
Method Article
Kulakçık ve karıncığa gelen kardiyak miyozitlerde Boru Membran Ağlarının Analizi
* Bu yazarlar eşit katkıda bulunmuştur
Bu Makalede
Özet
Kardiyak miyozitlerde, boru şeklindeki zar yapıları hücre içi ağları oluştururlar. Biz kalite kontrol, devlet-sanat-floresan mikroskop için ii) canlı hücre boyama dahil fare kalbinden miyositlerin i) izolasyonu için protokolleri optimize tanımlıyor, ve iii) doğrudan görüntü analizi bileşen karmaşıklığı ve hücre içi membran plastisite ölçmek için ağlar.
Özet
In cardiac myocytes a complex network of membrane tubules - the transverse-axial tubule system (TATS) - controls deep intracellular signaling functions. While the outer surface membrane and associated TATS membrane components appear to be continuous, there are substantial differences in lipid and protein content. In ventricular myocytes (VMs), certain TATS components are highly abundant contributing to rectilinear tubule networks and regular branching 3D architectures. It is thought that peripheral TATS components propagate action potentials from the cell surface to thousands of remote intracellular sarcoendoplasmic reticulum (SER) membrane contact domains, thereby activating intracellular Ca2+ release units (CRUs). In contrast to VMs, the organization and functional role of TATS membranes in atrial myocytes (AMs) is significantly different and much less understood. Taken together, quantitative structural characterization of TATS membrane networks in healthy and diseased myocytes is an essential prerequisite towards better understanding of functional plasticity and pathophysiological reorganization. Here, we present a strategic combination of protocols for direct quantitative analysis of TATS membrane networks in living VMs and AMs. For this, we accompany primary cell isolations of mouse VMs and/or AMs with critical quality control steps and direct membrane staining protocols for fluorescence imaging of TATS membranes. Using an optimized workflow for confocal or superresolution TATS image processing, binarized and skeletonized data are generated for quantitative analysis of the TATS network and its components. Unlike previously published indirect regional aggregate image analysis strategies, our protocols enable direct characterization of specific components and derive complex physiological properties of TATS membrane networks in living myocytes with high throughput and open access software tools. In summary, the combined protocol strategy can be readily applied for quantitative TATS network studies during physiological myocyte adaptation or disease changes, comparison of different cardiac or skeletal muscle cell types, phenotyping of transgenic models, and pharmacological or therapeutic interventions.
Giriş
Sağlıklı çizgisiz kas hücreleri, ana hücre eksenine dik "enine" oryantasyonundaki (T-tübül) ile boru şeklindeki zar yapıları bol miktarda bulunmaktadır. Sonuç olarak, T-tübülleri derin hücrenin merkezine doğru nüfuz sitoplazmada kas hücresi ana "yanal" yüzey zar (sarkolemma) sürekli uzantıları olarak karakterize edilmiştir. Dış yüzey membranı ile sürekli T-tübül fizyolojik rolü voltaj bağımlı L-tipi Ca nanometrik yakınlık bağlanması ile nispeten büyük bir kalp kası hücre hacmi boyunca SER organel temas alanları aracılığıyla oluşturulan uzak hücre içi bölmelerin elektrik bağlantı hızlı bir 2 + kanalları komşu riyanodin reseptörü (RYR2) SER Ca 2 + bültenleri aktive akımı (I Ca) içeri (Cav1.2). Junctional SER alanları ve T arasındaki ventriküler miyositlerde (VM), sürekli olmayan membran kontakları ("kavşaklar") olarak-tubules her hücrede 1 tek tek hücre içi Ca2 + serbest nanodomains binlerce kontrol edildiği düşünülmektedir.
Herhangi bir temas alanı için, yan yana zar bölümleri, T-tübül içinde ve çevre birimini (kavşak) SER, her birbirine yakın 15 nm, dolayısıyla nanodomain olarak tanımlanır yaklaşık olarak. Böylece, çok küçük bireysel sitoplazmik alt uzaylar yarı hücre özerk bölmesi davranışları sağlayacak hangi ayrılmış edilmektedir. Gelen bir hareket potansiyeli sanal makineye, nispeten küçük bir Ca T-tübüllerde Cav1.2 kanallarını aktive olduğunda 2 + içe akım hızlı attoliter boyutlu nanodomain 1 alt uzay Ca2 + konsantrasyonu [Ca2 +] S artacaktır. Sonra, [Ca 2 +] S artış 2+ -geçişli riyanodin reseptörleri (RYR2) yanyana SER membran kavşağında nanometre yakınlık içinde Ca harekete geçirir ve bu birleştirme işlemi, tüm elektriksel çift boyunca oluşurd miyositlerdeki nanodomains. RyR2s 5-10 RYR2 kanal 2 1 Cav1.2 kanal tahmini stokiyometri ile yoğun olarak çok kanallı kümeleri ortaya çıkar. SER-to-sitozolde beri [Ca 2 +] degrade çok dik (oranı 10 4: 1) ve RyR2s işlevsel birleştiğinde kümeler gibi yüksek iletkenlik Ca 2 + salım kanalları işlev, bir nicel büyük Ca RYR2 aktivasyon sonuçları 2+ 1-2 msn 3,4 olan 100 uM veya daha yüksek [Ca 2 +] S yerel altuzaya artan T-tübül birleştiğinde kavşak SER etki akım bırakın. Bu, kalp sinyal amplifikasyonu davranış, serbest Ca 2 + (KTKS) indüklenen Ca2 + olarak ifade edilir. Birlikte ele alındığında, T-tübülleri hızla eksitasyon-kasılma (EC) birleşme sırasında kavşak nanodomain SER kişileri ve hücre çapında KTKS aracılığıyla Ca 2 + sürümü sinyalleri aktive temel membran yapılar vardır.
T-tübül ek olarak, eksenel tübülana (uzunlamasına) hücre eksenine önemli ölçüde farklı bir yönelim paralel lar (A-tübüller) elektron mikroskobu (EM), confocal ve 2-foton mikroskopi çalışmaları ile belgelenmiştir. Örneğin, sarkomer Z hatlarının yakınında, T-tübül ile birbirlerine bağlanmış myofibrils arasında bir-tübüllerin bir hücre-hücre-dışı olarak geniş ve sürekli kafes ajanları ve sabit kobay VM 5 EM görüntüleme ile gösterilmiştir. Ekstrasellüler dekstran-bağlantılı floresein boyası kullanılarak ve sıçan VM'lerin 2-foton görüntüleme yaşamak, karmaşık bir retiküler 3D borucuk ağı ~ 60% T-tübüller ve ~ 40% A mikrotüplerin 6 oluşan görüntülendi. Bu çalışmada yalnızca, aynı zamanda EM görselleştirme için kesit doğal enine-eksenel borucuk sistemi (TATS) gibi karmaşık ve dinamik zar ağlarının analizi için sınırlı olduğunu gerçekleştirilmesi, bol A-Tübüllerin 3D görselleştirme yol değil. Sonuç olarak, TATS membran konfokal canlı hücre görüntüleme doğrudan ile boyanmış di-8-ANNEPS geliştirilmiştir. Eğer canlı hücreTATS ağlar Fourier dönüşümü ile analiz edilir, sarkomer Z hatlarının yakınında alanı T-tübül bileşenlerinin normal görünümünü çizgili sinyalleri 7 bir bölgesinden topluluğu güç spektrumu ile yansıtılır. Bu strateji, dolaylı analiz hastalık modellerinde 7 TATS bileşen düzenli hücre genelinde bölgesel değişiklikleri tespit etmek için kullanılmıştır. Örneğin, shRNA aracılı junctophilin-2 knock-down kalp yetmezliği ve izoformu spesifik protein eksikliği nanodomain Ca 2 + sürümü disfonksiyonu 8 T-tübül sonuçlanan yeniden neden oldu. Biz son zamanlarda doğrudan nicel yaklaşımlarla ve daha fazla uyarılmış emisyon tükenmesi (STED) nanoscopy 9 kullanılarak fare VM'lerde bireysel TATS bileşenlerin canlı hücre superresolution mikroskobu ile TATS membran ağlarının analizini artırdık. Enine 50:50 dağılımın yaklaşık küçük bireysel TATS bileşenlerin doğrudan analizi, izin nanometrik görüntü çözünürlüğüeksenel tübül yönelimleri karşısında, kantitatif sağlıklı fare kalpleri 9 iki bol henüz diferansiyel yönelimli bireysel TATS bileşenlerini doğrulayan. Bu stratejiler daha aşağıda protokol bölümünde özetlenen edilecektir.
Yetişkin merkezinde bol miktarda bir-tübül bileşenlerinin fizyolojik rolü anlaşılmaz kalmıştır da EM çalışmaları endojen Ca 2 +, gine domuzu ve sıçan VM 5,10 açma nanodomains süren bir-tübül ilişkili SER zar yapılarına belgelemiştir. Cav1.2 ve RYR2 Konfokal analizi, bir-tübül birleşme 10 ile yardımcı sınırlama yüksek derecede bulundu. VM T-tübülleri genellikle oluşmaz Z-line damarların nispeten uzak kökenli sıçan spontan Ca 2 + kıvılcımlar ~ 20% yana, tek argüman A-tübül ilişkili nanodomains gerçekten var ve fonksiyonu olabileceğini olmuştur Ca 2 + sürümü siteleri gibi 11,12. İlginç bir şekilde, T-tübül oluşumu ve olgunlaşma OCsadece doğumdan sonra sadece alıcılarında mümkündür ve P5 ile ön-madde sarkolemmal invajinasyonları filizlenerek yoluyla, örneğin kalp hücreleri, büyüme, paralellik ve olgunlaşmamış farelerde 13 P10 TATS ağ derlemeleri dallıdır. Bu Junctophilin-2 ShRNA knock-down beri doğum sonrası TATS ağ olgunlaşması için özellikle önemlidir gecikmeli Ca 2 + salınımı ve olgunlaşmamış bir-tübül hakim mimarileri ile tutarlı patolojik TATS örgütü lider SER kavşaklar T-tübül membranların ankrajını engelledi görünür VM 13. Bu gözlemler sonuçta kanıt-of-concept T-tübülleri ek ya da alternatif hücre içi mekanizmaları ile 14 morph olabilir A-tübüller ise membran invaginasyon süreçlerle oluşturmak yol açabilir.
Kalp hastalığı TATS membran değişikliklerinden Karakterizasyonu patofizyolojik sorular için önemli bir araştırma alanı haline gelmiştir. Pacing kaynaklı kalp failu bir köpek modelinde ilk raporlarYeniden T tübüller ve Cav1.2 akımı (I Ca) 15 bir kayıp göstermiştir. Iskemik kardiyomiyopati bir domuz model T-tübül yoğunluğu azaltılmış gösterdi ve hücre içi Ca azaltılmış sinkronisinde 16 serbest 2+. Kalp yetmezliği, kendiliğinden hipertansif sıçan (SHR) modeli kullanılarak T-tübüllerin bir kayıp "orphaning RYR2" nin bir mekanizma yolu ile Cav1.2 ve RYR2 azaltılmış nanodomain bağlanması ile ilişkili 7. T-tübül kaybı da iskemik dilate, hipertrofik kardiyomiyopati örnekleri 17 insan sanal makine içerisine gösterilmiştir. Ayrıca, A-tübül bir artışın insan dilate kardiyomiyopati 18 doku kesitlerinde bildirilmiştir. Miyokard enfarktüsü takiben, A-tübül bileşenlerin 9 artar aksine T-tübüller önemli düşüşlere fare sanal makinelerdeki TATS yeniden bir diferansiyel mekanizması gösterdi. Önemli bir şekilde, geliştirilmiş zar yerel kontrast canlı hücre superre yoluyla eldeçözüm STED'in mikroskobu genel TATS ağ uzunluğu artar ve karmaşıklığını 9 dallanma ile bir-Tübüllerin belirgin bir çoğalma gösterdi doğrudan ölçümler aracılığıyla detaylı nicel bileşen analizi sağladı. Ayrıca, miyokard infarktüsü sonra 19 ve KRT atriyal tachypacing kaynaklı kalp yetmezliği 20 köpeklerde T-tübül tersine yeniden biçimlenme yol açabilir sıçanlarda T-tübül biçimlenme ters olabileceğini egzersiz eğitimi gördü. Protokolde belirtilen ve aşağıda bölümleri sonuçları birlikte ele alındığında, hastalıklı insan ve hayvan VM'lerin yanı sıra potansiyel terapötik müdahalelerin hem çalışmalar tartışmasız kaliteli hücre izolasyon prosedürleri ve ayrıntılı kantitatif analiz stratejilerinden yararlanacaktır.
Katp kanal TATS membran kaçakçılığı 21 izoformlan karşı yanal yüzeyi ile son zamanlarda gösterdiği gibi Dahası, atriyal m dikkate almak önemlidiryocytes biyolojik olarak farklı olarak sanal makineye karşı karşılaştırmalı kalp hücre modeli olarak (AMS). T-tübüller yakın koyun ve insan AM'ler 22 belgelenmiştir. Mevcut kanıtlar birkaç T-tübülleri küçük kemirgenler 23 koyun ve insanlarda değil gibi büyük memelilerde tipik PM hücrelerde bulunur ve düşündürmektedir. VM'lere aksine, ams hücre içi Ca2 + serbest 2 + işaretli mekansal ve zamansal Ca ile sonuçlanan 23 gradyanlar hücrenin merkezine doğru difüzyon ile hücre yüzeyi ilerleyen meydana gelir gibi görünmektedir. Bu çerçevede bu tür atriyal fibrilasyon 24 gibi hücre içi Ca 2 + yaygın bir hastalık formları için istikrarsızlık sinyal mekanizmaları aydınlatmak için önemli gibi görünüyor. Özetle, sağlıklı ve hastalıklı kalpleri hem AM ve VM hücre izolasyonu ve genellikle her protokolleri kullanılmaktadır. Yeterli hücre kalitede mikroskobik belgelerine göre değerlendirildiği gibi hücre izolasyonu düzgün yapılır Sadece eğer, AM ve VM örnekleri ca olmalıKantitatif analiz TATS ileri rried. Bu duruma göre, aşağıdaki protokol kesitleri fare veya sağlam zarları TATS analiz etmek için bir canlı hücre mikroskobu ile takip diğer türlerden izole edilen yüksek kaliteli hücre kritik bağlıdır. Daha önce de belirttiğimiz gibi, TATS membranların karakterizasyonu tespit ve hazırlık eserler 6, ozmotik değişikliklere bağlı membran değişikliklerinden ve geleneksel ışık mikroskopisi 9 çözünürlük sınırlamaları için bir eğilimi ile zorlu bir araştırma alanıdır. Biz Ca 2 + görüntüleme ve yama-kelepçe ve hücre kültürü için sıçan VM'lerin insan AM'ler izolasyonu için son state-of-the-art protokolleri daha önce bu dergide 25,26 yayınlanmaktadır unutmayın.
Protokol
NOT: Tüm hayvan prosedürleri gözden ve laboratuar hayvanlarının insancıl bakım ve kullanım ile uyumlu Üniversitesi Tıp Merkezi Goettingen Kurumsal Hayvan Bakım ve Kullanım Kurulu tarafından kabul edildi.
Fare Kalpten Atriyal ve Ventriküler miyositlerin 1. İzolasyonu
- Mümkün olduğunca nazik hayvanları idare ve onaylanmış protokollere göre izole kardiyak hücreleri üzerinde Nörohormonal aşırılıkları potansiyel güçlü istenmeyen etkilerini önlemek için özellikle genel stresi en aza indirmek ve. Buna ek olarak, heparin (500 IE / kg vücut ağırlığı sc) belirgin bir izolasyon sırasında kalp hücreleri veriminin ve bütünlüğünü tehlikeye sokabilir ve kan pıhtılaşmasının önlenmesi için mikro embolilerin önce kalp ekstre en az 20 dakika, her fare enjekte edilir.
- , 12 hafta veya izofluran inhalasyon yoluyla üzeri fareler anestezi ağrı çekme reflekslerin yokluğu onaylamak ve servikal dislokasyon ile hayvanları euthanize.
- Daha önceki çalışmalarda hızlı uzman protokoller izlenerek, kalp Extract (örneğin, Kaestner et al. 26 ve Louch ve ark. 27). Gereksiz sıkma veya germe ile kulakçıklar herhangi kasıtsız zarar kaçının.
- Dikkatle başarılı kanülasyon ve kalp perfüzyon için önemli olan aort damar duvarında, kesintisiz bir çapraz kesme kenarı kurmak güdük forseps ve düz bir makas kullanarak proksimal asendan aort dokusunun korunması.
- (Tablo 1 e bakınız çözeltiler için) buz soğukluğunda hemen içine nominal olarak Ca 2 + serbest perfüzyon tamponu çıkarıldı kalp aktarın. Kalp tamamen batık kadar istenmeyen hava embolisi önlemek için tampon içine hava yoluyla transferi sırasında sıkıştırılmış büyük damarları tutun. Kalp kasılmasının inhibe etmek için, tampon çözelti ve buzla soğutulmuş Çözeltilerin BDM kullanın.
- Yeterli 3D illum ile binoküler mikroskop zoom kullanınasÙ.
- Tamamen tamponu ile dolduruldu gereken; (fare normal kalp ağırlığı dış çapı 0.81 mm), pürüzsüz bir yüzey ile kalbin panoramik stereovision altında aortu Cannulate 21 G kanül cilalı. Hızlı çözüm akış kontrolü sağlayan bir 2-yollu Luer vanadan bir yakın çözüm deposunu (örneğin, bir şırınga) bağlayarak kanül içinde hiçbir hava kabarcığı olmadığından emin olun.
- Kanül aort ve koroner arter dalları üzerinde yaklaşık 1 mm aort, içine doğru şekilde yerleştiğinden emin binoküler büyütme altında onaylayın. Kesinlikle herhangi bir geçişi veya kanül ile aort yanlışlıkla delinme (bu kalıcı aort kapak kapanması bozabilir ve dolayısıyla kardiyak perfüzyon bozacak) kaçının.
- İki ipek sütür kullanarak kanül sonuna ısmarlama, çevresel odaklı kaymaz oluklar yavaşça aort bağlayın. Koroner arterlerin yıkayın etmeyinherhangi bir noktada güçlü bir şekilde es. (; Aşağıda ayrıca tartışma bölümüne bakın ya sabit basınç veya sabit akışını kullanarak) çözelti dolu bir özelleştirilmiş ve önceden kalibre perfüzyon sistemi sıkı bir şekilde uyan çıkış konnektörüne aorta ve kalbe bağlı kanül, modifiye Langendorff kurulum aka bağlayın.
- 37 ° C (4 ml / dk bir hedef perfüzyon oranı), soğuk oksijenasyonlu perfüzyon tamponu kullanılarak 4 dakika boyunca mümkün olan en kısa olarak kalp serpmek. 37 ° C'de, 8-10 dakika boyunca sindirim tamponu (600 U / ml kolajenaz tip II) ihtiva eden kolajenaz perfüzyon geçerek sindirim başlatın. Bariz kalp yüzeyi boyunca artan opaqueness, yumuşaklık, ve gevşekliğe dahil olmak üzere benzer doku değişikliklerini onaylayarak doku sindirim ilerlemeyi izleyin.
- Gerektiği şu sindirim gibi kardiyak odaları teşrih. Binoküler mikroskop altında kanüle kalbi yerleştirin ve arka kalp duvarı görselleştirmek. Kalıntı kardiyak olmayan doku, örneğin, akciğer a teşrihnd damar parça mikro makas kullanılarak sindirim tamponu hücre kontaminasyonu önlemek için (örneğin, 8 mm düz bıçakları ile yaylı makas), Şekil 1'de gösterildiği gibi.
- Kollajenaz sindirilmiş kalbin belli odaları, bölgeler ve / veya hücrelerin hasat edilmesi gereken bir kontrol listesi (Şekil 1) takip edin: ve / veya sağ atrium, ücretsiz bir sol ve / veya sağ ventrikül duvar ve / veya ventriküler septum bıraktı.
- Belirli bir kardiyak dokularda diseksiyonundan için, silikon elastomer plastik bir kaç mm kalınlıkta bir tabaka ile kaplı nispeten geniş ve düz diseksiyon banyosu kullanın. Alt elastomer tabakası ince bir böcek çelik iğne ile kalbin apeks Fix.
- Sağ atriyal apendiks saptırmak ve sadece atriyoventriküler kapakların üzerinde sağ atrium teşrih. Sol atrium ile diseksiyon devam edin. Inceleyin ve elyaflı valf düzeneği atın. Son olarak, sol ve sağ ventrikül serbest duvarları ve septum ve / veya küçük teşrihdoku parçaları er gerektiği gibi.
- Stajyerler için sadece Not: non-sindirilmiş fare kalpleri ile başlar, pratik kazanmak için. Anatomik oryantasyon kolaylığı için, kollajenaz sindirilmiş fare kalpleri gibi devam, Şekil 1'de gösterildiği gibi binoküler görüş altında. 3D anatomi sonra elle taşıma binoküler görüş altında tüm ardışık diseksiyon adımlar dahil olmak üzere doku işleme uygulama ve diseksiyon adımlar yeterince tanıtılır Yukarıda özetlenen.
- Ventriküler miyositlerdeki (VM) hücre izolasyonu için: taze sindirim tamponu 2.5 ml ventriküler doku transferi. Birkaç organ parçaları, örneğin, kulakçık ve karıncığa eşzamanlı hücre izolasyonları teşebbüs edilirse, ikinci bir kişi, hücre ayrışma prosedürü tek bacak üzerinde sürebilir hem en aza indirmek ve çoklu kardiyak dokuların koordine elleçleme farelerin kullanımını optimize etmek. Adımlar bundan sonra 1.10 1.9.1-1.9.4 ile devam edin.
- Tüm ventriküler doku ya teşrihveya belirli bunların parçaları (örneğin, LV, RV, ücretsiz duvar ve / veya septum) keskin bir makas kullanarak 2.5 ml sindirim tamponu yaklaşık 1 mm 3 parçaya (örneğin, 8 mm düz bıçaklı yaylı makas) 60 mm Petri kabındaki .
- Yavaşça bir transfer pipeti ile doku parçalarının içinde ezilerek karıştırma, yavaş bir hücre süspansiyonu içine VM ayrışır. Hücre süspansiyonu herhangi bir hava kabarcıklarının oluşması kaçının.
- VM hücre süspansiyonu durdurma tamponu 8 ml ilave edilir ve 15 ml konik bir tüp içine hücre süspansiyonu aktarın. Kalan doku parçaları yaklaşık 15 saniye boyunca altta yerleşmek için izin, ama süspansiyon kalması izole edilmiş hücreler için yeterince kısa. Sonra, yeni bir 15 ml tüp süpernatant hacmi havalesi yoluyla VM süspansiyon hasat. Aşırı doku parçaları mevcut ise, alternatif olarak, hücre süspansiyonundan, doku parçaları ayırmak için az 200 mikron aralıklı naylon örgü kullanılır.
- VM hücre süspansiyonu, 15 ml eş dibe Let8 dakika boyunca yerçekimi ile nical tüpü.
- Adım Yıkama: süpernatant kaldırmak ve yavaşça perfüzyon tamponu, 10 ml geri kalan VM pelletini. Tekrarlayın yıkama adımı 1.9.5 (opsiyonel: yavaş yavaş gerektiği gibi Ca 2 + konsantrasyonunu artırmak için ek yıkama adımları ekleyebilirsiniz).
- 10 mi perfüzyon tampon maddesi içinde yerleşmiş VM pelet yeniden süspanse edin ve 1.5 ml mikrosantrifüj tüplerine (tüp başına yaklaşık 50,000 hücre VM) kalan hücre süspansiyonu hacmi dağıtın.
- Atriyal miyosit (AM) bir hücre izolasyonu: temiz sindirim tamponu 1 ml sindirilmiş / kesilmiş atriyal doku aktarın.
- Küçük bir petri kabının (örneğin, 60 mm çap) mikro makasları kullanılarak 1 mi sindirim tamponu içinde yaklaşık olarak 1 ile 3 mm parçalar halinde, kısmen sindirilmiş atriyal doku kesilir. Hafifçe zarar sıvı jetleri önlemek için bir kesme ucu olan bir 1 ml'lik plastik bir pipet ile öğütülmesinden kullanılarak hücre süspansiyonu içine Sindirilmiş doku parçaları üzerinden PM hücreleri ayırmak. SırasındaTritürasyon sıkı hücre süspansiyonu herhangi bir hava kabarcıklarının oluşması kaçının. Hücre süspansiyonu içinde herhangi geri kalan kollajenaz aktivitesini durdurmada (% 10 BCS 50 uM CaCl2) mekanik ajitasyon sonra, durdurma tamponu 4 ml.
- 15 ml konik tüp içine AM hücre süspansiyonu aktarın. Kalan doku parçaları yaklaşık 15 saniye boyunca altta yerleşmek için izin, ama süspansiyon kalmaya izole hücreler için yeterince kısa. Yeni bir 15 ml tüp çözüm havalesi yoluyla ücretsiz AM hücrelerini içeren süpernatant hacmi hasat.
- Oda sıcaklığında PM hücre süspansiyonu, örneğin, 20 x g'de 2 dakika santrifüj veya - tercihen membran çalışmaları için - hücreler, 15 ml konik bir tüp içinde 20 dakika boyunca yerçekimi ile yavaş yavaş yerleşmek sağlar.
- 5 ml perfüzyon tamponunda AM pelet hafifçe süpernatantı atmak ve: adım yıkayın. 1.10.4 tekrarlayın.
- Hücreler nazikçe yeniden süspanse PM 5 mi perfüzyon durdurulmuştur. 1.5 ml mikrosantrifüj küvet içinde hücre süspansiyonu hacmi dağıtmaES (tüp başına yaklaşık 1.000 PM hücreleri) içerir.
- Analiz ve tripan mavi lekelenmesi kullanılarak hücre verimi dahil olmak üzere her kalp için izole edilmiş hücre popülasyonu kalitesini belgelemek.
- Bunun için, 1 gibi hücre süspansiyonu 500 ul seyreltilmiş: 1 mi kesme pipet uçları kullanılarak 1 hacim / hacim ile, tripan mavisi çözeltisi (nihai konsantrasyon% 0.02). Çok / aşağı pipetlenmesini kadar yavaş yavaşça hücreleri ve mavilerin karıştırın. Hemen sitometresi geliştirilmiş bir Neubauer-tipi hücre süspansiyonu ihtiva eden tripan mavisi uygulamak ve bir ters mikroskop kullanılarak bütün miyositlerin sayısı.
- Belirgin hasarına, zar kabarcıklannın, hücre içi, tripan mavisi (Şekil 2) biriken kesintiye şeritlerin, kontraktürleri, ve hücreler ile herhangi bir hücre hariç. Ayrıca, kendiliğinden daha sonra hücre ölümüne karşı hassas olan hücrelerin, sözleşme dışlama. Süspansiyon bozulmamış hücrelerin sayısını değerlendirmek için, genelinde mavi tripan hariç düzenli şeritlerin sadece kardiyak miyositleri kullanınHücre hacmi.
- Iletilen ışık mikroskobu ile bireysel atriyal ve ventriküler miyositlerin bütünlüğünü yargıç. Dokümantasyon ve daha fazla analiz için bir tif dosyası olarak parlak bir alan görüntü kaydetme.
- Kardiyak hücre bütünlüğü analizi için aşağıdaki kriterleri kullanın:
- Görünür hücre hacmi boyunca düzenli şeritlerin varlığını doğrulamak;
- Her iki hücre tarafta yanal yüzey zarı myofilaments paralel sürekli bütünlüğünü doğrulamak;
- spesifik yüzey zar yapılarına bütünlüğünü yansıtan iki hücre tarafta birleştirilmiş diskler keskin izlerin görselleştirmek; ve
- aşağıdaki yatan hücreye özel parlak alan görüntü morfolojisi (örneğin, bileşik, bindirme imag olarak ImageJ her iki görüntüleri birleştirmek için lokalizasyon korelasyon bölüm 3'de tarif edildiği gibi TATS membran flüoresan sinyal (ya da imüno-kaveolin-3 protein ya da diğer zar belirteçler) görselleştirmeke).
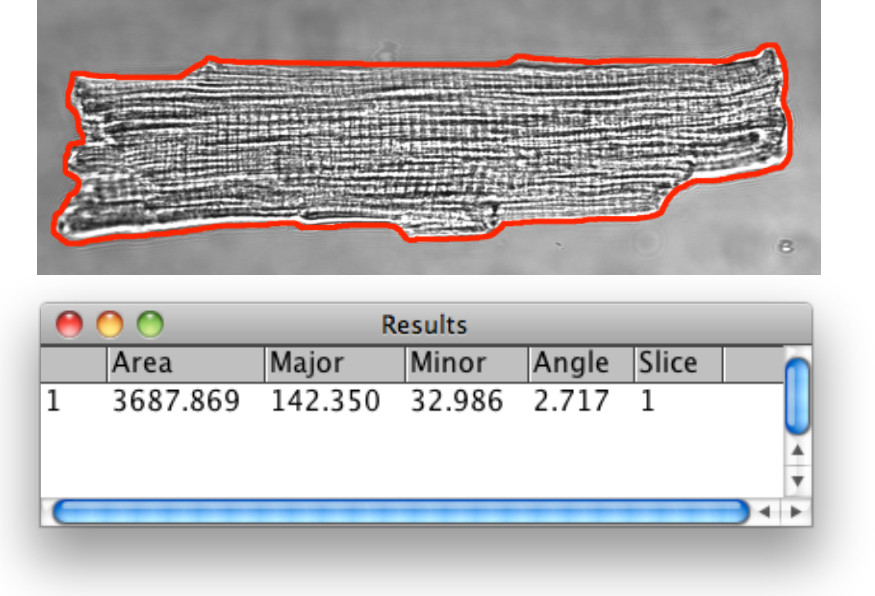
- Parlak bir alan görüntülerden sarkomer uzunluğunu belirleyin. Ortalama sarkomer uzunluğu için, sırayla hizalanmış sarkomer şeritlerin mesafeyi ölçmek ve sarkomer sayısına göre mesafeyi bölmek. Hücre başına en az iki konum ölçün. ImageJ ile ticari yazılımı veya çevrimdışı ile analiz gerçekleştirin.
NOT: ayrılması maddeler ile tedavi durumunda, fare kalp bozulmamış rahat VM ~ 1.9 um 28 bir ortalama sarkomer uzunluğunu gösterir.
- Kardiyak hücre bütünlüğü analizi için aşağıdaki kriterleri kullanın:
- Iletilen ışık mikroskobu görüntü morfolojisi ve bireysel atriyal ve ventriküler miyositlerin boyutlarını ölçmek. AM ve VM hücreler boyutu önemli ölçüde farklı olduğunu düşünün. Hücre uzunluğu, genişliği ve alanı ölçün ve uzunluğunu hesaplamak: genişlik oranını.
- Komutlar çokgen seçim aracını kullanarak ImageJ iletilen ışık görüntüden 2D hücre boyutlarını analiz ve Şekil 3'e, ROI yöneticisi benzer ekleyin. Morfolojik c iseHanges spesifik çalışma bağlamlar içinde bekleniyor, tüm hücreler için başka belge özel fare suşu, yaş, cinsiyet, kalp boyutu ve sonraki veri sınıflandırma için herhangi müdahaleler.
Yaşam Atriyal ve Ventriküler miyositlerde TATS Membranlarının 2. Boyama
- Görüntüleme odasına yerleştirin (örneğin, POC-R2) 42 mm cam lamel ile. Lamel kararlı miyosit tutunma için, fizyolojik perfüzyon tampon maddesi (nihai konsantrasyon 0.2 mg / ml) içinde, laminin stoklar 1:10 seyreltme laminin solüsyonu 20 ul hazırlama. Cam lamel üzerinde eşit laminin solüsyonu 20 ul yayıldı.
- Perfüzyon tampon maddesi içinde bir 50 uM, di-8-ANEPPS çözeltisi 800 ul hazırlayın. Bunun için, 780 ul fizyolojik tampon içinde, 2 mM di-8-ANEPPS stok solüsyonu, 20 ul seyreltilmiş.
- VM leke amacıyla hücreler, bir 1.5 ml'lik tepkime tüpü içinde, 8 dakika boyunca yerçekimi ile razı. AMS leke, ya yerçekimi sedimantasyon veya sp kullanmak için2 dakika boyunca hücre süspansiyonu (1.10.3 bakınız). Hücre topağı gereksiz karıştırmadan kaçınarak AMS VMS her ikisi için de, dikkatli bir şekilde süpernatant kaldırmak ve yavaşça 800 ul çözeltisi (50 uM) ihtiva eden di-8-ANEPPS hücre pelletini. Hemen görüntüleme odasında laminin kaplı lamel üzerine di-8-ANEPPS / miyositlerdeki süspansiyonu transfer.
- Karanlıkta oda sıcaklığında 15 dakika için VM süspansiyon Leke.
- Yavaşça elle pipet ile görüntüleme odasının iki yanında yukarı doğru sıvı menisküs ile fazla hacmi çıkarın. Çoğunluğu di-8-ANEPPS lekeli miyositler sıkıca laminin kaplanmış lamel bağlı kalır ve havaya maruz görmemesine emin olun. Daha sonra, yavaş yavaş yapışmayan hücreler dahil olmak üzere herhangi bir fazla sıvının uzaklaştırılması, ardından perfüzyon tampon maddesi 1 ml ilave bir kez bağlı miyosit süspansiyon yıkayın.
- Dikkatle yavaş yavaş t yan perfüzyon 1 ml tampon ile boyanmış ve yüzey ekli miyositleri Bindirmeo haznesi görüntülenmesi. Mikroskop sahnede görüntüleme odasına yerleştirin.
Yaşam Atriyal ve Ventriküler miyositlerde TATS Membran Yapıların 3. Görüntüleme
- Genel olarak, dikkatle TATS membran görüntüleme için mümkün olan en iyi floresan mikroskop seçeneği (ler) seçin. Konfokal görüntüleme için, son nesil optimize PMT dizi dedektörleri ve floresan sinyal yoğunluğu maksimize foton geri dönüşüm yolları ile modern bir floresan mikroskop düşünün. Tats küçük detayların konfokal görüntüleme için membran yapılar 63X 1.4 NA yağ objektif veya kullanmayın - durumuna göre -. Yüksek genel ilkeleri için Kohl ve arkadaşları 34 miyositlerin özel uygulamalar için yorum olarak küçük TATS detaylar için bir STED'in superresolution mikroskop kullanmak çözünürlüklü floresan mikroskopi, tartışma bölümüne bakın.
- , Ideal tüm di-8-ANEPPS lekeli hücre içi mem en algılamak için görüntüleme parametrelerini ayarlayınBelirli bir görüntü düzlemi içine miyosit membranlar. Maksimal lazer gücünün% 3, uyarma 458 nm örneğin;: odaklı lazer tarama mikroskopisi için başlangıç noktası olarak aşağıdaki parametreleri kullanın 550 nm ve 740 nm arasında yayılan sinyalin tespit edilmesi; dedektör kazanç (örneğin, ana komut 800); ve 900 nm'lik bir optik kesit kalınlığı için, iğne deliği 1 AB. Sinyal-gürültü optimize etmek bu parametreleri ayarlayın.
- Uygun (Şekil 4A ve 4B) olarak bozulmamış bir AM veya VM hücreyi seçmek için parlak bir alan modunu kullanın. Hücre çapında düzenli damarların ve eşit sarkomer aralığı, keskin yüzey kenarları tırtıklı ve dört hücre taraftan, yanal yüzey zarı sürekli bütünlüğü ve yokluğu: özetlendiği gibi hücreleri seçmek için hücre bütünlüğünü yargılamak için nasıl ilgili kriterleri 1,12 işaret bakın Herhangi bir zar kabarcıkları.
- Merkezi bir hücre içi miyositlerdeki bölümünde örnek bir resim çekin. ROI "hasat" fonksiyonu → kullanmak ayarlamak için, Son piksel boyutu 100 nm x 100 nm ölçen → kırpma penceresini ayarlamak. Miyositinin önemli (eksenel / boyuna) ekseni ile karşılık kırpma pencerenin x-eksenini ayarlayın.
- Nihai görüntüleme düzlemi seçin. Elle z-doğrultusunda uygun görüntüleme düzlem seçmek için tek bir görüntü karelerinin kullanın. T-tübül ve A-tübül bileşenler dahil TATS membranlar, odak düzleminde görsel olarak belirgin olduğunu onaylayın. Tipik bir hücre içi görüntüleme düzlemi içi referans noktası olarak çekirdeğini de olabileceğini unutmayın. Şekiller 4A ve 4B'de örneklere bakınız.
Not: genel olarak, mümkün olduğu kadar kısa lazer ışığına maruz kalmasını hücre. Mümkünse, kalp miyositlerde optimum odak düzlemi belirlemek için tek resim çerçeveleri kullanın. - Yaklaşık 0,5 mikro saniye olduğu için piksel bekleme süresini ayarlayın. 16x ortalamasını seçin ve anlık olarak görüntüyü kaydedin. Uygun görüntüleme düzlemi a kurmak için görüntü anlık adımı yineleyinGerektiğinde 3,5 olarak TATS zar yapılarına için belirtilen s.
- Son görüntüyü kaydedin ve dosya hedef klasörde kaydedilmiş onaylamak. Genel olarak, analiz yazılımı düzgün uygulanması için aynı formatta (örneğin, EKK) tüm resim dosyalarını kaydedin. Önceden herhangi bir görüntü analizi, bir kez daha (adım 1.12.1 altında listelenen kriterleri dikkate) off-line yeterli hücre bütünlüğünü onaylamak ve analizi hasarlı hücreleri hariç. Şekil 4C ve 4D örneklere bakınız.
TATS Membran Network ve Bileşenlerinin 4. Analizi
TATS membran bileşenlerin doğrudan analizi için aşağıdaki görüntü işleme adımları Şekil 5A ve 5B yukarıdan aşağıya akışı diyagram olarak özetlenmiştir.
- Fiji (http://fiji.sc/) bir di-8-ANEPPS lekeli miyositinin görüntü dosyasını açın, ImageJ bir serbestçe kullanılabilir varyant için gerekli analiz eklentileri içerengörüntü işleme. Daha fazla bilgi için Schindelin ark 29 bakın.
- Kaydet → Tif → görüntü Dosya → kaydedin.
- TATS zar bileşenlerini analiz etmek için, gösterildiği gibi, daha sonra İB sınır dış yüzey membran (sarkolemma) hariç ve TATS membran kısımları dahil olmak üzere hücre içi tanımlamak için "çokgen seçim" aracını kullanarak, dış yüzey zar sinyali dışında uygun ROI seçmek Şekil 5A (ROI). → Araçlar → ROI Yöneticisi analiz uygulayarak "ROI Manager" seçilen ROI ekleyin.
- , TATS bileşenlerin yönlendirme analizi için özel bir ROI seçmek ana eksene ve uzunlamasına, hücre paralel olarak görüntü x ekseni hizalayın. Hücre hafif kavisli ise, birkaç İB'leri seçin ve tek tek her ROI hizalayın. Analiz herhangi çekirdekleri hariç. Analiz herhangi bir aşırı kavisli hücreleri hariç, çünkü Ar hassas hizalamaOIs giderek daha zor hale gelir ve analiz sırasında yönlendirme hataları artacaktır.
- Dış yüzey zarı istenmeyen sinyal bilgilerini sil: Düzen → Temizle Dışında "seçilen ROI" (Şekil 5A) oluşturmak için. Seçilen ROI sadece TATS ağ ile karşılık hücre içi membran bölümlerini, içerdiğinden emin olun.
- Şekil 5A'da belirtildiği gibi daha sonraki kantitatif analiz öncesinde görüntü işleme adımlarla (komutları) aşağıdaki zinciri gerçekleştirin.
- Çıkart Arka → → Süreci tıklayın. 5 piksel Rolling top yarıçapı ayarlayın.
NOT: Görüntü analiz 100 nm x 100 nm bir piksel boyutuna sahip olması durumunda 5 piksel yuvarlanan bilye yarıçapı ayarlayın. Diğer piksel boyutları için yaklaşık 500 nm arasında bir fiziksel yarıçapına karşılık gelen piksel sayısı ile yuvarlanan bilye yarıçapı ayarlayın. - Yerel Kontrast geliştirin → → Süreci tıklayın (KLAHE). , 256, 49'a 3, maksimum eğim ve "yok" maske histogram kutuları blok boyutunu ayarlayın.
- Pürüzsüz → → Süreci tıklayın.
- → Eklentiler → Segmentasyon → İstatistiki Bölge Merging tıklayın. Göster Ortalamalar tıklayın → Q100 için parametreleri ayarlayın.
- Istatistiki bölge tamamen işlenmesini onaylayın yeni bir resim çerçevesi otomatik sunumu ile gösterilen ve etiket "SRM Q = 100" göründüğünden emin olun Birleştirme. Bu resim dosyası ile aşağıdaki adımları devam edin. → Görüntü → Tip → 8-bitlik tıklayın.
- → Eşiği ayarlayın → Görüntü → tıklayın. Düşük sinyal yoğunluğu TATS bileşenlerin özellikle kaçının dışlanmasına en tespit yeterince düşük eşik, ideal tüm TATS yapıları, seçin (başlangıç noktası olarak 40 eşiğini kullanın). Şekil 6 "Temsilcisi Sonuçlar" bölümünde, ayrıntılı veri çıkışı ve örnekler danışın(255 üst eşiği). Eşik parametrelerinin son seçim belgeleyin. Eşiğinin doğru seçimi nedeniyle arka plan gürültü yanlış pozitif sinyaller sadece belirli TATS membran yapılar üretmek değil gerektiğini unutmayın.
- Iskelet yapısının (gösterge) sürekliliğini aynı olmalıdır orta ve yüksek floresan sinyal seviyesinin özellikle ekstre iskelet veriler, karşı TATS görüntü detayları doğru superimposition onaylayın. Uygun bir eşik tespit edildikten sonra, analiz sırasında tüm görüntüleri bu aynı eşik uygulamak ve iskelet veri tutarlı yanlılık bu potansiyel kaynağını en aza indirmek için karşı orijinal sinyalin bindirme karşılaştırma tekrarlayın.
- Uygulama üzerine tıkla → "Eşik" başlığı altında, Şekil 5'te gösterildiği gibi, resim verileri ikili olur.
- → tıklayın Eklentileri → İskelet → iskeletini (2D / 3D). Tarafından Tif dosyası olarak (Şekil 5 de gösterildiği gibi) iskeletize 2D görüntü kaydetme→ Tif olarak kaydet → Dosya → tıklayarak. Eklentiler → tıklayarak nicel veri çıkışı için iskeletize görüntü dosyası analiz → İskeleti (2D / 3D) analiz. Hiçbiri: Erik döngüsü yöntemi seçin. Oluşan veri tablosunun otomatik nesil onaylayın.
- Txt dosyası olarak otomatik olarak oluşturulan veri tablosu kaydedin. İlgili nicel parametreler örneğin seçin, şube puan toplam sayısı veya Şekil 7'de örnek verileri tarafından gösterildiği gibi ortalama dal uzunluğu. Tamamlayıcı / destekleyici yazılım araçları Excel gibi ve gibi uygun olan başka veri analizi düşünün.
- Çıkart Arka → → Süreci tıklayın. 5 piksel Rolling top yarıçapı ayarlayın.
- Bireysel görüntüleri ve / veya görüntü partiler arasında analizini uyum için 4,5 altında açıklanan tüm gerekli adımları da dahil olmak üzere mümkün otomatik görüntü işleme rutinleri kullanmak için düşünün. Fiji için programlanmış bir görüntü işleme Makro (gerektiği gibi programlama ayarlayın) içeren örnek Ek Kod dosyası kullan.
- In Analiz"Directionality" eklentisi Fiji tarafından iskeletize görüntü verilerinden bütün seçin veya TATS ağ bileşenleri dividual yönelimleri. İlgili gerdirilmis A-tübül veya enine odaklı T-tübül bileşenleri 0 ° veya 90 ° bidonları tarafından temsil edildiği bir yönlülük histogram oluşturun. Görüntü yönü doğru başvurusu, ve Şekil 8 ve Örnek Sonuçlar gösterildiği gibi VM hücrenin ana (boyuna) 0 ° ekseni ile yakın karşılık görüntünün x-ekseni için önemli.
- Üzerine → → Direksiyonalite → Yöntem belirtmek Analiz tıklayın: Fourier bileşenleri, Nbins 180, Histogram -45 → başlamak Ekran masaya tıklayın.
- Txt dosyası olarak ilişkili histogram verileri de dahil yeni oluşturulan ve görüntülenen sonuçlar tabloyu kaydedin. Excel gibi ve gerektiği gibi ücretsiz yazılım araçları tarafından txt dosya verilerinin daha fazla analiz düşünün.
- Alt sınıflarını oluşturmak içinAynı durumda (ve potansiyel olarak diğer koşullar) altında tedavi edilen tüm hücreler için gruplandırılmış veri setleri örneğin gibi veri, analiz uygun olarak ilgili tüm görüntüler için 4,1-4,7 adımları tekrarlayın. Ortalama değerleri elde etmek için bir kombine Excel dosyası içine tüm görüntüleri iskeletize veri parametrelerini içe aktarın. Ayrıca, hesaplamak ve ortalama yöneltme histogram oluşturmak için kombine Excel dosyasına aynı gruplandırılmış veri kümesi tüm yönlülük histogram verilerini içe. Gerektiği gibi bireysel ya da arasında farklı tedavi grupları için ilgi ek TATS ağ parametrelerini analiz etmek gruplandırılmış veri setlerinin daha işleme düşünün.
Sonuçlar
Buna ek olarak, membran TATS ağ analizi için hücre içi Ca2 + görüntüleme, yama klempi elektrofizyoloji veya farmakolojik doz tepkisi çalışmaları gibi yaygın olarak kullanılan hücre biyolojisi teknikleri bir dizi kulakçıklar veya karıncık yüksek kaliteli birincil hücre izolasyonu sıkı sıkıya bağlıdır ya da seçmek Kalp dokusunun parçaları olgun yapısal, farklılaşmış ve fizyolojik bozulmamış kalp miyositlerin karakterizasyonu etkinleştirmek için. Bu nedenle, bölüm 1 de tarif AM ve VM hücreler için izolasyon ve kalite değerlendirme sağlam zarı ve hücre bütünlüğü eleştirel bağlıdır burada açıklanan TATS ağ analizi, dahil olmak üzere birçok farklı sorular için sonuçta yararlıdır.
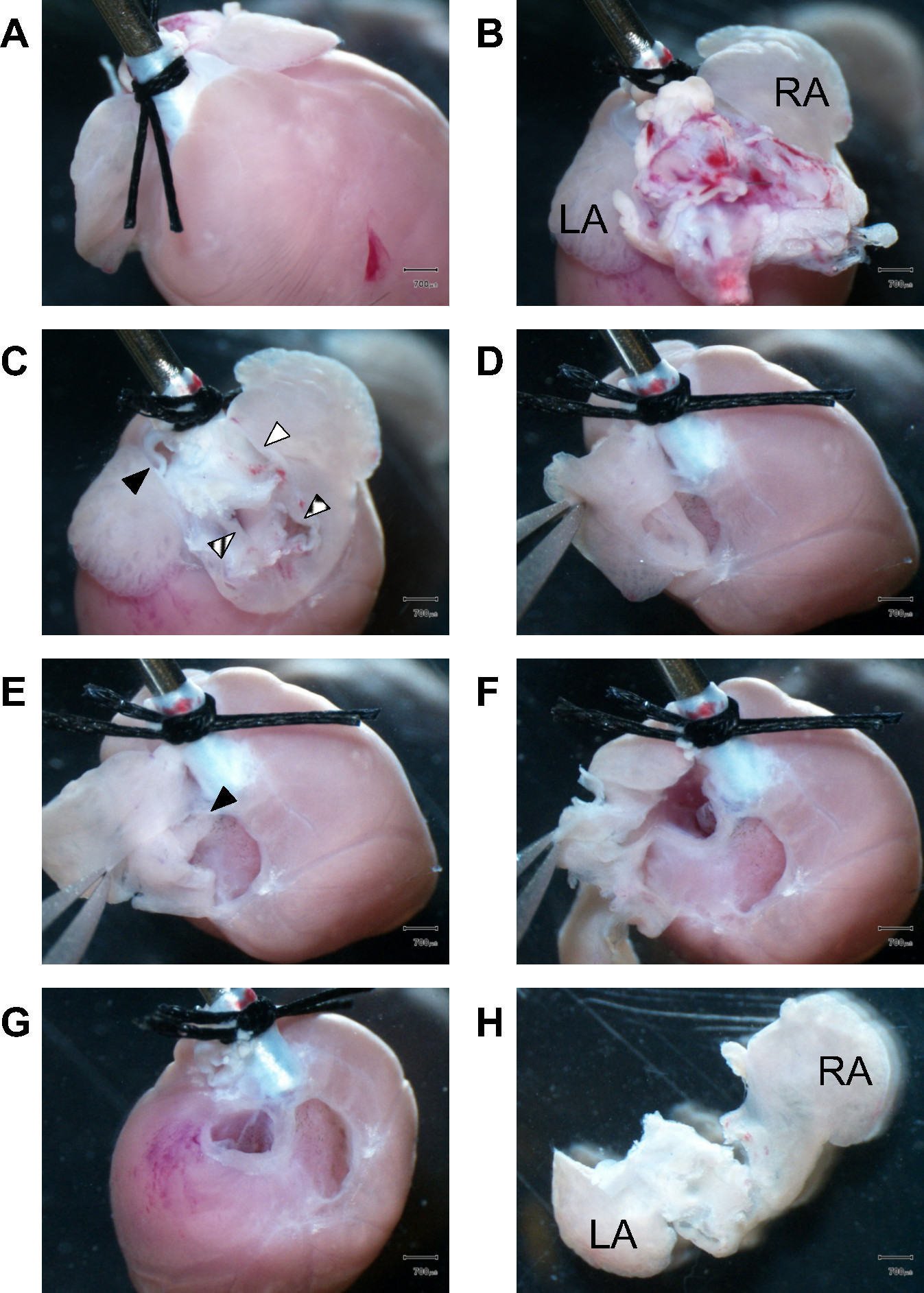
Şekil 1 kardiyak doku diseksiyonu fare kalbinde atriyal odaları ile başlayan ile devam etmek nasıl bir görüntü adım adım kılavuzu sağlar. Daha sonra, ventriküler bölmelerin ve septum hazırlanırlar ve birgerektiği gibi disseke d. Hassas seçimi ve doğru doku parçalarının hazırlanması güvenilir yeterli hücre saflıkta PM ve VM izolasyonların oluşturulması önemlidir. Kollajenaz sindirimi takiben de atriyal ve ventriküler dokunun arasındaki diseksiyon doğru hat belirlemek için göreceli olarak zor olabilir, ancak PM ve VM hücrelerin kontrolsüz hücre süspansiyonu karıştırıldıktan sonra, bu karma hücre grubunun tersine imkansızdır. Bu nedenle, anatomik oryantasyon, 3D doku görselleştirme, sindirilmiş doku işleme ile yeterli deneyim, özel doku parçaları ve diseksiyon hatları doğru belirlenmesi tüm hücre izolasyon başarısına katkıda bulunacaktır.

Atrial dokuda Şekil 1.Dissection. (A) kalbin ön kısmını bakan, iki cerrahi sutures bir 21 G çelik kanül güdük sonuna proksimal aort düzeltmek. diseksiyonu sırasında atriyal odacık görünümü engellemekten akciğer dokusunun kalan kalp temeli gösterileri doğru (B) Görünüm. LA, atrium sol; RA, sağ atrium. (C) Kalan lungtissue ve büyük damarların atriumlarda erişmek için çıkarıldı. Siyah bir üçgen, pulmoner arter Dolgulu; çizgili üçgen, pulmoner venler; beyaz üçgen, üst vena kava doldurdu; Kutulu üçgen, alt vena kava. (D) forseps atriyal apendiks tutarken Birincisi, sağ atriyal duvar disseke edilir. (E) sağ atriyal odacık boşluğuna gör. Siyah üçgen sağlam İnteratriyal septum işaretler. (F) diseksiyonu, sol atriyal boşluğa girmek için devam edilir. (G) sol ve sağ kulakçıklar tam diseksiyonu takiben, kalp kapakları görünür hale gelir. Lifli valf aparatı, kesilerek çıkarıldı ve subseq için uzaklaştırılmıştıruently sadece ventriküler kas dokusundan. izole sol ve sağ kulakçık (H) Posterior görünümü. LA, atrium sol; RA, sağ atrium.Scale barlar: 700 mikron.
Hücre izolasyonların kalitesini belirlemek için, Şekil 2 AM'ler ve sanal makineler için hem verim ve tipik çubuk veya çizgili bozulmamış miyositler şekilli tuğla canlılığı değerlendirilmesi sırasında tipik bir hücre örnekleri sağlar. Minik kolaylıkla bölümünde tarif edildiği gibi 1.11 tripan mavi eksklüzyonu dahil olmak üzere tespit edilebilir, anormal aşırı kavisli morfolojileri, veya anormal küresel şekillere sahip olan hücrelerdir görsel olarak göze çarpmayan olarak ciddi şekilde hasar görmemiş hücreler hasarlı. Mavi dışı Trypan maruz kaldığında bozulmamış miyositler parlak ve homojen striyate kalırken, hasarlı hücreler genellikle birden fazla zar kabarcıklannı göstermek ve / veya hızlı tripan mavi hücre belirten membran hasar birikir. Bununla birlikte, kendi başına, tripan mavi ile unnecessari hücrelere zarar verebilirly uzun inkübasyon ve acil hücre kalite değerlendirme nedenle zorunludur. Myofilament kontraktürün veya brüt yüzey hasarı ödün hücre bütünlüğü gibi hücre hasarının daha bariz biçimlerinin örnekleri Şekil 4C ve 4D gösterilmiştir. , bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.
{kind=link}

Şekil 2. Trypan mavi dışlama hücre boyama. İzole (A) AM ve (B) VM hücreleri tripan mavi ile süspansiyon içinde karıştırılır ve 40Xmagnification gösterilen ters ışık mikroskobu ile görüntülenmiştir. Leaka indicatesmembrane tripan mavisi alarak thatabnormal küresel (A) hücreleri ve (B) Notge ve yapısal hasar. Gösterildiği gibi aksine, sağlam membranlar ile centralAM ve VM hücreler mavilerin hariç. Ayrıca, bu bozulmamış AM unutmayın ve VM hücreleri, hücre hacmi boyunca sarkomer damarlar, nomembrane kabarcıklanm ve her iki yan taraflarında keskin kenarları ve her iki arakatgılı diskleri gösteriyor. Ölçek çubukları: 20 mikron.
10, 6 5 x 10 5 ikinci VM'lerin başarılı izolasyonu, hücre verimi izlenerek bir fare kalp sindirimden beklenebilir. AMS verimi, 3 x 10 3 4 10 3 x çubuk-şekilli, tripan mavisi hariç hücre için önemli oranda daha düşüktür. VM aksine, izolasyonları bazen hatta deneyimli ellerde başarısız AM. Adım 1.11 nasıl süspansiyon izole sağlıklı hücrelerin verimini tahmin etmek için prosedürleri özetlemektedir. Bir birey veya VM hücre izolatları için, Şekil 3'te gösterildiği gibi, ek olarak, adım 1.13 yoluyla ortalama hücre boyutlarını belirlemek veya VM hücre popülasyonlarının karşı yan yana (gerektiği gibi) AM karşılaştırmak. , bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.
{kind=link}

Kardiyak miyositlerin 3.Bright alan morfometrik analizi Şekil. VM kontur aşama 1.13 altında açıklanan görüntü analizi araçları tarafından tespit edildi. Işareti görsel olarak ve analizi için dış yüzeyi membran tarafından tanımlanan hücre dışı sınır tanımlamak için poligon seçim aracını kullanın. Reklam ROI yöneticisi ilgi (ROI) Seçili bölge 1D mesafe ölçümleri ile takip. Genişlik oranını: VM boyutları vs AM karşılaştırmalı çalışmalar için, hücre uzunluğu, genişliği ve alanı belge ve uzunluğunu hesaplamak için yararlıdır.
{kind=link}

Sağlam atriyal ve ventriküler myo Şekil 4.Live membran boyanmasılenfositler. ışık ve yaşam konfokal görüntüleri bulaşan karşı gelen di-8-ANEPPS bozulmamış (A) lekeli AM ve (B) VM hücreleri. Buna karşılık, 1.2 um'lik bir sarkomer uzunluğu ile kısmen daralarak potansiyel olarak zarar PM (C) 'de gösterilmiştir. Sözleşmeli miyositler genellikle anormal kısaltılmış göstermek ve çarpık TATS yapıları, bu nedenle daha fazla analiz dışında. Hücre membran kusurları için bir diğer önemli gösterge bir VM (D) 'de gösterildiği gibi zar kabarcıklannı (kırmızı üçgenler) vardır. Zar kabarcıkları bleblerini ile hasarlı yüzey membran yapılar ve hücreler ayrıca TATS analize dahil edilmelidir temsil eder. Ayrıca, VM görünüşte (yıldız ile işaretlenmiştir) sol alt bölümünün bir bütün parçası eksik bütün hasarı göstermektedir. Özetle, iletilen ışık ve konfokal görüntüleri karşılaştırarak, hücreler morfolojisi ve yüzey intakt belgelenmiş ve floresan sinyal bilgileri ile birleştirilir. 'N' işaretleri nuclei TATS membran leke analizde ihmal. Sarı çubuklar yukarıda sunulan aynı konfokal görüntü ROI'leri büyütülmüş göstermektedir. Ölçek çubukları:. 10 mikron bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.
{kind=link}
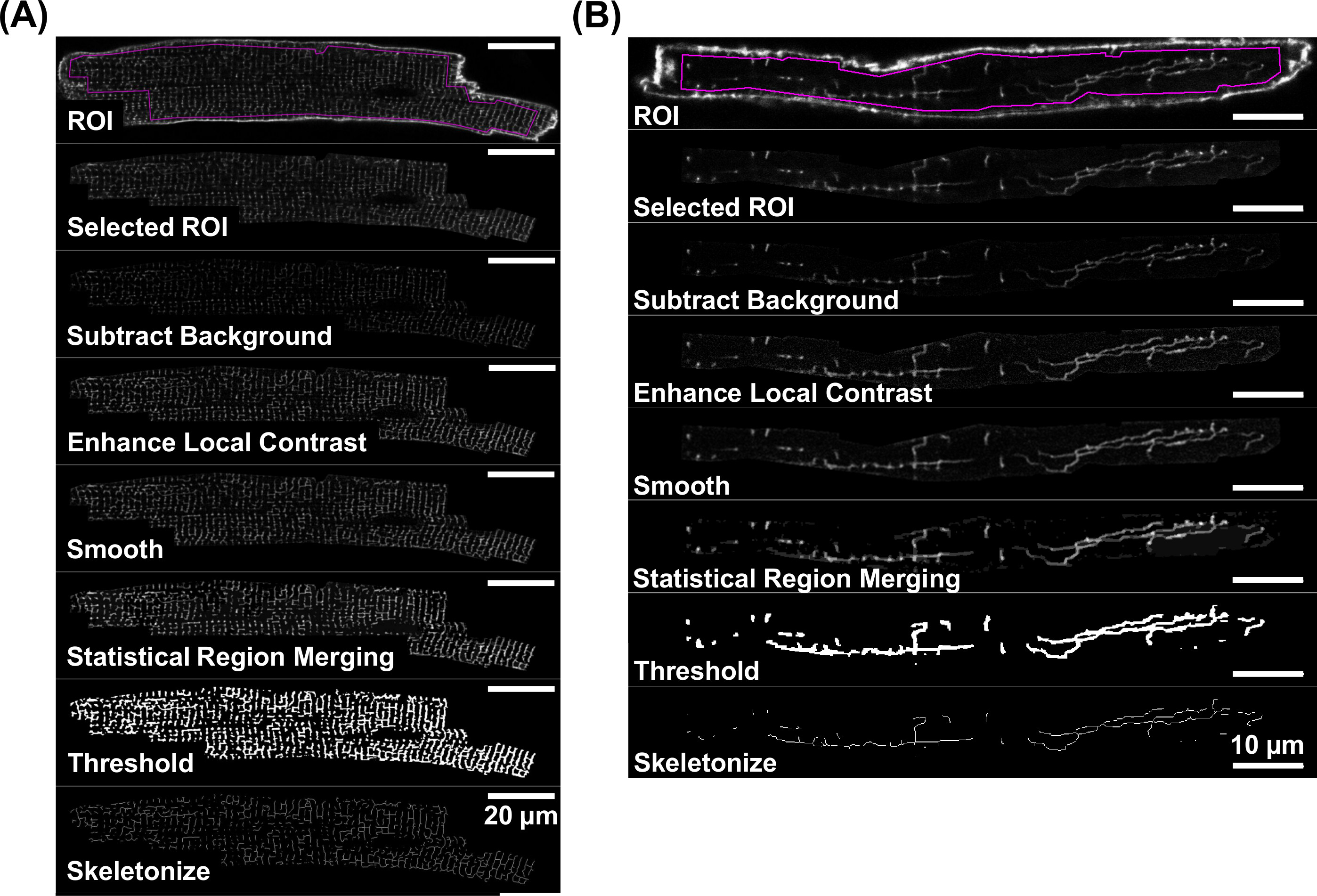
Şekil 4A ve 4B'de gösterildiği gibi, yeterli bir sinyal-gürültü oranına sahip TATS membran Konfokal görüntüler ayrıca nicel analiz için kabul edilir. TATS membran analizi flüoresan sinyal çizgisel maddeden türetilen iskeletize verilerine dayanmaktadır. 5 4,3-4,5 aşamalar ile detaylı olarak tarif edilmektedir, tek tek görüntü işleme aşamalarının iş akışı diyagramını göstermektedir. Bu adımlar her izole VM (Şekil 5A) için gösterildiği gibi doğrusal TATS membran ağları temsil iskeletize görüntüler üretir ve kurgu (hücreleri AMGüre 5B).

TATS ağın skeletonize bir resme neden Şekil 5.Workflow floresan TATS görüntülerin skeletonize için. Görüntü işleme adımları tek tek adım adım görüntü örnekler ile hem bir di-8-ANEPPS için temsil edilmiştir ve VM (A) ve AM lekeli (B). Bireysel resim işleme adım için, bölüm 4 bakın. Ölçek barlarda farklılıkları Not:. 20um (A) ve 10 um (B) , bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.
{kind=link}
Bölümünde 4,5 açıklandığı gibi görüntü işleme sırasında kritik bir adım, veri Binarization için uygun eşiğini belirler.6. Çıkan ikili görüntü sadece TATS ağdan gerçek membran sinyalleri değil arka sinyal gürültü hatalı eşikleme ile elde edilen sahte yapıları içermelidir. Ancak, bu eşik gerçek TATS bileşenler yanlışlıkla görüntü analizi sırasında kaybolmaz şekilde hepsi doğru TATS yapıları tespit için yeterince düşük olması önemlidir. 6 nasıl veri Binarization sırasında eşik seçmek için süreci göstermektedir Şekil. 40 eşik Şekil 6B'de gösterildiği ve 6C, sarı üçgenler ile belirtildiği gibi soluk eksenel zar yapılarına (ATS) algılamıyor Şekil 6A'da gösterildiği gibi, 60, daha yüksek bir eşik örneğin seçme tek tek tüm gerçek TATS yapıları tespit etmek için uygun göründüğü gibi iken . Tarafından gösterildiği gibi Şekil 6D'de gösterildiği gibi tersine, 20 daha düşük bir eşik değeri seçme, örneğin arka plan gürültü olarak yanlış pozitif TATS yapıların hatalı sezilmesinisarı üçgenler.

Şekil 6.How TATS görüntü veri skeletonizing sırasında sinyal eşiğini belirlemek için. Örnekler her (adım 4.5 tarif) veri Binarization sırasında farklı eşikleri için oluşturulan TATS iskeletler göstermektedir. Üst görüntüler: Fiji kullanarak eşik ayarlamaları gösterir. Alt görüntüler: gelen giriş floresan görüntü ile iskeletize görüntüleri ve büyütülmüş bölgelerin bindirme gösterildiği gibi. Yüksek eşik, örneğin, uygulanan 60 (A) büyütülmüş kısmında sarı üçgenler ile belirtildiği gibi, tüm gerçek TATS yapılarını tespit etmek için görünüşe uygun değildir. (B) ve (C) uygulanan 40 bir eşik düşük bir eşik örneğin, oysa arka plan gürültüsünü algılamıyor doğru tüm TATS yapıları algılar ve., 20 (D) yanlışlıkla mevcut olmayan zar yapılarının yanlış pozitif sinyaller üretir ve böylece TATS yapılar olarak arka plan gürültü tanımlar ve. Yanlış pozitif sinyaller büyütülmüş içerlek sarı üçgenler ile gösterilir. , bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.
{kind=link}
"İskelet (2D / 3D) Analiz" eklentisi iskeletize TATS yapıların detaylı analizini destekler. #branches, #junctions, # Son nokta voksellerden, ortalama dal uzunluğu, #triple puan, #quadruple puan ve maksimum şube uzunluğu: Bir kez eklenti aşağıdaki iskelet parametreleri ile bir veri tablosu oluşturur, idam. Tüm olası çıkış parametreleri ayrıntılı bilgi için lütfen bu adrese gidinhttp: //fiji.sc/wiki/index.php/AnalyzeSkeleton ve ilgili makaleleri 29-31. Tipik bir çıkış verileri tablosu, Şekil 7A'da gösterilmektedir.
parametreleri, başına toplam iskelet uzunluğu veya alan başına birleşme sayısını elde etmek için kullanılabilir. Ortalama dal uzunluğu ile çarpımı dalların sayısından örnek hesaplaması 2B'de sürekli iskeletin toplam uzunluğu verir:YG başına toplam iskelet uzunluğu:
Σ (#branches x ortalama dal uzunluğu) = 5155 PX = 515.5 um
Iskeletin toplam uzunluğu, görüntü bölgesine normalize olabilir. Aşağıda gösterildiği gibi, Şekil 7 0.64μm normalleştirilmiş iskelet uzunluğu gösterilen örnek için / um 2 ve her kavşak toplamı hesaplanır:
Normalize iskelet uzunluğu:
515.5 mm / 803,6 mikron 2 = 0.64 mm / mikron 2
Kavşaklarından normalize sayısı:
155 kavşak / 803,6 mikron 2 = 0.19 kavşaklar / mikron 2

Iskeletize görüntüleri 7.Automated veri çıkışı Şekil. (A) (B) gösterilen iskeletize görüntüden 'İskeleti analiz (2D / 3D)' eklentisi tarafından üretilen tipik bir veri elektronik tablo. Olası çıkış parametrelerinin ayrıntılı bir açıklama başvurun için http://fiji.sc/wiki/index.php/AnalyzeSkeleton . , bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.
{kind=link}
Şekil 5 4.5 altında tarif ve tasvir görüntü işleme adımları Ek Kod Dosyası olarak sağlanan bir Fiji makro kullanarak otomatik hale getirilebilir ong. Komutları reiterative adımlarda görüntü işleme tekrarlar tanımlar. Makro adımları 4.3 ve 4.4 ile üretilen görüntü giriş yığınlarını tamamlamak için uygulanabilir. Makro makro komutlarını kullanarak otomatik analiz için her bir bağımsız bir tedavi grubu için ayrı ayn giriş görüntü yığınlarını hazırlanmasıyla, örneğin, tam veri kümesi gruplarının analizi için avantajlı olabilir.
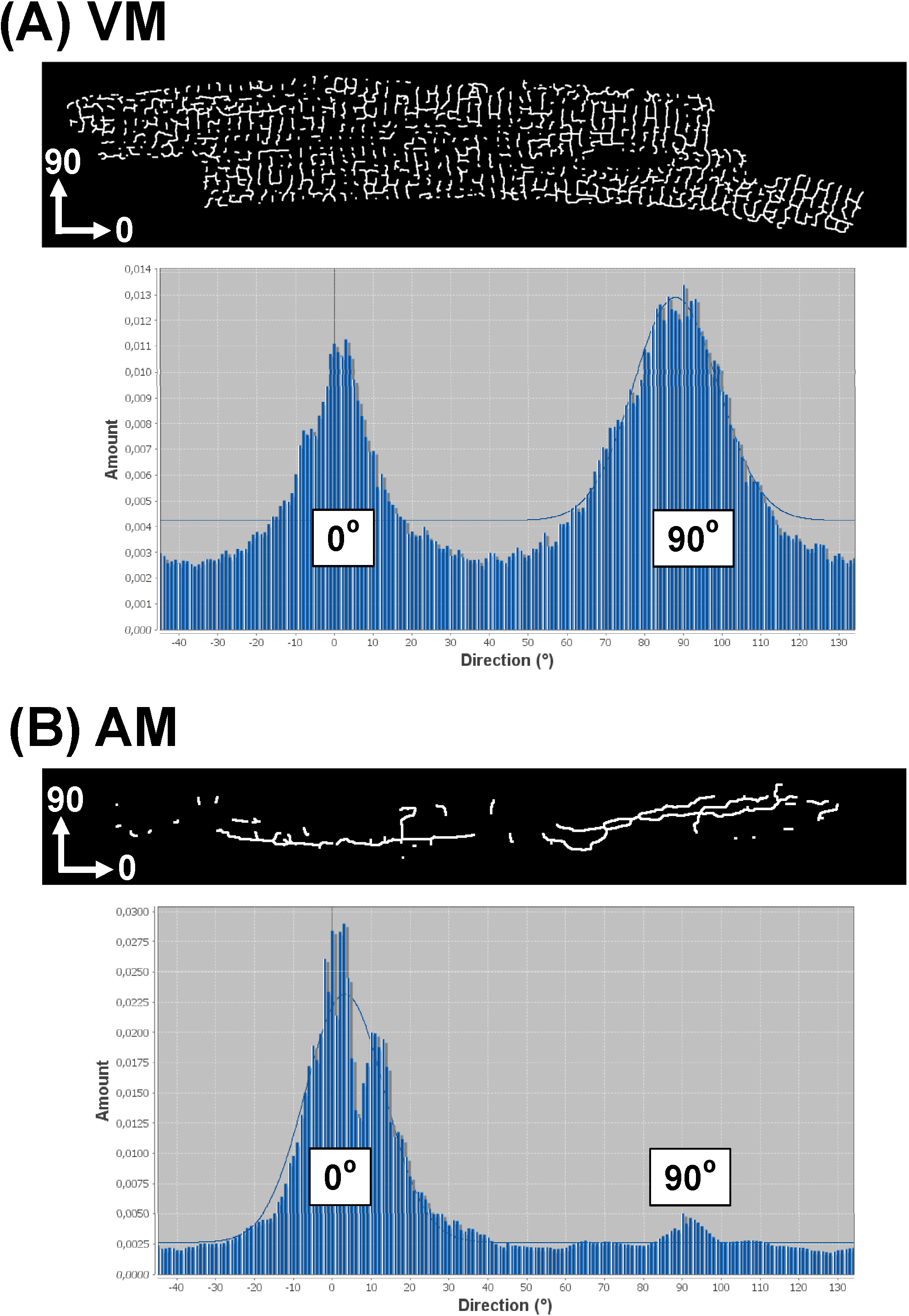
Özetlenen yazılım stratejisi daha tüm bileşenler için TATS ağ yönlendirme analiz sağlar. Bunun için, histogram verileri tüm tats bileşen yönelimleri yönelimi dağılımını gösteren üretir "Direksiyonalite" eklentisi (http://fiji.sc/Directionality) 29,31 kullanın. Giriş görüntüsünün x ekseni belirli bir PM veya VM hücrenin ana eksenine karşılık geliyorsa, çapraz parçalar 90 ve ile temsil edilecektir ise eksenel (boyuna) TATS parçalar, 0 ° bin ile temsil edilecektir# 176; bin. 8 örnek yönlülük bir AM (8B) hücrenin karşı bir VM (8A) için iskeletize TATS görüntüleri histogramlar göstermektedir. Tipik bir VM yöneltme histogram 0 ° ve 90 ° bir çift pik dağılımını gösterirken, AM histogram 0 ° tek bir baskın zirveye gösterir. Bu örnekler AM'ler yılında TATS bileşenler ağırlıklı olarak A-tübüllerinin oluşabilir oysa VM'lerin bireysel TATS bileşenleri neredeyse eşit, T-tübülleri ve A-tübüller arasında dağıtılır olduğunu daha önceki gözlemler ile uyum içindedir.

Tek tek hücrelerin TATS ağlardan 8.Representative yönlülük histogram Şekil. Direksiyonalite histogramlar AM karşı bireysel VM (A) iskeletize görüntüleri elde edildi (B ) TATS ağlar. T-tübül bileşenler 90 ° bin ile ifade edilen ise analiz görüntüsünün x ekseni gösterildiği gibi miyosit ana (boyuna) eksenine tekabül eden göz önüne alındığında, bir-tübül parçalar, 0 ° bin ile temsil edilmektedir. Gauss sadece arka plan grafik gibi önemli histogram peakasshown uyuyor. , bu rakamın büyük bir versiyonunu görmek için lütfen buraya tıklayınız.
{kind=link}
| Perfüzyon tamponu | mM |
| NaCl | 120.4 |
| KCl | 14.7 |
| KH 2 PO 4 | 0.6 |
| Na 2 HPO 4 | 0.6 |
| MgSO 4 | 1.2 |
| Hepes | 10 |
| NaHCO3 | 4.6 |
| Taurin | 30 |
| 2,3-Butandion-monoxime | 10 |
| Glikoz | 5.5 |
| pH 7.4 | |
| Sindirim tampon | mM (eğer belirtilmemişse) |
| NaCl | 120.4 |
| KCl | 14.7 |
| KH 2 PO 4 | 0.6 |
| Na 2 HPO 4 | 0.6 |
| MgSO 4 | 1.2 |
| Hepes | 10 |
| NaHCO3 | 4.6 |
| Taurin | 30 |
| 2,3-Butandion-monoxime | 10 |
| Glikoz | 5.5 |
| Kolajenaz, tip II | 600 U / ml |
| pH 7.4 | |
| Tampon | mM (eğer belirtilmemişse) |
| NaCl | 120.4 |
| KCl | 14.7 |
| KH 2 PO 4 | 0.6 |
| Na 2 HPO 4 | 0.6 |
| MgSO 4 | 1.2 |
| Hepes | 10 |
| NaHCO3 | 4.6 |
| Taurin | 30 |
| 2,3-Butandion-monoxime | 10 |
| Glikoz | 5.5 |
| CaCl2 | 0.0125 |
| buzağı serumu | % 10 |
| pH 7.4 | |
Tablo 1.Buffer solutiyonları. hücre izolasyonu ve görüntüleme için üç farklı fizyolojik tampon çözeltilerin içeriği özetlenmiştir.
Tartışmalar
Kardiyak miyositler izole ve yıllardır 32 için çalışılmasına rağmen, bir yeni yorum tutarlı, yüksek kaliteli miyositlerdeki hücre izolasyonları 27 zorlu kalması sonucuna varmıştır. Bu vis-à-vis ortak bir standart yaklaşımlar eksikliği, paylaşılan metadata ve şeffaf hücre kalite belgelerine primer kalp miyositlerin izolasyonu için nispeten karmaşık protokolleri yansıtır. Hücre izolasyonu protokolleri genellikle bireysel gruplar tarafından özelleştirilmiş, hücre izolatlarının, değişken sonuçlar üretmek bireysel modeli ayarlarıyla (örneğin, tür, yaş, eşlik eden kalp rahatsızlıkları) bağlıdır ve genellikle belirli deney koşulları için ayarlanır. Membran çalışmalar ve protokoller Burada yer alan sayısal tats, kalite değerlendirmesi ve belgelerin önemli bir seviyesinin kapsamında metabolik ve izolasyon protokole bağlı değişikliklere yatkındır Bireysel hücre zar yapılarına konfokal mikroskopi veya superresolution ilgilidir, her ikiAM veya VM. Önemlisi, hücre izolatların yüksek verimi sağlıklı sağlam miyositleri göstermese bile, araştırmacılar belgelemek ve eleştirel nedeniyle nedeniyle farklı türlerine özgü değişiklikler karşı izolasyon prosedürleri non-spesifik hasar karşı yüzey ve TATS membran bütünlüğünün morfolojik kriterlere karşı dikkatli her hücreyi yargılamak gerekir müdahalelerin gibi koşulları kontrol etmek için karşılaştırdık. Kalp hücre izolasyonu sırasında önemli bir değişkendir, belirli bir kolajenaz çok spesifik aktivitedir. Kolajenaz yeni bir çok seçmek için çeşitli örneklerin kolajenaz enzimatik aktivitesi, kardiyak miyosit, verim ve kaliteyi değerlendirmek ve üretici talimatlarına göre birbirlerine karşı test edilmelidir. İdeal olarak, kollajenaz, yeni bir sürü, önceki başarıyla kullanılmaktadır sürü benzer kollajenaz aktivitesi ile tanımlanır (olası enzim faaliyetlerinin uzun değerlendirilmesi için malzeme ve yöntemler tabloda "kollajenaz çok seçim aracı" bakınız). TBirlikte Aken, TATS membran görselleştirme nicel yaklaşımlar hücre izolasyon kalitesi ve tersi, izolasyon prosedürleri kritik gözden ve düzeltme tetiklemesi gerekir TATS mikroskopi ile belgelenen spesifik membran hasarına yol açan kardiyak hücre izolasyonları sıkı sıkıya bağlıdır. Hücre izolasyon kalitesi ve TATS membran görselleştirme ve miktar özünde bağlantılı olduğundan, bu makalede açıklanan protokolleri sürekli bir strateji olarak tüm önemli yönlerini kapsar.
Ek bir zorluk ve kardiyak çalışmalar, hücre hasarı ve / veya hücre kaybı ortak sorunu miyokard enfarktüsü 9 aşağıdaki metabolik ödün müdahaleler, örneğin nedeniyle meydana, henüz hücre izolasyonu sırasında fark hava embolisi aşağıdaki, potansiyel istenmeyen hasar örneğin karşı değerlendirilecektir gerekir. Hastalıklı kalpleri kardiyak miyositlerin izolasyonu ek, önemli hücre kaybına neden ve hücre verimleri azalmış olabilir. Bu nedenle, karşılaştırmalıhücre izolasyonu ve sayımı sürekli standart protokoller yoluyla uygulanır eğer kontrol ve hastalıklı kalpleri arasındaki izole edilmiş sağlam hücrelerinin toplam sayısının ison anlamlı olabilir. Sonuç olarak, mümkün olan en iyi miyosit hücre izolasyon kalitesini yansıttığı uygun bir kontrol grubu ile sağlanan hücre bütünlüğünü yargılamak için çok önemlidir. Daha da önemlisi, tek tek hücre kalitesi ve miyosit hastalıklı ve sağlıklı karşı canlı hücre mikroskopi yanlışlıkla anlamlı TATS membran ağların analiz etkileyebilir izolasyon prosedürü ile, hasarlı. Burada sunulan protokolleri bu nedenle hücre izolasyonu ve sağlam membranların canlı hücre mikroskopi sırasında fizyolojik membran bileşenlerinin bütünlüğü ve kararlılığı vurgulamak. Tüm iş akışı bu, membran bozulur membran tübüllere gibi izolasyon bağımlı membran eserler sergileyecek yana, başarmak ve hasarlı hücreleri hariç ise sağlam TATS zar bileşenlerini korumak için sürekli bir strateji olarak tasarlanmıştıre blebleri ve değişmiş TATS ağlar yanlışlıkla fazla nicel analiz kontrol koşulları altında ve uzlaşma. Tersi, aynı stratejiler TATS membran değişiklikleri ile gerçek hastalıklı hücreye karşı gerçek sağlıklı arasında anlamlı karşılaştırma kontrolleri sıkı sıkıya bağlıdır TATS membranlar, bozmaya potansiyeline sahip müdahale çalışmaları için çok önemlidir.
Ayrıca, AM hücrelerinin teknik olarak çok daha zorlu izolasyonu sağlamak için prosedürler ele. Ilerleme ve geliştirilmiş protokoller rağmen, o VM'lerin yüksek kalitede cep izolasyonların çoğaltmak önemsiz ve daha az güvenilir AM'ler için olmadığını vurgulamak önemlidir. Bu VM bir hücre hasarı hafif derecede bağlı olarak nispeten yüksek bir hücre, hücre süspansiyonu daha az belirgin olabilir, oysa bir hücre izolasyonu sırasında bile, küçük bir hata ya da varyasyonlar PM hücre izolasyonu başarısızlığı tamamlamak yol açabilir PM hücrelerin bütün olarak daha düşük verim nedeniyle numaraları AM karşılaştırıldığında. PM hücreleri c hale gelebilir yanaAşama 4.3 altında tarif edildiği üzere izole etme sonra urved çeşitli ROI analizi ile avantajlı olabilir. Hücre izolasyonu adımların ayrıntılı bir prosedür izlenerek, biz doğrudan integral membran boyama ve confocal veya VM ve AM'ler hem TATS ağlarının STED'in superresolution görüntüleme için bir protokol sağlar. Bu protokoller önce kurulmuş parametreleri aracılığıyla nicel analiz ve TATS membranların seçin bileşenlerinin farklılaşmasını sağlayacak. VM'lere karşılaştırıldığında, 3D organizasyon ve AM'ler atriyal TATS ağının işlevsel davranışları halen yeterince bilinmemektedir.
Canlı hücrelerde görüntü TATS zarlarına prosedürler (3,1-3,7 adımları) ticari confocal (Malzeme / Ekipman Tablo) ve ısmarlama STED'in floresan mikroskoplar 9 ile geliştirilmiştir. Floresan görüntü üretimi ve nicel TATS analiz için mikroskop ayarlarını optimize etmek için, aşağıdaki hususlar genel önem taşımaktadır:
- Amaç
Ampirik, TATS zar yapılarının küçük ayrıntıları çözmek hücrede derin birkaç mikrometre odaklanırken yüksek görüntü kalitesini sağlayan nesnel test etmek için. Bazı konfokal mikroskop burada kullanılan 63X 1.4 NA petrol amacı aksine, su ya da gliserol amaçlarına daha iyi yapabilir. 100X büyütme ile Amacı nanometrik çözünürlükte 34 için daha küçük bir görüş alanını kapalı ticaret, superresolution STED'in mikroskopi için kullanılır. - Uyarma ve Kazanç
Uyarım gücü ve detektör kazanç uygun ayarları mikroskop ışık yolu, lazer performansı ve örnek özelliklerine bağlıdır. İdeal olarak, lazer güç ve kazanç dedektörün tam kapsamlı istismar, henüz görüntü doygunluğu önlemek için ayarlanır. Ticari mikroskobu yazılım paketleri genellikle dinamik aralığının alt ve üst sınırı görselleştirmek arama tabloları sağlar. Bundan başka, boya beyazlatıcı mümkün olan en düşük l kullanılması en aza indirmek içinHala yeterli yapısal TATS membran ayrıntıları sağlar aser güç. Ayrıca, tahrik güç miyosit kontraktürleri ve ölüme yol kümülatif verilmedi hasarı önlemek için yeterince düşük olmalıdır. - Piksel Boyutu
Nyquist örnekleme ile uyumlu bir piksel boyutu, verilen ayarlarla elde yaklaşık yarısı çözünürlüğü kullanın. Konfokal görüntüleme için 100 nm x 100 nm'lik bir piksel boyutu, aynı zamanda, ağartma sınırlar olan uyumludur. Superresolution mikroskopi için önemli ölçüde daha küçük piksel boyutları STED'in mikroskopi 9 örneğin, 20 nm x 20 nm kullanılmaktadır. - Bekleme Süresi
Eşodaklı mikroskoplar bir ortalama fonksiyonu sağlamak. Genel olarak, sinyal ortalama örneğin birlikte ağartma önlemek için kısa piksel bekleme süresini kullanmak, hat-ortalama ≥ 8 sinyal-gürültü-oranı artırmak için. - Meta-veri ile Uygulamalı Mikroskopi Ayarları Belge
Nasıl ayarları kezTATS zar yapılarına görüntü bilgilerini belirli konfokal mikroskop güvenli duruma getirilmiş ve / veya ayarları (protokol meta-veri) belge edildi. Aynı amaca, uyarma gücü, kazanç, piksel boyutu, piksel bekleme süresi ve ortalama fonksiyonu ile hücrelerin bir (veya arasında) grup (lar) içindeki tüm görüntü kazanır. Eşit görüntüleme koşulları bir hücre (veya ile) grup (lar) içinde doğrudan karşılaştırılması ve nicelleştirilmesi için de izin verir. - Genel rehberlik ve esaslar konfokal mikroskopi uygulamaları hakkında daha fazla bilgi için, Biyolojik Konfokal Mikroskopi (Pawley JB, 3. Baskı, 2006, Springer Science + Business Media, LLC) El Kitabına bakınız.
Burada sunulan doğrudan analizi stratejileri, TATS membranlar ve hastalık ile ilgili değişiklikleri açıklayan önceki yayınlar, aksine Fourier transf dayalı nicel strateji 16,17 veya dolaylı bölgesel stratejileri gibi T-tübül yoğunluğu bölgesel agrega okumalarını kullanmışT-tübül bileşen düzenlilik 7 değerlendirmek amacıyla çizgili membran sinyallerin ormation analizi. Aksine, burada açıklanan nicel yaklaşımlar doğrudan bireyin TATS bileşenleriyle ilgili ve membran ağ özellikleri ve A-tübüller yüzdesi gibi özel bileşenler dahil olmak üzere ek parametreler bir dizi sağlamaktadır. Bundan başka, ağ yoğunluğu TATS YG alanı başına tüm ekstre iskeletin normalize uzunluğu belirlenebilir. Üç bireysel üçlü eklemlerin sayısı sürekli bağlı bileşenler tübül TATS zar ağın dallanma karmaşıklığı bir ölçüsü olarak kullanılabilir. Biz küçük TATS bileşenlerin herhangi bir analiz boyama prosedürleri bağlıdır unutmayın. Deneyimlerimize göre, 50 uM, di-8-ANEPPS çözeltisi 800 ul 50000 VM, 9 hücreleri ihtiva eden bir hücre peleti içerisindeki tam TATS ağları leke için yeterlidir. Ancak, hücre topağı kardiak miyositler daha az sayıda içeriyorsa, Güçlü bir floresan dedektör kullanılabilir ve eğer küçük membran bilgilerini ve niceliksel değişiklikler ilgilendiren ziyade genel TATS ağ dağıtım konfokal görüntüleme, düşük miktarda boya konsantrasyonları ampirik test göre de yararlanılabilmektedir. Son olarak, tarif edilen analiz için yazılmış bir yazılım makrosu farklı tedavi grupları (örneğin ilaçlar), hücre tipleri arasındaki karşılaştırma için özellikle yararlı olan daha büyük bir veri kümelerinin analizi kolaylaştırmak için görüntü işleme adımları otomatik olarak kullanılabilir (örneğin, VM karşı AM ) ve patofizyolojik müdahaleler (örneğin, miyokard enfarktüsü karşı plasebo).
TATS ağlarının görüntü analizi için, ilke adımların takip sırası uygulanır: 1) yuvarlanan bir top arka-çıkarma (4.5.1) arka plan yoğunluğu mekansal varyasyonları kaldırmak için; 2) Yerel kontrast geliştirme (4.5.2).; 3) görüntü yumuşatma (4.5.3); 4) İstatistiki bölge birleştirme (4.5.4); 5) tanımlayangörüntü Binarization eşik (4.5.6); iskelet verilerin ve 6) hesaplama (4.5.8). Floresan TATS görüntülerin skeletonize esnasında kritik bir safha, Şekil 6'da gösterilen görüntü çiftleme olan. Bağlantılı eşik aşamaları sonunda gerçek zar yapıları arka plan gürültü hata tarafından tespit edilen potansiyel yanlış yapıların karşı yatan TATS bileşenlerini temsil etmek için saptandığı tanımlar. Ikili görüntü analizi için uygun eşik tanımlanması konfokal mikroskopi ve superresolution yaklaşımlar için yeterince yüksek bir sinyal-gürültü oranı (SNR) oranına bağlı olarak, her gerçek tats zar yapıları ile uyumlu olmalıdır. Bu nedenle, yeterli bir görüntü kalitesi birinci ve belirtildiği gibi sonradan parlak alan görüntülerinin belgeleri dahil bireysel hücre kalitesinin kritik kararı ile kombine kurulmalıdır. Alternatif seçenekler belirli bir mikroskop veri çıkışı ve / veya PHYS için görüntü segmentasyonu protokolü adaptemalarla sorular görüntü dekonvulasyon ve İmageJ eklentileri olarak kullanılabilir "Otsu" veya "Iso-veri" gibi diğer eşikleme işlemleri içermektedir. Ne olursa olsun son segmentasyon prosedürünün, biz görüntü bindirmesi ile ekstre edilmiş ve ham veriler arasındaki karşılaştırmayı zorunlu kalite kontrol adım düşünün. Özetle, bireysel izole miyositler, hücre içi TATS membranların yeterli boyanması, floresan görüntüleme için parametre optimizasyonu ve ekstre iskelet veri bindirme kontrolü morfolojik ve membran bütünlüğü tüm floresan TATS görüntüleri ve kantitatif sonuçları kalitesine katkıda bulunacaktır.
Fare daha büyük türler hücre izolasyonu için kullanılır ise, hali hazırda protokoller uygun bir şekilde adapte edilebilir. Bir sonraki daha büyük türleri için, sıçan kalpleri kör bir 14 G kanül (dış çapı 2,1 mm) kanüllenmiş ve / dak, 8 ml perfüze. Anlamlı eski veya hastalıklı kalpleri daha büyük kanül boyutları gerektirebilir. Geninderal, kardiyak perfüzyon hazne ve aort arasında veya bir peristaltik pompa kullanılarak sabit akış ile 1 m yüksekliğinde bir su sütunu kullanılarak, örneğin sabit basınçlı yoluyla gerçekleştirilebilir. Kollajenaz sindirim sonunda sabit akış protokolleri tarafından bir dereceye kadar kontrol edilecek sızıntı damar yataklar aşırı perfüzyon oranları neden koroner direnç damarları bozacak beri fareler ve sıçanlar gibi küçük kemirgen kalpleri hücre izolasyonu için sabit akış avantajlı olabilir. Akış hızı ve doğru kanülasyon izlenmesi değiştirilmiş damar direnci davranışlı müdahale model yanı sıra hücre izolasyon prosedürlerinin eğitimi için yararlı olan bir öncelik vardır Buna zıt olarak, sabit basınç perfüzyon avantajlıdır.
Yukarıda belirtildiği gibi, yeterli hücre kalite endojen membran sistemleri kantitatif çalışmalar için çok önemlidir. Ancak kalp perfüzyon ve kollajenaz sindirim sırasında çok sayıda faktör cr olabiliritically protokol optimizasyonu veya sorun 27 atış sırasında asla küçümsenmemelidir hücre izolasyonu, kalitesini etkileyebilir. Özellikle, belirli bir kolajenaz aktivitesi çok çalışmanın kalan süresi boyunca muhafaza edilmesi ilgi örneğin, kulakçıklar veya izolasyonu koşulları belirlemek için deneysel çalışmalarda iyi niyetli yürütülmeden önce ventriküllerin spesifik doku için belirlenmelidir. Ayrıca, perfüzyon kurulum su kalitesi, pH, sıcaklık, optimizasyon ve temizleme kirletici ve emboli yersiz bir hasar riskini en aza indirecek, ve potansiyel olarak ek faktörler hücre izolasyonu sırasında optimal homeostatik koşullarını oluşturmak için izlenecek var. BDM (2,3-bütandion-monoxime) miyozin ATPaz çapraz köprü reversibl bir inhibitörüdür yaygın hücre izolasyonların verimini artırır kalp kas gevşemesini sürdürmek için doku diseksiyonu ve sindirim sırasında kullanılır. Bununla birlikte, araştırmacılar t gerekirO BDM hedef dışı etkiler, örneğin, belirli koşullar altında 33, Na + / Ca2 + değişimi akımlarının inhibisyonuna yol spesifik olmayan fosfataz etkinlikler sergileyebilen unutmayın. Bazı deneyler için, zehirli ve nispeten pahalı kardiyoplejik solüsyon, ancak, mikromolar konsantrasyonlarda miyozin için yüksek bir afiniteye sahip bir inhibitörü olarak Blebbistatin ile BDM değiştirilmesi avantajlı olabilir ve diğer hedef dışı etkilere sahip olabilir. İstirahat sağlıklı kardiyomiyositlerde ileri analiz dışlanmamalıdır elektrik stimülasyon ve bu hücrelerin yokluğunda bir kasılmaları göstermemelidir. Diğer taraftan, fizyolojik hücre dışı Ca2 + konsantrasyonlarında, elektrikli uyarılmaya yanıt olarak kardiyak miyosit kasılma ve gevşeme üzerinde sağlıklı kontrol grubuna kıyasla kalp hastalığı, fonksiyonel hücre kalitesi ve / veya anormal davranışlarını değerlendirmek için ek bir önlem olarak, normal kasılma hareketleri kurmak için kullanılabilir hücreler.
, tek bir hücre izolasyonu ve burada açıklanan kantitatif görüntü analizi için protokoller başarıyla hücrelerin 21 yanı sıra mikrotübül ağı kantitatif analiz VM 9 TATS membran ağın konfokal mikroskopi ve superresolution uygulanan ve AM edilmiştir sabit kardiyak miyositler (veriler gösterilmemiştir). Böyle farklı gelişim aşamalarında TATS membranların karakterizasyonu veya TATS ağı irtibata membran ilişkili protein veya organel yapıların analizi gibi deneysel sorular çeşitli yollar açabilir protokollerin ve gelecekteki uygulamaları son derece, etki alanı özel sinyalizasyon yerelleştirilmiş uygularken için AM ve VM hücrelerinde fonksiyon.
Açıklamalar
The authors declare that no conflict of interest exists.
Teşekkürler
This work received support through Deutsche Forschungsgemeinschaft SFB 1002 (subprojects A05 and B05 to S.E.L.) and KFO 155 (subproject 4 to S.E.L.), a Halbach Foundation award to S.E.L. supporting E.W.; a grant from the German Cardiac Society to S.B.; and a DAAD exchange program supporting T.K. as visitor at the University of Maryland. The research leading to these results has received funding from the European Community’s Seventh Framework Program FP7/2007-2013 under grant agreement No. HEALTH-F2-2009-241526, EUTrigTreat (to S.E.L.). S.E.L. is a principal investigator of the German Center of Cardiovascular Research (DZHK).
Malzemeler
| Name | Company | Catalog Number | Comments |
| Chemicals and Enzymes | |||
| 2,3-Butanedione monoxime | Sigma-Aldrich, Munich, Germany | B0753 | |
| Bovine calf serum | Thermo Scientific, Schwerte, Germany | SH30073 | Triple 0.1 µm sterile filtered. |
| CaCl2 | Sigma-Aldrich, Munich, Germany | 21115 | Diluted 1:10 in MQ water to obtain 100 mM CaCl2 stock concentration. |
| Collagenase type II | Worthington via Cell Systems, Troisdorf, Germany | on request | Enzymatic activity depends on individual collagenase batches. Collagenase II and other enzyme activities (Caseinase, Clostripain, Tryptic) can be assessed in the "collagenase lot selection tool". Determine cell yield and quality individually for each new lot of collagenase. |
| Glucose | Carl Roth, Karlsruhe, Germany | HN06.1 | |
| Heparin | Rotexmedica, Trittau, Germany | PZN-03862340 | Diluted in 0.9% NaCl and injected subcutaneuosly in abdominal skin. |
| HEPES | Carl Roth, Karlsruhe, Germany | 9105.4 | |
| Forene 100% (V/V) | Abbott, Libertyville, IL, USA | B506 | Active agent: isoflurane, 250 ml. Use approximately 2 Vol% in air/oxygen dispenser instrument. |
| KCl | Carl Roth, Karlsruhe, Germany | 6781.3 | |
| KH2PO4 | Carl Roth, Karlsruhe, Germany | 3904.2 | |
| Laminin (2 mg/ml) | BD Biosciences, Heidelberg, Germany | 354232 | Lamination is described under step 2.1. |
| MgCl2·6H2O | Carl Roth, Karlsruhe, Germany | 2189.2 | |
| MgSO4·7H2O | Carl Roth, Karlsruhe, Germany | 8283.2 | |
| Na2HPO4·2H2O | Carl Roth, Karlsruhe, Germany | 4984.2 | |
| NaHCO3 | Carl Roth, Karlsruhe, Germany | HN01.1 | |
| Taurin | Carl Roth, Karlsruhe, Germany | 4721.2 | |
| Dyes | |||
| Di-8-ANEPPS | Molecular Probes, Life Technologies, Darmstadt, Germany | D-3167 | Stock solution 2 mM in DMSO |
| Trypan blue | Sigma-Aldrich, Munich, Germany | T8154 | Trypan blue is gently mixed 1:1 via tip-cut 1 ml plastic pipette with cell suspension prior to cell counting in Neubauer cytometer. |
| Langendorff Perfusion Setup | |||
| Circulation thermostat | Lauda, Lauda-Königshofen, Germany | Please refer to Louch et al. (JMCC 2011). Heat up thermostat und buffers in perfusion tubing to 37 °C 15 min prior to use. | |
| Flexible silicone tubing Tygon for peristaltic pump | VWR, Darmstadt, Germany | 224-2252 | Tubing needs to be changed regularly. |
| Flexible silicone tubing Tygon for thermostat | VWR, Darmstadt, Germany | 228-4340 | |
| Heating coil surroundung perfusion tubing | Rettberg, Göttingen, Germany | custom-made | Heating coil and tubing needs to be cleaned thoroughly via MQ water after using. Do not use detergents. Glass components should be bathed regularly in 10 mM NaOH overnight. |
| Peristaltic pump | Ismatec, Wertheim, Germany | ISM830 | |
| Three way stop cock Discofix C Luer Lock 10 cm | Braun, Melsungen, Switzerland | 16500C | |
| Three way stop cock Discofix 3SC | Braun, Melsungen, Switzerland | 4095146 | |
| Instruments | |||
| 42 mm glass coverslips | Menzel Gläser via Thermo Scientific, Schwerte, Germany | on request | 0.13-0.16 mm thickness |
| Cannula 21 G | Becton, Dickinson and Company, Franklin Lake, NY, USA | 304432 | Cut to a length of ~5 mm, roughened with sandpaper. |
| Coverslips for Neubauer cytometer 24 x 24 mm | Menzel Gläser via Thermo Scientific, Schwerte, Germany | on request | 0.38-0.42 mm thickness |
| Graefe forceps, 0.5 mm tips, slight curve | Fine Science Tools, Heidelberg, Germany | 11151-10 | |
| LSM 710 NLO | Carl Zeiss, Jena, Germany | 63X 1.4 NA oil objective | |
| Neubauer improved cytometer | Labor Optik, Friedrichsdorf, Germany | 1100000 | Counting procedure: Wipe cytometer and coverslip provided with the counting chamber with 70 % ethanol. Press coverslip gently on the counting chamber so that the two glass surfaces are in contact and Newton's rings can be observed. Subsequently, 10-20 µl cell suspension can be applied to the edge of the coverslip to be sucked into the void by capillary action. Count the intact vs. defect myocytes using the squares of the cytometer grid which reflects 100 nl. Repeat counting procedure on the second grid provided on the cytometer. Calculate the density of cells in your original cell suspension by taking account of any dilutions and counting shortcuts. |
| POC-R2 Imaging Chamber | Pecon, Erbach, Germany | Cell suspension volume: 800 µl; desired plating density: ~1,000 AM and ~10,000 VM | |
| Spring scissors, 8 mm blades straight, blunt | Fine Science Tools, Heidelberg, Germany | 15025-10 | |
| Student dumont #7 forceps, inox | Fine Science Tools, Heidelberg, Germany | 91197-00 | |
| Student iris scissors, curved, 11.5 cm | Fine Science Tools, Heidelberg, Germany | 91461-11 | |
| Student iris scissors, straight, 11.5 cm | Fine Science Tools, Heidelberg, Germany | 91460-11 | |
| Student surcigal scissors, straight, sharp, 12 cm | Fine Science Tools, Heidelberg, Germany | 91402-12 | |
| Tissue forceps, 1 x 2 teeth, slim, 10 cm | Fine Science Tools, Heidelberg, Germany | 11023-10 | |
Referanslar
- Prosser, B. L., Ward, C. W., Lederer, W. J. Subcellular Ca2+ signaling in the heart the role of ryanodine receptor sensitivity. J Gen Physiol. 136 (2), 135-142 (2010).
- Wehrens, X. H., Lehnart, S. E., Marks, A. R. Intracellular calcium release and cardiac disease. Annu Rev Physiol. 67, 69-98 (2005).
- Cheng, H., Cannell, M. B., Lederer, W. J. Propagation of excitation-contraction coupling into ventricular myocytes. Pflugers Arch. 428 (3-4), 415-417 (1994).
- Williams, G. S., Chikando, A. C., Tuan, H. T., Sobie, E. A., Lederer, W. J., Jafri, M. S. Dynamics of calcium sparks and calcium leak in the heart. Biophys J. 101 (6), 1287-1296 (2011).
- Sperelakis, N., Rubio, R. orderly lattice of axial tubules which interconnect adjacent transverse tubules in guinea-pig ventricular myocardium. J Mol Cell Cardiol. 2 (3), 211-220 (1971).
- Soeller, C., Cannell, M. B. of the transverse tubular system in living cardiac rat myocytes by 2-photon microscopy and digital image-processing techniques. Circ Res. 84 (3), 266-275 (1999).
- Song, L. S., et al. et al. ryanodine receptors in the failing heart. Proc Natl Acad Sci USA. 103 (11), 4305-4310 (2006).
- Oort, R. J., et al. Disrupted junctional membrane complexes and hyperactive ryanodine receptors after acute junctophilin knockdown in mice. Circulation. 123 (9), 979-988 (2011).
- Wagner, E., et al. Stimulated emission depletion live-cell super-resolution imaging shows proliferative remodeling of T-tubule membrane structures after myocardial infarction. Circ Res. 111 (4), 402-414 (2012).
- Asghari, P., Schulson, M., Scriven, D. R., Martens, G., Moore, E. D. Axial tubules of rat ventricular myocytes form multiple junctions with the sarcoplasmic reticulum. Biophys J. 96 (11), 4651-4660 (2009).
- Lukyanenko, V., Ziman, A., Lukyanenko, A., Salnikov, V., Lederer, W. J. Functional groups of ryanodine receptors in rat ventricular cells. J Physiol. 583 (Pt 1), 251-269 (2007).
- Shacklock, P. S., Wier, W. G., Balke, C. W. Local Ca2+ transients (Ca2+ sparks) originate at transverse tubules in rat heart cells. J Physiol. 487 (Pt 3), 601-608 (1995).
- Reynolds, J. O., et al. Junctophilin-2 is necessary for T-tubule maturation during mouse heart development. Cardiovasc Res. 100 (1), 44-53 (2013).
- Di Maio, A., Karko, K., Snopko, R. M., Mejia-Alvarez, R., Franzini-Armstrong, C. T-tubule formation in cardiac myocytes two possible mechanisms. J Muscle Res Cell Motil. 28 (4-5), 231-241 (2007).
- He, J., et al. Reduction in density of transverse tubules and L-type Ca(2+) channels in canine tachycardia-induced heart failure. Cardiovasc Res. 49 (2), 298-307 (2001).
- Heinzel, F. R., et al. Remodeling of T-tubules and reduced synchrony of Ca2+ release in myocytes from chronically ischemic myocardium. Circ Res. 102 (3), 338-346 (2008).
- Lyon, A. R., et al. Loss of T-tubules and other changes to surface topography in ventricular myocytes from failing human and rat heart. Proc Natl Acad Sci USA. 106 (16), 6854-6859 (2009).
- Crossman, D. J., Ruygrok, P. N., Soeller, C., Cannell, M. B. Changes in the organization of excitation-contraction coupling structures in failing human heart. PLoS One. 6 (3), e17901(2011).
- Kemi, O. J., et al. The effect of exercise training on transverse tubules in normal, remodeled, and reverse remodeled hearts. J Cell Physiol. 226 (9), 2235-2243 (2011).
- Sachse, F. B., et al. Subcellular structures and function of myocytes impaired during heart failure are restored by cardiac resynchronization therapy. Circ Res. 110 (4), 588-597 (2012).
- Arakel, E. C., et al. Tuning the electrical properties of the heart by differential trafficking of KATP ion channel complexes. J Cell Sciences. 127 (Pt 9), 2106-2119 (2014).
- Richards, M. A., et al. Transverse tubules are a common feature in large mammalian atrial myocytes including human. Am J Physiol Heart Circ Physiol. 301 (5), H1996-H2005 (2011).
- Trafford, A. W., Clarke, J. D., Richards, M. A., Eisner, D. A., Dibb, K. M. Calcium signalling microdomains and the t-tubular system in atrial mycoytes potential roles in cardiac disease and arrhythmias. Cardiovasc Res. 98 (2), 192-203 (2013).
- Greiser, M., Schotten, U. Dynamic remodeling of intracellular Ca2+ signaling during atrial fibrillation. J Mol Cell Cardiol. 58, 134-142 (2013).
- Voigt, N., Zhou, X. B., Dobrev, D. Isolation of human atrial myocytes for simultaneous measurements of Ca2+ transients and membrane currents. J Vis Exp. (77), 10-3791 (2013).
- Kaestner, L., et al. Isolation and genetic manipulation of adult cardiac myocytes for confocal imaging. J Vis Exp. (31), (2009).
- Louch, W. E., Sheehan, K. A., Wolska, B. M. Methods in cardiomyocyte isolation, culture, and gene transfer. J Mol Cell Cardiol. 51 (3), 288-298 (2011).
- King, N. M., et al. Mouse intact cardiac myocyte mechanics cross-bridge and titin-based stress in unactivated cells. J Gen Physiol. 137 (1), 81-91 (2011).
- Schindelin, J., et al. Fiji, an open-source platform for biological-image analysis. Nat Methods. 9 (7), 676-682 (2012).
- Arganda-Carreras, I., Fernandez-Gonzalez, R., Munoz-Barrutia, A., Ortiz-De-Solorzano, C. 3D reconstruction of histological sections Application to mammary gland tissue. Microsc Res Tech. 73 (11), 1019-1029 (2010).
- Liu, Z. Q. Scale space approach to directional analysis of images. Appl Opt. 30 (11), 1369-1373 (1991).
- Powell, T., Twist, V. W. A rapid technique for the isolation and purification of adult cardiac muscle cells having respiratory control and a tolerance to calcium. Biochem Biophys Res Commun. 72 (1), 327-333 (1976).
- Watanabe, Y., et al. Inhibitory effect of 2,3-butanedione monoxime (BDM) on Na(+)/Ca(2+) exchange current in guinea-pig cardiac ventricular myocytes. Br J Pharmacol. 132 (6), 1317-1325 (2001).
- Kohl, T., Westphal, V., Hell, S. W., Lehnart, S. E. Superresolution microscopy in heart - cardiac nanoscopy. J Mol Cell Cardiol. 58, 13-21 (2013).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır