
Method Article
Analisi della membrana tubolare reti in Cardiac miociti da atri e ventricoli
* Questi autori hanno contribuito in egual misura
In questo articolo
Riepilogo
In miociti cardiaci, strutture a membrana tubolare formano reti intracellulari. Descriviamo protocolli per i) l'isolamento dei miociti da cuore del mouse, tra cui il controllo di qualità, ii) la colorazione delle cellule vive per microscopia state-of-the-art fluorescenza ottimizzato, e iii) analisi di immagine diretta di quantificare la complessità dei componenti e la plasticità della membrana intracellulare reti.
Abstract
In cardiac myocytes a complex network of membrane tubules - the transverse-axial tubule system (TATS) - controls deep intracellular signaling functions. While the outer surface membrane and associated TATS membrane components appear to be continuous, there are substantial differences in lipid and protein content. In ventricular myocytes (VMs), certain TATS components are highly abundant contributing to rectilinear tubule networks and regular branching 3D architectures. It is thought that peripheral TATS components propagate action potentials from the cell surface to thousands of remote intracellular sarcoendoplasmic reticulum (SER) membrane contact domains, thereby activating intracellular Ca2+ release units (CRUs). In contrast to VMs, the organization and functional role of TATS membranes in atrial myocytes (AMs) is significantly different and much less understood. Taken together, quantitative structural characterization of TATS membrane networks in healthy and diseased myocytes is an essential prerequisite towards better understanding of functional plasticity and pathophysiological reorganization. Here, we present a strategic combination of protocols for direct quantitative analysis of TATS membrane networks in living VMs and AMs. For this, we accompany primary cell isolations of mouse VMs and/or AMs with critical quality control steps and direct membrane staining protocols for fluorescence imaging of TATS membranes. Using an optimized workflow for confocal or superresolution TATS image processing, binarized and skeletonized data are generated for quantitative analysis of the TATS network and its components. Unlike previously published indirect regional aggregate image analysis strategies, our protocols enable direct characterization of specific components and derive complex physiological properties of TATS membrane networks in living myocytes with high throughput and open access software tools. In summary, the combined protocol strategy can be readily applied for quantitative TATS network studies during physiological myocyte adaptation or disease changes, comparison of different cardiac or skeletal muscle cell types, phenotyping of transgenic models, and pharmacological or therapeutic interventions.
Introduzione
Nelle cellule muscolari striate sane, strutture a membrana tubolari con orientamenti "trasversali" (tubuli T) perpendicolari all'asse delle cellule principali sono abbondanti. Di conseguenza, T-tubuli sono stati caratterizzati da continue estensioni della cellula muscolare principale "laterale" membrana superficiale (sarcolemma), che penetrano profondamente citosol verso il centro della cellula. Il ruolo fisiologico di tubuli T continue con la membrana superficie esterna è l'accoppiamento elettrico rapido di compartimenti intracellulari remoti formate da domini contatto SER organelli tutto il volume relativamente grande cardiaco cellule muscolari per accoppiamento vicinanza nanometrica di tensione attivati tipo L Ca 2 + canali (Cav1.2) verso l'interno corrente (I Ca) attivando recettori ryanodine adiacente (RyR2) SER Ca 2 + release. In miociti ventricolari (VM), i contatti di membrana non continui ("nodi") tra i domini giunzionale SER e T-tubules sono pensati per controllare migliaia di singoli nanodomini Ca 2 + intracellulare di rilascio in ogni cella 1.
Per ogni dominio contatto, le porzioni di membrana giustapposte ciascuno dei T-tubulo e l'(giunzionale) SER periferico sono di circa 15 nm vicini gli uni agli altri, quindi definiti come nanodomain. In tal modo, molto piccoli singoli sottospazi citoplasmatici vengono separati che consentono comportamenti vano cella-autonoma quasi. Quando un potenziale d'azione in arrivo attiva i canali Cav1.2 nel T-tubuli di macchine virtuali, una relativamente piccola Ca 2 + verso l'interno corrente aumenterà rapidamente il sottospazio Ca 2 + concentrazione [Ca 2 +] S nel nanodomain dimensioni attoliter 1. Accanto, la S aumento [2 + Ca] attiva Ca 2 + -gated recettori rianodinici (RyR2) in prossimità nanometri nella giunzione membrana SER giustapposti, e questo processo di accoppiamento si verifica in tutta coppia elettricamented nanodomini miociti. RyR2s si verificano come densi cluster multi-canale con una stechiometria stimato di 1 canale Cav1.2 per 5-10 canali RyR2 2. Dal momento che il SER-to-citosol [Ca 2 +] pendenza è molto ripida (rapporto 10 4: 1) e RyR2s Funzione come ad alta conduttanza canali Ca 2 + rilascio in ammassi funzionalmente accoppiati, RyR2 risultati attivazione di un grande quantitativo di Ca 2 + rilasciare corrente da T-tubulari accoppiati domini SER giunzionali crescenti sottospazio locale [Ca 2 +] S a 100 micron o superiore in 1-2 msec 3,4. Questo cardiaco comportamento amplificazione del segnale è indicato anche come Ca 2 + indotto da Ca 2 + rilascio (CICR). Nel loro insieme, T-tubuli sono strutture di membrana essenziali che attivano rapidamente Ca 2 + segnali di rilascio attraverso giunzionali contatti nanodomain SER e livello cellulare CICR durante l'eccitazione-contrazione (EC) di accoppiamento.
Oltre ai tubuli T, tubulo assiales (A-tubuli) con una significativamente diverso orientamento parallelo alle principali (longitudinale) Asse cellulari sono stati documentati mediante microscopia elettronica (EM), confocale e studi di microscopia a 2 fotoni. Ad esempio, un reticolo continuo a livello delle cellule di A-tubuli tra miofibrille interconnessi con T-tubuli vicino Z-linee sarcomero è stato dimostrato da traccianti extracellulari e EM imaging cavia fisso VM 5. Utilizzando extracellulare destrano-linked fluoresceina colorazione e vivere 2-Photon Imaging di ratto macchine virtuali, una reticolare complessa rete tubulo 3D è stato visualizzato costituito da ~ 60% di T-tubuli e ~ 40% A-tubuli 6. Questo studio non solo ha portato alla visualizzazione 3D di abbondanti A-tubuli, ma anche alla realizzazione che di sezionamento per la visualizzazione EM è intrinsecamente limitato per l'analisi di reti di membrana complessi e dinamici come il sistema tubulo trasverso-assiale (TATS). Di conseguenza, confocale imaging cellulare dal vivo di membrane tatuaggi direttamente macchiato di di-8-ANNEPS è stato sviluppato. Se il cellulare dal vivoReti Tats sono analizzati da Fourier trasformazione, l'aspetto regolare dei componenti T-tubulo nello spazio vicino Z-linee sarcomero si riflette lo spettro di potenza insieme da una regione di segnali striati 7. Questa strategia di analisi indiretta è stata utilizzata per rilevare le modifiche regionali a livello di cellulari in TATS componente regolarità in modelli di malattia 7. Ad esempio, shRNA mediata junctophilin-2 knock-down ha causato insufficienza cardiaca e la carenza di proteina specifica isoforma portato a T-tubulo riorganizzazione con nanodomain Ca 2 + rilascio disfunzioni 8. Abbiamo recentemente esteso l'analisi delle reti di membrana tatuaggi attraverso approcci quantitativi diretti e oltre, dal vivo al microscopio SuperResolution cella di componenti tatuaggi individuali in topo macchine virtuali utilizzando deplezione emissione stimolata (STED) nanoscopia 9. Risoluzione nanometrica consentito per l'analisi diretta dei componenti tatuaggi individuale più piccoli, che approssimano la distribuzione delle 50:50 trasversalirispetto orientamenti tubuli assiali, quantitativamente confermando ancora due differenzialmente orientati componenti tatuaggi individuali abbondanti nei cuori sani topo 9. Tali strategie saranno ulteriormente illustrate nella sezione del protocollo di seguito.
Mentre il ruolo fisiologico delle abbondanti componenti A-tubulari nel cuore adulto è rimasto enigmatico, studi EM hanno documentato strutture a membrana SER connessi con A-tubuli suggerendo endogena Ca 2 + nanodomini rilascio di cavia e nel ratto VM 5,10. Analisi confocale di Cav1.2 e RyR2 trovato un alto grado di colocalization agli incroci A-tubulo 10. Poiché ~ 20% dei spontanee Ca 2 + scintille nel ratto VM avuto origine relativamente lontano da striature Z-linea, dove in genere si verificano T-tubuli, un argomento è stato che nanodomini A-tubulo associati possono effettivamente esistere e funzionare come Ca 2 + siti di rilascio 11,12. È interessante notare che, la formazione di T-tubuli e la maturazione occagnacci solo dopo la nascita e paralleli la crescita delle cellule cardiache, per esempio, attraverso la germinazione di invaginazioni precursore sarcolemma a P5 e immaturo ramificati assemblee rete TATS a P10 nei topi 13. Sembra che Junctophilin-2 è particolarmente importante per la maturazione postnatale rete TATS dal shRNA knock-down impedito l'ancoraggio delle membrane T-tubuli per giunzioni SER conduce al ritardato rilascio di Ca 2 + e organizzazione TATS patologica coerente con architetture immaturo A-tubulo dominato in VM 13. Queste osservazioni possono sfociare proof-of-concept che T-tubuli formano attraverso processi di invaginazione considerando che un tubuli possono trasformarsi attraverso ulteriori o addirittura alternative meccanismi intracellulari 14.
Caratterizzazione dei tatuaggi variazioni di membrana nella malattia di cuore è diventato un settore importante della ricerca per le domande fisiopatologiche. I primi rapporti in un modello canino di stimolazione indotta cuore failure ha mostrato una perdita di T-tubuli e attuale Cav1.2 (I Ca) 15. Un modello suino di cardiomiopatia ischemica ha mostrato una riduzione di densità T-tubulo e una sincronia ridotta intracellulare di Ca 2 + rilascio 16. Utilizzando un ratto spontaneamente ipertesi (SHR) modello di scompenso cardiaco, una perdita di T-tubuli è stata associata con una ridotta accoppiamento nanodomain di Cav1.2 e RyR2 dal meccanismo proposto di "RyR2 orfano" 7. Una perdita di T-tubuli è stato dimostrato anche in macchine virtuali umana dal ischemici, dilatati, e ipertrofiche campioni cardiomiopatia 17. Inoltre, un aumento di A-tubuli è stato segnalato in sezioni di tessuto di cardiomiopatia dilatativa umana 18. A seguito di infarto miocardico, abbiamo mostrato un meccanismo differenziale di TATS riorganizzazione del mouse VM con diminuzioni significative di T-tubuli in contrasto con incrementi di componenti A-tubulo 9. È importante sottolineare che, migliorato il contrasto locale di membrana ottenuta attraverso superre cellule vivesoluzione microscopia STED abilitato dettagliata analisi componente quantitativa attraverso misure dirette, che hanno mostrato la proliferazione significativa degli A-tubuli con aumenti globali della lunghezza della rete TATS e ramificazione complessità 9. Inoltre, è stato dimostrato che l'esercizio fisico può invertire T-tubulo rimodellamento nei ratti dopo infarto miocardico 19 e che la terapia di risincronizzazione cardiaca può portare ad invertire il rimodellamento di T-tubuli in cani con tachypacing atriale indotta insufficienza cardiaca 20. Nel loro insieme, gli studi sia in potenziali interventi terapeutici umana e animale malato macchine virtuali così come potranno probabilmente beneficiare di procedure di isolamento delle cellule di alta qualità e strategie di analisi quantitativi dettagliati, come indicato nel protocollo e risultati sezioni di seguito.
Inoltre, come dimostrato recentemente dalla superficie laterale rispetto TATS traffico di membrana del canale KATP isoforme 21, è importante considerare m atrialeyocytes (AMS) come biologicamente distinti, nonché comparativa modello di cellule cardiache rispetto a macchine virtuali. T-tubuli sono state recentemente documentate negli ovini e umano AM 22. L'evidenza attuale suggerisce che esistono pochi tubuli T in cellule AM e in genere nei mammiferi più grandi come le pecore e gli esseri umani, ma non nei piccoli roditori 23. A differenza di macchine virtuali, in AMS rilascio intracellulare di Ca 2 + sembra verificarsi dalla propagazione superficie cellulare per diffusione verso il centro della cellula, che si traduce in contrassegnato Ca spaziale e temporale 2+ gradienti 23. In questo quadro mi sembra importante chiarire i meccanismi di Ca 2 + intracellulare di segnalazione di instabilità per le forme di malattie comuni come la fibrillazione atriale 24. In sintesi, sia AM e VM isolamento delle cellule e ciascuno per i cuori sani e malati sono comunemente impiegati protocolli. Solo se l'isolamento delle cellule è fatto correttamente, come giudicato da documentazione microscopica di cellule sufficiente qualità, AM e VM campioni devono essere carried avanti per l'analisi quantitativa TATS. Di conseguenza, le sezioni seguenti protocolli sono criticamente dipendenti da cellule di alta qualità isolati da mouse o altre specie seguite da microscopio cellule in vivo per analizzare membrane Tats intatte. Come sottolineato in precedenza, la caratterizzazione delle membrane Tats è un'area di ricerca impegnativo con una propensione per il fissaggio e la preparazione di manufatti 6, cambiamenti di membrana a causa di alterazioni osmotiche, e le limitazioni di risoluzione di microscopia ottica convenzionale 9. Notiamo che i recenti protocolli di state-of-the-art per l'isolamento dei macrofagi alveolari umani per Ca 2 + imaging e patch-clamp e di ratto macchine virtuali per colture cellulari sono stati precedentemente pubblicati in questa rivista 25,26.
Protocollo
NOTA: Tutte le procedure sugli animali sono stati esaminati e approvati dalla cura e l'uso degli animali Comitato Istituzionale della University Medical Center di Goettingen in conformità alla cura umana e l'uso di animali da laboratorio.
1 Isolamento di miociti atriali e ventricolari dal Cuore del mouse
- Maneggiare gli animali il più delicatamente possibile e secondo protocolli approvati per ridurre al minimo lo stress in generale e specificamente per evitare potenziali potenti effetti involontari da eccessi neuro-ormonali su cellule cardiache isolate. Inoltre, iniettare ogni mouse con eparina (500 UI / kg di peso corporeo sc) almeno 20 minuti prima dell'estrazione cuore per prevenire la coagulazione del sangue e micro emboli che possono compromettere significativamente la resa e l'integrità delle cellule cardiache durante l'isolamento.
- Anestetizzare topi dai 12 settimane di età per inalazione isoflurano, confermare l'assenza di riflessi ritiro dolore, e l'eutanasia degli animali da dislocazione cervicale.
- Estrarre il cuore rapidamente seguendo protocolli di esperti stabiliti in precedenza (ad esempio, vedere Kaestner et al. 26 e Louch et al. 27). Evitare qualsiasi possibile danno ai suoi atri da spremitura inutili o stiramento.
- Conservare con cura il tessuto della aorta ascendente prossimale utilizzando un paio di pinze e forbici moncone dritto per stabilire un tagliente trasversale continua attraverso la parete del vaso aortico, che è importante per la cannulazione successo e perfusione del cuore.
- Trasferire il cuore asportato immediatamente in ghiaccio-freddo nominalmente Ca 2 + buffer di perfusione gratuito (per le soluzioni vedi tabella 1). Tenere i grandi vasi fissati al suolo durante il trasferimento attraverso l'aria nel buffer per evitare involontarie embolia gassosa fino a quando il cuore è completamente sommerso. Utilizzare BDM nelle soluzioni tampone e ghiaccio raffreddato per inibire la contrazione cardiaca.
- Utilizzare un microscopio binoculare con zoom sufficiente illum 3Dinazione.
- Incannulare l'aorta sotto stereovisione panoramica del cuore con una superficie liscia e lucida 21 G cannula (esterno diametro 0,81 millimetri, per il normale peso del cuore del mouse) che deve essere completamente riempito con tampone. Assicurarsi che non ci siano bolle d'aria nella cannula collegando una soluzione serbatoio prossimale (ad esempio, una siringa) attraverso una valvola Luer 2 vie che consente un rapido controllo del flusso della soluzione.
- Confermare sotto ingrandimento binoculare che la cannula sia posizionata correttamente all'interno dell'aorta, che è di circa 1 mm al di sopra delle valvole aortiche e rami dell'arteria coronaria. Assolutamente evitare qualsiasi passaggio o perforazione accidentale delle valvole aortiche con la cannula (questo sarà definitivamente interrompere la chiusura della valvola aortica e di conseguenza interrompere la perfusione cardiaca).
- Legare l'aorta delicatamente per scanalature antiscivolo su misura, circonferenza orientata verso la fine della cannula utilizzando due punti di sutura in seta. Non lavare la arteri coronaricaes forza in qualsiasi punto. Collegare la soluzione riempito cannula legata alla aorta e il cuore di un connettore di deflusso strettamente montaggio di un sistema di perfusione su misura e pre-calibrato, aka il setup Langendorff modificata (sia con pressione costante o costante flusso, vedere anche la sezione discussione sotto).
- Profumato cuore appena possibile per 4 min utilizzando tampone perfusione ossigenato a 37 ° C (target frequenza perfusione: 4 ml / min). Avviare la digestione passando la perfusione di collagenasi contenente tampone di digestione (600 U / ml di collagenasi di tipo II) per 8-10 min a 37 ° C. Monitorare l'avanzamento della digestione dei tessuti confermando cambiamenti del tessuto simili, tra cui l'aumento opacità, morbidezza, e flaccidità tutta la superficie apparente cuore.
- Sezionare le camere cardiache come necessari i seguenti digestione. Mettere il cuore cannulata sotto un microscopio binoculare e visualizzare la parete posteriore del cuore. Sezionare residuo ad esempio tessuto non-cardiaco, polmonare unnd parti di nave per evitare la contaminazione delle cellule nel buffer di digestione con micro forbici (per esempio, le forbici a molla 8 mm lame dritte), come mostrato in Figura 1.
- Seguire una lista di controllo che particolare le camere, le regioni, e / o cellule del cuore collagenasi digerito dovrebbero essere raccolti (Figura 1): a sinistra e / o atrio destro, sinistro libero e / o parete del ventricolo destro, e / o del setto ventricolare.
- Per la dissezione di specifici tessuti cardiaci, utilizzare un bagno dissezione relativamente ampio e piatto rivestito con uno strato spesso diversi millimetri di plastica elastomero siliconico. Fissare l'apice del cuore con un bel perno in acciaio insetto allo strato di elastomero fondo.
- Deviare il diritto appendice atriale e sezionare l'atrio destro appena sopra le valvole atrioventricolari. Continuare la dissezione con l'atrio sinistro. Staccare e gettare l'apparato valvola fibrosa. Infine, sezionare le pareti ventricolari liberi destro e sinistro e del setto e / o piccoleer le parti di tessuto, se necessario.
- Nota solo per i tirocinanti: per guadagnare pratica, iniziare con il cuore non digeriti mouse. Per facilità di orientamento anatomico, praticare la manipolazione dei tessuti comprese tutte le fasi di dissezione consecutivi sotto la visione binoculare, come mostrato in Figura 1. Volta l'anatomia 3D, movimentazione manuale sotto visione binoculare, e passi di dissezione sono sufficientemente familiarità, continuare con le collagenasi digerito cuori mouse descritto sopra.
- Per ventricolare miociti (VM) isolamento delle cellule: trasferire il tessuto ventricolare in 2,5 ml di tampone di digestione fresco. Se isolamenti cellulari simultanee provenienti da più parti d'organo, ad esempio, atri e ventricoli è tentato, di una seconda persona può assumere una gamba della procedura di dissociazione cellulare, sia per minimizzare e ottimizzare l'utilizzo di topi attraverso la gestione coordinata di molteplici tessuti cardiaci. Continuare con i punti 1.9.1-1.9.4, successivamente 1.10.
- Sezionare sia l'intero tessuto ventricolareo di sue parti specifiche (ad esempio, LV, RV, pareti libere, e / o setto) in circa 1 millimetro 3 pezzi in 2,5 ml di tampone di digestione con forbici affilate (ad esempio, forbici a molla 8 mm lame dritte) in un piatto 60 millimetri Petri .
- Dissociarsi delicatamente VM in sospensione cellulare per lenta triturazione dei pezzi di tessuto con una pipetta di trasferimento. Evitare qualsiasi gorgogliare aria in sospensione cellulare.
- Aggiungere 8 ml di tampone di arresto alla sospensione cellulare VM e trasferire la sospensione cellulare in una provetta conica da 15 ml. Lasciare i pezzi di tessuto residui di depositarsi sul fondo per circa 15 secondi, ma abbastanza breve per cellule isolate di rimanere in sospensione. Successivamente, raccogliere la sospensione VM tramite trasferimento del volume surnatante in una nuova provetta da 15 ml. Se i pezzi di tessuto eccessive sono presenti, in alternativa, utilizzare una rete di nylon mini 200 micron distanziati per separare i pezzi di tessuto dalla sospensione cellulare.
- Lasciate che la sospensione cellulare VM depositano sul fondo di un 15 ml cotubo nica per gravità per 8 min.
- Lavare passo: rimuovere il surnatante e risospendere delicatamente il restante VM pellet in 10 ml di tampone di perfusione. Ripetere il lavaggio punto 1.9.5 (facoltativo: aggiungere ulteriori fasi di lavaggio per aumentare gradualmente la concentrazione di Ca 2 +, se necessario).
- Risospendere il pellet VM si stabilì a buffer di perfusione 10 ml e distribuire la cellula restante volume di sospensione in 1,5 ml provette da microcentrifuga (circa 50.000 cellule VM per provetta).
- Per atriale miociti (AM) isolamento delle cellule: trasferire il / tessuto atriale sezionato digerito in 1 ml di tampone di digestione fresco.
- Tagliare il tessuto atriale parzialmente digerito in ca. 1 mm 3 pezzi in tampone di digestione 1 ml utilizzando micro forbici in una piccola scatola di Petri (ad esempio, diametro 60 mm). Dissociare delicatamente le cellule sono fuori dei pezzi di tessuto digerito in sospensione cellulare mediante triturazione con una pipetta da 1 ml con una punta di taglio per evitare getti di fluido dannosi. Durantetriturazione rigorosamente evitare qualsiasi gorgogliare aria in sospensione cellulare. A seguito di agitazione meccanica, aggiungere 4 ml di tampone di arresto (50 mM CaCl 2, 10% BCS) per arrestare qualsiasi attività collagenasi rimanendo in sospensione cellulare.
- Trasferire la sospensione cellulare AM in un tubo da 15 ml. Lasciare i pezzi di tessuto residui di depositarsi sul fondo per circa 15 secondi, ma abbastanza breve per le cellule isolate di rimanere in sospensione. Raccogliere il volume di supernatante contenente le cellule sono libero tramite trasferimento soluzione in un tubo da 15 ml.
- Centrifugare la sospensione cellulare AM, ad esempio, 2 min a 20 xg a temperatura ambiente o - preferibile per gli studi di membrana - lasciare che le cellule si depositano lentamente per gravità per 20 minuti in un tubo da 15 ml.
- Lavaggio: scartare il surnatante e delicatamente risospendere il pellet in AM 5 ml di tampone di perfusione. Ripetere 1.10.4.
- Risospendere le cellule AM delicatamente nel buffer di perfusione 5 ml. Distribuire il volume di sospensione di cellule in 1,5 ml microcentrifuga vascaì (circa 1.000 AM cellule per tubo).
- Analizzare e documentare la qualità isolato popolazione di cellule per ogni cuore compreso il rendimento cella utilizzando trypan colorazione blu.
- Per questo, diluire 500 ml di sospensione cellulare come 1: 1 vol / vol con trypan soluzione di blu (concentrazione finale 0,02%) con 1 ml punte taglio pipette. Mescolare le cellule e il blu trypan delicatamente molto rallentare pipettaggio / giù. Applicare immediatamente il trypan blu contenente sospensione cellulare ad un Neubauer di tipo migliorato citometro e contare i miociti intatti utilizzando un microscopio invertito.
- Escludi eventuali cellule con danno apparente, vesciche membrana, striature di turbativa, contratture, e le cellule accumulando intracellulare trypan blu (Figura 2). Escludono anche spontaneamente contrarre le cellule, che sono suscettibili di successiva morte cellulare. Per valutare il numero di cellule intatte in sospensione, utilizzare solo i miociti cardiaci con striature regolari che escludono trypan blu tuttoil volume cellulare.
- Giudicare l'integrità dei singoli miociti atriali e ventricolari al microscopio a luce trasmessa. Salvare l'immagine in campo chiaro come file Tif per la documentazione e ulteriori analisi.
- Utilizzare i seguenti criteri per l'analisi dell'integrità delle cellule cardiache:
- confermare la presenza di striature regolari in tutto il volume cellulare visibile;
- confermare l'integrità continuo della membrana superficie laterale su entrambi i lati delle celle paralleli ai miofilamenti;
- visualizzare dentellature taglienti ai dischi intercalari su entrambi i lati delle cellule che riflettono l'integrità delle specifiche strutture a membrana di superficie; e
- visualizzare i segnali fluorescenti di membrane tatuaggi (o immunolabeled caveolina-3 proteine o altri marcatori di membrana), come descritto nel capitolo 3 per la localizzazione correlazione accanto al campo luminoso sottostante immagine specifica morfologia delle cellule (ad esempio, combinare entrambe le immagini con ImageJ come overlay imag compostoe).
- Determinare la lunghezza sarcomero dalle immagini in campo chiaro. Per la lunghezza sarcomero media, misurare la distanza della sequenza allineata striature sarcomero, e dividere la distanza per il numero di sarcomeri. Misurare almeno due posizioni per cella. Eseguire l'analisi con software commerciale o non in linea con ImageJ.
NOTA: Se trattati con agenti disaccoppiamento, intatto rilassato VM dal cuore di topo mostrano una lunghezza sarcomero media di ~ 1.9 micron 28.
- Utilizzare i seguenti criteri per l'analisi dell'integrità delle cellule cardiache:
- Quantificare la morfologia e le dimensioni dei singoli miociti atriali e ventricolari dall'immagine microscopia a luce trasmessa. Si consideri che le cellule AM e VM differiscono significativamente in termini di dimensioni. Misurare la lunghezza delle cellule, la larghezza, e la zona, e calcolare la lunghezza: rapporto larghezza.
- Analizzare le dimensioni delle celle 2D dall'immagine luce trasmessa in ImageJ utilizzando lo strumento di selezione comandi poligono e aggiungere al gestore ROI, analoga alla figura 3. Se morfologica cHANGES sono attesi all'interno di contesti specifici di studio, ulteriore documento per tutte le cellule del ceppo specifico del mouse, l'età, il sesso, le dimensioni del cuore, ed eventuali interventi per la successiva classificazione dei dati.
2 La colorazione di tatuaggi Membrane in Living miociti atriali e ventricolari
- Caricare la camera di imaging (ad esempio, POC-R2) con un coprioggetto di vetro 42 mm. Per myocyte fissaggio stabile al coprioggetto, preparare 20 ml di soluzione laminina dalla diluizione 1:10 della laminina magazzino in tampone di perfusione fisiologica (concentrazione finale di 0,2 mg / ml). Distribuire 20 ml di soluzione di laminina uniformemente sul coprioggetto di vetro.
- Preparare 800 ml di una soluzione 50 mM di-8-ANEPPS in tampone di perfusione. Per questo, diluire 20 ml di di-8-ANEPPS soluzione madre 2 mM in 780 microlitri di buffer fisiologico.
- Per macchiare VM, lasciare che le cellule si depositano per gravità per 8 minuti in una provetta da 1,5 ml di reazione. Per macchiare AM, utilizzare sedimentazione gravità o spnella sospensione di cellule per 2 minuti (vedi 1.10.3). Per entrambi emendamenti e VM, rimuovere con attenzione il surnatante evitando inutili agitazione del pellet e risospendere delicatamente il pellet cellulare in 800 ml di di-8-ANEPPS contenente soluzione (50 micron). Trasferire immediatamente il di-8-ANEPPS / sospensione miociti sul coprioggetto laminina rivestite in camera di imaging.
- Colorare la sospensione VM per 15 min a RT al buio.
- Rimuovere lentamente il volume in eccesso attraverso il menisco di fluido verso l'alto ai lati della camera di imaging con una pipetta manuale. Verificare che miociti la maggior parte dei di-8-ANEPPS macchiati rimane fermamente attaccata al coprioggetto laminina rivestite e non rimangano esposti all'aria. Successivamente, lavare la sospensione dei miociti allegata una volta aggiungendo lentamente 1 ml di tampone di perfusione seguiti da rimuovere il liquido in eccesso comprese le cellule non aderenti.
- Sovrapporre accuratamente i miociti colorati e di superficie collegati con 1 ml di tampone di perfusione lentamente dal lato di tche di imaging camera. Posizionare la camera di imaging sul palco microscopio.
3. Imaging delle strutture TATS membrana in Living miociti atriali e ventricolari
- In generale, scegliere con attenzione la migliore opzione possibile microscopio a fluorescenza (s) disponibile per l'imaging TATS membrana. Per l'imaging confocale, considerare recente generazione, moderni microscopi a fluorescenza con rivelatori a serie di PMT ottimizzate e percorsi di riciclaggio fotone che massimizzano l'intensità del segnale fluorescente. Per l'imaging confocale dei dettagli più piccoli di TATS strutture a membrana utilizzano un obiettivo 63X olio 1.4 NA o - a seconda della disponibilità - utilizzare un microscopio SuperResolution STED per i più piccoli particolari come tatuaggi recensioni, foto e applicazioni specifiche miociti da Kohl ed altri 34 dei principi generali di alta. risoluzione microscopia a fluorescenza, fare riferimento alla sezione di discussione.
- Impostare i parametri di imaging per rilevare la maggior parte, idealmente tutti mem intracellulare di-8-ANEPPS macchiatobrane all'interno di un determinato piano di imaging miociti. Utilizzare i seguenti parametri come punto di partenza per microscopio confocale a scansione laser: 458 nm di eccitazione ad esempio, al 3% della potenza laser massima; rilevare il segnale emesso tra 550 nm e 740 nm; guadagno del rivelatore (ad esempio, il comando maestro 800); e pinhole 1 UA per uno spessore fetta ottica di 900 nm. Regolare questi parametri per ottimizzare il rapporto segnale-rumore.
- Utilizzare la modalità di campo luminoso per selezionare AM intatto o cella VM a seconda dei casi (Figure 4A e 4B). Fare riferimento al punto 1.12 di criteri pertinenti come giudicare l'integrità cella per selezionare le celle come sintetizzato: striature regolari a livello di cellule e spaziatura uguale sarcomero, spigoli vivi e superfici dentellature su tutti e quattro i lati delle cellule, l'integrità continuo della membrana superficie laterale, e l'assenza di eventuali vesciche membrana.
- Prendete una immagine di esempio di una sezione miociti intracellulare centrale. Per regolare il ROI utilizzare il "raccolto" funzione →, Regolare la finestra di ritaglio → la dimensione finale del pixel misura 100 x 100 nm nm. Regolare l'asse x della finestra di ritaglio per corrispondere con la maggiore (assiale / longitudinale) asse del miociti.
- Selezionare il piano di imaging finale. Usare fotogrammi singoli di immagine per selezionare manualmente il piano di imaging appropriato nella direzione z. Verificare che le membrane tatuaggi, tra cui T-tubuli e componenti di A-tubulo, sono visivamente evidenti nel piano focale. Si noti che un piano di imaging intracellulare tipico può includere un nucleo come punto di riferimento intracellulare. Fare riferimento agli esempi delle figure 4A e 4B.
NOTA: In generale, mantenere l'esposizione delle cellule alla luce laser più breve possibile. Se possibile, utilizzare fotogrammi singoli di immagine per determinare il piano focale ottimale in miociti cardiaci. - Regolare il tempo di permanenza dei pixel di circa 0,5 msec. Selezionare 16x media e registrare l'immagine come istantanea. Ripetere il passo immagine istantanea di stabilire imaging appropriato piano di uns delineato per strutture a membrana tatuaggi in 3,5 se necessario.
- Salvare l'immagine finale e verificare che il file è stato salvato nella cartella di destinazione. In generale, salvare tutti i file di immagine nello stesso formato (ad esempio, LSM) per l'applicazione uniforme del software di analisi. Prima di qualsiasi analisi delle immagini, ancora una volta confermare l'integrità delle cellule sufficiente off-line (in considerazione criteri elencati nel punto 1.12.1) ed escludere eventuali cellule danneggiate da analisi. Fare riferimento agli esempi delle figure 4C e 4D.
4 Analisi della TATS membrana rete e dei suoi componenti
Le seguenti operazioni di elaborazione delle immagini per l'analisi diretta dei componenti della membrana tatuaggi sono riassunti come top-down diagramma del flusso di lavoro nelle Figure 5A e 5B.
- Aprire il file immagine di un miociti macchiato di-8-ANEPPS in Fiji (http://fiji.sc/), una variante liberamente disponibile di ImageJ, che contiene i plugin essenziali per l'analisielaborazione delle immagini. Per ulteriori informazioni consultare Schindelin et al 29.
- Salvare l'immagine → File → Salva con nome → Tif.
- Per analizzare le componenti della membrana Tats, selezionare il ROI adeguato escludendo il segnale di membrana superficie esterna, quindi utilizzare lo strumento "poligono di selezione" per delimitare il confine ROI escludendo la superficie esterna della membrana (sarcolemma) e comprendente le porzioni intracellulari delle membrane tatuaggi come mostrato in Figura 5A (ROI). Aggiungere il ROI selezionato alla "ROI Manager" applicando Analizza → Strumenti → ROI Manager.
- Per selezionare una ROI specifico per l'analisi orientamento dei componenti tatuaggi, allineare l'asse principale longitudinale cella e l'immagine asse x in parallelo. Se la cella è leggermente curva, selezionare vari ROI e allineare ogni ROI singolarmente. Escludere eventuali nuclei dall'analisi. Escludere tutte le cellule eccessivamente curvi dall'analisi a causa preciso allineamento della RIO diventerà sempre più difficile e aumentare gli errori di orientamento durante l'analisi.
- Eliminare tutte le informazioni segnale indesiderato dalla membrana superficie esterna: Modifica → Cancella Fuori per generare il "ROI selezionato" (Figura 5A). Assicurarsi che il ROI selezionata contiene solo le porzioni di membrana intracellulare, che corrispondono con la rete TATS.
- Eseguire la seguente catena di passaggi di elaborazione delle immagini (comandi) prima della successiva analisi quantitativa come documentato nella Figura 5A.
- Clicca su → Processo → Sfondo Sottrai. Impostare il raggio della sfera di rotolamento a 5 pixel.
NOTA: Impostare il raggio della sfera di rotolamento di 5 pixel se l'immagine deve essere analizzata ha una dimensione di pixel di 100 nm x 100 nm. Per altre dimensioni di pixel impostare il raggio di rotolamento della sfera per il numero di pixel che corrispondono approssimativamente ad un raggio fisica di 500 nm. - Clicca su → Processo → Migliora contrasto locale (CLAHE). Impostare la dimensione del blocco a 49, i bidoni istogramma a 256, la pendenza massima di 3, e la maschera di "nessuno".
- Clicca su → Processo → liscio.
- Clicca su → Plugin → Segmentazione → Regione statistico Fusione. Impostare i parametri da Q100 → fare clic su Mostra Medie.
- Confermare l'elaborazione completa della regione statistica merging indicato dalla presentazione automatica di una nuova cornice immagine e confermare che appare la label "SRM Q = 100". Continuare le seguenti operazioni con il file immagine. Fare clic su → Immagine → Tipo → 8 bit.
- Fare clic su → Immagine → Regola → Soglia. Scegli la soglia sufficientemente bassa per rilevare la maggior parte, idealmente tutte le strutture tatuaggi, in particolare evitare l'esclusione di componenti di tatuaggi con una bassa intensità di segnale (utilizzare una soglia di 40 come punto di partenza). Consultare la sezione "Rappresentante dei risultati" per l'output dettagliato di dati e gli esempi in Figura 6(Soglia superiore di 255). Documentare la scelta finale dei parametri di soglia. Si noti che la scelta corretta di soglia dovrebbe produrre solo specifiche strutture a membrana TATS ma non i segnali falsi positivi a causa del rumore di fondo.
- Confermare la corretta sovrapposizione di tatuaggi dettagli dell'immagine rispetto dati estratti scheletro, in particolare per i livelli di segnale intermedio e alto fluorescenza, che dovrebbe corrispondere al (display) la continuità della struttura scheletro. Una volta che una soglia appropriata è stata identificata, applicare questa stessa soglia a tutte le immagini durante l'analisi e ripetere il confronto sovrapposizione del segnale originale rispetto a dati scheletro per minimizzare costantemente questa potenziale fonte di bias.
- Fare clic su → Applica: i dati immagine diventa binario come mostrato in Figura 5 sotto "Threshold".
- Clicca su → Plugin → Skeleton → skeletonize (2D / 3D). Salvare l'immagine 2D scheletrato (come mostrato in Figura 5) in formato Tif dacliccando su → File → Salva con nome → Tif. Analizzare il file di immagine scheletrato per l'output dei dati quantitativi cliccando su → Plugin → Analizza Skeleton (2D / 3D). Scegli il metodo del ciclo Prune: nessuno. Confermare la generazione automatica della tabella dati risultanti.
- Salvare la tabella di dati generati automaticamente come file txt. Selezionare i parametri quantitativi relativi ad esempio, il numero totale di punti di ramificazione o la lunghezza media del ramo, come mostrato dai dati esemplari in figura 7. Considerare ulteriori analisi dei dati con supporto / strumenti software complementari come Excel e come del caso.
- Clicca su → Processo → Sfondo Sottrai. Impostare il raggio della sfera di rotolamento a 5 pixel.
- Considerare di utilizzare le routine automatiche di elaborazione delle immagini, quando possibile includendo tutte le misure necessarie descritte al 4,5 per armonizzare l'analisi tra le singole immagini e / o lotti di immagini. Utilizzare l'esempio File Codice supplementare che contiene una macro di elaborazione delle immagini programmato per Fiji (regolare la programmazione in base alle esigenze).
- Analizzare l'inorientamento individuale di tutti o selezionare componenti di rete TATS dai dati di immagine scheletrato da parte del Fiji plugin "Direzionalità". Genera un istogramma direzionalità in cui la rispettiva A-tubulo assialmente orientate o componenti T-tubulo trasversalmente orientati sono rappresentati dai 0 ° o 90 ° bidoni. Notare il riferimento corretto di orientamento dell'immagine e che è importante per l'asse x dell'immagine da corrispondere a stretto contatto con i principali (longitudinali) 0 ° asse della cella VM come mostrato in figura 8 e Risultati rappresentativi.
- Fare clic su Analizza → → → Direzionalità specificare Metodo: componenti di Fourier, Nbins 180, Istogramma avviare -45 → cliccare sulla tabella di visualizzazione.
- Salvare la tabella dei risultati appena generato e visualizzato compresi i dati di istogramma associati come file txt. Considerare ulteriori analisi dei dati del file txt da strumenti software gratuiti come Excel e come necessario.
- Per generare sottoclassi didati come set di dati raggruppati, ad esempio per tutte le cellule trattate sotto la stessa condizione (e potenzialmente di altre condizioni), ripetere l'analisi i passaggi 4,1-4,7 per tutte le immagini pertinenti a seconda dei casi. Importare i parametri dei dati scheletrato da tutte le immagini in un unico file di Excel combinati al fine di ricavare valori medi. Inoltre, importare tutti i dati direzionalità istogramma dallo stesso insieme di dati raggruppati in un file di Excel combinata per calcolare e generare l'istogramma media direzionalità. Prendere in considerazione l'ulteriore elaborazione di set di dati raggruppati per analizzare i parametri di rete TATS aggiuntivi di interesse per i gruppi di trattamento individuali o tra diversi a seconda delle necessità.
Risultati
Inoltre, per l'analisi di rete membrana TATS una serie di tecniche di biologia delle cellule comunemente usate come intracellulare di Ca 2 + di imaging, patch-clamp elettrofisiologia, o studi di dose-risposta farmacologici dipendono criticamente su di alta qualità di isolamento cellula primaria dagli atri o ventricoli o selezionare parti di tessuto cardiaco per consentire la caratterizzazione di maturi miociti cardiaci intatti differenziati, strutturalmente e fisiologici. Pertanto, la valutazione di isolamento e di qualità per AM e VM cellule descritte nel paragrafo 1 sono in ultima analisi, utili per molte questioni diverse, tra cui l'analisi di rete TATS qui descritta, che dipende in modo critico su membrana intatta e l'integrità delle cellule.
La figura 1 fornisce un manuale graduale di immagini come procedere con la dissezione del tessuto cardiaco a partire dalle camere atriali nel cuore del mouse. Successivamente, le camere ventricolari e del setto sono preparati und sezionato come necessario. Selezione precisa e preparazione dei pezzi di tessuto corrette è importante stabilire in modo affidabile AM e VM isolamenti con sufficiente purezza delle cellule. Dopo digestione con collagenasi può essere relativamente difficile individuare la corretta linea di dissezione tra atriale e ventricolare tessuto, ancora una volta cellule AM e VM sono mescolati incontrollata in sospensione cellulare, è impossibile invertire la popolazione cellulare mista. Pertanto, l'orientamento anatomico, la visualizzazione del tessuto 3D, sufficiente esperienza con la manipolazione del tessuto digerito, corretta identificazione delle parti di tessuto specifici e le loro linee di dissezione saranno tutti contribuire al successo di isolamento delle cellule.

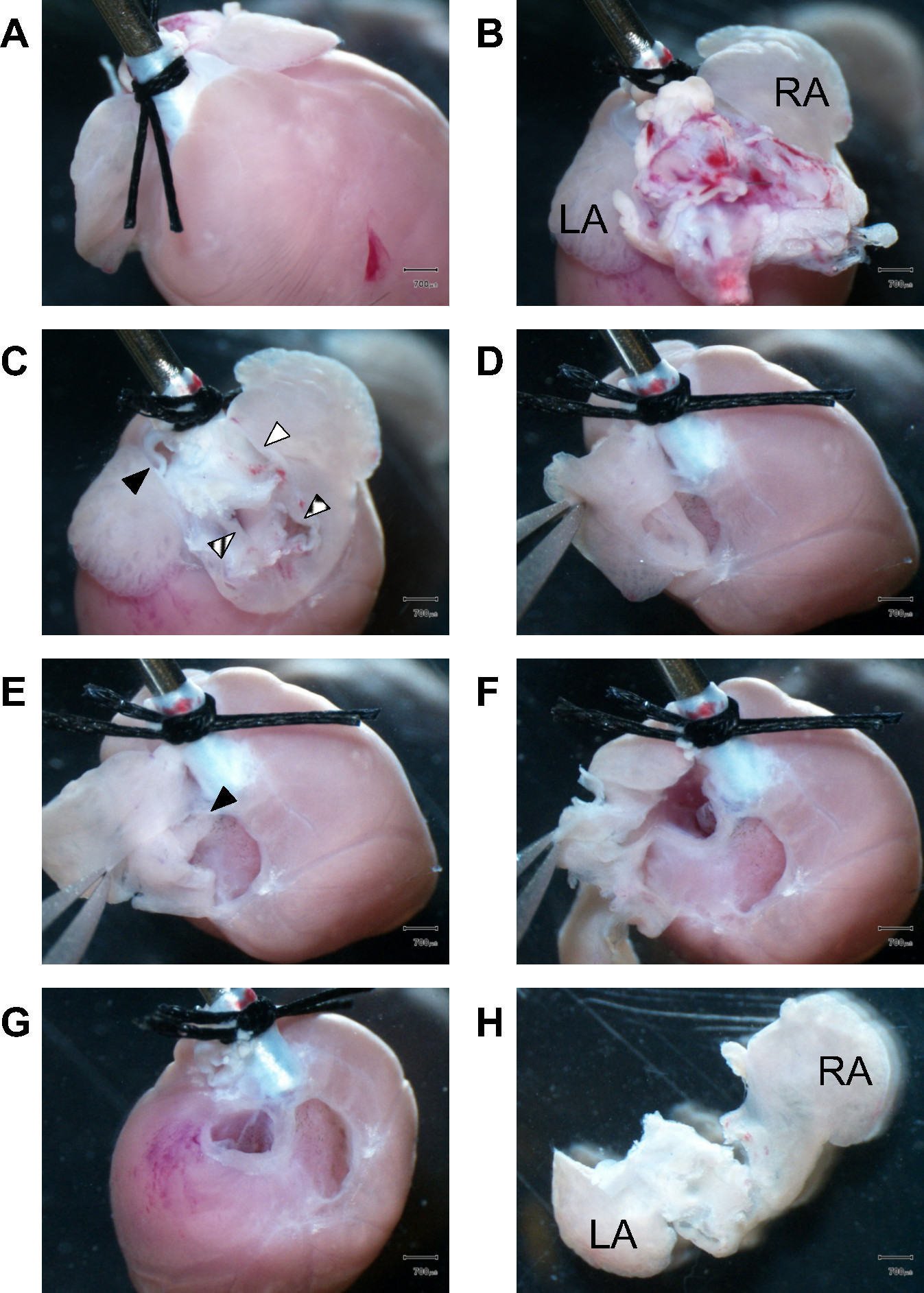
Figura 1.Dissection del tessuto atriale. (A) rivolto verso la parte anteriore del cuore, due s chirurgiciutures fissano l'aorta prossimale alla fine moncone di una cannula 21 G in acciaio. (B) Vista verso gli spettacoli di base cuore rimanenti tessuto polmonare ostruire la vista della camera atriale durante la dissezione. LA, atrio sinistro; RA, atrio destro. (C) rimanente lungtissue e dei grossi vasi sono stati rimossi per accedere alle camere atriali. Riempito triangolo nero, arteria polmonare; triangolo striato, vene polmonari; pieno triangolo bianco, vena cava superiore; triangolo boxed, vena cava inferiore. (D) In primo luogo, la parete atriale destra è sezionato, mentre la pinza tenere l'appendice atriale. (E) Visualizza nella giusta cavità atriale. Il triangolo nero segna il setto interatriale intatto. (F) La dissezione è continuato a entrare sinistra atriale cavità. (G) Dopo completa dissezione degli atri destro e sinistro, le valvole atrioventricolari diventano visibili. L'apparato valvola fibrosa è sezionato e scartato a Colluently raccogliere solo tessuto muscolare ventricolare. vista (H) posteriore degli isolati atri destro e sinistro. LA, atrio sinistro; RA, destra atrium.Scale bar: 700 micron.
Per determinare la qualità di isolamenti cellulari, Figura 2 fornisce esempi di cellule tipiche durante la valutazione del rendimento e la redditività di canna a forma di tipico o miociti intatti striati di mattoni, sia per AM e VM. Poco danneggiato, visivamente appariscente così come le cellule gravemente danneggiate con morfologie eccessivamente curve anomale, o cellule anomale a forma sferica, può essere facilmente identificato tra cui trypan blu come descritto nella sezione 1.11. Mentre miociti intatti rimangono luminoso e omogeneo striata se esposti a trypan blu extracellulare, le cellule danneggiate mostrano in genere più vesciche di membrana e / o rapidamente si accumulano trypan blu intracellulare che indica i danni della membrana. Tuttavia, blu trypan di per sé in grado di danneggiare le cellule attraverso unnecessarily lunga incubazione e immediata valutazione della qualità delle cellule è quindi obbligatoria. Esempi di forme più evidenti di danno cellulare come contrattura myofilament o danni superficiali compromettere l'integrità delle cellule lordo sono riportati nelle figure 4C e 4D. Cliccate qui per vedere una versione più grande di questa figura.
{kind=link}

Figura 2 Trypan colorazione della cellula esclusione blu. Isolato (A) AM e cellule (B) VM sono mescolati in sospensione con trypan blu e visualizzati da un microscopio ottico invertito mostrato a 40Xmagnification. Nota thatabnormal cellule sferiche in (A) e (B) che occupano trypan blu, che indicatesmembrane leakage e danni strutturali. Al contrario, le cellule centralAM e VM con membrane intatte escludono blu trypan come mostrato. Inoltre, notare che AM intatta e le cellule VM mostrano striature sarcomero in tutto il loro volume cellulare, blebs nomembrane e spigoli vivi ai due lati laterali e due dischi intercalari. Barre di scala: 20 micron.
Dopo rendimenti isolamento, cellulari di successo su macchine virtuali da 5 x 10 Maggio-10 Giugno ci si può aspettare da un singolo digestione cuore del mouse. La resa di AMS è significativamente inferiore nell'ordine di esclusione celle 3 x 10 3 a 3 x 10 4 a forma di bastoncello, blu trypan. In contrasto con VM, AM isolamenti a volte falliscono anche in mani esperte. Passo 1.11 riassume le procedure come stimare la resa di isolate cellule sane in sospensione. Inoltre, determinare le dimensioni medie delle celle attraverso passo 1.13 come mostrato in figura 3 per AM individuale o ceppi cellulari VM o per confrontare AM contro popolazioni cellulari VM side-by-side (se necessario). Cliccate qui per vedere una versione più grande di questa figura.
{kind=link}

Figura 3.Bright analisi morfometrica campo dei miociti cardiaci. Il profilo VM è stato rilevato dagli strumenti di analisi delle immagini descritte al punto 1.13. Utilizzare lo strumento di selezione poligono visivamente marchio e definire il confine extracellulare delle cellule definito dalla membrana superficie esterna per l'analisi. Ad regione selezionata di interesse (ROI) al gestore ROI seguita da misure di distanza 1D. Per gli studi comparativi di AM vs dimensioni VM, è utile per documentare la lunghezza delle cellule, la larghezza, e la zona, e per calcolare la lunghezza: rapporto larghezza.
{kind=link}

Figura 4.Live marcatura di membrana delle atriale e ventricolare intatto myociti. corrispondenti trasmessi luce e immagini confocali di vita di-8-ANEPPS macchiato intatto (A) AM e cellule (B) VM. Al contrario, un AM parzialmente contratta e potenzialmente danneggiato con una lunghezza sarcomero di 1,2 micron è mostrato in (C). Miociti contratti in genere mostrano anormalmente accorciati e strutture Tats distorte, pertanto esclusi da ulteriori analisi. Un altro indicatore importante per i difetti della membrana cellulare sono vesciche di membrana (triangoli rossi) come mostrato in una VM in (D). Vesciche membrana rappresentano strutture a membrana superficiali danneggiate e cellule con vesciche dovrebbero essere esclusi da ulteriori analisi TATS. Inoltre, la VM mostra danno grave a quanto pare manca una parte intera della sua porzione in basso a sinistra (contrassegnato da asterisco). In sintesi, confrontando luce trasmessa e immagini confocali, le cellule morfologia e l'integrità della superficie è documentato e combinati con le informazioni segnale fluorescente. Segni 'N' nuCLEI omesso dall'analisi della macchia membrana TATS. Le barre gialle indicano ingrandite ROI dalla stessa immagine confocale presentata sopra. Barre di scala:. 10 micron Cliccate qui per vedere una versione più grande di questa figura.
{kind=link}
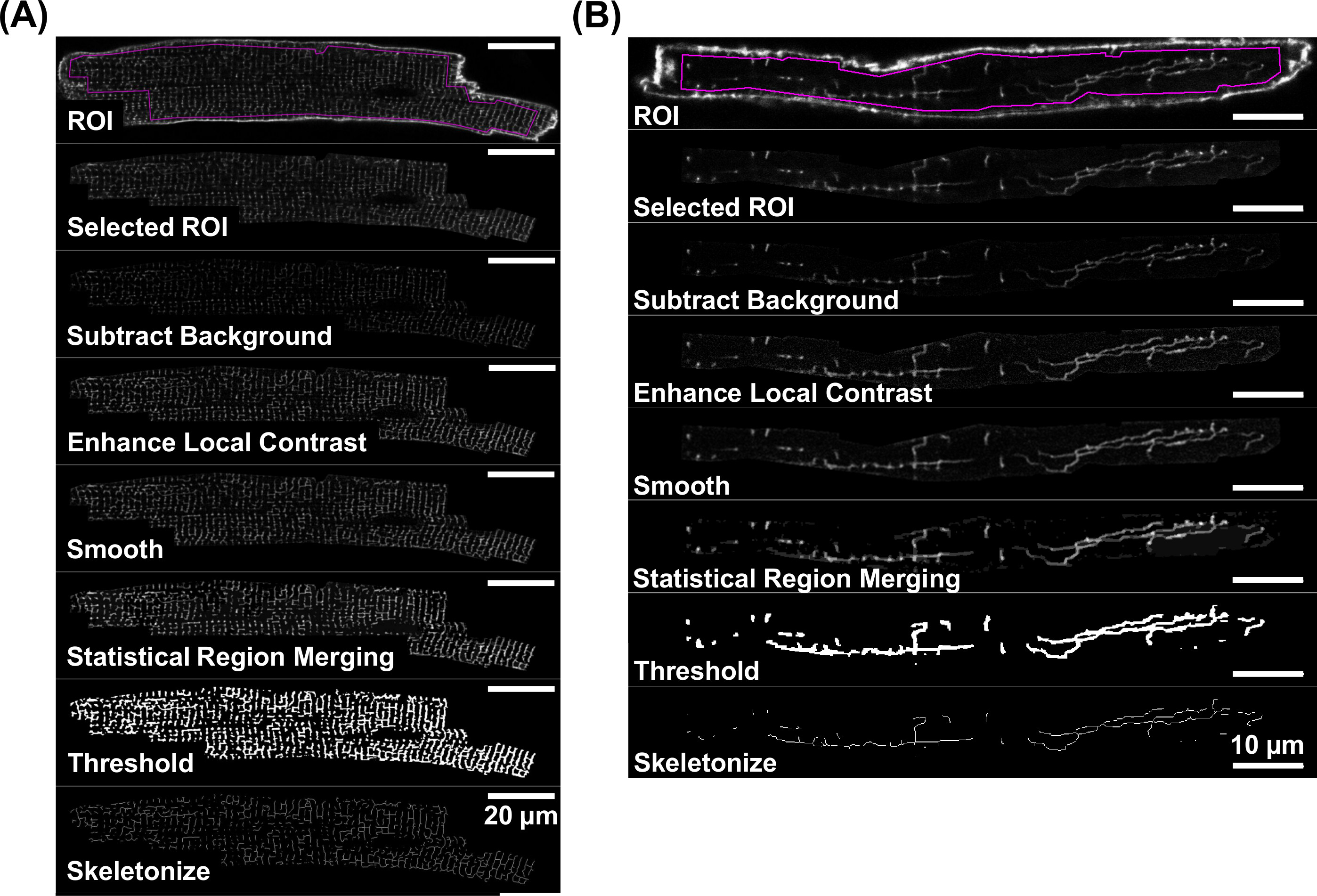
Immagini confocali di membrane tatuaggi con un sufficiente rapporto segnale-rumore, come mostrato nelle figure 4A e 4B sono accettati per ulteriori analisi quantitativa. L'analisi membrana TATS si basa sui dati skeletonizzati derivati da componenti del segnale rettilinee fluorescenti. Figura 5 mostra il diagramma di flusso di lavoro dei singoli passi di elaborazione dell'immagine che sono descritti in dettaglio da passaggi 4,3-4,5. Questi passaggi producono immagini scheletrato che rappresentano TATS rettilinei reti di membrana, come indicato per ciascun isolato VM (Figura 5A) e AM cellule (Fifigura 5B).

Figura 5.Workflow per scheletrizzazione di immagini di tatuaggi fluorescenti. Fasi di lavorazione Immagine che portano ad un'immagine scheletrato della rete TATS sono rappresentate da singoli step-by-step esempi di immagini sia per una di-8-ANEPPS macchiato VM (A) e AM (B). Per la fase di lavorazione singola immagine, si prega di fare riferimento alla sezione 4. Nota le differenze di barre di scala:. 20μm (A) e 10μm (B) Si prega di cliccare qui per vedere una versione più grande di questa figura.
{kind=link}
Un passo fondamentale durante l'elaborazione dell'immagine, determinare la soglia adeguata per binarizzazione dati come descritto nella sezione 4.5.6. L'immagine binaria risultante dovrebbe includere solo i segnali di membrana veri dalla rete TATS ma non falsi strutture derivate dalla soglia erronea dal rumore segnale di fondo. Tuttavia, è importante che la soglia è sufficientemente bassa per rilevare tutte le strutture tatuaggi veri tale che i componenti veri tatuaggi non sono erroneamente persi durante l'analisi dell'immagine. Figura 6 illustra il processo come selezionare la soglia durante binarizzazione dati. Mentre una soglia di 40 come mostrato nelle figure 6B e 6C sembra opportuno rilevare tutte le strutture tatuaggi veri singolarmente, scegliendo una soglia più elevata per esempio, di 60 come mostrato nella Figura 6A non rileva strutture a membrana debole assiali (ATS) come indicato da triangoli gialli . Al contrario, la scelta di una soglia inferiore esempio, 20 come mostrato nella Figura 6D porta alla rilevazione errata di strutture tatuaggi come falsi-positivi rumore di fondo come indicato dallatriangoli gialli.

Figura 6.How per determinare la soglia di segnale durante skeletonizing dei tatuaggi dati di immagine. Gli esempi mostrano scheletri Tats ogni generato per diverse soglie durante binarizzazione dati (di cui al punto 4.5). Immagini superiori: mostrano le regolazioni di soglia utilizzando Fiji. Immagini inferiori: sovrapposizione di immagini scheletrato l'immagine in ingresso fluorescenza corrispondente con le regioni e ingrandite come indicato. Un alto esempio di soglia, 60 applicato in (A) non è apparentemente idonei a rilevare tutte le strutture tatuaggi veri come indicato da triangoli gialli in sezione ingrandita. Una soglia di 40 applicata a (B) e (C) rileva tutte le strutture tatuaggi e correttamente non rileva il rumore di fondo, mentre una bassa soglia per esempio., 20 in (D) identifica erroneamente il rumore di fondo come strutture tatuaggi e, quindi, produce segnali falsi-positivi di strutture a membrana non-esistenti. Segnali falsi-positivi sono indicati da triangoli gialli nel riquadro ingrandita. Cliccate qui per vedere una versione più grande di questa figura.
{kind=link}
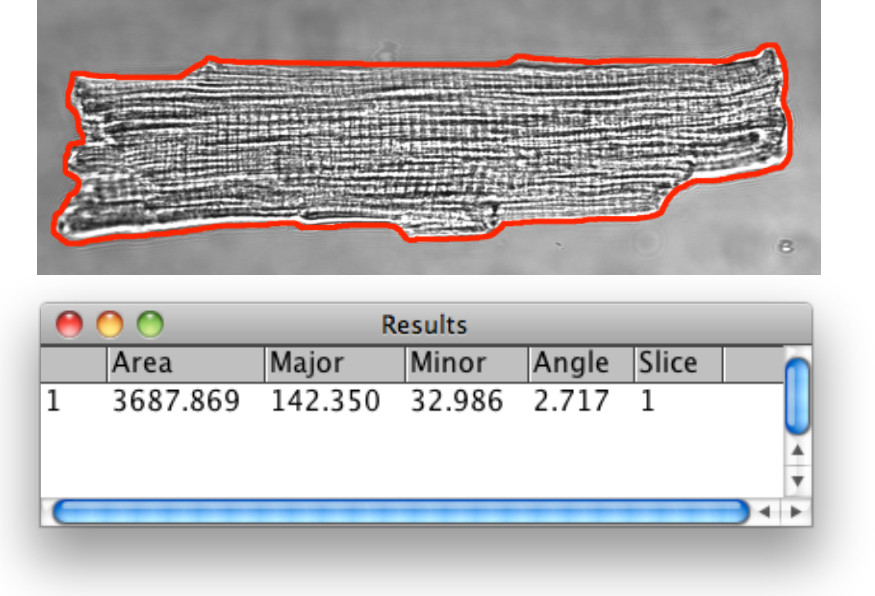
Il "Analizza Skeleton (2D / 3D)" plugin supporta l'analisi dettagliata delle strutture Tats scheletrato. Una volta eseguito, il plugin genera una tabella di dati con i seguenti parametri: scheletro #branches, #junctions, # end-point voxel, lunghezza media ramo, punti #triple, punti #quadruple, e la massima lunghezza del ramo. Per una descrizione dettagliata di tutti i possibili parametri di uscita consultare sitohttp: //fiji.sc/wiki/index.php/AnalyzeSkeleton e articoli correlati 29-31. Un tipico output della tabella dati è mostrato in Figura 7A.
I parametri possono essere ulteriormente utilizzati per ricavare la lunghezza scheletro totale per area o il numero di giunzioni per area. Calcolo esemplare del numero di rami moltiplicato per la lunghezza media ramo dà la lunghezza totale dello scheletro continua in 2D:Lunghezza scheletro totale per ROI:
Σ (#branches x lunghezza media branch) = 5155 px = 515,5 micron
La lunghezza totale dello scheletro può essere normalizzato alla zona dell'immagine. Per l'esempio mostrato nella figura 7 la lunghezza normalizzata scheletro di 0.64μm / micron 2 e la somma di tutte le giunzioni sono calcolate come segue:
Normalizzato Lunghezza scheletro:
515,5 micron / 803,6 micron 2 = 0.64 micron / micron 2
Numero normalizzato di giunzioni:
155 nodi / 803,6 micron 2 = 0,19 giunzioni / micron 2

Figura uscita dati 7.Automated da immagini scheletrato. (A) Un tipico foglio di dati generati dal 'Analizza Skeleton (2D / 3D)' plugin dal scheletrato immagine mostrata in (B). Per una descrizione dettagliata dei possibili parametri di output consultare http://fiji.sc/wiki/index.php/AnalyzeSkeleton . Cliccate qui per vedere una versione più grande di questa figura.
{kind=link}
Le fasi di elaborazione delle immagini descritte in 4.5 e illustrati nella figura 5 possono essere automatizzati utilizzando una macro Fiji fornito come Codice supplementare File . I comandi definiscono ripetizioni di elaborazione delle immagini attraverso passaggi reiterativi. La macro può essere applicato a completare pile di immagini in ingresso prodotte attraverso fasi 4.3 e 4.4. La macro può essere vantaggioso per l'analisi dei gruppi completi set di dati, ad esempio predisponendo singole pile di immagini in ingresso per ciascun gruppo di trattamento indipendente per l'analisi automatizzata utilizzando i comandi di macro.
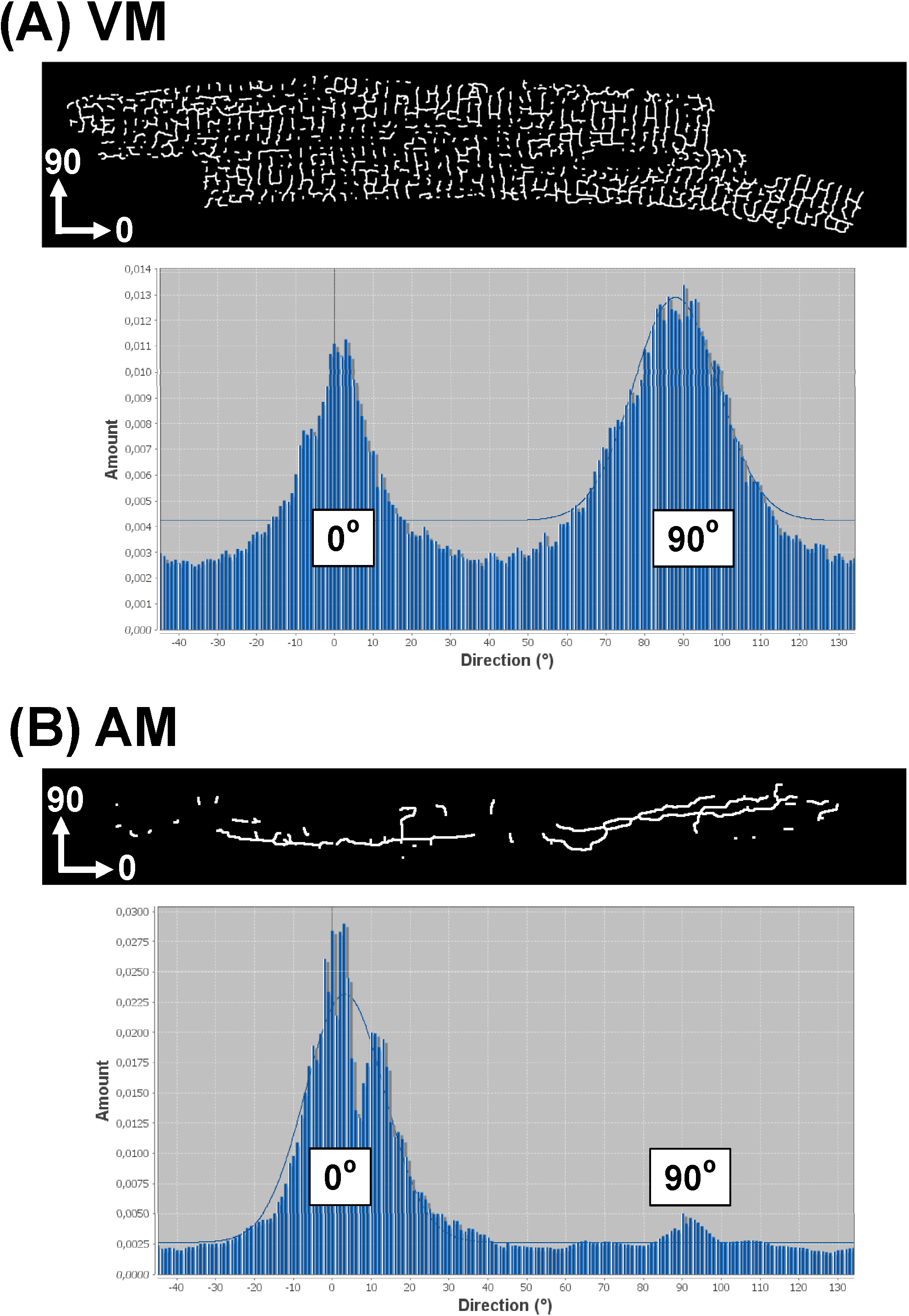
La strategia delineata software consente inoltre l'analisi orientamento rete TATS per tutti i componenti. Per questo, utilizzare il plugin "Direzionalità" (http://fiji.sc/Directionality) 29,31 che produce i dati di istogramma che mostra la distribuzione di orientamento di tutti i TATS orientamenti componenti. Se l'asse x dell'immagine in ingresso corrisponde all'asse principale di un determinato AM o VM cella, il assiale (longitudinale) componenti tatuaggi sarà rappresentato dallo 0 ° bin, che gli elementi trasversali saranno rappresentati dal 90 &# 176; bin. figura 8 mostra direzionalità esemplare istogrammi di immagini Tats scheletrato per una VM (8A) contro una (8B) cell AM. Mentre l'istogramma direzionalità di un tipico VM mostra una distribuzione doppio picco a 0 ° e 90 °, l'istogramma AM mostra un singolo picco dominante a 0 °. Questi esempi sono in accordo con precedenti osservazioni che i singoli componenti tatuaggi di macchine virtuali sono quasi equamente distribuiti tra i tubuli T e A-tubuli, mentre le componenti tatuaggi in AMS possono essere prevalentemente composti da A-tubuli.

Figura 8.Representative direzionalità istogramma da Tats reti di cellule singole. Istogrammi Direzionalità sono stati generati da immagini scheletrato dei singoli VM (A) rispetto a AM (B ) reti Tats. Dato che l'asse x dell'immagine analizzata corrisponde alla principale (longitudinale) asse del miociti come mostrato, componenti A-tubulo sono rappresentati dal 0 ° bidone, mentre componenti T-tubulari sono rappresentati dal bidone 90 °. Gauss si adatta solo per la grande peakasshown istogramma come sfondo grafico. Cliccate qui per vedere una versione più grande di questa figura.
{kind=link}
| Buffer di perfusione | mM |
| NaCl | 120.4 |
| KCl | 14.7 |
| KH 2 PO 4 | 0.6 |
| Na 2 HPO 4 | 0.6 |
| MgSO 4 | 1.2 |
| HEPES | 10 |
| NaHCO3 | 4.6 |
| Taurina | 30 |
| 2,3-butanedione-monoxime | 10 |
| Glucosio | 5.5 |
| pH 7.4 | |
| Buffer di digestione | mM (se non specificato) |
| NaCl | 120.4 |
| KCl | 14.7 |
| KH 2 PO 4 | 0.6 |
| Na 2 HPO 4 | 0.6 |
| MgSO 4 | 1.2 |
| HEPES | 10 |
| NaHCO3 | 4.6 |
| Taurina | 30 |
| 2,3-butanedione-monoxime | 10 |
| Glucosio | 5.5 |
| Collagenase tipo II | 600 U / ml |
| pH 7.4 | |
| Tampone di arresto | mM (se non specificato) |
| NaCl | 120.4 |
| KCl | 14.7 |
| KH 2 PO 4 | 0.6 |
| Na 2 HPO 4 | 0.6 |
| MgSO 4 | 1.2 |
| HEPES | 10 |
| NaHCO3 | 4.6 |
| Taurina | 30 |
| 2,3-butanedione-monoxime | 10 |
| Glucosio | 5.5 |
| CaCl 2 | 0,0125 |
| siero bovino di vitello | 10% |
| pH 7.4 | |
Tabella 1.Buffer solutioni. Il contenuto di tre diverse soluzioni fisiologiche tampone per l'isolamento delle cellule e di imaging sono riassunte.
Discussione
Anche se miociti cardiaci sono stati isolati e studiati per decenni 32, una recente revisione ha concluso che l'alta qualità isolamenti cellulari dei miociti coerenti rimangono impegnativo 27. Ciò riflette i protocolli relativamente complessi per l'isolamento dei miociti cardiaci primari nei confronti di una mancanza di approcci standard comuni, metadati condivisi, e la documentazione di qualità cellulare trasparente. Protocolli di isolamento delle cellule di solito sono personalizzate da singoli gruppi, produrre risultati variabili di cellule isolati, dipende dalle impostazioni individuali del modello (ad esempio, specie, età, coesistenti malattie cardiache), e di solito sono regolati per particolari condizioni sperimentali. Nel contesto del TATS quantitativa studi e protocolli di membrana qui presentati, un livello essenziale di valutazione della qualità e della documentazione riguarda la confocale o SuperResolution microscopio di singole strutture della membrana cellulare inclini a metaboliche e protocolli isolamento variazioni dipendenti, siain AM o VM. È importante sottolineare che, anche se alte rese di ceppi cellulari suggeriscono miociti sana ed intatta, gli investigatori hanno bisogno di documentare e valutare criticamente attentamente ogni singola cella in base a criteri morfologici di superficie e l'integrità della membrana TATS contro il danno non specifico a causa di procedure di isolamento rispetto a specifici cambiamenti dovuti a diversi tipi degli interventi rispetto alle condizioni di controllo. Una variabile importante durante l'isolamento delle cellule cardiache è l'attività specifica di un dato lotto collagenasi. Per selezionare un nuovo lotto di collagenasi, l'attività enzimatica di diversi campioni collagenasi deve essere testato contro l'altro valutando la resa dei miociti cardiaci e di qualità, e secondo le istruzioni del produttore. Idealmente, un nuovo lotto di collagenasi si identifica con l'attività collagenasi simile a precedenti lotti utilizzati con successo (per la valutazione estesa di possibili attività enzimatiche fare riferimento al "strumento di selezione collagenasi molto" nella tabella materiali e metodi). TAken insieme, approcci quantitativi di visualizzazione membrana TATS dipendono criticamente sulla qualità isolamento delle cellule e, viceversa, isolamenti cellule cardiache che portano a danni della membrana non specifica come documentato mediante microscopia TATS dovrebbe attivare revisione critica e correzione delle procedure di isolamento. Dal momento che la qualità di isolamento delle cellule e la visualizzazione della membrana TATS e la quantificazione sono intrinsecamente legate, i protocolli descritti in questo articolo coprono tutti i principali aspetti come strategia di continuo.
Un'ulteriore sfida e problema comune di studi cardiaci, danni cellulari e / o perdita di cellule si verificano a causa di interventi metabolicamente compromettenti per esempio, a seguito di infarto miocardico 9, ma devono essere giudicati da possibili danni accidentali ad esempio, a seguito di embolia gassosa inosservato durante l'isolamento delle cellule. Isolamento dei miociti cardiaci da cuori malati può portare a ulteriore, significativa perdita di cellule e una diminuzione dei rendimenti delle celle. Dunque, comparIson del numero totale di isolate cellule intatte tra controllo e cuori malati può essere significativo se l'isolamento e il conteggio delle cellule vengono costantemente applicati attraverso protocolli standardizzati. Di conseguenza, è fondamentale per giudicare l'integrità cellulare attraverso un gruppo di controllo adeguato, che riflette la migliore qualità possibile isolamento delle cellule miociti. È importante sottolineare che la qualità singola cella e microscopia cellulare dal vivo di un sano rispetto malata contro miociti inavvertitamente danneggiati dalla procedura di isolamento può influenzare in modo significativo l'analisi dei tatuaggi reti di membrana. I protocolli qui presentati sottolineano quindi l'integrità e la stabilità dei componenti della membrana fisiologiche durante l'isolamento delle cellule e microscopia di cellule in vivo delle membrane intatte. L'intero flusso di lavoro è concepito come una strategia continua per raggiungere e conservare componenti della membrana TATS intatte mentre escludendo le cellule danneggiate, dal momento che questi esporranno isolamento manufatti membrana dipendenti come tubuli di membrana perturbato, a membranaE blebs e reti tatuaggi alterati erroneamente in condizioni di controllo e di compromettere ulteriormente l'analisi quantitativa. Viceversa, le stesse strategie sono fondamentali per gli studi di intervento con il potenziale di distruggere le membrane Tats, che dipendono criticamente sui controlli confronto fra vero sano contro la vera cellula malata con tatuaggi membrana modifiche.
Inoltre, ci rivolgiamo procedure per realizzare l'isolamento tecnicamente molto più impegnativo di cellule AM. Nonostante i progressi e protocolli migliorati, è importante sottolineare che non è banale riprodurre isolamenti cellule di alta qualità di macchine virtuali e ancor meno affidabile per AM. Ciò è dovuto al rendimento complessivo inferiore di cellule AM dove anche piccoli errori o variazioni durante l'isolamento delle cellule può portare al fallimento completo di isolamento cellulare AM, mentre un lieve grado di danno cellulare VM potrebbe essere meno evidente in sospensione cellulare a causa della relativamente alta cella numeri rispetto al mattino. Poiché le cellule AM potrebbero diventare curved dopo l'isolamento, l'analisi attraverso diversi ROI può essere vantaggiosa come indicato sotto passo 4.3. A seguito di una procedura dettagliata di misure di isolamento delle cellule, mettiamo a disposizione un protocollo per la colorazione diretta integrale di membrana e confocale o STED SuperResolution imaging reti tatuaggi sia per VM e AMS. Questi protocolli consentono sia l'analisi quantitativa e la differenziazione di selezionare i componenti delle membrane tatuaggi attraverso i parametri precedentemente stabiliti. Rispetto al VM, l'organizzazione 3D e comportamenti funzionali della rete TATS atriale in AMS sono attualmente meno capito.
Le procedure per membrane immagine Tats in cellule viventi (passi da 3.1 a 3.7) sono stati sviluppati con confocale commerciale (Tabella dei materiali / attrezzature) e STED microscopi a fluorescenza misura 9. Per ottimizzare le impostazioni del microscopio per la generazione di immagini a fluorescenza e analisi TATS quantitativa, i seguenti punti sono di importanza generale:
- Obiettivo
Al fine di risolvere i piccoli dettagli di tatuaggi strutture a membrana, empiricamente testare obiettivo che fornisce la più alta qualità dell'immagine, mentre la messa a fuoco diversi micrometro profondità nella cella. Alcuni microscopi confocale possono funzionare meglio con gli obiettivi d'acqua o glicerolo, in contrasto con l'obiettivo 63X 1.4 NA olio usato qui. Obiettivi con ingrandimento 100X sono utilizzati per SuperResolution STED microscopia, la negoziazione fuori un campo più piccolo di vista per la risoluzione nanometrica 34. - Eccitazione e Gain
Le impostazioni ottimali della potenza di eccitazione e il guadagno del rivelatore dipendono dal percorso ottico del microscopio, le prestazioni del laser, e le proprietà del campione. Idealmente, potenza del laser e del guadagno vengono regolati per sfruttare l'intera gamma del rivelatore, ma evitare la saturazione dell'immagine. Pacchetti software microscopia commerciali in genere forniscono le tabelle di ricerca che visualizzano il limite inferiore e superiore della gamma dinamica. Inoltre, per ridurre al minimo colorante sbiancamento impiegare la più bassa possibile lpotere Aser che offre ancora sufficienti dettagli strutturali di membrana TATS. Inoltre, il potere di eccitazione dovrebbe essere sufficientemente basso per evitare danni foto cumulativo portando a contratture miociti e la morte. - Pixel Size
Utilizzare una dimensione del pixel compatibile con il campionamento di Nyquist, circa la metà della risoluzione ottenuta con le impostazioni date. Per l'imaging confocale una dimensione di pixel di 100 nm x 100 nm è compatibile, che si limita anche lo sbiancamento. Per la microscopia SuperResolution dimensioni dei pixel significativamente più piccole sono usate per esempio, 20 nm x 20 nm per la microscopia STED 9. - Tempo di sosta
Microscopi confocale forniscono una funzione di media. In generale, utilizzare il minor tempo di permanenza dei pixel possibile evitare sbiancamento in combinazione con il valore medio es, linea media ≥ 8 per migliorare il rapporto segnale-rumore rapporto. - Documentare le impostazioni di Microscopia applicata attraverso meta-dati
Una volta che le impostazioni comedettagli delle immagini di strutture a membrana tatuaggi sono stati ottimizzati su un particolare microscopio confocale, cassetta di sicurezza e / o documentare le impostazioni del protocollo (meta-dati). Acquisire tutte le immagini all'interno di un gruppo (o tra) (s) di cellule con lo stesso obiettivo, il potere di eccitazione, il guadagno, la dimensione dei pixel, pixel tempo di sosta, e la funzione media. Condizioni di imaging Pari permettono un confronto diretto e quantificazione all'interno di un gruppo (o tra) (s) delle cellule. - Per una guida generale e ulteriori dettagli circa i principi e le applicazioni della microscopia confocale fare riferimento al manuale del biologico Microscopia confocale (Pawley JB, 3 ° edizione, 2006, Springer Science + Business Media, LLC).
In contrasto con le strategie di analisi diretti qui presentati, precedenti pubblicazioni che descrivono le membrane tatuaggi e correlati alla malattia, i cambiamenti hanno utilizzato letture aggregati regionali di densità T-tubulo come strategia quantitativa 16,17, o strategie regionali indirette sulla base di Fourier trasfanalisi ormazione dei segnali di membrana striati al fine di valutare T-tubulo componente regolarità 7. Al contrario, gli approcci quantitativi qui descritti sono direttamente correlati ai componenti Tats individuali e fornire una serie di parametri aggiuntivi tra cui membrana proprietà di rete e componenti specifici come la percentuale di A-tubuli. Inoltre, la densità di rete TATS può essere quantificato come la lunghezza normalizzata dell'intero scheletro estratta per area ROI. Il numero delle giunzioni triple di tre singoli, componenti tubulari costantemente collegati può essere usato come misura della complessità ramificazione della rete membrana TATS. Notiamo che qualsiasi analisi di componenti più piccoli tatuaggi dipende dalle procedure di colorazione. Nella nostra esperienza, 800 ml di una soluzione di-8-ANEPPS 50 micron sono sufficienti a macchiare le reti Tats completi in un pellet di cellule contenente 50.000 cellule VM 9. Tuttavia, se il pellet cellulare contiene un minor numero di miociti cardiaci, Se i rivelatori di fluorescenza potenti sono disponibili, e se l'imaging confocale della distribuzione complessiva rete TATS piuttosto che piccoli dettagli di membrana e le modifiche quantitative sono di interesse, concentrazioni più basse di tintura può essere utilizzato sulla base di prove empiriche. Infine, una macro software scritto per l'analisi descritto può essere utilizzato per automatizzare le fasi di elaborazione dell'immagine per facilitare l'analisi di insiemi di dati di grandi dimensioni, che è particolarmente utile per il confronto tra i diversi gruppi di trattamento (ad esempio, farmaci), tipi di cellule (ad esempio, AM contro VM ), e gli interventi fisiopatologici (ad esempio, farsa contro infarto del miocardio).
Per l'analisi delle immagini delle reti di tatuaggi, la seguente sequenza di passi principio si applica: 1) palla background-sottrazione (4.5.1) per rimuovere variazioni spaziali di intensità di sfondo; 2) miglioramento del contrasto locale (4.5.2).; 3) smoothing immagine (4.5.3); 4) regione statistica fusione (4.5.4); 5) che definisce laSoglia di binarizzazione dell'immagine (4.5.6); e 6) calcolo dei dati scheletro (4.5.8). Un passo fondamentale durante la scheletrizzazione di immagini tatuaggi fluorescenti è l'binarizzazione immagine mostrato nella Figura 6. I passi thresholding associati infine definire quali strutture a membrana veri vengono rilevati a rappresentare i componenti tatuaggi sottostanti rispetto al potenzialmente falsi strutture individuate per errore dal rumore di fondo. Identificazione della soglia corretta per l'analisi delle immagini binaria deve corrispondere con i veri TATS strutture di membrana, che dipende da una (SNR) rapporto sufficientemente elevato segnale-rumore ogni approcci microscopia confocale e SuperResolution. Pertanto, una qualità dell'immagine sufficiente dovrebbe essere stabilito prima e successivamente combinate con giudizio critico di qualità cella individuale, compresa la documentazione per immagini in campo chiaro come indicato. Opzioni alternative per adattare il protocollo di segmentazione delle immagini per una data di uscita e / o grandezza fisica dati microscopiodomande iological includono deconvoluzione delle immagini e altre procedure thresholding come "Otsu" o "Iso-dati", disponibile come plugin di ImageJ. Indipendentemente dalla procedura di segmentazione finale, consideriamo il confronto tra i dati estratti grezzi e da sovrapposizione immagine un passaggio obbligatorio controllo di qualità. In sintesi, l'integrità morfologica e la membrana dei singoli miociti isolati, sufficiente colorazione delle membrane intracellulari tatuaggi, ottimizzazione dei parametri per l'imaging di fluorescenza, e il controllo di sovrapposizione dei dati scheletro estratti saranno tutti contribuire alla qualità delle immagini tatuaggi fluorescenti e risultati quantitativi.
Se si utilizzano le specie più grandi rispetto del mouse per l'isolamento delle cellule, i protocolli possono essere facilmente adattate a seconda dei casi. Per i prossimi specie più grandi, cuori di ratto possono essere cannulati con un ottuso 14 G cannula (diametro esterno 2,1 millimetri) e perfusi a 8 ml / min. Significativamente cuori più anziani o malati possono richiedere dimensioni cannula ancora più grandi. In general, perfusione cardiaca può essere effettuata sia dalla pressione costante ad esempio, utilizzando una colonna di acqua alta 1 m tra serbatoio e aorta o dal flusso costante utilizzando una pompa peristaltica. Per l'isolamento di cellule da piccoli cuori roditori come topi e ratti flusso costante può essere vantaggiosa in quanto collagenasi digestione alla fine distruggere vasi di resistenza coronarici che portano a tassi di perfusione eccessivi di perdite letti navi che saranno controllate in una certa misura dai protocolli di flusso costante. Al contrario, la perfusione pressione costante è vantaggioso se il monitoraggio della portata e corretta incannulazione sono una priorità, che è vantaggioso per i modelli di intervento con comportamenti resistenza vaso sanguigno alterato nonché per la formazione delle procedure di isolamento cellulare.
Come indicato sopra, la qualità delle cellule sufficiente è molto importante per studi quantitativi di sistemi a membrana endogeni. Tuttavia, durante la perfusione cardiaca e collagenasi digestione numerosi fattori possono critically influenzano la qualità del isolamento delle cellule, che non dovrebbe mai essere sottovalutato durante l'ottimizzazione di protocollo o di risoluzione dei problemi 27. In particolare, l'attività di un dato lotto di collagenasi deve essere determinato per il tessuto specifico di interesse per esempio, atri o ventricoli prima dell'esecuzione degli studi in buona fede sperimentali per stabilire le condizioni di isolamento deve essere mantenuta per tutto il resto dello studio. Inoltre, la qualità dell'acqua, il pH, la temperatura, l'ottimizzazione e la pulizia del setup perfusione sarà ridurre al minimo il rischio di danni accidentali da contaminanti e emboli, e fattori potenzialmente complementari devono essere monitorati per stabilire condizioni omeostatiche ottimali durante l'isolamento delle cellule. BDM (2,3-butanedione-monoxime), un inibitore reversibile della miosina ATPasi trasversali dei ponti è comunemente usato durante la dissezione dei tessuti e la digestione per sostenere il rilassamento del muscolo cardiaco, che aumenta il rendimento di isolamenti cellulari. Tuttavia, gli investigatori hanno bisogno di to essere consapevoli che BDM può esercitare attività di fosfatasi non specifici che portano a effetti fuori bersaglio ad esempio, l'inibizione della Na + / Ca 2 + correnti di scambio in determinate condizioni 33. Per alcuni esperimenti potrebbe essere vantaggioso sostituire BDM da blebbistatin come soluzione cardioplegic, un inibitore con alta affinità per miosina a concentrazioni micromolari che è, tuttavia, tossici e relativamente costoso e può avere altri effetti off-target. Riposo cardiomiociti sani non devono mostrare alcun contrazioni in assenza di stimolazione elettrica e tali cellule dovrebbero essere esclusi da ulteriori analisi. D'altra parte, la contrazione dei miociti cardiaci e relax in risposta alla stimolazione elettrica a fisiologici extracellulare di Ca 2 + concentrazioni possono essere utilizzati per stabilire il normale comportamento contrattile come misura supplementare per valutare la qualità delle cellule funzionali e / o comportamento anomalo nelle malattie cardiache rispetto ai controlli sani cellule.
In sintesi, i protocolli per l'isolamento di singole cellule e l'analisi quantitativa delle immagini qui descritto è stato applicato con successo per la microscopia confocale e SuperResolution della rete membrana TATS in VM 9 e 21 AM cellule così come per l'analisi quantitativa di reti microtubuli in miociti cardiaci fissi (dati non mostrati). Queste e future applicazioni dei protocolli possono aprire strade per una serie di questioni sperimentali come la caratterizzazione di membrane tatuaggi a diversi stadi di sviluppo o l'analisi delle proteine di membrana associata o organelli strutture che contattano la rete TATS di esercitare altamente localizzata, segnalazione dominio specifico funzioni nelle cellule AM e VM.
Divulgazioni
The authors declare that no conflict of interest exists.
Riconoscimenti
This work received support through Deutsche Forschungsgemeinschaft SFB 1002 (subprojects A05 and B05 to S.E.L.) and KFO 155 (subproject 4 to S.E.L.), a Halbach Foundation award to S.E.L. supporting E.W.; a grant from the German Cardiac Society to S.B.; and a DAAD exchange program supporting T.K. as visitor at the University of Maryland. The research leading to these results has received funding from the European Community’s Seventh Framework Program FP7/2007-2013 under grant agreement No. HEALTH-F2-2009-241526, EUTrigTreat (to S.E.L.). S.E.L. is a principal investigator of the German Center of Cardiovascular Research (DZHK).
Materiali
| Name | Company | Catalog Number | Comments |
| Chemicals and Enzymes | |||
| 2,3-Butanedione monoxime | Sigma-Aldrich, Munich, Germany | B0753 | |
| Bovine calf serum | Thermo Scientific, Schwerte, Germany | SH30073 | Triple 0.1 µm sterile filtered. |
| CaCl2 | Sigma-Aldrich, Munich, Germany | 21115 | Diluted 1:10 in MQ water to obtain 100 mM CaCl2 stock concentration. |
| Collagenase type II | Worthington via Cell Systems, Troisdorf, Germany | on request | Enzymatic activity depends on individual collagenase batches. Collagenase II and other enzyme activities (Caseinase, Clostripain, Tryptic) can be assessed in the "collagenase lot selection tool". Determine cell yield and quality individually for each new lot of collagenase. |
| Glucose | Carl Roth, Karlsruhe, Germany | HN06.1 | |
| Heparin | Rotexmedica, Trittau, Germany | PZN-03862340 | Diluted in 0.9% NaCl and injected subcutaneuosly in abdominal skin. |
| HEPES | Carl Roth, Karlsruhe, Germany | 9105.4 | |
| Forene 100% (V/V) | Abbott, Libertyville, IL, USA | B506 | Active agent: isoflurane, 250 ml. Use approximately 2 Vol% in air/oxygen dispenser instrument. |
| KCl | Carl Roth, Karlsruhe, Germany | 6781.3 | |
| KH2PO4 | Carl Roth, Karlsruhe, Germany | 3904.2 | |
| Laminin (2 mg/ml) | BD Biosciences, Heidelberg, Germany | 354232 | Lamination is described under step 2.1. |
| MgCl2·6H2O | Carl Roth, Karlsruhe, Germany | 2189.2 | |
| MgSO4·7H2O | Carl Roth, Karlsruhe, Germany | 8283.2 | |
| Na2HPO4·2H2O | Carl Roth, Karlsruhe, Germany | 4984.2 | |
| NaHCO3 | Carl Roth, Karlsruhe, Germany | HN01.1 | |
| Taurin | Carl Roth, Karlsruhe, Germany | 4721.2 | |
| Dyes | |||
| Di-8-ANEPPS | Molecular Probes, Life Technologies, Darmstadt, Germany | D-3167 | Stock solution 2 mM in DMSO |
| Trypan blue | Sigma-Aldrich, Munich, Germany | T8154 | Trypan blue is gently mixed 1:1 via tip-cut 1 ml plastic pipette with cell suspension prior to cell counting in Neubauer cytometer. |
| Langendorff Perfusion Setup | |||
| Circulation thermostat | Lauda, Lauda-Königshofen, Germany | Please refer to Louch et al. (JMCC 2011). Heat up thermostat und buffers in perfusion tubing to 37 °C 15 min prior to use. | |
| Flexible silicone tubing Tygon for peristaltic pump | VWR, Darmstadt, Germany | 224-2252 | Tubing needs to be changed regularly. |
| Flexible silicone tubing Tygon for thermostat | VWR, Darmstadt, Germany | 228-4340 | |
| Heating coil surroundung perfusion tubing | Rettberg, Göttingen, Germany | custom-made | Heating coil and tubing needs to be cleaned thoroughly via MQ water after using. Do not use detergents. Glass components should be bathed regularly in 10 mM NaOH overnight. |
| Peristaltic pump | Ismatec, Wertheim, Germany | ISM830 | |
| Three way stop cock Discofix C Luer Lock 10 cm | Braun, Melsungen, Switzerland | 16500C | |
| Three way stop cock Discofix 3SC | Braun, Melsungen, Switzerland | 4095146 | |
| Instruments | |||
| 42 mm glass coverslips | Menzel Gläser via Thermo Scientific, Schwerte, Germany | on request | 0.13-0.16 mm thickness |
| Cannula 21 G | Becton, Dickinson and Company, Franklin Lake, NY, USA | 304432 | Cut to a length of ~5 mm, roughened with sandpaper. |
| Coverslips for Neubauer cytometer 24 x 24 mm | Menzel Gläser via Thermo Scientific, Schwerte, Germany | on request | 0.38-0.42 mm thickness |
| Graefe forceps, 0.5 mm tips, slight curve | Fine Science Tools, Heidelberg, Germany | 11151-10 | |
| LSM 710 NLO | Carl Zeiss, Jena, Germany | 63X 1.4 NA oil objective | |
| Neubauer improved cytometer | Labor Optik, Friedrichsdorf, Germany | 1100000 | Counting procedure: Wipe cytometer and coverslip provided with the counting chamber with 70 % ethanol. Press coverslip gently on the counting chamber so that the two glass surfaces are in contact and Newton's rings can be observed. Subsequently, 10-20 µl cell suspension can be applied to the edge of the coverslip to be sucked into the void by capillary action. Count the intact vs. defect myocytes using the squares of the cytometer grid which reflects 100 nl. Repeat counting procedure on the second grid provided on the cytometer. Calculate the density of cells in your original cell suspension by taking account of any dilutions and counting shortcuts. |
| POC-R2 Imaging Chamber | Pecon, Erbach, Germany | Cell suspension volume: 800 µl; desired plating density: ~1,000 AM and ~10,000 VM | |
| Spring scissors, 8 mm blades straight, blunt | Fine Science Tools, Heidelberg, Germany | 15025-10 | |
| Student dumont #7 forceps, inox | Fine Science Tools, Heidelberg, Germany | 91197-00 | |
| Student iris scissors, curved, 11.5 cm | Fine Science Tools, Heidelberg, Germany | 91461-11 | |
| Student iris scissors, straight, 11.5 cm | Fine Science Tools, Heidelberg, Germany | 91460-11 | |
| Student surcigal scissors, straight, sharp, 12 cm | Fine Science Tools, Heidelberg, Germany | 91402-12 | |
| Tissue forceps, 1 x 2 teeth, slim, 10 cm | Fine Science Tools, Heidelberg, Germany | 11023-10 | |
Riferimenti
- Prosser, B. L., Ward, C. W., Lederer, W. J. Subcellular Ca2+ signaling in the heart the role of ryanodine receptor sensitivity. J Gen Physiol. 136 (2), 135-142 (2010).
- Wehrens, X. H., Lehnart, S. E., Marks, A. R. Intracellular calcium release and cardiac disease. Annu Rev Physiol. 67, 69-98 (2005).
- Cheng, H., Cannell, M. B., Lederer, W. J. Propagation of excitation-contraction coupling into ventricular myocytes. Pflugers Arch. 428 (3-4), 415-417 (1994).
- Williams, G. S., Chikando, A. C., Tuan, H. T., Sobie, E. A., Lederer, W. J., Jafri, M. S. Dynamics of calcium sparks and calcium leak in the heart. Biophys J. 101 (6), 1287-1296 (2011).
- Sperelakis, N., Rubio, R. orderly lattice of axial tubules which interconnect adjacent transverse tubules in guinea-pig ventricular myocardium. J Mol Cell Cardiol. 2 (3), 211-220 (1971).
- Soeller, C., Cannell, M. B. of the transverse tubular system in living cardiac rat myocytes by 2-photon microscopy and digital image-processing techniques. Circ Res. 84 (3), 266-275 (1999).
- Song, L. S., et al. et al. ryanodine receptors in the failing heart. Proc Natl Acad Sci USA. 103 (11), 4305-4310 (2006).
- Oort, R. J., et al. Disrupted junctional membrane complexes and hyperactive ryanodine receptors after acute junctophilin knockdown in mice. Circulation. 123 (9), 979-988 (2011).
- Wagner, E., et al. Stimulated emission depletion live-cell super-resolution imaging shows proliferative remodeling of T-tubule membrane structures after myocardial infarction. Circ Res. 111 (4), 402-414 (2012).
- Asghari, P., Schulson, M., Scriven, D. R., Martens, G., Moore, E. D. Axial tubules of rat ventricular myocytes form multiple junctions with the sarcoplasmic reticulum. Biophys J. 96 (11), 4651-4660 (2009).
- Lukyanenko, V., Ziman, A., Lukyanenko, A., Salnikov, V., Lederer, W. J. Functional groups of ryanodine receptors in rat ventricular cells. J Physiol. 583 (Pt 1), 251-269 (2007).
- Shacklock, P. S., Wier, W. G., Balke, C. W. Local Ca2+ transients (Ca2+ sparks) originate at transverse tubules in rat heart cells. J Physiol. 487 (Pt 3), 601-608 (1995).
- Reynolds, J. O., et al. Junctophilin-2 is necessary for T-tubule maturation during mouse heart development. Cardiovasc Res. 100 (1), 44-53 (2013).
- Di Maio, A., Karko, K., Snopko, R. M., Mejia-Alvarez, R., Franzini-Armstrong, C. T-tubule formation in cardiac myocytes two possible mechanisms. J Muscle Res Cell Motil. 28 (4-5), 231-241 (2007).
- He, J., et al. Reduction in density of transverse tubules and L-type Ca(2+) channels in canine tachycardia-induced heart failure. Cardiovasc Res. 49 (2), 298-307 (2001).
- Heinzel, F. R., et al. Remodeling of T-tubules and reduced synchrony of Ca2+ release in myocytes from chronically ischemic myocardium. Circ Res. 102 (3), 338-346 (2008).
- Lyon, A. R., et al. Loss of T-tubules and other changes to surface topography in ventricular myocytes from failing human and rat heart. Proc Natl Acad Sci USA. 106 (16), 6854-6859 (2009).
- Crossman, D. J., Ruygrok, P. N., Soeller, C., Cannell, M. B. Changes in the organization of excitation-contraction coupling structures in failing human heart. PLoS One. 6 (3), e17901(2011).
- Kemi, O. J., et al. The effect of exercise training on transverse tubules in normal, remodeled, and reverse remodeled hearts. J Cell Physiol. 226 (9), 2235-2243 (2011).
- Sachse, F. B., et al. Subcellular structures and function of myocytes impaired during heart failure are restored by cardiac resynchronization therapy. Circ Res. 110 (4), 588-597 (2012).
- Arakel, E. C., et al. Tuning the electrical properties of the heart by differential trafficking of KATP ion channel complexes. J Cell Sciences. 127 (Pt 9), 2106-2119 (2014).
- Richards, M. A., et al. Transverse tubules are a common feature in large mammalian atrial myocytes including human. Am J Physiol Heart Circ Physiol. 301 (5), H1996-H2005 (2011).
- Trafford, A. W., Clarke, J. D., Richards, M. A., Eisner, D. A., Dibb, K. M. Calcium signalling microdomains and the t-tubular system in atrial mycoytes potential roles in cardiac disease and arrhythmias. Cardiovasc Res. 98 (2), 192-203 (2013).
- Greiser, M., Schotten, U. Dynamic remodeling of intracellular Ca2+ signaling during atrial fibrillation. J Mol Cell Cardiol. 58, 134-142 (2013).
- Voigt, N., Zhou, X. B., Dobrev, D. Isolation of human atrial myocytes for simultaneous measurements of Ca2+ transients and membrane currents. J Vis Exp. (77), 10-3791 (2013).
- Kaestner, L., et al. Isolation and genetic manipulation of adult cardiac myocytes for confocal imaging. J Vis Exp. (31), (2009).
- Louch, W. E., Sheehan, K. A., Wolska, B. M. Methods in cardiomyocyte isolation, culture, and gene transfer. J Mol Cell Cardiol. 51 (3), 288-298 (2011).
- King, N. M., et al. Mouse intact cardiac myocyte mechanics cross-bridge and titin-based stress in unactivated cells. J Gen Physiol. 137 (1), 81-91 (2011).
- Schindelin, J., et al. Fiji, an open-source platform for biological-image analysis. Nat Methods. 9 (7), 676-682 (2012).
- Arganda-Carreras, I., Fernandez-Gonzalez, R., Munoz-Barrutia, A., Ortiz-De-Solorzano, C. 3D reconstruction of histological sections Application to mammary gland tissue. Microsc Res Tech. 73 (11), 1019-1029 (2010).
- Liu, Z. Q. Scale space approach to directional analysis of images. Appl Opt. 30 (11), 1369-1373 (1991).
- Powell, T., Twist, V. W. A rapid technique for the isolation and purification of adult cardiac muscle cells having respiratory control and a tolerance to calcium. Biochem Biophys Res Commun. 72 (1), 327-333 (1976).
- Watanabe, Y., et al. Inhibitory effect of 2,3-butanedione monoxime (BDM) on Na(+)/Ca(2+) exchange current in guinea-pig cardiac ventricular myocytes. Br J Pharmacol. 132 (6), 1317-1325 (2001).
- Kohl, T., Westphal, V., Hell, S. W., Lehnart, S. E. Superresolution microscopy in heart - cardiac nanoscopy. J Mol Cell Cardiol. 58, 13-21 (2013).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati