
Method Article
Toplanması ve analizi
Bu Makalede
Özet
Floem sap bileşimi yanı sıra yükleme ve uzun mesafe taşıma mekanizmasının Bilgi bitki gelişimi ve stres / patojen yanıt olarak ve ulaşım asimile uzun mesafe sinyal anlaşılması için gereklidir. Bu el yazması EDTA-kolaylaştırdı yöntemi kullanılarak floem eksuda toplanması açıklar.
Özet
Bitki floem uzun mesafe taşımacılığı (fotoğraf-) ve biyotik veya abiyotik stres taşıma sinyallerinin olarak özümseyen için gereklidir. Bu şekerler, amino asitler, proteinler, RNA, lipid ve diğer metabolitleri içerir. Böylece kompozisyonu ve fonksiyon floem ki, bu moleküllerin birçok rolü ve anlamada büyük bir ilgi varken, bitki gelişimi ve stres önemleri yanıt belirlenecek henüz. Floem analizi için bir engel üzerine floem mühürler kendisini yaraladı gerçeği yatıyor. Sonuç olarak, floem sap elde edilebileceği bitkilerin sayısı sınırlıdır. Eklendi ekipman olmadan çeşitli bitki türlerinden floem eksuda toplanması sağlayan bir yöntem EDTA-kolaylaştırdı floem eksuda koleksiyonu burada açıklanan olduğunu. Kullanımı kolay iken, hücrelerin yaralama yol yapar ve bakım hasarlı hücrelerin, içeriği kaldırmak için alınması gereken. Buna ek olarak, çok sayıda kontrol eksüda saflığı kanıtlamakgereklidir. Oldukça içeriğini sadece göreceli miktar oluşabilir floem sap (birçok türde mümkün değil) doğrudan bir koleksiyon daha bir sızma olduğu için. Diğerleri üzerinde bu yöntemin avantajı birçok otsu veya odunsu bitki türlerinin (Perilla, Arabidopsis, kavak, vb) kullanılan ve minimal ekipman ve eğitim gerektirir olmasıdır. Bu proteinler, şekerler, lipitler, RNA virüsleri ve metabolitlerin sonraki analiz için kullanılabilir dışkılarının oldukça büyük bir miktarda yol açar. Bu, hem araştırma hem de bir öğretim laboratuarda kullanılabilir kadar basittir.
Giriş
Bitkiler olumsuz koşullar kaçmak için hareket edemez. Sonuç olarak, onlar, çevresel streslere tespit etmek için mekanizmalar geliştirmek ortaya çıkarmak ve bitki boyunca ilgili sinyal iletimi, uygun geliştirme ayarlamak zorunda kaldı. İki taşıma sistemleri su, besin ve diğer (sinyal) bileşiklerin dağıtımı için var. İlk ksilem değil, genellikle su taşır ve mineraller bitki boyunca kökleri tarafından alınır hangi. İkinci floem olduğunu. Floem görüşünü RNA için basit asimile taşıma sistemi bir kanal değişti, protein, virüs, lipidler ve diğer küçük moleküller. Bu önemli bir rol oynar asimile ve besin taşıma, biyotik ve abiyotik stres yanıt, hem de bitki büyüme ve gelişme gibi. Şimdi bitki 18 "bilgi otoyolu" olarak adlandırılır.
Floem parankimi, hem de özel arkadaşı CE: floem çeşitli hücre tipleri oluşurlls ve elek elemanları. Elek elemanı uzun mesafe hareketin sitesidir. Engelsiz uzunlamasına akışı için izin vermek için, elek elemanlarının yanı çekirdekleri gibi birçok organeller eksik ve iyi sınırlı bir öteleme makine 21, 33 içeren düşünülmektedir. Bu hücreleridir proteinler ve floem akışında seyahat eden diğer bileşikler, sentez olduğu düşünülmektedir. Bu bileşikler daha sonra plasmodesmata ile elek elemanı içine taşınır ve uzun mesafe sinyalleri 4,16 olarak işlev görebilir.
Bileşiklerin birkaç grup floem sıvılarını bulunabilir:
- Şeker genellikle fotosentez ürünlerdir ve depolama için bitkinin diğer kısımlarına "enerji taşıyan" molekülleri olarak ya da yapı taşları olarak taşınmaktadır. Ancak, onlar da antifriz veya bileşikler sinyal olarak işlev görebilir.
- Proteinler de floem eksüda bulunabilir. Bu metabolik enzimler arasında, ancak aynı zamanda fonksiyon sinyalizasyon var. Bir exampgelişimsel bir sinyal olarak hizmet veren bir protein le çiçeklenme indüksiyon yanı sıra bitkilerde mevsimsel yaprak kesilme sinyalleri Çiçekli odağı T proteindir. 6
- Nükleik asitler mRNA, küçük ve mikro RNA'lar ve viral RNA'lar şeklinde floem sızıntıdaki bulunmaktadır. Bunlar büyük ölçüde sinyal 27, 28, 39 dahil olmak görünür. Aynı zamanda, rRNA ve hemen hemen tüm amino asitler için tRNA'lar kabakgiller 42 floem sap gözlenmiştir. Bunlar seçmeli olarak elek boru sistemine aktarılması gibi görünmektedir. TRNA'lar translasyonel cihaz için bir amino asit aktarma yeteneği için gerekli değişiklikler göstermektedir. Ancak, oldukça yapıldı katılan, daha da bu süreçte inhibe ederler. Alternatif olarak, bu sitokinin bir kaynağı olarak hizmet ya da bitkinin 42 metabolik durumu sinyal olabilir.
- Yağ asitleri, oxylipins ve diğer lipidler de floem eksüda 3, 13, 14, 23 olarak tespit edilmiştir. JasmoNIC asit, patojen enfeksiyonu 22, 29, 31, 32 ile ilgili tepkinin bir parçası olarak, floem aracılığıyla bir oxylipin hareket eder. En floem lipidlerin rolü henüz belirsiz olduğundan, bazıları muhtemelen fonksiyonu 4 sinyal var.
Bitki floem ile çalışma meydan yaralama üzerine kendisine mühür kapasitesini yatıyor. Orada floem eksuda toplamak için kullanılan dört ana yöntem vardır, ama onlar seçin türlerinde sadece çalışır:
1) cucurbits yılında yaprak sapı kesim ile floem eksüda makul miktarlarda elde etmek mümkündür. Yaralı hücre kontaminasyon başlangıç damla kaldırıldıktan sonra, saf floem sap 1, 15 arasında oldukça büyük bir miktarda elde etmek mümkündür. Ancak, artan toplama zaman, sıvı kromatografi yaklaşımlar (yayınlanmamış) için giderek uygun olmayan bu eksuda kalınlaşır yapma. Son yayınlar türüne göre, bu floem sap eithe türetilmiştir olduğunu, göstermektedirbu elek elemanları (FP) için "mobil" floem sap içeren yapar iken r fasikül (FP) veya extrafascicular floem (AP) ve bu, aynı zamanda ksilem 40 dahil olmak üzere diğer hücre tipleri, 41 kirlenme eğilimli .
2) İkinci bir yaklaşım kök veya yaprak sapı sığ kesik veya delikler ile floem sap elde etmektir. Bu yöntem, acı bakla 17, 25, 36 kabakgiller ve Brassica napus 12 başarılı bir şekilde kullanılmıştır. Burada yaralı hücrelerinden kirlenme az ve çok saf dışkıların. Ancak, bitkiler çok seçici delinme sadece elek elemanları için zor olduğu için sağlıklı ve sulak olması gerekir. Ksilem damarları çentikli ise, tüm eksuda ksilem akışı içine çekilir. Bu çok kırılgan veya yüksek Odunlaşmış sapı veya sapları ile bitkiler için uygun yöntemi yapar.
3) Yaprak biti stylectomy yaprak bitleri elek elemanları içine stile eklemek için izin verirn bir lazer ile yaprak biti kaldırır. Floem sap kalan Stile 2, 9, 10, 35, 38 boyunca yayılır. Teorik olarak, yaprak bitleri tarafından enfekte edilebilir herhangi bir bitki bu yaklaşım için kullanılabilir. Ancak, çoğu sera ya da büyüme odası yöneticileri patojenin kullanımını desteklemez. Buna ek olarak, yaprak bitleri kendi tükürük 19, 33 ile floem içine birkaç proteinlerin tanıtmak. Bu 30 yeniden programlama sınırlı bir transkripsiyonel yol açar ve floem kompozisyon 26 değiştirme potansiyeline sahiptir.
4) Burada anlatılan yöntemi floem sap EDTA-kolaylaştırdı sızma olduğunu. Bu yöntem, floem 20 sızdırmazlık önlemek için EDTA kullanmaktadır. EDTA Ca 2 + aksi floem mühür bu süreçleri katılacak iyonları şelat. EDTA hücre hasarı ile 30 yol açabilir, çeşitli gruplar, hücre ince yapısı veya phloe 20 mM kullanılan bu yöntem, 10 mM EDTA, konsantrasyonlarının olumsuz etkisi gözlemledikm yükleme ve taşıma 5, 24. Herhangi bir zararlı etkisi yanı sıra kromatografisi ve jel elektroforezi ile EDTA girişimi azaltmak için, bitkiler 1 saat sonra suya taşınır ve eksüda sadece sonraki bölümünde 14 kullanılır. Bu nedenle, yerine EDTA içine eksudasyon daha, floem su (EDTA-kolaylaştırdı sızma) içine exudated edilir. Bu floem eksuda toplanması için bir düz ileri, düşük maliyetli ve düşük teknolojili bir yöntemdir. Floem SAP bu şekilde proteinler, küçük moleküller, lipidler ve RNA'lar analiz etmek için kullanılabilecek pek çok bitki elde edilir ve başarılı bir şekilde kullanılmıştır. Deneyler sınırlı bir miktarda monokotlarda 11 gerçekleştirilirken, bu yöntem dikotlar (Perilla 17, 20, Arabidopsis, 8, 13, 14, Kavak 7) için daha uygun gibi görünmektedir. Eksuda toplanması terleme yoluyla eksuda kaybını önlemek için nemli bir ortam meydana zorundadır. Bitki bağlı olarak, 1-2 saat boyunca, EDTA inkübasyon olduğufloem / elek elemanları sızdırmazlık önlemek için yeterli. Toplama sonra su içine oluşabilir. Bu EDTA hücre yapısı ve istikrar üzerindeki olumsuz etkisi önlenmesi yararı vardır. Ayrıca, HPLC ya da SDS-page gibi yöntemler ile EDTA müdahale ortadan kaldırır. Bu Arabidopsis için geçerli gibi gösterilir. Daha büyük tesisler için, EDTA ve eksüda koleksiyonlarında inkübasyonu ölçeklenmesi sahiptir ve oldukça 1.7 ml'lik tepkime tüpleri daha kap içinde gerçekleştirilir.
Protokol
1. Bitkiler hazırlanması
- Arabidopsis tohum doğrudan toprağa veya MS plakalar üzerinde çimlenmiş olabilir. 12 saat fotoperiyodun (22 ° C gün, 15 ° C gece 14) ile büyüme odalarında dört ila altı hafta için bitkiler büyümek. Su bitkileri bir kez haftada iki kez. Sulama için alt tepsi kullanın, üstten değil su bitkileri yok, tepsiler yeniden sulama önce kurumasını bekleyin. Bitkilerin hasat sırasında bol sıvı olduğundan emin olun.
2. Çözümler ve Yemekleri hazırlanması
- K 2-EDTA çözeltisi: buzdolabında tutulabilir bir 100 mM K 2-EDTA çözeltisi stok solüsyonu, olun. Toplama gününde, 4:59 (bir kısım EDTA solüsyonu, dört kısım su), 20 mM K 2-EDTA çözeltisi elde etmek için çözeltinin.
- Yarım K 2-EDTA çözeltisi 20 mM ile bir cam veya Petri kabı (çapı 7-15 cm) doldurun. Eğer farklı tedaviler (ex için örnekler toplamabol kontrolü ve tuz stresli bitkiler veya farklı genotipleri), her tedavi için ayrı yemekler hazırlamak.
- 20 mM K 2-EDTA çözeltisi, 1.4 ml 1.7 ml'lik tepkime tüpleri, istenilen sayıda doldurun. 1.4 ml reaksiyon tüpleri eşit sayıda doldurun deiyonize veya Millipore su otoklava.
- Üzerinde bitki ayarlamak için bankta kağıt havlu koyun; yeni bir jilet almak ve eldiven koymak.
3. Floem eksudata toplanması (Şekil 1'de Akış Şeması in tasvir)
- Hasat rozet rozet ortasına yaprak sapı yakın dibinde jilet ile kesme yılına kadar dört ila altı haftalık-eski bitkilerden bırakır.
- Hemen 2-EDTA K 20 mM içeren yemekler yaprakları yerleştirin. Yaprak sapı, kesilen ucu (Şekil 2) çözüm altında olduğundan emin olun.
- Bir kez 15 yaprak hafifçe birbirine bu t üzerine yaprak yığını, (yaklaşık 3-4 bitkiler) toplanmıştırHat kesme sapı birbirleri ile hizalanır. Sapı (yaklaşık 1 mm) tabanına yeniden kesim ve hemen K 2-EDTA solüsyonu, 20 mM ile reaksiyon tüpleri içine bir yaprak aktarın. Onlar biraz sıvamış ise yaprakları kolay uyum. Bu hücrelerin hasar ve, böylece, yaprak hücre içeriğinin toplanması yol açar neden olabilir bu yana en yaprakları sıkma kaçının.
- Daha fazla malzeme gerekiyorsa, nazikçe bir ıslak kağıt havlu ile örnekleri kapsayacak. Eğer farklı tedaviler veya genotip örnekler toplanması için yemekler yeni bir dizi kullanın.
- Sonra, tüm numuneler, karanlıkta ya da ışık sıvılarını toplamak, reaksiyon tüpleri konumlandırılmıştır.
- Karanlık, çizgi ıslak kağıt havlu ile büyük bir sığ çanak katta toplanması için. Yaprak dolu tepki çanak tüpleri ve ıslak kağıt havlu ile kapak ya ile rafa yerleştirin veya havalandırma için yarıklar ile bir siyah plastik geri koymak. Bir kabine tüm kurulum veya karanlık bir odada yer.
- Işığında toplama, ıslak kağıt havlu ile net bir pleksiglas kabın dibine satır. Kapta yaprak dolu reaksiyon tüpleri ile raf yerleştirin ve kapatın.
- 1 saat sonra, kaldır konteyner ve reaksiyon tüpleri yavaşça bırakır ve tüm EDTA kaldırmak için, distile deiyonize veya Millipore su ile iyice yıkayın. Bu adım, üç amaca hizmet eder: (1) bu hücrelere hasar minimize etmek için EDTA kaldırır (2), SDS-PAGE ve kromatografi ile EDTA müdahaleyi ortadan kaldıran, ve (3) kesme biriken / zarar veren herhangi bir bileşik kaldırır hücreler. Hemen otoklava su içeren hazırlanan reaksiyon tüpleri içine aktarmak ve eksuda toplanması için nemlendirilmiş toplama kapları dönmek. Eksüda RNA ekstraksiyonu için kullanılması gerekmektedir, bu noktada, RNaz inhibitörü ekleyin. İsteğe bağlı: protein analizi istendiği takdirde proteinaz inhibitörü ilave edilebilir.
- Amaçlanan toplama süre sonra, çekin ve Akıntıyard yaprakları ve kullanım için sıvı N 2 floem eksuda dondurma. Uzun süre depolama gerekiyorsa, -80 örnekleri ve mağaza ° C lyophilize
- Arabidopsis durumunda, metabolitler önce 1 saat boyunca dışkılarının toplandıktan sonra tespit edilebilir. Daha az oranda mevcut bileşenlerin analizi planlanan Ancak, 5-8 saat için toplama tavsiye edilir. Perilla gibi büyük tesisler için, uzun toplama saatleri (8 saat) önerilir.
15 yapraklardan toplanmıştır eksüda bir mRNA'nın çıkarılması için SDS-PAGE jel (proteinler), GC-MS (şeker ve metabolitler) üzerinde tek bir şerit için genellikle yeterlidir. Lipid analizi için 2-3 örneklerin havuzu (30-45 yapraklarından eksüda) tavsiye edilir ve daha net sonuçlara yol açar.
4. Sonraki Analizi (Video için Biz sadece 4.4 göster)
- Protein analizi için, 15 ul su ve 30 ul LÄMMLI tampon tekrar süspansiyon örnek, 95 ısı ° C 5 dakika, hızlı bir şekilde herhangi p aşağı doğru döndürünrecipitates ve SDS-PAGE jel (45 ul yükleme cepler) üzerine tüm örnek yükleyin. Protein bolluk çok düşük olduğu için örnekleri Kolloidal Coomassie veya diğer hassas lekeleri ile boyanmış olmalıdır.
- Şeker ve küçük metabolit analizi için, literatürde 14'te tarif edildiği gibi kuru numune ajanlar Türevlendirme ekleyin.
- mRNA RT-PCR, mikroarray veya RNA-Sıra analizi kullanılarak analiz edilebilir.
- Lipit analizi için hemen havuzu 2-3 örnekleri ve kloroform karşı bölüm aşağıdaki gibidir: Bir çeker ocak içinde metanol (1:1) ile: 4 dk için her bir örnek için Metanol, bir kaç saniye boyunca girdap ve santrifüj: eşit Kloroform miktarı eklemek 400 x g. Teflon kapağı ve üst faz ile tekrar bölümü üç kez daha bir cam tüp içinde alt (organik) faz toplayın. N2 akışı altında bir araya getirilen organik fazlar kurutun ve (İnce tabaka kromatografisi daha fazla analiz için göndermek TLC 14, 37; sıvı kromatografisi-kütle spektrometrisi: MC-MS 14).
Sonuçlar
Bu floem sap karmaşıklığı bitkiler en iyi yanıt gelişim ve stres sinyalleri değişen göndermek nasıl sorusuna cevap olabilir ki son birkaç yıl içinde netlik kazandı. EDTA-kolaylaştırdı floem eksüdasyon kullanan diğer yaklaşımlar için uygun değil, ekonomik ya da fizyolojik ilginizi çekebilecek bitkilerden floem eksuda analiz etmek için fırsat sağlayabilir.
Yeterli floem ve hasat floem-lokalize proteinleri belirlemek ve (Şekil 3) geliştirme veya strese yanıt olarak düzeylerini değişiklikleri algılamak için eksuda için bu yöntemi sağlar. Rakam gösterir, protein çok düşük bolca olduğu için, henüz düzeylerini LC-MS/MS kullanarak sonraki proteomik deneyler için veya Western Blot analizi için yeterince yüksek. Kabakgiller bulgular kök kesme su potansiyeli denge bozulması ve daha sonra bir su akışına yol açıyor ve düşündürmektedirapoplastı 41 mümkün kirletici. : Yine bir ila sekiz saat arasında değişen zamanlar için toplama floem protein profili / kompozisyon Arabidopsis'te (. Yalnızca mutlak miktarı Guelette ark değişir) toplandı, protein miktarı ve bulunan protein deseni etkilemez türleri (Arabidopsis arasında değişmektedir de; Perilla, şerit I ve NI) ve tedaviler (Perilla farklı bir gün uzunluklarında büyüdü, şerit I ve NI). Sonuç protein sinyalleri görüntülenebilir gibi, tespit ve takip.
mRNA ve mikro RNA'lar, aynı zamanda bu yöntemle floem sıvılarında tespit edilebilir. Bununla birlikte, RNaz inhibitörü ile tedaviye genişletilmiş eksudasyon süresinde mRNA bozulmasını önlemek için gereklidir. Bu elek elemanlar işlevsel kloroplast, RUBISCO küçük ya da büyük bir alt birim (sırasıyla, eritrositler veya rbcL), içermediği floem çok bilinen mRNA, negatif kontroller olarak kullanılır-Lokalize Ubiquitin-bağlayıcı enzim gibi mRNA bir pozitif kontrol (Şekil 4) olarak kullanılabilir.
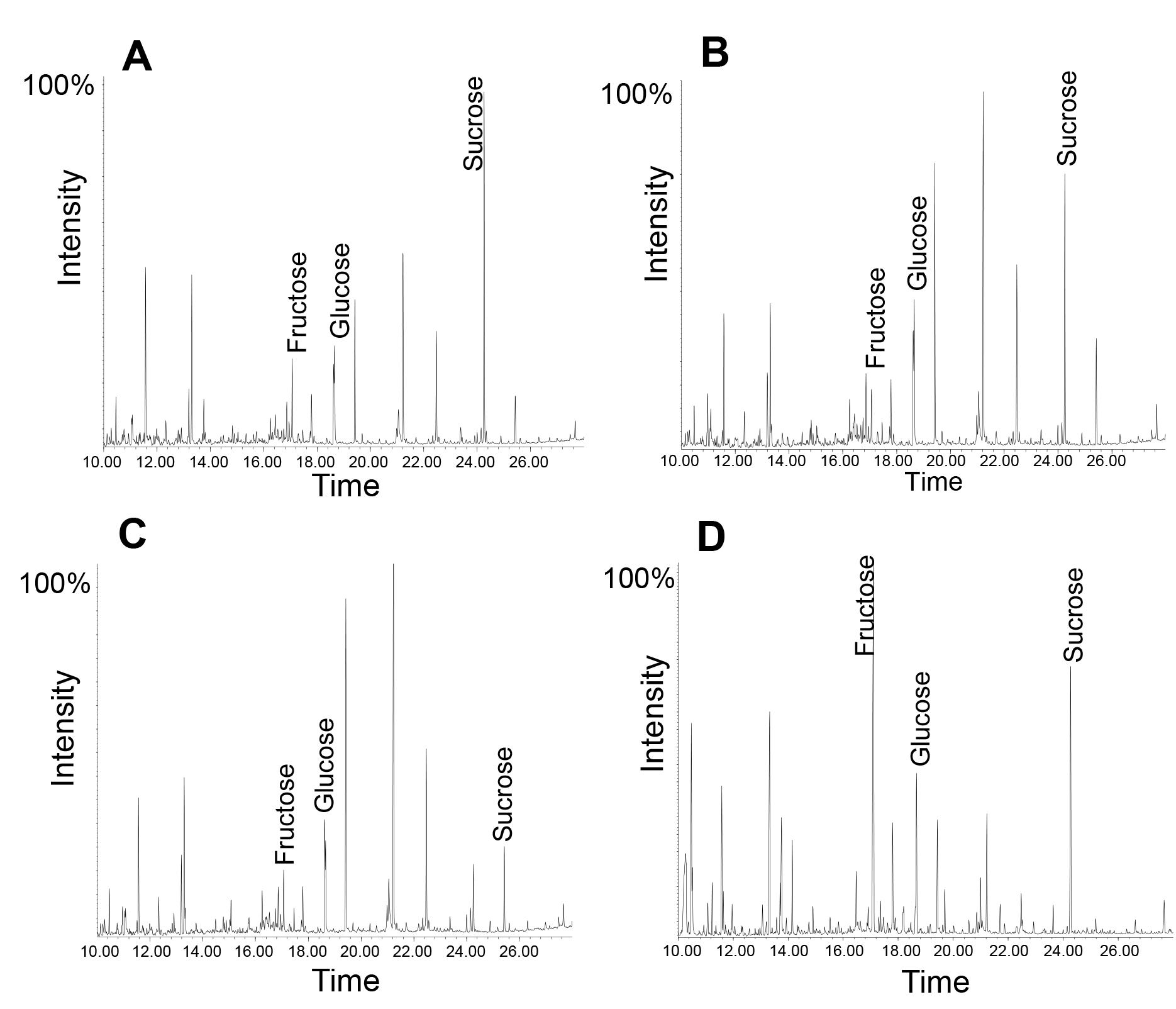
Benzer şekilde, şeker ya da küçük metabolitleri GC-MS ve LC-MS kullanılarak analiz edilebilir. Burada floem eksüda neredeyse tüm glikolitik yolun 12 de dahil olmak üzere fonksiyonel bir enzim, çok sayıda içerdiği belirtilmelidir. Bu nedenle, birkaç zaman noktalarını kullanarak eksuda toplama sırasında metabolik süreçleri ortaya çıkarabilir. Bir örneği Şekil 5'te gösterilmektedir: Tüm sızıntıdaki, sukroz en bol metaboliti, bu 1 saat boyunca toplandıktan sonra çok açıktır. Bununla birlikte, beş saat sonra fruktoz oranı sakaroz biraz azalır. Aynı eksüda sonraki dört saat için, bir saat için, ve tezgah üzerinde bırakılırsa, oran fruktoza sukroz floem aktif enzimler oda sıcaklığına de sakaroz bozulmaya yol dışkıların düşündüren, çok daha büyük bir ölçüde azaltılırsıcaklık (daha yoğun yorumlar için 14). Bu, sistemin canlılığını 34 kaybedene kadar floem yükleme ve elek elemanları içine arkadaşı hücrelerinden moleküllerin taşıma sızma sırasında devam edebilir bulgularla tutarlıdır.
Floem eksuda lipid ve, uzantısı, uzun mesafe lipid sinyal bitki bilimi ilgi oldukça yeni bir alan vardır. Hidrofobik doğası nedeniyle, lipidlerin düşük konsantrasyonlarda mevcut olan ve yalnızca çözündürme için diğer moleküllere bağlı olabilir. Yine de, EDTA-kolaylaştırdı eksudasyon görselleştirmek için yeterli malzeme toplanmasına olanak (TLC: Şekil 6) ve (LC-MS; es Şekil 7) tespit birkaç bitki türünden floem lipidleri bulunmaktadır. Şekilde görüldüğü gibi birkaç lipid türleri belirlemek ve ayırmak mümkündür. LC-MS farklı lipid türleri tanımlamak için ve differe bir lipid profili içinde değişiklik izlemenizi sağlarnt genotipleri veya tedavi. Bu bitki gelişimi ve stres tepkisi sırasında lipid rolünün çalışılması için olanak sağlar.

Şekil 1. Arabidopsis veya Perilla bir floem dışkılarının toplanması akış şeması. Farklı yollar mavi gösterilir farklı son ürünler, yol. RNA, RNaz inhibitörü (100 U / ml, Roche) nın hazırlanması için, floem eksüda toplandığı içine su ilave edilir.
Büyük resim görmek için buraya tıklayın .
{kind=link}

Şekil 2. Toplamak floem eksüda için malzeme Kurulumiyon. Kurulum protokol adımları 3,1-3,4 için gerekli tüm malzemeler görüntüler.
Büyük resim görmek için buraya tıklayın .
{kind=link}

. Moleküler ağırlık belirteci (şeritli 2): MW; floem bulundu Şekil 3 Proteinler iki farklı bitki türleri, Arabidopsis (At) ve Perilla (; farklı bir gün uzunlukları I ve NI) bir eksuda. Proteinler,% 10-20 gradyan SDS-PAGE kullanılarak ayrıldı.
Büyük resim görmek için buraya tıklayın .
{kind=link}

Şekil 4,. AnaliziRubisco küçük ve büyük alt ünitesi için mRNA (sırasıyla, eritrositler ve rbcL) ve ubikuitin-bağlayıcı enzim (UBC) varlığı. mRNA Arabidopsis yaprağı (L), yaprak sapı (P) ve floem eksüda (Ph) toplanmıştır, PCR kullanılarak görüntülendi transkripsiyonu ve belirli transkript ters. Bu rakam Guelette ark güncellenmiştir. 14.
Büyük resim görmek için buraya tıklayın .
{kind=link}

Floem Şekil 5. GC-MS profilini kez farklı miktarda tahsil eksudatları. Not nasıl toplama süresi (B), 5 saat için 1 saat toplama (A) 'dan sukroz değişiklikler nispi miktarı. Odasında "kuluçka" ardından 1 saat toplandıktan sonra Profil eksüda Dört saat boyunca sıcaklığında (RT) (° C). Yaprak metaboliti profili (D).
Büyük resim görmek için buraya tıklayın .
{kind=link}

Şekil 6. İnce tabaka Perilla (solda) ve Arabidopsis (sağ) yaprakları ve floem eksüda lipidler karşılaştırarak kromatogram. Yıldız eksüda floem spesifik lipidler göstermektedir. Dgdg: digalactosyldiacylglycerol; PG: fosfatidilgliserol; MGDG: monogalactosyldiacyglycerol; Arabidopsis lipidler gösteren rakam kısmı Guelette ark değiştirildi 14..
Büyük resim görmek için buraya tıklayın .
{kind=link}

Şekil 7. LC-MS / negatif iyon modunu kullanarak Arabidopsis floem eksuda (kloroform faz) Lipid profili. En iyi olarak Grafikler (mutantı A) ve alt (vahşi tip B) iki farklı genotipleri (oklar ile gösterildiği gibi) arasında lipid farklılıklar göstermektedir.
Büyük resim görmek için buraya tıklayın .
{kind=link}
Tartışmalar
Floem eksuda EDTA-kolaylaştırdı toplama, çok basit az ekipman ihtiyacı ve birçok bitki için geçerlidir. Genel olarak, bu yöntem, daha fazla türe floem eksüda analiz yeteneği uzanır. Eksuda seyreltilmiş olmasına rağmen, malzeme büyük miktarda büyütmek için birçok farklı örnekleri toplamak ya da birçok bitkilerden kolaydır. Bu da çok düşük bolca ve aksi takdirde göz ardı edilebilir eden yeni bileşiklerin saptanması için izin verir.
Bu yöntem, birden fazla bitki türleri için kullanılan ve bu yazının listelenen olanlar için tarif olarak çalışıyor olabilir. Yeni tür incelenmiştir ise, değişiklik gerekebilir iki yönü kullanılan yaprak ve eksuda sırasında sayısıdır. Çoğu durumda, bir 5-8 saat eksudasyon uygun olmalıdır. Yeni bitki türleri için bu yöntemin kullanımını doğrulamak için, eksüda protokolü parçası 4 N tarif edildiği gibi toplanır ve sonraki analiz bir veya daha fazla olması gerekireeds yapılacak. Gözlenen sinyal yoğunluğuna bağlı olarak, toplama hacmi büyütülüyor gerekebilir. Eski floem eksuda daha az bol olduğundan Genel olarak, lipid ve protein analizi şeker ve metaboliti analizi daha fazla malzeme gerektirir.
EDTA kolaylaştırılan akıntı tahsilat kesilmiş yüzeyler yanı sıra, EDTA, potansiyel olarak zararlı etkisi içerir için, birkaç nokta dikkat edilmesi gerekir: (1) EDTA ile bir saat inkübe edildikten sonra, yaprak sapı iyice yıkanması gerekir. Bu yaralı hücreleri, hücrelerin zarar verebilir ya da çıkarma engelleyebilir EDTA kendisi, elde edilen herhangi bir bileşik kaldırır. (2) Pozitif ve negatif kontroller elde edilen veriler floem sap elde sağlamak için dahil edilmesi gereken değil, yaralı hücrelerinden (aşağıdaki kritik adımlar bakınız). (3) Floem sap eksuda toplama sırasında aktif olabilir çeşitli enzimler, içermiyor. Bu nedenle, bazı durumlarda, bunun için toplamak için yararlı olabilirZaman r farklı miktarlarda. Yöntem, su içine sızma dahil yana (4), bileşiklerin mutlak değerlendirilmesi mümkün değildir.
Kritik adımda: (. Ref 14) uygun kontrol herhangi bir hücrelerin yanı sıra kullanım zarar vermez için numune taşıma sırasında bu yöntemin başarılı kullanımı için önemli dikkatli kullanılmasıdır. Bir uygun kontrol EDTA ile önceden tedavi olmadan floem dışkılarının topluluğudur. Bu durumda floem kendisi mühür ve hiçbir eksuda toplanabilir. Yaralı hücreleri gelen herhangi bir tehlike bileşikler burada algılanır. İkinci bir kontrol eksüda şekerlerin analizi olacaktır. Arabidopsis, sukroz 1:04 ve 1:08 (ürün 8, 14) arasında bulunan sakaroz oranına sahip fruktoz, floem SAP içinde baskın metaboliti olmalıdır.
RNA ekstraksiyonu için, bir RNaz inhibitörü kullanılması gereklidir. Daha önce describ Buna ek olarak, bir pozitif kontroled floem-lokalize mRNA (UBC9: Ubiquitin bağlayıcı enzim 9, At4g27960, 8, 14) ve örneğin bir negatif kontrol bir kloroplast mRNA (Rubisco LSU) tavsiye edilir.
Eksuda aktif enzimler içerdiğinden, bir süre ders toplama zaman farklılıklar sadece nedeniyle değil floem profilinde emin değişiklik yapmak için tavsiye edilir. Bazı durumlarda, proteinaz inhibitörleri, kullanmak yararlı olabilir. Ancak, araştırmacılar toplanan eksuda konsantre ve eklenen bileşikler büyük ölçüde konsantre olacağını olacağını, akılda tutmak gerekir. Protokol içinde ikinci bir kritik adım EDTA altında yaprak sapı en recutting olduğunu. Bu adım ikili bir amaca hizmet eder: Yeni böylece floem sap serbest akışını sağlayan fişler oluşumunu önlerken İlk olarak, herhangi bir kapalı elek plakaları kaldırır. İkincisi, kesim sırasında oluşan ksilem içinde kavitasyon kabarcık kaldırır ve böylece ksilem taşıma için izin verir. İkinci kesim büyüklüğü bağlıdırkullanılan bitkiler ile ilgili. Büyük sapı ile tesislerde bu Arabidopsis gibi bitkilerde ilk kesim, yukarıda birkaç milimetre (en fazla 1 cm) olmalı, 1-2 mm yeterlidir.
Bu yöntemin kullanımı, gelişim veya biyomedikal etkileri sinyal olabilir floem sızıntıdaki yeni bileşiklerin keşfine yol açabilir. Bu floem erişim eksikliği nedeniyle bitki bilimi içinde az çalışılmış bir alan oldu uzun mesafe lipid sinyal, de derinlemesine bakmak daha yol açtı.
Açıklamalar
Biz ifşa hiçbir şey yok.
Teşekkürler
Bu çalışma, Ulusal Bilim Vakfı NSF-IOS hibe # 1144391 için SHB tarafından desteklenmiştir.
Malzemeler
| Name | Company | Catalog Number | Comments |
| K2-EDTA | Sigma-Aldrich | ED2P-500G | |
| Shallow Glass or plastic Petri Dish (7-15cm) | PYREX or Corning | Any clean, shallow dish will work | |
| Chloroform | EMD | CX1054-1 | Only open containers in fume hood |
| Methanol | J.T.Baker | 9070-03 | |
| Screw cap tubes | VWR International | 53283-800 | |
| Screw cap tubes | Sun Sri | 13-425 | |
| Eppendorf tubes | Denville | C2170 |
Referanslar
- Balachandran, S., Xiang, Y., Schobert, C., Thompson, G. A., Lucas, W. J. Phloem sap proteins from Cucurbita maxima and Ricinus communis have the capacity to traffic cell to cell through plasmodesmata. Proc. Natl. Acad. Sci. USA. 94, 14150-14158 (1997).
- Barnes, A., Bale, J., Constantinidou, C., Aston, P., Jones, A., Pritchard, J. Determining protein identity from sieve element sap in Ricinus communis L. by quadrupole time of flight (Q-TOF) mass spectrometry. J. Exp. Bot. 55, 1473-1481 (2004).
- Behmer, S. T., Grebenok, R. J., Douglas, A. E. Plant sterols and host plant suitability for a phloem feeding insect. Functional Ecology. , (2010).
- Benning, U. F., Tamot, B., Guelette, B. S., Hoffmann-Benning, S. New aspects of phloem-mediated long-distance lipid signaling in plants. Front.Plant.Sci. 3, 53> (2012).
- Chen, S., Petersen, B. L., Olsen, C. E., Schulz, A., Halkier, B. A. Long-distance phloem transport of glucosinolates in Arabidopsis. PlantPhysiol. 127, 194-201 (2001).
- Corbesier, L., et al. FT protein movement contributes to long distance signalling in floral induction of Arabidopsis. Science. 316, 1030-1033 (2007).
- Dafoe, N. J., Gowen, B. E., &Constabel, C. P. Thaumatin-like proteins are differentially expressed and localized in phloem tissues of hybrid poplar.BMC. Plant Biology. 10, (2010).
- Deeken, R., Ache, P., Kajahn, I., Klinkenberg, J., Bringmann, G., Hedrich, R. Identification of Arabidopsis thaliana phloem RNAs provides a search criterion for phloem-based transcripts hidden in complex datasets of microarray experiments. The Plant Journal. 55, 746-759 (2008).
- Doering-Saad, C., Newbury, H. J., Bale, J. S., Pritchard, J. Use of aphid stylectomy and RT-PCR for the detection of transporter mRNAs in sieve elements. J. Exp. Bot. 53, 631-637 (2002).
- Fisher, D. B., Wu, Y., Ku, M. S. B. Turnover of soluble-proteins in the wheat sieve tube. Plant Physiol. 100, 1433-1441 (1992).
- Gaupels, F., Buhtz, A., Knauer, T., Deshmukh, S., Waller, F., van Bel, A. J. E., Kogel, K. -. H., Kehr, J. Adaptation of aphid stylectomy for analyses of proteins and mRNAs in barley phloem sap. J Exp Bot. 59, 3297-3306 (2008).
- Giavalisco, P., Kapitza, K., Kolasa, A., Buhtz, A., Kehr, J. Towards the proteome of Brassica napus phloem sap. Proteomics. 6, 896-909 (2006).
- Guelette, B. S., Chamberlin, B., Benning, U. F., Hoffmann-Benning, S., Benning, C., Ohlroggeeds, J. Indications of lipids/lipid signaling in the phloem exudates of Arabidopsis thaliana and Perilla ocymoides. , 92-95 (2007).
- Guelette, B. S., Benning, U. F., Hoffmann-Benning, S. Identification of lipids and lipid-binding proteins in phloem exudates from Arabidopsis thaliana. J. Exp. Bot. 63, 3603-3616 (2012).
- Haebel, S., Kehr, J. Matrix-assisted laser desorption/ionization time of flight mass spectrometry peptide mass fingerprints and post source decay: a tool for the identification and analysis of phloem proteins from Cucurbita maxima Duch. separated by two-dimensional polyacrylamide gel electrophoresis. Planta. 214, 3328 (2001).
- Hayashi, H., Fukuda, A., Suzui, N., Fujimaki, S. Proteins in the sieve element-companion cell complexes: their detection, localization and possible functions. Aust. J. Plant Physiol. 27, 489-496 (2000).
- Hoffmann-Benning, S., Gage, D. A., McIntosh, L., Kende, H., Zeevaart, J. A. D. Comparison of peptides in the phloem sap of flowering and non-flowering Perilla and lupine plants using microbore HPLC followed by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Planta. , 216-2140 (2002).
- Jorgensen, R. A., Atkinson, R. G., Forster, R. L., Lucas, W. J. An RNA-Based Information Superhighway in Plants. Science. 6, 1486-1487 (1998).
- Kehr, J. Phloem sap proteins: their identities and potential roles in the interaction between plants and phloem-feeding insects. J. Exp. Bot. 57, 767-774 (2006).
- King, R. W., Zeevaart, J. A. Enhancement of phloem exudation from cut petioles by chelating-agents. Plant Physiol. 53, 96-103 (1974).
- Lin, M. -. K., Lee, Y. -. J., Lough, T. J., Phinney, B., Lucas, W. J. Analysis of the pumpkin phloem proteome provides functional insights into angiosperm sieve tube function. Mol. Cell. Proteomics. 8, 343-356 (2009).
- Lough, T. J., Lucas, W. J. Integrative plant biology: role of phloem long-distance macromolecular trafficking. Annu.Rev. Plant Biol. 57, 203-232 (2006).
- Madey, E., Nowack, L. M., Thompson, J. E. Isolation and characterization of lipid in phloem sap of canola. Planta. 214, 625-634 (2002).
- Maeda, H., Song, W., Sage, T. L., DellaPenna, D. Tocopherols playa crucial role in low-temperature adaptation and phloem loading in Arabidopsis. The Plant Cell. 18, 2710-2732 (2006).
- Marentes, E., Grusak, M. A. Mass determination of low-molecular-weight proteins in phloem sap using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. J. Exp. Bot. 49, 903-911 (1998).
- Prado, E., Tjallingii, W. Behavioral evidence for local reduction of aphid-induced resistance. Journal ofInsect Science. 7, 48 (2007).
- Ruiz-Medrano, R., Xoconostle-Cázares, B., Lucas, W. J. Phloem long-distance transport of CmNACP mRNA: implications for supracellular regulation in plants. Development. 126, 4405-4419 (1999).
- Ryabov, E. V., Robinson, D. J., Taliansky, M. E. A plant virus-encoded protein facilitates long-distance movement of heterologous viral RNA. Proc. Natl. Acad. Sci. USA. 96, 1212-1217 (1999).
- Schilmiller, A. L., Howe, G. A. Systemic signaling in the wound response. Curr.Opin. Plant Biol. 8, 369-377 (2005).
- Thompson, G. A., Goggin, F. L. Transcriptomics and functional genomics of plant defence induction by phloem-feeding insects. J. Exp. Bot. 57, 755-766 (2006).
- Thorpe, M. R., Ferrieri, A. P., Herth, M. M., Ferrieri, R. A. (11)C-imaging: methyl jasmonate moves in both phloem and xylem, promotes transport of jasmonate, and of photoassimilate even after proton transport is decoupled. Planta. (11), 226-541 (2007).
- Truman, W., Bennett, M. H., Kubigsteltig, I., Turnbull, C., Grant, M. Arabidopsis systemic immunity uses conserved defense signaling pathways and is mediated by jasmonates. Proc. Natl. Acad. Sci. USA. 104, 1075-1080 (2007).
- van Bel, A. J. E., Knoblauch, M. Sieve element and companion cell: the story of the comatose patient and the hyperactive nurse. Aust. J. Plant Physiol. 27, 477-487 (2000).
- van Bel, A. J. E., Hess, P. H. Hexoses as phloem transport sugars: the end of a dogma. J. Exp. Bot. 59, 261-272 .
- Walz, C., Juenger, M., Schad, M., Kehr, J. Evidence for the presence and activity of a complete antioxidant defence system in mature sieve tubes. Plant J. 31, 189-197 (2002).
- Walz, C., Giavalisco, P., Schad, M., Juenger, M., Klose, J., Kehr, J. Proteomics of curcurbit phloem exudate reveals a network of defence proteins. Phytochemistry. 65, 1795-1804 (2004).
- Wang, Z., Benning, C. Arabidopsis thaliana Polar Glycerolipid Profiling by Thin Layer Chromatography (TLC) Coupled with Gas-Liquid Chromatography (GLC). J. Vis. Exp. (49), e2518 (2011).
- Will, T., van Bel, A. J. E. Physical and chemical interactions between aphids and plants. J. Exp. Bot. 57, 729-737 (2006).
- Yoo, B. C., Kragler, F., Varkonyi-Gasic, E., Haywood, V., Archer-Evans, S., Lee, Y. M., Lough, T. J., Lucas, W. J. A systemic small RNA signaling system in plants. Plant Cell. 16, 1979-2000 (2004).
- Zhang, B., Tolstikov, V., Turnbull, C., Hicks, L. M., Fiehn, O. Divergent metabolome and proteome suggest functional independence of dual phloem transport systems in cucurbits. PNAS USA. 107, 13532-13537 (2010).
- Zhang, C., Yu, X., Ayre, B. G., Turgeon, R. The Origin and Composition of Cucurbit "Phloem" Exudate. Plant Physiology. 158, 1873-1882 (2012).
- Zhang, S., Sun, L., Kragler, F. The Phloem-Delivered RNA Pool Contains Small Noncoding RNAs and Interferes with Translation. Plant Physiol. 150, 378-387 (2009).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır