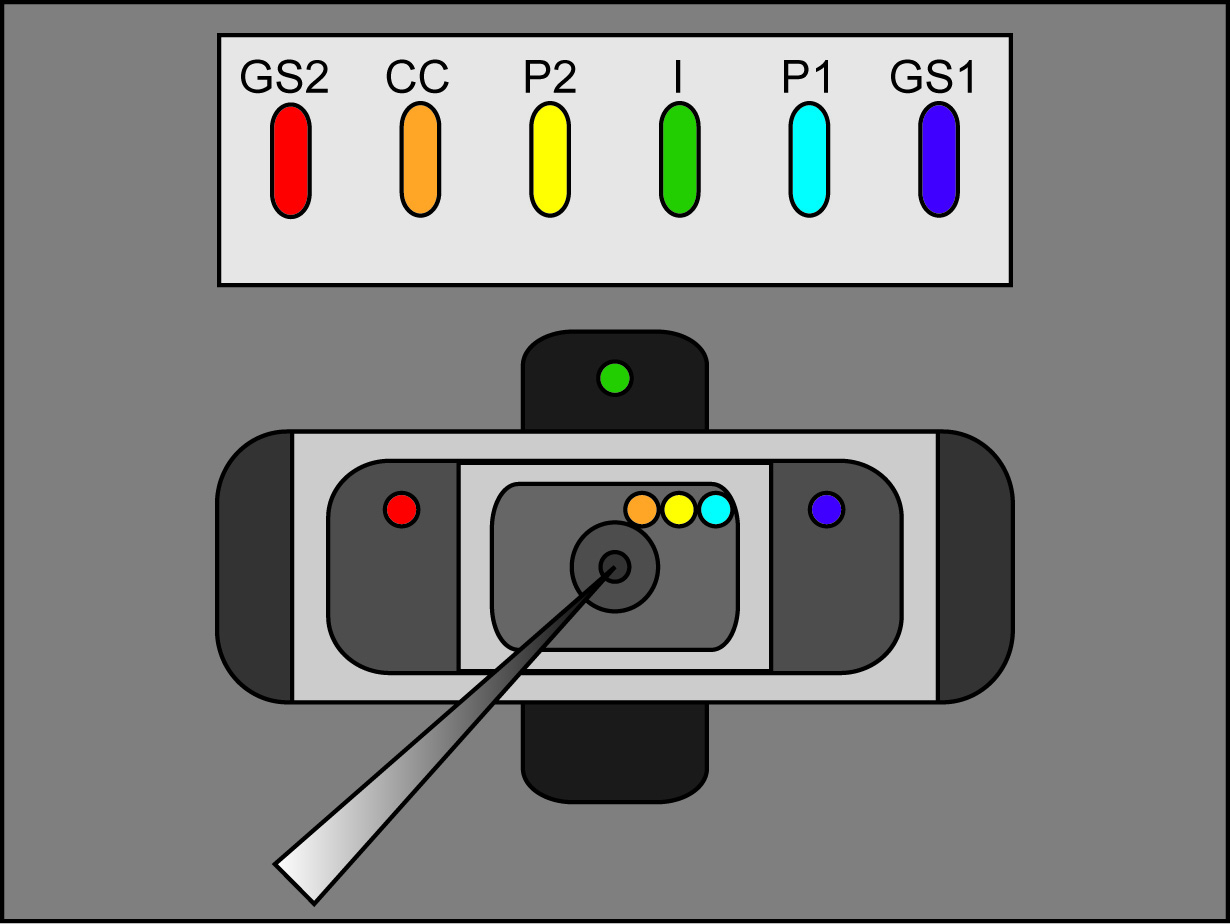
- Припой шесть Ag / AgCl гранул к длинам шестидюймовыми от 24 AWG провода. Для одного из этих длин (для подключения к P1), соединение в второй проволоки, чтобы сформировать "Y". На концах каждого провода припаять золотую BNC штифт, который входит с усилителем.
- Подключите пять Ag / AgCl гранулы спаянные до 24 AWG провода к headstage Ванна / Guard (P1, P2, CC, GS1 и GS2). Подключите «я» Ag / AgCl окатышей на "I" headstage и второго провода сходит Р1 к V2 headstage.
- Подключите усилитель с блоком сбора данных в соответствии с указаниями в руководствах к оборудованию.
- Поместите и эпоксидные контроллер термисторы температуры. Заправьте блок термистор через отверстие в металлической scaffolд непосредственно над центром элемента нагрева / охлаждения. Место и эпоксидные ванны термистор в отверстие, просверленное в корпусе температуры проведение нижнюю камеру в непосредственной близости, но не контактирование раствора.
2. Ооцитов и предварительной подготовки
- Для записи гетерологично выраженную канал например HNA V 1.5, синтезировать мРНК (производный от hSCN5a) и ввести его в яйцеклетку Xenopus около 4-5 дней, прежде чем выполнять протокол 4. Для hSCN5a, пик выражение после инкубации в течение 4-5 дней при 19 ° C. См. Ричардс и Dempski 14 и Коэн и др.. 15 Подробные инструкции по яйцеклетки, подготовка мРНК, и инъекции ооцитов.
- Хлорида провод AgCl и AgCl гранул перед началом протокола 4. Чтобы сделать это, поместите один конец проволоки и гранул в отбеливателя в течение не менее 20 мин и до тех пор, O / N. После того, как гранулы были хлоридноред, прикрепить их к коллектору с помощью клея.
Примечание: ток возбуждения через провода также можно использовать к хлориду провод и гранулы. Эта техника будет увеличить скорость хлорирование, но и требуют больше оборудования. См. Методы для хлорирующим серебряные проволочки для дальнейшего обучения 16.
3. Агар Мост Подготовка
- Сделать по меньшей мере шесть агар мосты, нагревая один конец боросиликатного капиллярной трубки в среднем огне. Убедитесь, что конец капиллярной трубки находится в верхней части голубого пламени. Сделать дополнительные мосты в случае повреждения оригиналов.
- После того, как капиллярная трубка нагрелась, использовать пинцет, чтобы сделать угол изгиба 90 ° в трубке. Цель для изгиба с гладкой кривизны, а не резким углу, или он может значительно уменьшить внутренний диаметр стакана, что делает более трудным заполнение и увеличивает сопротивление моста.
- Сделать второй 90 ° изгибв том же направлении 25 мм вниз капиллярной трубки от первого изгиба, используя те же самые шаги.
Примечание: Точная длина моста не имеет значения до тех пор, пока размер мост согласуется, но в конечном счете длины должны быть пригодны для установки они будут использоваться на. Имейте в виду, что сопротивление моста пропорционально ее длине и должны быть сведены к минимуму. - После того, как капиллярные пробирки остыли, использовать алмазный стеклорез, чтобы урезать "Эпический" моста примерно 5 мм.
- Вставьте длины платиновой проволоки в капиллярных трубках трех «токоподводящими« мостов для повышения производительности за счет снижения сопротивления в агар 17. Обрежьте излишки платиновой проволоки, так что нет провод подвергается снаружи трубки.
Примечание: В связи с высокой стоимостью платины, извлекать и повторно использовать любой провод от сломанных мостов. - Нажмите платиновую проволоку дальше в капиллярной трубке с острым концом реализации таких как микропипетки тIP так, что провод 1 мм короче, чем стекло на обоих концах капилляра.
- Сделать 100 мл 1 М NMDG буферизированные с 1,2 г HEPES. С помощью рН-метра и добавить MES гидратов порошка до рН 7,4 не будет достигнута (~ 10 г). Как только величина рН 7,4 достигнута, удалить рН электрод. Установите в сторону 40 мл раствора, чтобы держать в виде раствора для хранения.
- Добавить гранулированного агар для получения 2-3% агара смеси. Перемешать и тепла до раствора агара растворяют и ясно. Не перегревать или кипятят раствор, как он станет слишком вязкой и заполнения мосты будет трудно.
- Перемещение агар решение нового стакан и добавить небольшое мешалкой. Продолжить нагрева и перемешивания с умеренной скоростью.
- Добавить капилляра Мосты по одному с ног вверх. Со временем мосты наполнит агара. С другой стороны, заполнение мосты, нажав раствора агара с помощью шприца, присоединенного к небольшим наконечником пипетки.
- Как только там нет пузырьков в бриДГЭС, извлекать мосты от раствора агара и поместите мосты на бумажное полотенце, чтобы высушить. Любые мосты с остаточными пузырьков можно перемешивать щипцами для облегчения пузырь выход.
Примечание: Ставка агар с пузырьками может быть полностью удалена путем погружения мосты в кипящую воду. После того, как агар удален, использовать вакуумную линию для удаления остаточной воды. Bridges затем могут быть повторно использованы для лечения агара. - Удалите излишки агар с мостов, когда сухие. Добавить 60 мл воды в резервный раствор 40 мл и поместите мосты в растворе для хранения.
4. Вырезать открыть Рог Подготовка
- Включите источник воды для регулятора температуры, а затем выключатель питания на контроллер температуры. Подождите, пока температура ванны не достигнет заданной температуры (19 ° С).
- Потяните микроэлектродов из боросиликатного капиллярной трубки с микроэлектродной съемника к сопротивлению 0,2-0,5 МОм.
Примечание: снижение пипсСопротивление Ette повышает скорость зажима. Тем не менее, более низкие пипетки сопротивления, скорее всего, привести к повреждению яйцеклетки. Эксперименты требуется определить оптимальное значение сопротивления пипетки для каждого приложения. - Готовят раствор сапонина путем смешивания 0,125 г сухого сапонина с 50 мл внутреннего раствора. Это приведет к 0,25%-ного раствора. Переверните аккуратно перемешать.
- Под микроскопом рассекает нанести небольшое количество вазелина вокруг края отверстия на верхней стороне средней камеры и нижней стороне верхней камеры с очень остроконечного предмета.
Примечание: "пончик" вазелина поможет сохранить яйцеклетки в месте над отверстием и поможет в формировании печатью. Однако, слишком много Вазелин будут улавливать пузыри и предотвратить решения, достигающий поверхности яйцеклетки. - Добавить раствор 3 М KCl в коллектор слотов, имеющих Ag / AgCl гранул, так что нет переполнение из пазов но гранулы и концы мост ножные бухтакрасный. Очистка дополнительные капельки KCl для предотвращения нежелательных электрических соединений между слотами.
- Добавить внешнее решение для нижнего и среднего камер ванны.
- Поместите агара мосты в слоты в осадке коллектора AgCl так, чтобы одна нога за моста в каждом слоте. Другая нога из мостов в дальнейшем будут помещены в соответствующих камерах (P1, P2, CC лучших; GS1, GS2 средний (охранник), я снизу). Убедитесь, что платиновой проволоки мосты в GS2, P2, и слоты ввода.
Примечание: Убедитесь, мосты промывают дистиллированной водой и полностью высушить перед установкой в камеру ванной. - Включите систему сбора данных и ПК. Запустите программное обеспечение для записи.
5. Вырезать открыть Порядок
- Установите высшего и среднего камер ооцитов без яйцеклетки. Сдвиньте верхнюю камеру не по центру так, чтобы отверстия в двух палат не перекрываются. Заполните все камеры с внешнего решения и поставить все электроды в своих камерах.
Примечание: Не нажимайте верхнюю камеру все, вплоть при установке камеры. Убедитесь очень небольшой зазор остается между двумя камерами. Устройство не по центру увеличивает сопротивление системы камеры, чтобы лучше имитировать наличие клетке. Этот процесс, называемый "балансировки мосты", компенсирует смещения потенциалов, которые могут возникнуть в результате неоднородности среди агара мостов.- Выключите внешнюю команду и оба зажима. Проверьте текущее показание на усилителе. Отрегулируйте с помощью маленькой отвертки P зачете на задней панели ванна / охранник головной стадии к нулевом токе.
- Переведите переключатель ванна / охранник на усилителе к активным и отрегулируйте GS смещения для получения нулевой ток.
- Повторите между «активной» и «пассивной», пока оба не являются достаточно близко к нулю (<100 нА).
- Удалите верхнюю камеру и передавать ооцит в среднюю камеру ванны с помощью пипетки насос. Убедитесь, что oocyТе расположен над отверстием в центре ванны.
Примечание: При подготовке яйцеклетки для VCF, разместить меченых клеток в камеру с анимальной (темная сторона) вверх. Ориентация сотовый не имеет значения, если VCF не выполняется. - Удаление избыточного внешнего раствора из нижнего бане с помощью аспиратора, чтобы создать уплотнение между ооцита и поверхностью ванны.
- Поместите верхнюю камеру над ванной ооцита так, чтобы отверстие в камере по центру в верхней части ооцита. Использование большим и средним пальцами, медленно надавливать вниз по камере, пока не будет плотно прижимается ооцита так, чтобы открыть лишь небольшую часть мембраны к верхней ванны через отверстие.
Примечание: яйцеклетка может выпячиваться под давлением верхней камеры. Не прилагайте чрезмерных усилий к началу камеры, так как это вызовет ооцитов к разрыву. Пинцеты советы Расположенные диагональных углов верхней камеры может быть использована в качестве альтернативы пальцами, чтобы Applу понижательное давление. - Добавить внешнюю решение верхней и нижней ванны, пока они не почти полный.
- Поместите бесплатно ноги агара мостов в наружный раствор каждого ванной, как показано на рисунке 2 (размещения мост). Убедитесь, что каждый мост отдыхает в правильном месте ванной. (Я мост в нижней ванной, GS1 и GS2 мостов в средней ванной, и P1, P2, и CC мостов в верхней ванной). Переведите переключатель ванна / охранник на усилителе к активным.
Примечание: Убедитесь, нет 3 М KCl связи между мостами, их скважин, и запись камер. Кроме того, убедитесь, что мосты вылеплены так, чтобы они поднимаются над записи камерных решений.
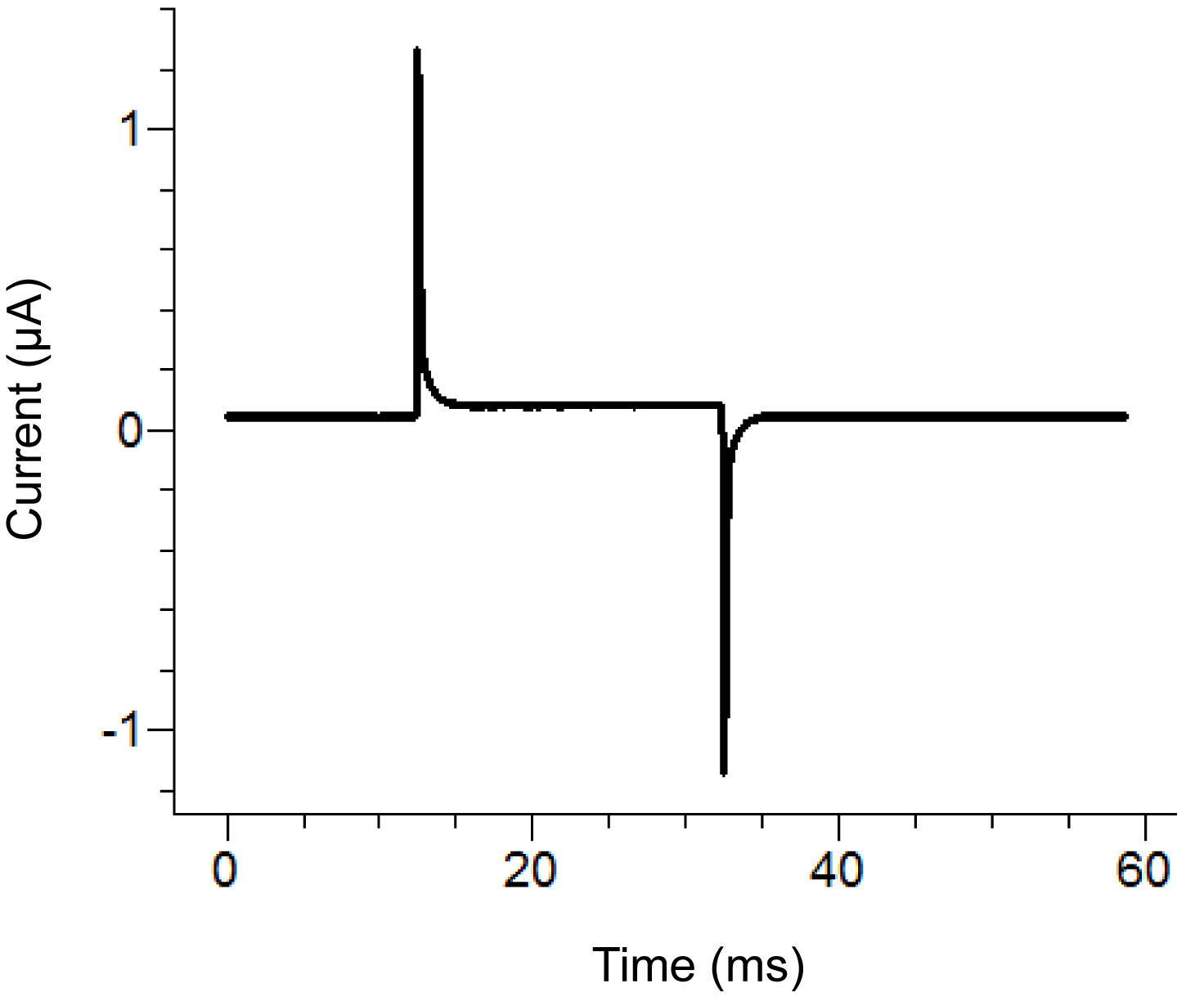
Рисунок 2. Места Агар мост Настройка Схема расположения. Размещение свободных концов агара мостовв различных банях. Нажмите здесь, чтобы увеличить изображение . - Начать протокол тестирования в программном обеспечении для записи. Если импульс показывает вертикальное перемещение горизонтальной секции между двумя пиками при приложении импульса 100 мВ, которое больше 100 нА (что соответствует 0,3 МОм с ванной / охраны в пассивный), то увеличить герметичность крышки ванны. См. Рисунок 3 для примера идеального тестового импульса.
Примечание: протокол испытаний испускает импульс напряжения, чтобы увидеть, если крышка ванны достаточно жесткой и все компоненты были собраны правильно. Кроме того, тест функции усилителя можно использовать.
Рисунок 3. Идеально тестового импульса от Software Запись. Тестовый импульсдолжна быть похожа на выше импульса в зависимости от протокола прикладного. Нынешний холдинг (основная линия в центре) должна быть близка к нулю. Нажмите здесь, чтобы увеличить изображение . - Снимите внешнее решение в нижней ванной и заменить сапонина решения. Будьте осторожны, чтобы не создавать пузырьки при добавлении сапонины. Для обеспечения максимальной замены, применять всасывания на противоположном конце нижней бане при добавлении раствора.
- После того, как сапонины как раствор был добавлен, наблюдать повторяющуюся тестовый импульс. Если пик протокола испытаний уменьшает или исчезает, то это признак того, что есть пузырь находится ниже яйцеклетки. В этом случае полностью удалить сапонина раствора, а затем заменить его.
Примечание: Пермеабилизации обычно завершается в течение 30 секунд с свежим раствором сапонина. Решения могут иметь трудности в достижении ячейку, если есть скопившийсяили если фолликулярная слой остается на плохо переваренной яйцеклетки. Ооцитов был проницаемыми (открыта), когда наклон всплеска напряжения уменьшается (увеличение постоянной времени распада).
Рисунок 4. Тест Пульс Следы Во ооцитов пермеабилизации. Выбранные следы от протокола испытаний импульса после 0,25% раствор сапонина был введен в нижней ооцитов камеры. Увеличение постоянной времени распада видели в следах демонстрирует увеличение ооцитов пермеабилизации. Нажмите здесь, чтобы увеличить изображение . - Как только клетка проницаемыми, удалите сапонина решение и заполнить ванну с внутренним решением. Остановите протокола испытаний.
Примечание: Несмотря на то, сапонины позволяет получить доступ кВнутренняя часть клетку permeabilizing мембрану, уравновешивание концентраций ионов между нижней ванной и цитоплазму ооцита путем диффузии является очень медленным процессом. Этот процесс может потребовать несколько десятков минут в зависимости от условий (рис. 5). - Проверьте, нет ли высокий уровень решения в банях и кристаллизуют KCl между скважинами многообразия, поскольку они могут привести к короткому замыканию и странное поведение.
- Используйте модифицированный шприц для введения 3 М КСl в микроэлектродом. Флик Микроэлектрод несколько раз одним пальцем, пока готовятся с другими пальцами.
Примечание: Этот шаг необходим, чтобы удалить скопившийся воздух в микроэлектродом. - Установите KCl заполненный электрод на микроманипулятора руку, вставив провод нить в конце открытой микроэлектродной. Нажмите на конце микроэлектродом в держатель нити и убедитесь, что электрод не потерять. Затянуть электрода застежку.
Примечание: Маке в том, что провод имеет четное покрытие для AgCl электрода нормально функционировать. - Свинг руку в положение над ооцитов ванны и затяните зажимы для предотвращения дальнейшего движения рук.
- Использование манипулятора ручки, ходить электрод вниз в ванну. Убедитесь, что ни один тестовый импульс не применяется, и что функция тест мембрана не активирован в этой точке.
Примечание: Перед конец электрода вставляется в жидкости, V1-V2 на зажиме напряжения будет считывать положительное напряжение. После того, как кончик электрода вставляется в жидкости, измеритель напряжения на зажиме напряжения должен измениться на значение, близкое к нулю. Для VCF записей, электрод должен подойти ячейку на достаточно небольшим углом, чтобы оставить место для объектива. Накалывание ячейку с электродом не по центру, ближе к краю изолированной мембраны патч также помогает избежать столкновения целью с электрода. - Остановить ходить электрод вниз. Установите электрод смещенияпотенциал к нулю, нажав кнопку V1, а затем снижения напряжения V1 к нулю, регулируя смещение V1. Кроме того, выполняют те же регулировки для V2. Разность потенциалов V1-V2 следует читать 000 мВ.
- Вернитесь к V1 и превратить Z-тест на измерение сопротивления электродов. Значение будет постепенно падать и приближаться фактическое сопротивление. Стремитесь к значению сопротивления 0,2-0,5 МОм.
- Продолжайте движение электрода вниз к видимой патч ооцита в верхней ванны. После того, как микроэлектрода в непосредственной близости от ооцита, смотреть показание V1-V2, чтобы увидеть, когда электрод входит в ооцит; напряжение V1-V2 станет отрицательным, когда микроэлектрода проникает в клетку.
Примечание: Значение, показанное на данный момент является мембранный потенциал клетки и будет зависеть от каналов выраженных и решений, используемых. Установка микроэлектрод слишком далеко повредит клеточную мембрану. - Откройте протокол сбора данных взаписи программного обеспечения.
- Флип на коммутаторе зажима на зажим напряжения и регулировать потенциал, чтобы соответствовать команду (например, -100 мВ), регулируя ручку, расположенную на "I" headstage.
- Флип на коммутаторе емкости и компенсации сопротивления.
- Переверните "Тест" включении в "Команды" области зажима напряжения. Используйте осциллограф для вывода сигнала. Отрегулируйте ручки компенсации см в сегменте формирования сигналов для уменьшения емкостных переходных процессов в осциллографа. Не слишком компенсировать пики до точки, где дополнительные обратные пики иметь место или пики начинают развиваться сигмоидальную кривизну, которая может ввести артефакт в записи.
- После того, как емкость была вручную снижена до удовлетворительного уровня, выключите переключатель тестирования.
- Начать протокол записи данных в программном обеспечении для записи.
{kind=link}
{kind=link}
{kind=link}
6. Уборка
- При записи есть пчелан завершена, выключите все различные переключатели на зажим напряжения, включая зажим и ванной / охранник переключателей.
- Используйте пинцет, чтобы удалить агар мосты из различных ванн.
- Снимите верхнюю ванну и аспирации все решения и яйцеклетки из всех ванн.
- Использование бутылку деионизированной воды для полоскания все ванны и затем аспирации ванны с вакуумом. Повторите этот шаг 3-5x.
- Протрите кристаллизованного KCl от мостов и поместите мосты в решение для хранения данных. Мосты могут быть повторно использованы в течение многих недель, пока они хранятся надлежащим образом.
- Аспирируйте раствора хлорида калия из коллектора скважин и промойте коллектор деионизированной водой несколько раз.
- Выключите все различные устройства, включая контроль температуры и программному обеспечению для записи.
7. Добавление напряжения Зажим флуорометрии
- Следуйте инструкциям в разделе 1 по 6 в ранее опубликованной протокола Юпитер 16 Examining конформационной динамики мембранных белков на месте с сайта-направленного флуоресценции маркировки: Щелкните здесь для просмотра страницы.
- Выполните действия, описанные в разделе 4 через 5,22 вышеупомянутого протокола CoVG использованием VCF микроскоп установлен на уровне 4-кратным фокусе.
Примечание: VCF записи требуют больших камер ооцитов ванны, чем требуется в измерениях CoVG. (Размеры пользовательских VCF камер расположены в списке материалов.) Это больше, VCF камера должна быть способна одновременно размещения объектива, микроэлектрод и агар мосты. Кроме того, животное полюс (темная сторона яйцеклетки) должно быть направлено вверх в камере для низкого фона VCF записей. - Доведите верхнюю часть ооцита в фокус с использованием водно-иммерсионной 40X цели.
Примечание: Переключение с 4X к цели 40X требует определенного GEometry срезанных открыть компонентов и тщательного внимания, чтобы не ударить электродов, мосты или камеры при опускании Объектив 40х. Кроме того, в связи с увеличением объема в верхней охранника, убедитесь, что объем ванны с верхней охраны не связано с средней стражи, когда цель 40X установлен на место. - Сосредоточьтесь на кольце по периметру открытой поверхности яйцеклетки, так что очень верхней части яйцеклетки немного выше плоскости фокуса.
Примечание: Регулировка в плоскости может быть необходимо, чтобы поле зрения в основном заполнены мембраны и не камеры. Перевод XY легче всего достигается путем размещения микроскопом на этапе трансляции. - Перемещение куб фильтра в оптическом пути и переключения пути света от окуляра к детектору (диод).
- Включите источник VCF света.
- Выключите верхний свет, волоконно света подсветки и другие источники света.
Примечание: В идеале, VCFЗаписи должны быть выполнены в совершенно темной комнате. - Запустите протокол флуоресценции в программном обеспечении для записи.
Результаты
Рисунок 4 показывает изменение проницаемости ооцита в виде раствора сапонина применяется к нижней части ооцита. Рисунок 5 демонстрирует скорость обмена внутриклеточного раствора путем диффузии следующее сапонина проницаемости. 20-40 мин обязаны прийти к стационарной...
Обсуждение
Ооцит Вазелин техника напряжение на промежутке зажим отключение открытой позволяет быстро разрешения данных, низкий уровень шума, возможность постоянного контроля за внутренним решением и внешнего состава раствора, и стабильные записи над относительно длинных протоколов 19. Эт...
Раскрытие информации
Авторы не имеют ничего раскрывать.
Благодарности
Все члены Вашингтонского университета в Сент-Луисе Сердечная молекулярной инженерии Lab. Берроуз Здравствуйте фонд Карьера премии в Научно-интерфейс - 1010299 (с JS).
Материалы
| Name | Company | Catalog Number | Comments |
| External Solution | Brand | Catalog Number | [Final], weight, or volume |
| N-methyl-D-glucamine (NMDG) | Sigma-Aldrich | M2004 | 25mM |
| MES Sodium Salt | Sigma-Aldrich | M5057 | 90mM |
| HEPES | Research Products International | H75030 | 20mM |
| Calcium hydroxide | Sigma-Aldrich | 239232 | 2mM |
| MES Hydrate | Sigma-Aldrich | M8250 | variable (pH to 7.4) |
| Internal Solution | |||
| N-methyl-D-glucamine (NMDG) | Sigma-Aldrich | M2004 | 105mM |
| MES Sodium Salt | Sigma-Aldrich | M5057 | 10mM |
| HEPES | Research Products International | H75030 | 20mM |
| Ethylene glycol-bis(2-aminoethylether)-N,N,N',N'-tetraacetic acid (EGTA) | Sigma-Aldrich | E4378 | 2mM |
| MES Hydrate | Sigma-Aldrich | M8250 | variable (pH to 7.4) |
| Depolarizing Solution | |||
| KCl | Sigma-Aldrich | 221473 | 110mM |
| Magnesium chloride | Sigma-Aldrich | M8266 | 1.5mM |
| Calcium Chloride | Caisson | C021 | 0.8mM |
| HEPES | Research Products International | H75030 | 10mM |
| Pipet Solution | |||
| KCl | Sigma-Aldrich | 221473 | 3M |
| Saponin Solution | |||
| Saponin | Sigma-Aldrich | 47036 | 0.125g |
| Internal Solution | See above | 50mL | |
| Agar Bridge Solution | |||
| N-methyl-D-glucamine (NMDG) | Sigma-Aldrich | M2004 | 100ml of 1M |
| HEPES | Research Products International | H75030 | 1.2g |
| MES Hydrate | Sigma-Aldrich | M8250 | variable (pH to 7.4) |
| Granulated Agar | Research Products International | A20250 | 3% |
| NMDG Storage Solution | |||
| NMDG, HEPES, MES Hydrate solution | see above | 40ml | |
| Water | 60ml | ||
| Name of Material/ Equipment | Company | Catalog Number | Comments/Description |
| High Performance Oocyte Clamp | Dagan | CA-1B | |
| Data Acquisition System | Axon CNS | Digidata 1440A | |
| Oscilloscope | Tektronix | TDS 210 | |
| Rack Power Filter | APC | G5 | |
| Heating/Cooling Bath Temperature Controller | Dagan | HCC-100A | |
| PC | Dell | Optiplex 990 | |
| pCLAMP 10.3 Voltage Clamp Software | Molecular Devices, LLC | pCLAMP10.3 | |
| TMC Vibration Control TableTop Platform | TMC | 64 SERIES | |
| TMC Vibration Control Air Table | TMC | 20 Series | |
| V1/I Electrode Data Collector | Dagan | part of CA-1B | |
| MX10L Micromanipulator | Siskiyou | MX10L | |
| Bath/Guard (I/V) Headstage (with appropriate connectors) | Dagan | part of CA-1B | |
| Microscope | Omano | OM2300S-JW11 | |
| Temperature Control Bath | Custom or Dagan | Custom or HE-204C | Custom chamber made from materials from Cool Polymers (D-series). Dagan also provides a prefeabricated stage (HE-204C). |
| Custom AgCl Pellet Container | Custom | Custom | Custom machined |
| Ag/AgCl electrode, pellet, 2.0 mm | Warner | E-206 | |
| External Oocyte Bath | Custom or Dagan | Custom or CC-1-T-LB | Custom machined or purchased from Dagan |
| Internal Oocyte Bath | Custom or Dagan | Custom or CC-TG-ND | Custom machined or purchased from Dagan |
| Capillaries for Agar Bridges and Pulled Electrodes | Warner | G150T-4 | |
| Rotatable Mounts for the Microscope, Micromanipulator, and Bath | Siskiyou | SD-1280P | |
| Fiber-Lite | Dolan-Jenner | LMI-600 | |
| Regular Bleach | Clorox | 470174-764 | |
| Xenopus laevis Oocytes | Nasco | LM535M (sexually mature females) | |
| 90 Na+ External Solution | See Solutions sheet | ||
| 10 Na+ Internal Solution | See Solutions sheet | ||
| 3 M KCL | See Solutions sheet | ||
| Saponin | Sigma-Aldrich | 47036 | |
| NMDG Storage Solution | See Solutions sheet | ||
| 5mL transfer pipets | SciMart | GS-52 | |
| Modified KCl electrode injector | BD | 309659 | Plastic syringe tip melted to allow for injection of solution into electrodes. Alternatively, a Microfil by WPI can be purchased. |
| Microvaccum | Custom | Custom | |
| Forceps | VWR | 63040-458 | |
| Oocyte Handling Tools (Pipette Pump) | VWR | 53502-222 | |
| Deionized Water Squirt Bottle | VWR | 16649-911 | |
| Vaseline Petroleum Jelly | Fisher Scientific | 19-086-291 | |
| Additional Materials Required for VCF Recordings: | |||
| VCF Microscope | Nikon | Eclipse FN1 | |
| Nikon CFI APO 40XW NIR Objective | Nikon | N40X-NIR | |
| X-Y Translator System for Fixed-Stage Upright Microscopes | Sutter Instruments | MT500-586 | |
| External VCF Oocyte Bath | Custom | Custom machined. The chamber dimensions are 2.7 x 1.9 x 0.4 cm. | |
| Internal VCF Oocyte Bath | Custom | Custom machined. The chamber dimensions are 1.6 x 1.6 x 0.4 cm. | |
| Modified Temperature Control Bath | Custom | Custom chamber made from materials from Cool Polymers (D-series). The chamber dimensions of the modified temperature controller bath are 2.7 x 1.9 x 0.3 cm for the horizontal chamber, and 1 x 2.5 x 0.5 cm for the vertical chamber. |
Ссылки
- Kalstrup, T., Blunck, R. Dynamics of internal pore opening in KV channels probed by a fluorescent unnatural amino acid. Proc. Natl. Acad. Sci. U.S.A. 110, 8272-8277 (2013).
- Stefani, E., Bezanilla, F. Cut-open oocyte voltage-clamp technique. Methods Enzymol. 293, 300-318 (1998).
- Muroi, Y., Chanda, B. Local anesthetics disrupt energetic coupling between the voltage-sensing segments of a sodium channel. J. Gen. Physiol. 133, 1-15 (2009).
- Stefani, E., Toro, L., Perozo, E., Bezanilla, F. Gating of Shaker K+ channels: I. Ionic and gating currents. Biophys. J. 66, 996-1010 (1994).
- Wang, S., Liu, S., Morales, M. J., Strauss, H. C., Rasmusson, R. L. A quantitative analysis of the activation and inactivation kinetics of HERG expressed in Xenopus oocytes. J. Physiolt. 502 (Pt 1), 45-60 (1997).
- Neely, A., Garcia-Olivares, J., Voswinkel, S., Horstkott, H., Hidalgo, P. Folding of active calcium channel beta(1b) -subunit by size-exclusion chromatography and its role on channel function. J. Biol. Chem. 279, 21689-21694 (2004).
- Silva, J. R., Goldstein, S. A. Voltage-sensor movements describe slow inactivation of voltage-gated sodium channels I: wild-type skeletal muscle. Na(V)1.4. J. Gen. Physiol. 141, 309-321 (2013).
- Silva, J. R., Goldstein, S. A. Voltage-sensor movements describe slow inactivation of voltage-gated sodium channels II: a periodic paralysis mutation in Na(V)1.4 (L689I). J. Gen. Physiol. 141, 323-334 (2013).
- Taglialatela, M., Toro, L., Stefani, E. Novel voltage clamp to record small, fast currents from ion channels expressed in Xenopus oocytes. Biophys. J. 61, 78-82 (1992).
- Clare, J. J., Trezise, D. J. . Expression and analysis of recombinant ion channels : from structural studies to pharmacological screening. , (2006).
- Cha, A., Zerangue, N., Kavanaugh, M., Bezanilla, F., Susan, G. A. . Methods in enzymology. 296, 566-578 (1998).
- Lakowicz, J. R. . Principles of fluorescence spectroscopy. 3rd edn. , (2006).
- Cha, A., Bezanilla, F. Characterizing voltage-dependent conformational changes in the Shaker K+ channel with fluorescence. Neuron. 19, 1127-1140 (1997).
- Richards, R., Dempski, R. E. Examining the conformational dynamics of membrane proteins in situ with site-directed fluorescence labeling. J. Vis. Exp. , (2011).
- Cohen, S., Au, S., Pante, N. Microinjection of Xenopus laevis oocytes. J. Vis. Exp. , (2009).
- Raynauld, J. P., Laviolette, J. R. The silver-silver chloride electrode: a possible generator of offset voltages and currents. J. Neurosci. Methods. 19, 249-255 (1987).
- Gagnon, D. G., Bissonnette, P., Lapointe, J. Y. Identification of a disulfide bridge linking the fourth and the seventh extracellular loops of the Na+/glucose cotransporter. J. Gen. Physiol. 127, 145-158 (2006).
- Pantazis, A., Olcese, R., Roberts, G. . Cut-Open Oocyte Voltage-Clamp Technique. In: Roberts G. (Ed.) Encyclopedia of Biophysics: SpringerReference. , (2013).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены