
Method Article
管式膜网络在心肌细胞的心房和心室分析
摘要
在心肌细胞,管式膜结构形成细胞内的网络。我们描述了优化方案从小鼠心脏肌细胞I)的隔离,包括质量控制,二)活细胞染色国家的最先进的荧光显微镜,及iii)直接图像分析来量化成分的复杂性和膜细胞的可塑性网络。
摘要
In cardiac myocytes a complex network of membrane tubules - the transverse-axial tubule system (TATS) - controls deep intracellular signaling functions. While the outer surface membrane and associated TATS membrane components appear to be continuous, there are substantial differences in lipid and protein content. In ventricular myocytes (VMs), certain TATS components are highly abundant contributing to rectilinear tubule networks and regular branching 3D architectures. It is thought that peripheral TATS components propagate action potentials from the cell surface to thousands of remote intracellular sarcoendoplasmic reticulum (SER) membrane contact domains, thereby activating intracellular Ca2+ release units (CRUs). In contrast to VMs, the organization and functional role of TATS membranes in atrial myocytes (AMs) is significantly different and much less understood. Taken together, quantitative structural characterization of TATS membrane networks in healthy and diseased myocytes is an essential prerequisite towards better understanding of functional plasticity and pathophysiological reorganization. Here, we present a strategic combination of protocols for direct quantitative analysis of TATS membrane networks in living VMs and AMs. For this, we accompany primary cell isolations of mouse VMs and/or AMs with critical quality control steps and direct membrane staining protocols for fluorescence imaging of TATS membranes. Using an optimized workflow for confocal or superresolution TATS image processing, binarized and skeletonized data are generated for quantitative analysis of the TATS network and its components. Unlike previously published indirect regional aggregate image analysis strategies, our protocols enable direct characterization of specific components and derive complex physiological properties of TATS membrane networks in living myocytes with high throughput and open access software tools. In summary, the combined protocol strategy can be readily applied for quantitative TATS network studies during physiological myocyte adaptation or disease changes, comparison of different cardiac or skeletal muscle cell types, phenotyping of transgenic models, and pharmacological or therapeutic interventions.
引言
在健康横纹肌细胞,管状膜结构与"横向"方向(T管)垂直于主细胞轴是丰富的。因此,T管已被定性为肌肉细胞主要的"横向"的表膜(肌膜)持续扩展,深入渗透朝小区中心的细胞质。的T管连续的外表面的膜的生理作用是通过SER细胞器接触结构域在整个较大的心肌细胞体积的电压活化的L-型Ca纳米邻近耦合形成远程胞内区室的快速电耦合离子通道(Cav1.2)内向电流(I Ca)的激活相邻兰尼碱受体(RyR2的)SER的钙离子释放。在交界SER域和T之间的心室肌细胞(虚拟机)中,非连续的膜的接触("结")-tubules被认为控制数千个体细胞内的 Ca 2 +释放纳米畴的每个单元1中。
对于任何给定的接触领域,在并置的膜的每个T形小管和外周(连接)的SER的部分是大约彼此接近15纳米,因此定义为nanodomain。因此,极个别小细胞质子空间隔离这使准小区自治仓行为。当传入的动作电位活化的T管虚拟机,一个相对较小的Ca Cav1.2 离子通道内向电流会迅速增加的子空间中钙离子浓度的钙离子浓度S IN的attoliter大小nanodomain 1。接下来,[ 钙 ] S增加激活钙离子门控的兰尼碱受体(RyR2的)中的并列SER膜纳米路口附近,并且这种耦合过程发生在所有电连接ð心肌细胞纳米畴。 RyR2s发生致密多通道簇的1 Cav1.2通道的估计的化学计量为5-10的RyR2通道2。由于SER到胞浆钙离子浓度梯度很陡(10比4:1)和RyR2s作为高电导钙离子的功能结合型集群的释放渠道,在数量上大RyR2的钙激活导致2 +释放的T小管加上交界的SER域增加局部子空间的[Ca 2 +] S以100微米或更高在1-2毫秒3,4电流。这种心脏信号放大行为也被称为钙离子诱导的Ca 2 +释放(CICR)。总之,T管是必要的膜结构,可以快速激活兴奋收缩(EC)的耦合过程中通过交界nanodomain SER的接触和细胞性CICR 钙释放的信号。
除了T管,轴向细管S(A-小管)平行于主(纵向)细胞轴的显著不同的方向已经证实了通过电子显微镜(EM),共聚焦和双光子显微镜的研究。举例来说,肌原纤维相互连接的带T管附近肌Z-线之间的A-肾小管细胞的宽连续晶格所表现出细胞外示踪剂和固定豚鼠虚拟机5的EM图像。使用外葡聚糖联荧光素染色和生活大鼠虚拟机的双光子成像,复杂的三维网状小管网络中包含可视〜60%T管和〜40%,小管6。这项研究不仅导致了大量的A-小管三维可视化,同时也实现了切片的EM可视化本质上是有限的复杂和动态膜网络,如横向轴小管系统(TATS)的分析。因此,TATS膜的共聚焦活细胞成像直接沾上二8 ANNEPS开发。如果活细胞TATS网络由傅里叶变换分析,经常出现在靠近肌Z-线空间的T-细管部件是通过从条纹信号7的区域中的集合功率谱反射。这种间接的分析策略已经被用于检测在疾病模型7中TATS成分规律性小区宽的区域的变化。例如,shRNA的介导junctophilin-2击倒导致心脏衰竭和亚型特异性蛋白质不足造成的T小管重组与nanodomain 钙释放功能障碍8。我们通过直接定量方法,并进一步利用受激发射损耗(STED)纳米显微镜9在小鼠单独的虚拟机TATS组件的活细胞超高分辨率显微镜最近延长TATS膜网的分析。允许直接分析较小的单个TATS组件,其中接近50:50的横向分布的纳米图像分辨率与小管的轴向方向,定量确定2丰富的差异还面向个人TATS在健康小鼠心脏9成分。这些策略将在下面的协议部分中进一步概述的。
而丰富的A-小管组成部分,在成人心脏的生理作用仍然神秘莫测,新兴市场的研究已经证明了A-小管提示内源性钙豚鼠和大鼠的VM 5,10释放纳米畴相关的SER膜结构。 Cav1.2和RyR2的共聚焦分析发现,高度共存的A-小管路口10。由于自发钙火花在大鼠虚拟机起源比较远的地方,从T管,通常会出现Z线条纹〜20%,有一个参数一直是A-小管相关的纳米畴可能确实存在,并作为钙释放位点11,12。有趣的是,T-细管形成和成熟OC只有出生后小人和P5平行心脏细胞, 例如生长,通过对前体肌膜内陷发芽和未成熟的支TATS网络组件在P10小鼠13。看来,Junctophilin-2是产后TATS网络成熟特别重要,因为的shRNA敲除抑制T型小管膜锚定到SER路口,导致延迟钙释放及病理TATS组织,在不成熟的A-小管为主的架构一致虚拟机13。这些意见最终可能导致验证性的概念,T管通过膜内陷的过程,而A-小管形成可变形通过额外的,甚至替代细胞内的机制14。
在心脏疾病TATS膜特性的变化已成为一个重要的研究领域病理生理问题。在起搏诱发心脏failu狗模型初步报告再次表明了T管和Cav1.2电流(I Ca)的 15亏损。缺血性心肌病的动物模型表现出减少的T小管密度和细胞内钙的减少同步离子释放16。采用自发性高血压大鼠(SHR)模型心脏衰竭中,T管的亏损与Cav1.2和RyR2的的减少nanodomain耦合关联由"RyR2的孤儿"所提出的机制7。的T管的亏损也已缺血,扩张型,肥厚型心肌病样17表明在人类的虚拟机。此外,增加了在A-肾小管报道,在人的扩张型心肌病18的组织切片。心肌梗死后,我们发现TATS重组小鼠虚拟机与T管对比显著降低差分机制中的A-小管组件9的增加。重要的是,改善了当地的膜对比,通过活细胞superre实现解决方案STED显微镜启用了详细的定量成分分析通过直接测量,这表明了A-小管与TATS网络长度的整体增加,复杂的分支9显著增殖。此外,还表明,运动训练可逆转大鼠的T-细管重塑后心肌梗死19和心脏再同步治疗可能导致反向的T管重构犬心房快速起搏诱导的心脏衰竭20。合在一起,无论是在患病的人和动物的VM以及潜在的治疗性干预的研究将可以说从高质量细胞分离程序和详细的定量分析策略受益在协议中所概述和下面的效果的部分。
此外,最近由侧面证明了对ATP敏感性钾通道的TATS膜贩运亚型21,考虑心房m是重要的yocytes(AMS)作为生物鲜明以及比较心肌细胞模型与虚拟机。 T管最近被记录在羊和人的肺泡22。目前的证据表明,存在于细胞上午几个T管,通常像羊和人类,而不是较大的哺乳动物在小型啮齿动物23。相反,虚拟机,在肺泡巨细胞内的 Ca 2 +释放出现从细胞表面传播通过向小区中心其导致标记的空间和时间的 Ca 2 +梯度23扩散发生。在这个框架内就阐明细胞内Ca2 +信号不稳的常见病,多发病形式的机制,如房颤24显得重要。总之,无论是上午和VM细胞的分离和分别用于健康和患病的心是常用的协议。只有当细胞分离做正确作为判断的足够的细胞质量的微观文件,AM和虚拟机的样品应该是CArried向前定量TATS分析。因此,下面的协议部分是极其依赖于高质量细胞从小鼠或其它物种接着活细胞显微术来分析完整TATS膜分离。正如前面所指出的,TATS膜的特性是有倾向的固定和编制文物6时,由于渗透压的改变细胞膜的变化,传统的光学显微镜9的分辨率限制,一个具有挑战性的研究领域。我们注意到,国家的最先进的最新协议,对人类肺泡大鼠虚拟机用于细胞培养的钙成像和膜片钳和隔离此前已发表 在该刊25,26。
研究方案
注:所有动物的程序进行了审查,并符合实验动物的人文关怀和使用经大学医学中心哥廷根的机构动物护理和使用委员会。
1,隔离心房和心室的小鼠心脏的心肌细胞
- 处理动物尽量轻柔,并根据批准的协议,一般尽量减少应激,特别要避免对离体心肌细胞神经内分泌的过度潜在的强大的意外效果。此外,注入每只小鼠用肝素(500 IU / kg体重,SC)至少20分钟前,心脏提取,以防止血液凝固和微栓子的隔离期间可以显著损害心肌细胞的产量和完整性。
- 麻醉年龄在12周或更旧异氟醚吸入小鼠,证实没有痛撤反射,并通过颈椎脱位安乐死的动物。
- 提取迅速按照先前制定的专家协议心脏( 例如 ,见Kaestner 等 26 Louch 等 27)。避免到心房由不必要的挤压或拉伸的任何无意的伤害。
- 用一对镊子残端和直剪刀的建立通过主动脉血管壁,这对于成功的插管和心脏灌注重要的连续横向切削刃小心保持近端升主动脉的组织。
- 马上转移切除心脏入冰冷的名义上的 Ca 2 +免费灌注缓冲液(解决方案见表1)。请转移至空气进入缓冲区时夹紧大血管,以避免意外的空气栓塞,直至心脏被完全淹没。使用BDM在缓冲器和冰浴冷却的溶液,以抑制心脏收缩。
- 使用双筒变倍显微镜具有足够的3D ILLUMination。
- 具有光滑,表面导管插入下心脏的全景立体视觉主动脉抛光21克套管(外径0.81毫米;正常小鼠心脏重量)必须完全充满缓冲区。确保没有气泡套管通过一个2路鲁尔阀提供快速解决方案,流量控制连接近端溶液槽( 如注射器)。
- 下双眼放大该插管的主动脉,这大约为1毫米以上的主动脉瓣和冠状动脉分支内正确定位确认。绝对避免通过任何通道或主动脉瓣与插管不慎穿孔(这会永久破坏主动脉瓣关闭,从而破坏心肌灌注)。
- 轻轻地绑在主动脉定做,圆周方向取向的防滑槽附近使用两种丝线插管的末端。不要冲洗冠状动脉arteri的es强行在任何点。连接解决方案,填补套管绑在主动脉和心脏,以定制和预校准灌注系统的紧密配合流出连接器,又名修改后的Langendorff设置(使用恒压或恒流;另见下文讨论部分)。
- 使用含氧灌注缓冲液中于37℃(4毫升/分钟的目标灌注速度)尽快灌注心脏为4分钟。通过切换灌注胶原酶含有消化缓冲液(600 U / ml的胶原酶II型)为8-10分钟,在37℃下开始消化。监察组织消化的通过确认类似组织变化包括增加不透明性,柔软性,并且在整个表观心脏表面痿的进展。
- 解剖心腔需要如下消化。将空心心脏双目显微镜下可视化后心脏壁。解剖残留非心脏组织如肺中第二容器的部分,以避免细胞污染中使用微型剪刀的消化缓冲液( 例如 ,弹簧剪刀8毫米直叶片), 如图1。
- 跟随该特定腔室,区域,和/或胶原酶消化心脏的细胞应收获查找列表( 图1):左和/或右心房,游离左和/或右心室壁和/或心室中隔膜。
- 对于具体的心脏组织的解剖,用一个比较宽扁清扫浴涂有几个毫米厚的一层有机硅塑料弹性体。修复心脏的心尖用细虫钢针底部弹性体层。
- 偏转右心耳和解剖右心房的正上方的房室瓣。继续与左心房解剖。解剖和丢弃纤维状阀装置。最后,解剖左,右自由心室壁和室间隔和/或小呃组织的部分需要。
- 只对学员请注意:获得实践,先从非消化小鼠心脏。为了便于解剖定位,实行组织处理,包括在双眼视力都连续清扫步骤, 如图1所示。一旦3D解剖,人工搬运下的双眼视觉,和解剖步骤足够熟悉,继续与胶原酶消化小鼠心脏的上文所述。
- 心室肌细胞(VM)的细胞分离:转移心室组织成2.5毫升新鲜的消化缓冲液。如果从多个器官部位, 如心房和心室同步单元隔离尝试,第二人可接管细胞分离过程的一条腿,这样可以尽量减少,并通过多种心脏组织协调处理优化利用小鼠。继续进行步骤1.9.1-1.9.4,此后1.10。
- 解剖或者是整个心室组织或特定的零件( 例如 ,LV,RV,游离壁和/或隔膜)成约1mm 3件在2.5ml消化缓冲液使用锋利的剪刀( 例如 ,弹簧剪刀8毫米直叶片)在60毫米陪替氏培养皿。
- 轻轻地分解成虚拟机细胞悬液通过组织块用移液缓慢研磨。避免空气泡在细胞悬液。
- 加入8毫升停止缓冲到VM的细胞悬浮液,并将细胞悬浮液转移入15ml锥形管中。使剩余的组织碎片沉淀在底部为约15秒,但是对于分离的细胞保持在悬浮液足够短。接下来,通过收获上清体积的转移虚拟机悬挂到一个新的15ml试管中。如果过度的组织片都存在,可选地使用微创200μm的间隔开的尼龙网的组织碎片从细胞悬浮液中分离出来。
- 让VM的细胞悬液沉降到15毫升的合作底部nical管通过重力为8分钟。
- 洗步骤:除去上清液,并轻轻悬浮在剩余的VM沉淀在10ml的灌注缓冲液中。重复洗涤步骤1.9.5(可选:添加额外的洗涤步骤,以逐渐增加的 Ca 2 +的浓度根据需要)。
- 重悬结算VM沉淀在10ml灌注缓冲和分发在1.5ml微量离心管(每管大约50000 VM细胞)中剩余的细胞悬浮液的体积。
- 心房肌细胞(AM)细胞分离:转移消化/解剖心房组织在1ml新鲜的消化缓冲液。
- 采用切开一个小培养皿( 例如 ,直径60毫米),显微剪的部分消化的心房组织成约1mm 3个1毫升消化缓冲液。使用研磨以1毫升的塑料吸管切尖,以免损坏流体喷射轻轻游离细胞上午从消化组织块制成细胞悬液。在研磨严格避免空气泡在细胞悬液。继机械搅拌,加入4毫升挡车器(50微米氯化钙 10%BCS)的逮捕细胞悬液任何剩余的胶原酶活性。
- 转让上午细胞悬液注入15毫升锥形管。使剩余的组织碎片沉淀在底部为约15秒,而对于分离的细胞保持在悬浮液足够短。收获含有通过解决方案转移的自由上午细胞到一个新的15ml试管中的上清液的体积。
- 离心AM细胞悬浮液, 例如 ,在20 XG 2分钟,在室温或-优选为膜的研究-让细胞在15ml锥形管中沉淀下来慢慢由重力20分钟。
- 洗步骤:弃去上清液并轻轻悬浮在AM沉淀在5ml灌注缓冲液中。重复1.10.4。
- 上午悬浮细胞轻轻地在5毫升灌注缓冲。散布在1.5ml微量离心桶的细胞悬浮液体积ES(每管约1000上午细胞)。
- 分析和记录每一个心脏,包括利用台盼蓝染色的细胞产量的分离的细胞群的质量。
- 对于此,稀释500μl的细胞悬浮液为1:1体积/体积用锥虫蓝用1ml切割枪头溶液(终浓度0.02%)。通过非常缓慢的向上/向下吹打混合细胞,台盼蓝平缓。立即应用的含有细胞悬浮液到一个纽鲍尔式细胞仪提高了锥虫蓝和使用倒置显微镜计数的完整细胞。
- 排除任何细胞明显损伤,膜泡,干扰条纹,挛缩,和细胞内累积锥虫蓝( 图2)。还排除自发性收缩细胞,其很容易受到随后的细胞死亡。为了评估完整的细胞悬浮液的数量,只能使用心肌细胞与普通的条纹贯穿其中不包括台盼蓝细胞体积。
- 判断个人的心房和心室肌细胞通过透射光显微镜的完整性。明场图像保存为TIF文件的文档和进一步的分析。
- 使用下列标准的心肌细胞完整性分析:
- 确认的定期条纹整个可见细胞体积的存在;
- 确认在两个小区边的侧表面的膜平行于肌丝的连续完整性;
- 可视化尖锐的锯齿,在对两种细胞两侧反映特定表面的膜结构的完整性的闰盘;和
- 如在第3描述了用于定位相关旁边的底层小区特有亮场图像的形态( 例如 ,用ImageJ的结合两个图像作为叠加化合物IMAG可视TATS膜的荧光信号(或免疫标记小窝蛋白3蛋白或其它膜标记)E)。
- 确定来自亮场图像的肌节的长度。对于平均小节的长度,测量的顺序排列肌条纹的距离,和由肌节的数量划分的距离。测量每个单元的至少两个位置。进行分析的商业软件或离线使用ImageJ的。
注:如果用解偶联药物治疗的,完整的虚拟机轻松从小鼠心脏显示的1.9〜28微米,平均肌节长度。
- 使用下列标准的心肌细胞完整性分析:
- 量化的形态和个体心房和心室肌细胞的尺寸从该透射光显微镜图像。想想看,上午和VM细胞体积显著不同。测量电池的长度,宽度和面积,并计算出长度:宽度比。
- 使用该命令,多边形选择工具中的ImageJ分析从透射光图像的二维细胞的尺寸和增加的投资回报率管理器,类似于图3,如果形态çhanges被具体研究背景中预期,进一步文件中所有单元的具体的小鼠品系,年龄,性别,心脏大小,任何人干预用于随后的数据分类。
2,染色膜TATS生活中的心房和心室肌细胞
- 装入成像室( 如 POC-R2),配有42毫米的玻璃盖玻片。为稳定肌细胞附着到盖玻片,用1:10稀释的层粘连蛋白的库存在生理灌注缓冲液(终浓度为0.2毫克/毫升)制备20微升的层粘连蛋白溶液。差价20微升的玻璃盖玻片的层粘连蛋白均匀溶液。
- 准备800微升的灌注缓冲50μm的二8 ANEPPS解决方案。对于此,稀释20微升2mM的二-8-ANEPPS储备溶液在780微升的生理缓冲液中。
- 染色的虚拟机,让细胞在1.5ml反应管解决了重力8分钟。染色AMS,或者使用重力沉降或SP在细胞悬浮液中,持续2分钟(参考1.10.3)。为AMS和VM时,小心地取出上清液,同时避免将细胞沉淀的不必要的搅拌,轻轻重悬细胞沉淀在800微升二-8-ANEPPS含有溶液(50μM)。紧接在成像室中的二-8-ANEPPS /肌细胞悬浮液转移到层粘连蛋白包被的盖玻片。
- 染色的虚拟机悬挂在室温下15分钟在黑暗中。
- 慢慢地通过向上的液体弯月面在成像室的两侧具有手动移液管除去过量的体积。确认大部分二8 ANEPPS染细胞仍然牢固地附着在层粘连蛋白包被的盖玻片,不要变成暴露在空气中。接着,慢慢加入1 ml的灌注缓冲液,然后除去任何多余的流体包括非粘附细胞洗附着肌细胞悬浮液一次。
- 小心覆盖在染色和表面附着细胞用1ml的灌注缓冲液慢慢从吨的侧他成像室。放置在显微镜工作台上成像室。
3,成像TATS膜结构在生活心房和心室肌细胞
- 一般情况下,慎重选择最佳的荧光显微镜选项(S)供TATS膜的成像。对于共焦成像,可以考虑最近的一代,现代的荧光显微镜与优化的光电倍增管阵列检测器和光子循环通路,最大限度地提高荧光信号强度。对于TATS的小细节聚焦成像膜结构使用63X 1.4 NA油浸物镜或-视情况而定-使用STED超分辨率显微镜最小TATS细节为心肌细胞的具体应用由科尔等[34]对高一般原则审查。分辨率荧光显微镜,参阅讨论部分。
- 置的摄像参数,以检测大多数,最好是全部二-8-ANEPPS染细胞内的纪念品一个给定的心肌细胞成像平面内branes。使用下面的参数作为起点,共聚焦激光扫描显微镜:激发458纳米,例如 ,以最大激光功率的3%;检测550nm和740纳米之间的发射信号;检测器增益( 例如 ,主命令800);和针孔1非盟为900纳米的光切片厚度。调节这些参数,以优化信号 - 噪声。
- 使用明场模式中选择一个完整的AM或VM细胞作为适当的( 图4A和4B)。请参考1.12点相关的标准如何判断细胞的完整性,选择细胞作为总结:在所有四个小区边小区范围内的普通条纹和平等的肌间隔,尖锐的表面和边缘锯齿,侧膜表面的连续完整,没有任何膜泡。
- 以中央心肌细胞内部分的样本图像。要调整投资回报率使用"裁剪"功能→;调整裁剪窗口→最终的像素尺寸测量100纳米×100纳米。调整裁剪窗口的x轴,以与主要的(轴向/纵向)的心肌细胞的轴线相对应。
- 选择最终的成像平面上。使用单个图像帧,以在z方向上手动选择适当的成像平面上。确认TATS膜,包括T-细管和A-小管组件,是在焦平面明显可见。需要注意的是一个典型的细胞内成像平面可以包括核细胞内的参考点。参阅图4A和4B的实施例。
请注意:一般来说,保持细胞暴露于激光尽可能地短。如果可能,使用单个图像帧,以确定在心肌细胞中的最佳聚焦平面。 - 调整的像素停留时间约为0.5微秒。选择16倍的平均和记录图像的快照。重复图像的快照的步骤,以建立相应的成像平面上的根据需要Š概述了TATS膜结构3.5。
- 保存最终的图像,并确认该文件被保存在目标文件夹中。一般情况下,保存所有的图像文件进行分析的软件统一应用相同的格式( 例如 ,LSM)。在此之前的任何图像分析,再次确认足够的细胞的完整性离线(考虑下步1.12.1列出的标准),并从分析中排除任何受损的细胞。参阅图4C和4D中的示例。
在TATS膜网络及其组件的四分析
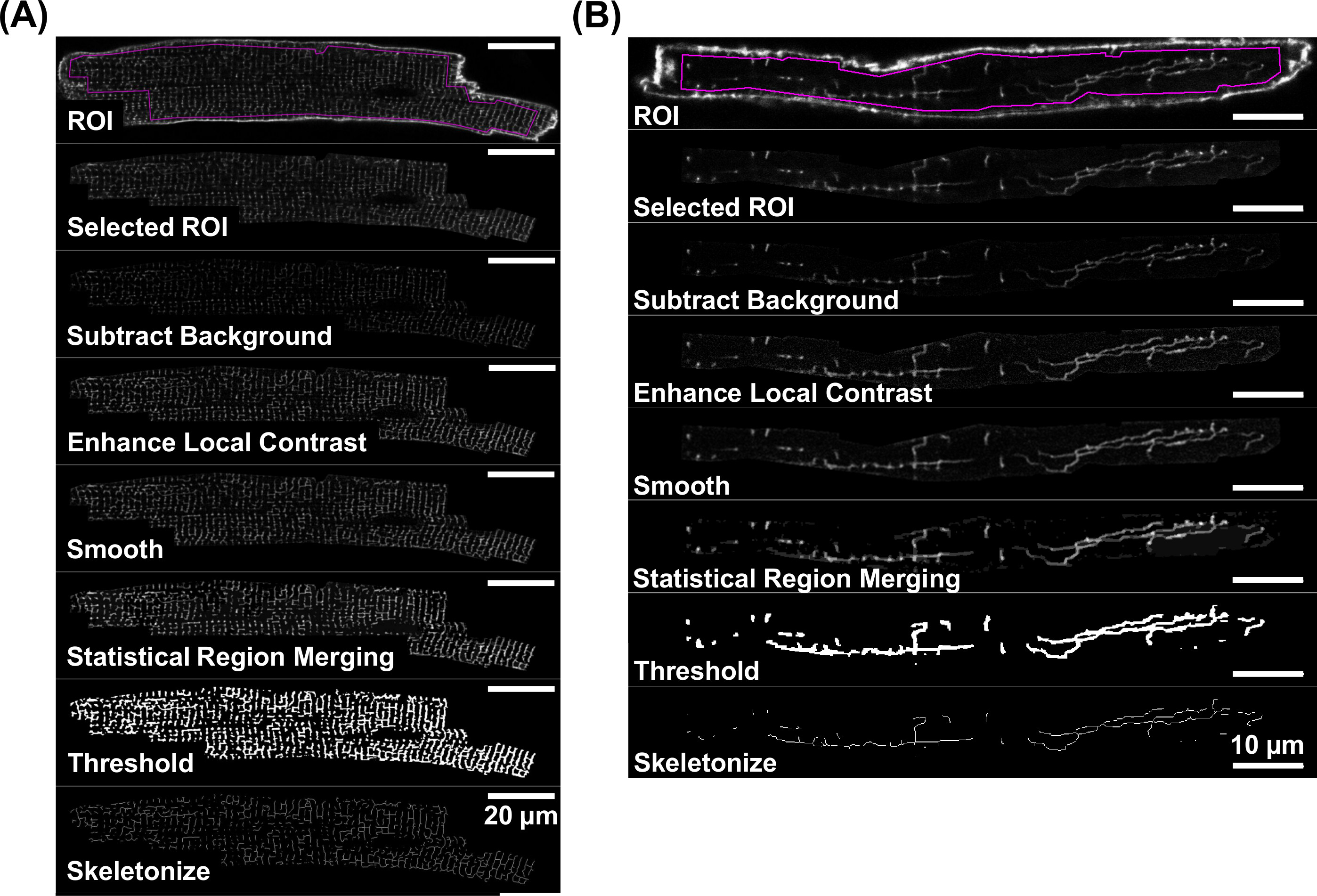
为TATS膜元件直接分析如下图像处理步骤归纳在图5A和5B的自上而下的工作流程图。
- 打开斐济(http://fiji.sc/)二-8-ANEPPS染心肌细胞的图像文件,免费提供的ImageJ的变体,它包含了分析插件必不可少图像处理。欲了解更多信息,请参阅Schindelin 等29。
- 保存图像→文件→另存为→tif格式。
- 分析TATS膜元件,选择适当的投资回报率,不包括外表面的膜信号,然后使用"多边形选择"工具来划定边界的投资回报率不包括外表面的膜(肌膜),并包括TATS膜的胞内部分,如图图5A(投资回报率)。通过应用分析→工具→投资回报率经理所选择的投资回报率添加到"经理的投资回报率"。
- 选择用于TATS组分的取向分析的特定的ROI,对齐的主要纵向细胞轴和所述图像的x轴平行。如果细胞是稍弯曲,选择几个投资回报并分别调整每个投资回报率。排除在分析任何核。从分析中排除任何过度弯曲的细胞因的R精确对准机会性感染将变得越来越困难,在分析过程中增加的方向错误。
- 删除从外表面的膜任何不需要的信号信息:编辑→清除外,产生了"选择的投资回报率"( 图5A)。确保所选择的ROI仅包含细胞内的膜部,它与TATS网络通讯。
- 执行前的后续定量分析作为记录在图5A中的图像处理步骤(命令)下列链。
- 请点击→→工艺减去背景。设置滚动的球半径为5像素。
注:设置滚动的球的半径为5个像素,如果要分析的图像具有100纳米×100nm的像素大小。对于其它的像素尺寸设置滚动的球半径到大约对应于500nm的物理半径的像素的数目。 - 请点击→→工艺增强局部对比度(CLAHE)。将块大小设置到图49中,直方图单元256,最大坡度为3,掩模为"无"。
- 请点击→→过程顺利。
- 点击→插件→分割→统计区域合并。设置参数Q100→点击查看平均值。
- 确认统计区域的完整拼合处理指示的自动演示了新的图像帧,确认标签"SRM Q = 100"出现。继续这个映像文件下面的步骤。点击→图片→类型→8位。
- 点击→图像→调整→阈值。选择门槛足够低,以检测最多,最好所有的TATS结构,在TATS零部件的特定避免排斥与低信号强度(用40阈值作为起点)。请参考"代表性成果"部分,详细的数据输出和示例如图6(255上限阈值)。文档的阈值参数的最终选择。需要注意的是正确的选择阈值应该只产生特定TATS膜结构,但不因背景噪声的假阳性信号。
- 确认的TATS图象细节与提取轮廓数据,特别是对于中间和高的荧光信号水平,这应与骨架结构的(显示)的连续性正确叠加。一旦合适的阈值已经被确定,在分析过程中应用此相同的阈值,所有的图像和重复原始信号的覆盖比较与骨架数据一致地减少这种潜在的偏差来源。
- 点击→应用:图像数据变成二进制下的"门槛", 如图5。
- 点击→插件→骷髅→缩略语(2D / 3D)。保存镂空2D图像通过( 如图5),为TIF文件点击→文件→另存为→tif格式。点击→插件定量数据输出的镂空图像文件进行分析→分析骨架(2D / 3D)。选择修剪周期法:无。确认自动生成所得到的数据表中的。
- 保存自动生成的数据表为txt文件。选择相关的量化参数,例如 ,分支点的总数,或由示例性的数据在图7所示的平均分支长度。考虑进一步的数据分析以像Excel和适当补充/支持的软件工具。
- 请点击→→工艺减去背景。设置滚动的球半径为5像素。
- 请考虑使用自动图像处理程序尽可能包括根据4.5所述,协调单个图像和/或图像批次间分析的所有必要步骤。使用含有编程斐济图像处理的宏(如需要调整规划)的例子补充代码文件。
- 分析中从斐济的镂空图像数据的全部或TATS选择网络组件的灵敏度独立方向插件"方向性"。生成其中相应的轴向定向的A-小管或横向定向的T细管部件由0°或90°仓表示的方向性的直方图。注意,图像方向的正确的参考,并且它以如图8和代表结果与主(纵向)为0°的虚拟机单元的轴紧密地对应为图像的x轴重要。
- 点击→分析→方向性→指定方法:傅立叶分量,Nbins 180,柱状图开始-45→单击显示表。
- 保存新生成并显示结果表包括相关的直方图数据为txt文件。考虑到TXT文件中的数据通过Excel等,并根据需要免费的软件工具进一步分析。
- 要生成的子类数据分组的数据集,例如 ,对于相同的条件(和潜在的其他条件)下进行处理的所有小区,重复分析步骤4.1-4.7为所有相关的图像作为合适的。导入以得出平均值从所有图像为一个合并的Excel文件中的镂空数据参数。此外,从相同的分组数据集中导入所有方向性的直方图数据组合成组合的Excel文件,计算并生成平均方向性的直方图。考虑分组的数据集进行进一步的处理需要分析的个人之间或不同的治疗组更多有趣的TATS网络参数。
结果
此外,向TATS膜网络分析若干像细胞内的 Ca 2 +成像,膜片钳电生理学或药理学剂量反应研究中常用的细胞生物学技术从心房或心室严重地依赖于高品质的初级细胞分离或选择心脏组织的部分,以使成熟的分化,结构和生理完好心肌细胞的表征。因此,隔离和质量评估第1节上午和VM细胞是许多不同的问题,包括这里所描述TATS网络分析,这主要取决于完整的膜和细胞的完整性,最终很有用。
图1提供了图像的逐步手册如何进行心脏组织解剖开始,在小鼠心脏的心房室。随后,心室腔和隔膜都准备了ð需要解剖。可靠地建立调幅和VM隔离以足够的细胞纯度精确选择和制备正确的组织部位是很重要的。以下胶原酶消化它可以是相对难以确定心房和心室组织之间夹层的正确路线,但一旦AM和VM细胞混合不受控制的细胞悬浮液中,这是不可能的扭转混合细胞群。因此,解剖定位,三维组织可视化,足够的经验与消化的组织处理,正确识别的特定组织部位及其解剖线将所有向细胞分离的成功。

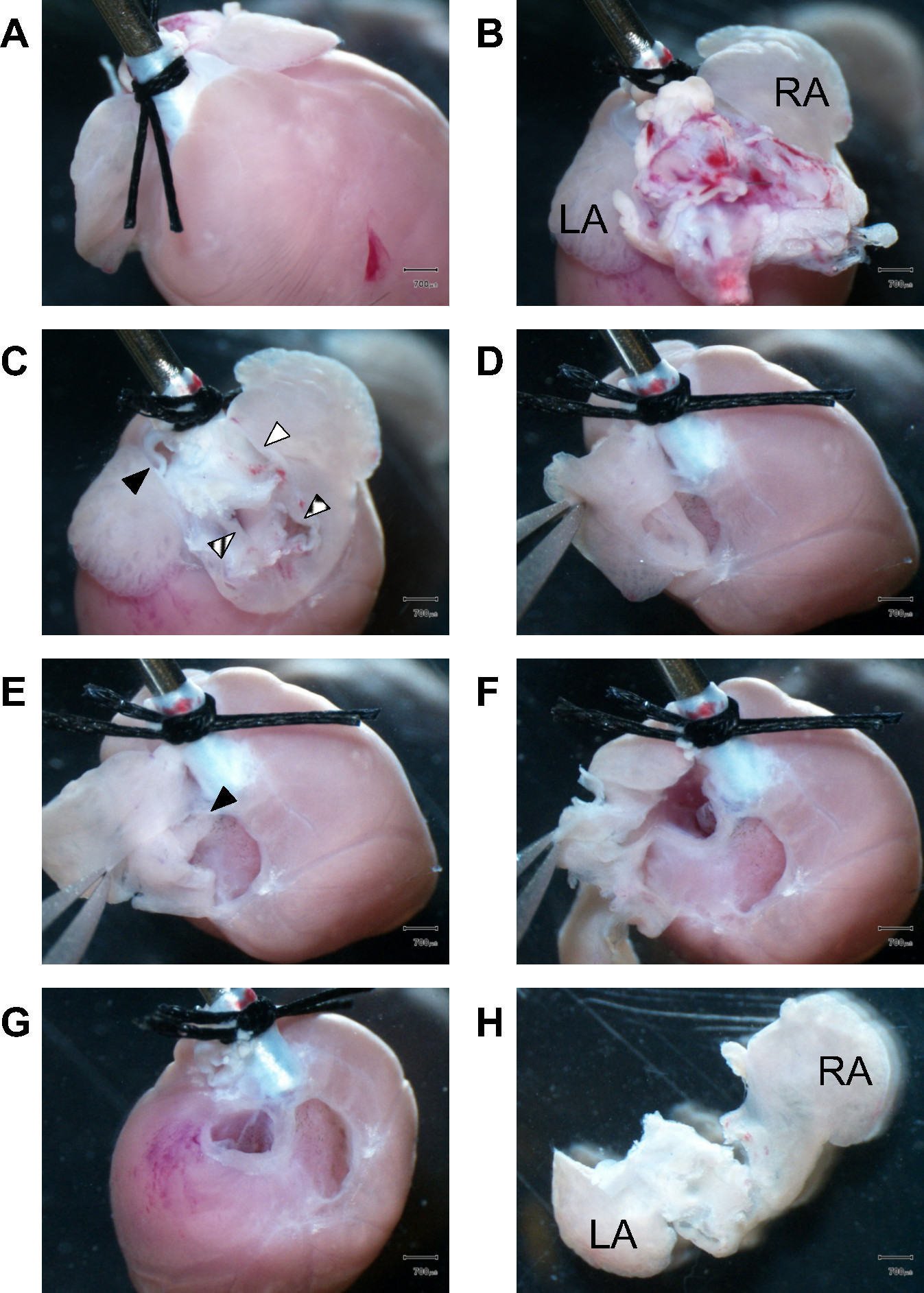
心房组织的图1.Dissection。 (一)面对心脏的前部,两种手术Šutures修复近端主动脉到21克钢套管的残肢末端(B)对剩余肺组织解剖阻碍在心房腔心切的心脏基础影音浏览。洛杉矶,左心房;类风湿性关节炎,右心房(C)剩余lungtissue及大血管被拆除访问心房室。填充黑色三角形,肺动脉;三角纹,肺静脉;填充白色三角形,上腔静脉;盒装三角形,下部腔静脉(D)首先,在右心房壁被切开,而钳握住心耳(E)查看进入右心房室腔。黑色三角形标记完好房间隔(F)的解剖继续进入左心房室腔(G)以下的左,右心房的完全剥离,房室瓣变得可见。纤维气门传动装置被解剖并丢弃到SUBSEQuently收获仅心室肌肉组织。分离的左和右心房(H)的后视图。洛杉矶,左心房;类风湿性关节炎,右atrium.Scale条:700微米。
以确定细胞隔离的质量, 图2提供了典型的细胞的例子的产率和典型的杆或砖状横纹肌完整细胞的存活力的评估过程中,无论是用于AMS和VM。小损坏,在视觉上不明显,以及与异常过度弯曲的形态,或不规则球面状细胞严重受损的细胞,可以容易地确定包括台盼蓝排除在1.11节中描述。虽然完整的细胞保持明亮,均匀条纹,当暴露于外台盼蓝,受损细胞通常显示多个小泡膜和/或迅速积累台盼蓝细胞内表明膜的损伤。不过,台盼蓝本身会伤害过unnecessari细胞LY长潜伏期和直接的细胞质量评估,因此强制性的。对像肌丝挛缩或总表面损伤影响细胞的完整性细胞损伤较为明显的实例是如图4C和4D, 请点击这里查看该图的放大版本。
{kind=link}

如图2台盼蓝排除细胞染色。隔离(A) 的 AM和(B)中的VM的细胞混合在悬浮液用锥虫蓝和通过在40Xmagnification所示的倒置光学显微镜下观察。注意thatabnormal球形细胞(A)和(B)占台盼蓝,这indicatesmembrane leakaGE和结构损坏。与此相反,centralAM和VM细胞与完整的膜排斥台盼蓝,如图所示。此外,请注意,完整的AM和VM细胞显示肌条纹整个细胞体积,nomembrane泡,并在两侧边尖锐的边缘和两闰盘。比例尺为20μm。
下面的VM的成功分离,细胞产量从5×5月 10日至6月 10日可从单个小鼠心脏消化可以预期的。 AMS产率是在3×10 3〜3×10 4个杆形,台盼蓝排除单元的顺序显著更低。相反,虚拟机,甚至有经验的人员隔离偶尔失败。步骤1.11总结程序如何估计分离的健康细胞在悬浮液中的产率。另外,判断为如图3所示的单个AM或VM细胞株通过步骤1.13的平均泡孔尺寸或者比较调幅与VM的细胞群并排侧(如需要)。 请点击这里查看该图的放大版本。
{kind=link}

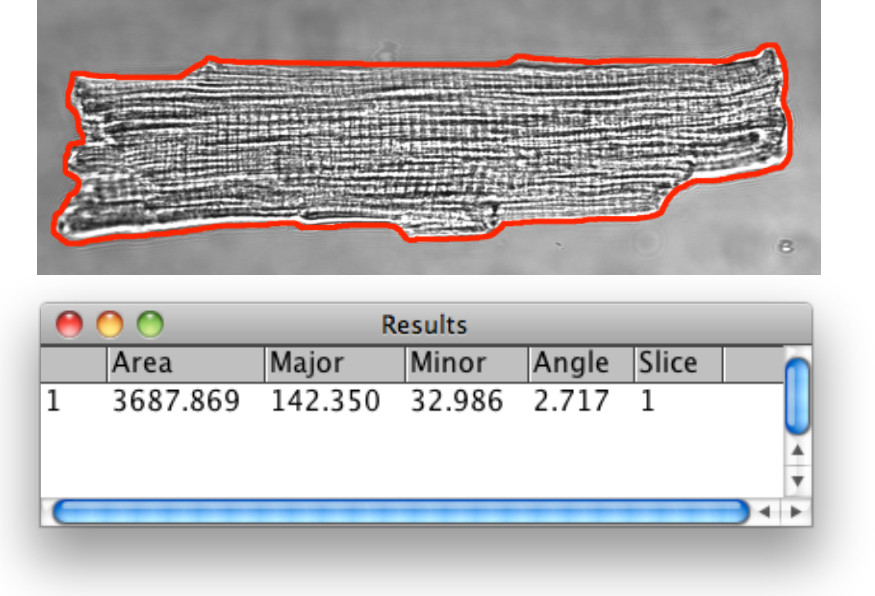
图心肌细胞3.Bright场形态分析,虚拟机的轮廓由下步1.13中描述的图像分析工具检测到。使用多边形选择工具以可视记号和定义由外表面的膜用于分析所定义的细胞的细胞外边界。广告的感兴趣区域(ROI)的投资回报率管理器所选择的区域接着1D距离测量。宽度比调幅与VM尺寸的比较研究,它以记录单元的长度,宽度和面积,并计算出的长度是有用的。
{kind=link}

完整的心房和心室肌的图4.Live膜染色核细胞。对应的透射光及生活的共焦图象二- 8-ANEPPS染色的完整的(A)的 AM和(B)VM细胞。相反,部分收缩,并可能损坏调幅为1.2微米的肌节长度显示在(C)。承包细胞通常显示异常缩短,因而扭曲TATS结构,排除进一步分析。对于细胞膜缺陷另一个重要指标是膜泡(红色三角形)所示在虚拟机中(D)。膜泡代表破坏表面的膜结构和细胞泡应排除进一步TATS分析。此外,虚拟机显示了毛损坏显然缺少其左下部分的整个部分(标以星号)。总之,通过比较透射光及共焦图象,细胞形态和表面完整性是记录和结合荧光信号的信息。 "N"标记NUCLEI从TATS膜染色的分析被删去。黄色柱线表示从上面介绍的相同的共聚焦图像放大投资回报。比例尺:10微米,请点击这里查看该图的放大版本。
{kind=link}
如示于图4A和4B TATS膜具有足够的信号-噪声比的共焦图象接受进一步的定量分析。该TATS膜的分析是基于从荧光直线的信号分量导出镂空的内容图5示出了这是由以下步骤进行详细说明4.3至4.5的个别图像的处理步骤的流程图。这些步骤产生代表直线TATS膜网镂空图像,示出了每个独立的虚拟机( 图5A)和AM的细胞( 连接gure 5B)。

有图5.Workflow的荧光TATS图像骨架。图像处理步骤,这导致TATS网络的骷髅图像由单独的一步一步的图像例子都为二-8-ANEPPS代表的虚拟机染色(A)和AM (B)。对于单个图像处理步骤,请参见第4。注意比例尺的不同:20微米(A)和10微米( 二) 请点击这里查看该图的放大版本。
{kind=link}
图像处理过程中的关键步骤,确定合适的阈值二值化数据如第4.5节所述。6,得到的二进制图像应包括TATS网络唯一真正的膜信号,但不是从背景信号中的噪声阈值的错误虚假的衍生结构。然而,重要的是,该阈值是足够的低,以检测所有真TATS结构使得真TATS组件不会错误地在图像分析丢失。 图6示出的过程中如何选择数据进行二值化时的阈值。而40的阈值, 如图6B和6C似 乎是恰当的,以检测所有真TATS结构单独地,选择较高的阈值,例如 ,60, 如图6A中没有检测到微弱的轴向膜结构(AT)的由黄色三角形所指示。与此相反,选择一个较低的阈值,例如 ,20, 如图6D导致误检测的本底噪声视为假阳性TATS结构由所指示的黄色三角形。

图6.How确定skeletonizing TATS图像数据的过程中的信号的阈值的例子显示每个所产生的数据进行二值化过程中不同的阈值(在步骤4.5中所述)TATS骨架。上图片:显示阈值调整使用斐济。下图片:骷髅图像与相应的输入荧光图像和放大区域的覆盖所示。高门槛,例如 ,60英寸采用(A)是显然不适合检测所有真正的TATS结构,由黄色三角形的放大部分所示。 40中的(B)和(C)应用阈值检测所有TATS结构和正确不检测背景噪音,而低门槛,例如,在20(D)的误识别的背景噪声如TATS结构及由此产生的不存在的膜结构的假阳性信号。假阳性信号由黄色三角形的放大的插图所示。 请点击这里查看该图的放大版本。
{kind=link}
"分析骷髅(2D / 3D)"插件支持镂空TATS结构的详细分析。一旦执行,该插件生成一个数据表具有以下骨架参数:#branches,#junctions,#终点体素,平均枝长,#triple点,#quadruple点,最大分支长度。对于所有可能的输出参数的详细说明,请参考tohttp://fiji.sc/wiki/index.php/AnalyzeSkeleton及相关物品29-31。一个典型的数据表的输出被示于图7A。
参数可以进一步用于导出每个区域的总骨架长度或单位面积结的数目。分支乘以平均分支长度的数目的示例性的计算给出了连续的骨架中的2D的总长度:总长度为骨架每投资回报率:
Σ(#branches点¯x平均枝长)= 5155像素= 515.5微米
骨架的总长度可以是归一化的图像区域。对于图7中的归一化的骨架长度0.64μm的中所示的示例个/μm2和所有路口的总和来计算,如下所示:
归骨架长度:
515.5微米/ 803.6微米2 = 0.64微米/微米2
归号路口:
155路口/ 803.6微米2 = 0.19路口/微米2

图从镂空的图像输出7.Automated数据。 (一)由"分析骨架(2D / 3D)"插件从(B)所示的镂空图像生成一个典型的数据表格。对于可能的输出参数的详细说明请参考http://fiji.sc/wiki/index.php/AnalyzeSkeleton 。 请点击这里查看该图的放大版本。
{kind=link}
在图5中在4.5中描述和图示的图像处理步骤,可以使用作为补充代码文件斐济宏实现自动化。该命令通过反复的步骤定义图像处理的重复。宏可以应用来完成堆叠通过以下步骤4.3和4.4产生的输入图像。宏可以通过制备单独的输入图像栈用于使用宏命令每个独立治疗组用于自动分析是有利的完整的数据集组的分析,例如。
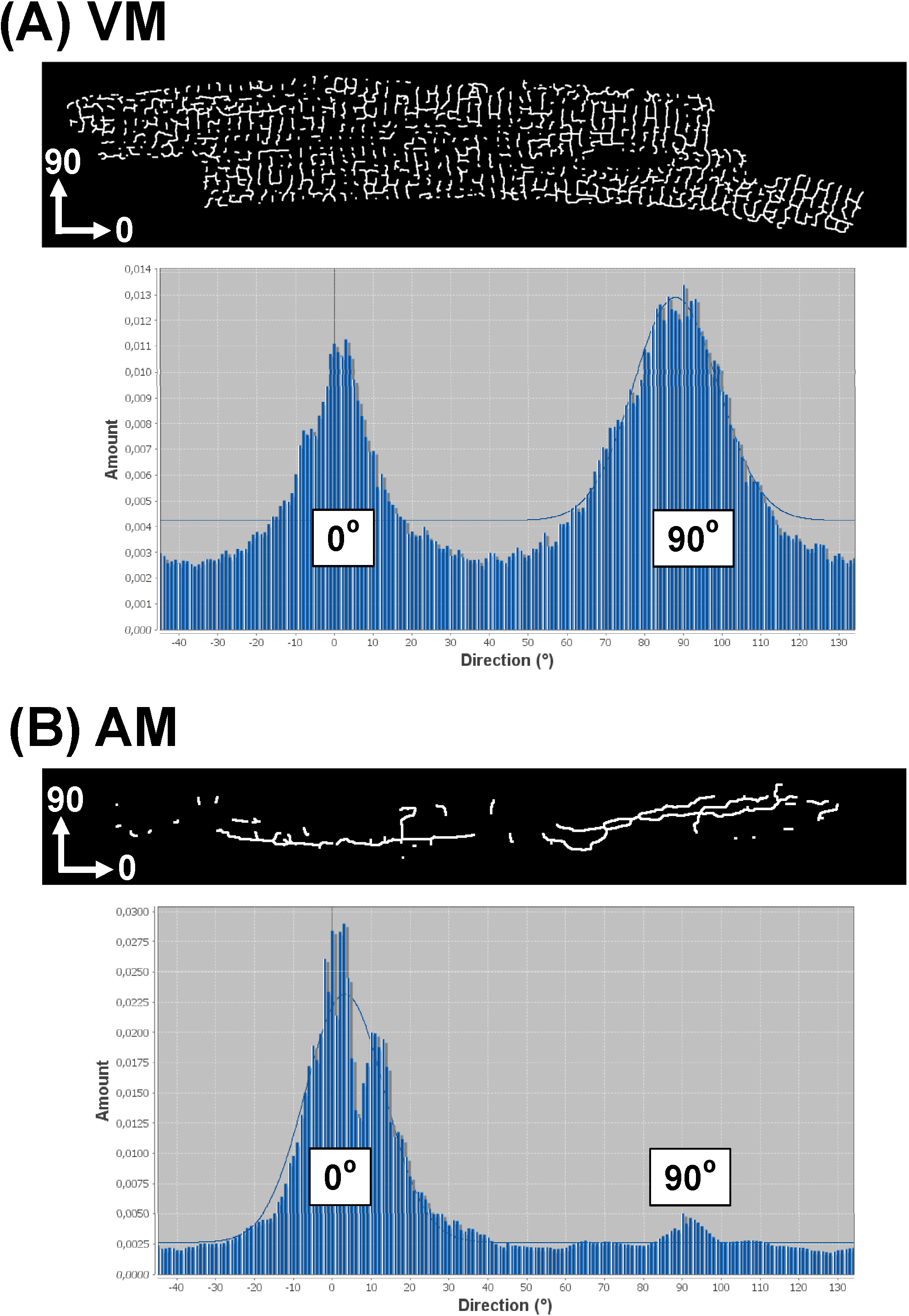
概述的软件战略进一步使TATS网络导向分析的所有组件。为此,使用"方向性"插件(http://fiji.sc/Directionality)29,31产生直方图数据显示所有TATS组件取向的取向分布。如果输入图像的x轴对应于一个给定的AM或虚拟小区的主轴线,轴向(纵向)TATS部件将由0°仓表示,而横向的部件用90来表示#176;宾图8显示了镂空TATS图像的虚拟机(8A)与调幅(8B)电池示范方向性直方图。而典型的VM的方向性的直方图示出了在0°和90°的双峰值分布,则AM直方图显示在0℃显性单峰。这些实施例中与先前观察该虚拟机的单个TATS分量几乎相等的T-管和A-肾小管间分布,而在肺泡的TATS组分可以主要成分的A-肾小管协议。

图8.Representative方向性直方图从单个细胞TATS网络从单个VM(一 )与AM的骷髅图像产生方向性直方图(B )TATS网络。因为所分析的图像的x轴对应于如图所示的心肌细胞的主要(纵)轴,A小管部件由0°仓表示,而T-细管部件用90°的槽来表示。高斯只适合于大型直方图peakasshown作为背景图。 请点击这里查看该图的放大版本。
{kind=link}
| 灌注缓冲 | 毫 |
| 氯化钠 | 120.4 |
| 氯化钾 | 14.7 |
| KH 2 PO 4的 | 0.6 |
| 的Na 2 HPO 4 | 0.6 |
| 硫酸镁 | 1.2 |
| HEPES | 10 |
| 碳酸氢钠 | 4.6 |
| 牛磺酸 | 30 |
| 2,3 - 丁二酮,肟 | 10 |
| 血糖 | 5.5 |
| pH值7.4 | |
| 消化缓冲液 | 毫(如果未指定) |
| 氯化钠 | 120.4 |
| 氯化钾 | 14.7 |
| KH 2 PO 4的 | 0.6 |
| 的Na 2 HPO 4 | 0.6 |
| 硫酸镁 | 1.2 |
| HEPES | 10 |
| 碳酸氢钠 | 4.6 |
| 牛磺酸 | 30 |
| 2,3 - 丁二酮,肟 | 10 |
| 血糖 | 5.5 |
| 胶原酶II型 | 600 U / ml的 |
| pH值7.4 | |
| 挡车器 | 毫(如果未指定) |
| 氯化钠 | 120.4 |
| 氯化钾 | 14.7 |
| KH 2 PO 4的 | 0.6 |
| 的Na 2 HPO 4 | 0.6 |
| 硫酸镁 | 1.2 |
| HEPES | 10 |
| 碳酸氢钠 | 4.6 |
| 牛磺酸 | 30 |
| 2,3 - 丁二酮,肟 | 10 |
| 血糖 | 5.5 |
| 氯化钙 | 0.0125 |
| 小牛血清 | 10% |
| pH值7.4 | |
表1.Buffer SOLUT离子。三种不同的生理缓冲液的细胞分离和成像的内容进行了总结。
讨论
虽然心肌细胞已被分离,并研究了几十年的32,最近一次检讨的结论是一致的高质量的心肌细胞的隔离仍然充满挑战27。这反映了主要的心肌细胞VIS-à-VIS缺乏共同的标准方法,共享的元数据,而透明细胞质量文件的隔离相对复杂的协议。细胞分离的协议通常是由单独的组定制,产生可变结果的细胞株,取决于个体模式设置( 例如 ,物种,年龄,并存心脏疾病),通常调整为特定的实验条件。在定量TATS膜的研究和协议这里提出,质量评估和文档的基本水平的情况下涉及单个细胞的膜结构容易出现代谢和隔离协议相关变动或共聚焦显微镜超高分辨率,无论是在上午或VM。重要的是,即使细胞株高产建议健康完好的细胞,研究人员需要记录和批判仔细判断每个细胞对抗与非特异性损害表面和TATS细胞膜的完整性,由于分离过程对由于不同类型的具体变化形态标准干预措施相比,控制的条件。在心肌细胞的分离的一个重要变量是一个给定的胶原酶大量的特定活性。以选择一组新的胶原酶,胶原酶几个样品的酶活性,应互相通过评估心肌细胞产率和质量,并根据制造商的说明书进行试验。理想情况下,一个新的大量的胶原酶是确定与类似于以前成功地使用大量的胶原酶活性(对可能的酶活性进一步评估是指在材料和方法表中的"胶原酶很多选择工具")。牛逼阿肯一起的TATS膜的可视化定量方法严重依赖于细胞的分离和质量,反之亦然,心肌细胞隔离导致非特异性膜破坏记录在案的TATS显微镜应触发的隔离程序,严格审查和纠正。由于细胞分离质量和TATS膜的可视化和量化有着内在的联系,本文所讨论的协议,涵盖所有主要方面作为一个连续的策略。
一个额外的挑战和心脏研究中,细胞损伤和/或细胞损失的常见问题的发生是由于代谢损害的干预,例如 ,心肌梗死后如图9所示 ,还需要针对潜在的意外损坏,例如判断,下列过程中细胞分离忽视空气栓塞。从患病心脏的心肌细胞的分离可能导致额外的,显著细胞损失和降低细胞的产量。因此,对比例如果细胞的分离和计数是通过标准化的协议一致地施加控制和病态心脏之间分离的完整细胞的总数的ISON可以是有意义的。因此,重要的是通过适当的对照组,这反映了最佳的心肌细胞的分离的质量来判断细胞的完整性。重要的是,单个细胞的质量和健康与患病细胞与活细胞显微镜的分离过程中不慎损坏,可能显著影响TATS膜的网络分析。这里介绍的协议,在细胞的分离和胎膜完整的活细胞显微因此强调生理膜元件的完整性和稳定性。整个工作流被设计为一个连续的策略来实现并保持完好TATS膜元件同时排除损伤的细胞,因为这些将表现出隔离依赖性膜器物等破坏膜小管,卷材胶粘ê泡,并改变TATS网络错误地控制条件和损害进一步定量分析。反之,同样的策略是至关重要的与潜在的干预研究扰乱TATS膜,它主要取决于真正健康与同TATS膜变化真患病细胞之间进行有意义的比较对照。
此外,我们要解决的程序,实现了技术上更有挑战性的隔离上午细胞。尽管进步和改善的协议,它强调的是,这是不平凡重现虚拟机的高品质小区隔离,甚至不太可靠的肺泡是很重要的。这是由于整体产量较低AM的细胞,其中在细胞的分离,即使小的误差或变化可能导致完全的调幅细胞分离失败,而VM的细胞损伤轻微程度可能是在细胞悬浮液中不那么明显,由于相对高的细胞数字相比,PM。由于上午细胞可能成为ç分离后urved,通过几个感兴趣区分析可能是有利的根据步骤4.3所概述。继细胞分离步骤的详细过程,我们提供了一个协议,直接整合膜染色和共聚焦或TATS都为虚拟机和AMS网络的STED超分辨成像。这些协议使通过先前建立的参数定量分析和TATS膜的选择组件的分化。相比于虚拟机,三维组织和肺泡心房TATS网络功能的行为是目前还不了解。
该程序形象TATS膜在活细胞中(步骤3.1至3.7)的开发与商业焦(表材料/设备)和定制的受激发射损耗荧光显微镜9。以优化荧光图像生成和定量分析TATS显微镜设置,以下几点是具有普遍意义:
- 目的
为了解决TATS膜结构的小细节,实证检验的目标提供最高的图像质量,同时注重几个微米深的细胞。某些共焦显微镜可以与水或甘油的目标,以此处使用的63X 1.4 NA油浸物镜有更好的表现,与此相反。目标与100X的放大倍率用于STED超分辨显微术,权衡的视场较小的纳米级分辨率的34。 - 激发和增益
励磁功率和检测器增益的最佳设置取决于显微镜光路,激光表演和样本性质上。理想情况下,激光功率和增益调节以利用全范围的检测器的,但避免图像的饱和度。商业显微镜软件包通常提供了可视化的动态范围的下限和上限的查找表。此外,为了最小化染料漂白采用尽可能低的升ASER电源仍然提供足够的结构TATS膜的详细信息。此外,激励功率应足够低,以避免导致心肌细胞挛缩和死亡的累积光损伤。 - 像素大小
使用像素大小与奈奎斯特采样兼容,大约有一半实现了与给定设置的分辨率。对于共焦成像的100纳米×100nm的像素大小是兼容的,这也将限制漂白。对于超分辨率显微镜显著更小的像素尺寸,例如使用20纳米×20纳米的STED显微镜9。 - 停留时间
共焦显微镜提供了一个平均功能。在一般情况下,用最短的像素停留时间,以避免漂白结合信号平均化,例如 ,线路均≥8,以改善信号噪声比。 - 文档的应用显微设置通过元数据
一旦设置如何TATS膜结构的图像细节进行了优化,在一个特定的激光共聚焦显微镜,安全和/或文档的设置(协议的元数据)。掌握细胞内的(或间)组(S)的所有图像具有相同的目标,激励功率,增益,像素尺寸,像素停留时间和平均功能。平等的成像条件允许的范围内细胞(或间)组(S)直接比较和定量。 - 对于一般的指导和有关原则和共聚焦显微镜的应用程序的详细信息请参阅生物共轭焦显微镜(帕利JB,第3版,2006年,施普林格科学+商业媒体有限责任公司)的手册。
相反,这里介绍的直接分析的策略,描述TATS膜与疾病相关的变化以前的出版物中,使用了T型小管密度区域总读数定量策略16,17,或间接的区域战略基于傅立叶互感器纹状的膜的信号,以评估T-细管部件规律性7本署可能分析。相反,此处所描述的定量方法是直接关系到个人TATS组件,并提供了一些额外的参数,包括膜的网络性能和像的A-肾小管的百分数特定组件。此外,TATS网络密度可被量化为每ROI区域的整个提取骨架的归一化长度。三个单独的三联点数目,连续地连接小管组件可以用来作为TATS膜网络的分支的复杂性的度量。我们注意到,最小TATS组件的任何分析依赖于染色程序。根据我们的经验,800微升的50微米二8 ANEPPS解决方案,足以完成染色TATS网络,包含50000 VM 9细胞的细胞沉淀。但是,如果将细胞沉淀中含有心肌细胞的下数如果强大的荧光检测器是可用的,并且如果根据经验测试可以使用整体TATS网络分布的共焦成像,而不是最小的膜细节和量的变化是感兴趣的,低级的染料浓度。最后,所描述的分析编写的软件的宏可用于自动化图像处理步骤,以促进分析的更大的数据集,这对于不同的治疗组( 例如 ,药物),细胞类型之间的比较是特别有用的( 例如 ,AM与VM ),和病理生理干预( 例如 ,假与心肌梗死)。
为TATS网络的图像分析,主要步骤如下顺序应用:1)滚动的球背景减法(4.5.1),以除去在背景强度的空间变化; 2)局部对比度增强(4.5.2); 3)图像平滑(4.5.3); 4)统计区域合并(4.5.4); 5)定义图像二值化的阈值(4.5.6);和6)计算该骨架数据的(4.5.8)。的荧光TATS图像的骨架化过程中的一个关键步骤是在图6所示的图像的二值化的阈值相关联的步骤,最终确定被检测到表示底层TATS部件与从背景噪声确定错误可能的错误结构,其真正的膜结构。正确的阈值进行二值化图像分析,识别要与真实TATS膜结构,它依赖于足够高的信号 - 噪声(SNR)的各比值为共焦和超高分辨率显微镜方法对应。因此,足够的图像质量,应首先建立,随后结合单个细胞的质量包括亮场图像文件的关键判断,概括。替代方案,以适应图像分割协议对于给定的显微镜数据输出和/或物理iological问题包括图像去卷积和像"大津"或"ISO数据"作为ImageJ的插件,其他的阈值程序。不管最终的分割过程中,我们考虑了图像叠加的强制性质量控制步骤提取原始数据之间的比较。总之,个人的离体心肌细胞,足以染色细胞TATS膜,用于荧光成像参数优化,并提取骨架数据的叠加控制形态和膜的完整性都将有助于荧光TATS图像和定量结果的质量。
如果较大的物种比小鼠用于细胞的分离,该协议可以很容易地修改为适当的。为下一个较大的物种,大鼠心脏可插管的钝地下14套管(外2.1mm直径),并灌注以8毫升/分钟。显著年长或患病的心脏,可能需要更大的套管尺寸。在基因拉尔,心脏灌注可以或者由恒压例如进行,使用储层和主动脉之间或通过使用蠕动泵恒定流动的高1米水柱。用于从小型啮齿动物的心像小鼠和大鼠细胞分离恒定流可能是有利的,因为胶原酶消化最终将破坏冠状动脉阻力血管,导致过度的灌注速度漏出血管床将由恒定流的协议来控制到某种程度。与此相反,恒压灌注是有利的,如果监视的流量和正确的插管是一个优先,这是有利的干预模式具有改变血管阻力的行为,以及用于在细胞分离程序的培训。
如以上概述的,足够的细胞质量为内源性膜系统的定量研究很重要。然而,在心脏灌注和胶原酶消化众多因素CRitically影响细胞的分离,这不应该在协议优化和故障排除27被低估的质量。特别是,在给定的胶原酶大量的活性应为兴趣如心房或在执行前的实验真正研究建立隔离条件心室的特定组织被确定为保持在整个研究的剩余部分。此外,水的质量,pH值,温度,灌注设置的优化和清洗会无意中破坏污染物和栓塞的风险最小化,并有可能附加因素必须被监视,以在细胞分离建立最佳稳态条件。 BDM(2,3 - 丁二酮-肟)肌球蛋白ATP横桥可逆抑制剂的组织解剖和消化过程中是常用的,以维持心脏肌肉松弛,从而增加细胞分离物的产率。不过,调查人员需要吨ö意识到BDM可能施加导致脱靶效应,例如 ,在某些条件下33抑制钠 / 钙交换电流的非特异性磷酸酶的活性。对于一些实验中,可能有利的由II型肌球蛋白的心脏停搏液,抑制剂与肌球蛋白在微摩尔浓度的高亲和性然而这是,中毒性且相对昂贵的更换BDM,并且可以具有其他的脱靶效应。搁健康心肌不应显示在没有电刺激与这些细胞的任何收缩应被排除在进一步分析。另一方面,响应于在生理细胞外的 Ca 2 +浓度的电刺激心肌细胞的收缩和舒张,可用于建立正常的收缩行为作为一种附加措施来评估心脏疾病与健康控制功能单元的质量和/或反常的行为细胞。
总之,对于单个细胞的分离和在此描述的定量图像分析的协议已被成功地应用在TATS膜网络的共焦和超高分辨率显微术中的VM 9和AM单元21以及用于微管网络在定量分析定心肌细胞(数据未示出)。的协议,这些和未来的应用可能会打开渠道,多种实验问题,如TATS膜在不同发育阶段的特征或相关膜蛋白或细胞器结构的联系TATS网络的分析,以发挥高度本地化的,特定的域信号功能在上午和VM细胞。
披露声明
The authors declare that no conflict of interest exists.
致谢
This work received support through Deutsche Forschungsgemeinschaft SFB 1002 (subprojects A05 and B05 to S.E.L.) and KFO 155 (subproject 4 to S.E.L.), a Halbach Foundation award to S.E.L. supporting E.W.; a grant from the German Cardiac Society to S.B.; and a DAAD exchange program supporting T.K. as visitor at the University of Maryland. The research leading to these results has received funding from the European Community’s Seventh Framework Program FP7/2007-2013 under grant agreement No. HEALTH-F2-2009-241526, EUTrigTreat (to S.E.L.). S.E.L. is a principal investigator of the German Center of Cardiovascular Research (DZHK).
材料
| Name | Company | Catalog Number | Comments |
| Chemicals and Enzymes | |||
| 2,3-Butanedione monoxime | Sigma-Aldrich, Munich, Germany | B0753 | |
| Bovine calf serum | Thermo Scientific, Schwerte, Germany | SH30073 | Triple 0.1 µm sterile filtered. |
| CaCl2 | Sigma-Aldrich, Munich, Germany | 21115 | Diluted 1:10 in MQ water to obtain 100 mM CaCl2 stock concentration. |
| Collagenase type II | Worthington via Cell Systems, Troisdorf, Germany | on request | Enzymatic activity depends on individual collagenase batches. Collagenase II and other enzyme activities (Caseinase, Clostripain, Tryptic) can be assessed in the "collagenase lot selection tool". Determine cell yield and quality individually for each new lot of collagenase. |
| Glucose | Carl Roth, Karlsruhe, Germany | HN06.1 | |
| Heparin | Rotexmedica, Trittau, Germany | PZN-03862340 | Diluted in 0.9% NaCl and injected subcutaneuosly in abdominal skin. |
| HEPES | Carl Roth, Karlsruhe, Germany | 9105.4 | |
| Forene 100% (V/V) | Abbott, Libertyville, IL, USA | B506 | Active agent: isoflurane, 250 ml. Use approximately 2 Vol% in air/oxygen dispenser instrument. |
| KCl | Carl Roth, Karlsruhe, Germany | 6781.3 | |
| KH2PO4 | Carl Roth, Karlsruhe, Germany | 3904.2 | |
| Laminin (2 mg/ml) | BD Biosciences, Heidelberg, Germany | 354232 | Lamination is described under step 2.1. |
| MgCl2·6H2O | Carl Roth, Karlsruhe, Germany | 2189.2 | |
| MgSO4·7H2O | Carl Roth, Karlsruhe, Germany | 8283.2 | |
| Na2HPO4·2H2O | Carl Roth, Karlsruhe, Germany | 4984.2 | |
| NaHCO3 | Carl Roth, Karlsruhe, Germany | HN01.1 | |
| Taurin | Carl Roth, Karlsruhe, Germany | 4721.2 | |
| Dyes | |||
| Di-8-ANEPPS | Molecular Probes, Life Technologies, Darmstadt, Germany | D-3167 | Stock solution 2 mM in DMSO |
| Trypan blue | Sigma-Aldrich, Munich, Germany | T8154 | Trypan blue is gently mixed 1:1 via tip-cut 1 ml plastic pipette with cell suspension prior to cell counting in Neubauer cytometer. |
| Langendorff Perfusion Setup | |||
| Circulation thermostat | Lauda, Lauda-Königshofen, Germany | Please refer to Louch et al. (JMCC 2011). Heat up thermostat und buffers in perfusion tubing to 37 °C 15 min prior to use. | |
| Flexible silicone tubing Tygon for peristaltic pump | VWR, Darmstadt, Germany | 224-2252 | Tubing needs to be changed regularly. |
| Flexible silicone tubing Tygon for thermostat | VWR, Darmstadt, Germany | 228-4340 | |
| Heating coil surroundung perfusion tubing | Rettberg, Göttingen, Germany | custom-made | Heating coil and tubing needs to be cleaned thoroughly via MQ water after using. Do not use detergents. Glass components should be bathed regularly in 10 mM NaOH overnight. |
| Peristaltic pump | Ismatec, Wertheim, Germany | ISM830 | |
| Three way stop cock Discofix C Luer Lock 10 cm | Braun, Melsungen, Switzerland | 16500C | |
| Three way stop cock Discofix 3SC | Braun, Melsungen, Switzerland | 4095146 | |
| Instruments | |||
| 42 mm glass coverslips | Menzel Gläser via Thermo Scientific, Schwerte, Germany | on request | 0.13-0.16 mm thickness |
| Cannula 21 G | Becton, Dickinson and Company, Franklin Lake, NY, USA | 304432 | Cut to a length of ~5 mm, roughened with sandpaper. |
| Coverslips for Neubauer cytometer 24 x 24 mm | Menzel Gläser via Thermo Scientific, Schwerte, Germany | on request | 0.38-0.42 mm thickness |
| Graefe forceps, 0.5 mm tips, slight curve | Fine Science Tools, Heidelberg, Germany | 11151-10 | |
| LSM 710 NLO | Carl Zeiss, Jena, Germany | 63X 1.4 NA oil objective | |
| Neubauer improved cytometer | Labor Optik, Friedrichsdorf, Germany | 1100000 | Counting procedure: Wipe cytometer and coverslip provided with the counting chamber with 70 % ethanol. Press coverslip gently on the counting chamber so that the two glass surfaces are in contact and Newton's rings can be observed. Subsequently, 10-20 µl cell suspension can be applied to the edge of the coverslip to be sucked into the void by capillary action. Count the intact vs. defect myocytes using the squares of the cytometer grid which reflects 100 nl. Repeat counting procedure on the second grid provided on the cytometer. Calculate the density of cells in your original cell suspension by taking account of any dilutions and counting shortcuts. |
| POC-R2 Imaging Chamber | Pecon, Erbach, Germany | Cell suspension volume: 800 µl; desired plating density: ~1,000 AM and ~10,000 VM | |
| Spring scissors, 8 mm blades straight, blunt | Fine Science Tools, Heidelberg, Germany | 15025-10 | |
| Student dumont #7 forceps, inox | Fine Science Tools, Heidelberg, Germany | 91197-00 | |
| Student iris scissors, curved, 11.5 cm | Fine Science Tools, Heidelberg, Germany | 91461-11 | |
| Student iris scissors, straight, 11.5 cm | Fine Science Tools, Heidelberg, Germany | 91460-11 | |
| Student surcigal scissors, straight, sharp, 12 cm | Fine Science Tools, Heidelberg, Germany | 91402-12 | |
| Tissue forceps, 1 x 2 teeth, slim, 10 cm | Fine Science Tools, Heidelberg, Germany | 11023-10 | |
参考文献
- Prosser, B. L., Ward, C. W., Lederer, W. J. Subcellular Ca2+ signaling in the heart the role of ryanodine receptor sensitivity. J Gen Physiol. 136 (2), 135-142 (2010).
- Wehrens, X. H., Lehnart, S. E., Marks, A. R. Intracellular calcium release and cardiac disease. Annu Rev Physiol. 67, 69-98 (2005).
- Cheng, H., Cannell, M. B., Lederer, W. J. Propagation of excitation-contraction coupling into ventricular myocytes. Pflugers Arch. 428 (3-4), 415-417 (1994).
- Williams, G. S., Chikando, A. C., Tuan, H. T., Sobie, E. A., Lederer, W. J., Jafri, M. S. Dynamics of calcium sparks and calcium leak in the heart. Biophys J. 101 (6), 1287-1296 (2011).
- Sperelakis, N., Rubio, R. orderly lattice of axial tubules which interconnect adjacent transverse tubules in guinea-pig ventricular myocardium. J Mol Cell Cardiol. 2 (3), 211-220 (1971).
- Soeller, C., Cannell, M. B. of the transverse tubular system in living cardiac rat myocytes by 2-photon microscopy and digital image-processing techniques. Circ Res. 84 (3), 266-275 (1999).
- Song, L. S., et al. et al. ryanodine receptors in the failing heart. Proc Natl Acad Sci USA. 103 (11), 4305-4310 (2006).
- Oort, R. J., et al. Disrupted junctional membrane complexes and hyperactive ryanodine receptors after acute junctophilin knockdown in mice. Circulation. 123 (9), 979-988 (2011).
- Wagner, E., et al. Stimulated emission depletion live-cell super-resolution imaging shows proliferative remodeling of T-tubule membrane structures after myocardial infarction. Circ Res. 111 (4), 402-414 (2012).
- Asghari, P., Schulson, M., Scriven, D. R., Martens, G., Moore, E. D. Axial tubules of rat ventricular myocytes form multiple junctions with the sarcoplasmic reticulum. Biophys J. 96 (11), 4651-4660 (2009).
- Lukyanenko, V., Ziman, A., Lukyanenko, A., Salnikov, V., Lederer, W. J. Functional groups of ryanodine receptors in rat ventricular cells. J Physiol. 583 (Pt 1), 251-269 (2007).
- Shacklock, P. S., Wier, W. G., Balke, C. W. Local Ca2+ transients (Ca2+ sparks) originate at transverse tubules in rat heart cells. J Physiol. 487 (Pt 3), 601-608 (1995).
- Reynolds, J. O., et al. Junctophilin-2 is necessary for T-tubule maturation during mouse heart development. Cardiovasc Res. 100 (1), 44-53 (2013).
- Di Maio, A., Karko, K., Snopko, R. M., Mejia-Alvarez, R., Franzini-Armstrong, C. T-tubule formation in cardiac myocytes two possible mechanisms. J Muscle Res Cell Motil. 28 (4-5), 231-241 (2007).
- He, J., et al. Reduction in density of transverse tubules and L-type Ca(2+) channels in canine tachycardia-induced heart failure. Cardiovasc Res. 49 (2), 298-307 (2001).
- Heinzel, F. R., et al. Remodeling of T-tubules and reduced synchrony of Ca2+ release in myocytes from chronically ischemic myocardium. Circ Res. 102 (3), 338-346 (2008).
- Lyon, A. R., et al. Loss of T-tubules and other changes to surface topography in ventricular myocytes from failing human and rat heart. Proc Natl Acad Sci USA. 106 (16), 6854-6859 (2009).
- Crossman, D. J., Ruygrok, P. N., Soeller, C., Cannell, M. B. Changes in the organization of excitation-contraction coupling structures in failing human heart. PLoS One. 6 (3), e17901(2011).
- Kemi, O. J., et al. The effect of exercise training on transverse tubules in normal, remodeled, and reverse remodeled hearts. J Cell Physiol. 226 (9), 2235-2243 (2011).
- Sachse, F. B., et al. Subcellular structures and function of myocytes impaired during heart failure are restored by cardiac resynchronization therapy. Circ Res. 110 (4), 588-597 (2012).
- Arakel, E. C., et al. Tuning the electrical properties of the heart by differential trafficking of KATP ion channel complexes. J Cell Sciences. 127 (Pt 9), 2106-2119 (2014).
- Richards, M. A., et al. Transverse tubules are a common feature in large mammalian atrial myocytes including human. Am J Physiol Heart Circ Physiol. 301 (5), H1996-H2005 (2011).
- Trafford, A. W., Clarke, J. D., Richards, M. A., Eisner, D. A., Dibb, K. M. Calcium signalling microdomains and the t-tubular system in atrial mycoytes potential roles in cardiac disease and arrhythmias. Cardiovasc Res. 98 (2), 192-203 (2013).
- Greiser, M., Schotten, U. Dynamic remodeling of intracellular Ca2+ signaling during atrial fibrillation. J Mol Cell Cardiol. 58, 134-142 (2013).
- Voigt, N., Zhou, X. B., Dobrev, D. Isolation of human atrial myocytes for simultaneous measurements of Ca2+ transients and membrane currents. J Vis Exp. (77), 10-3791 (2013).
- Kaestner, L., et al. Isolation and genetic manipulation of adult cardiac myocytes for confocal imaging. J Vis Exp. (31), (2009).
- Louch, W. E., Sheehan, K. A., Wolska, B. M. Methods in cardiomyocyte isolation, culture, and gene transfer. J Mol Cell Cardiol. 51 (3), 288-298 (2011).
- King, N. M., et al. Mouse intact cardiac myocyte mechanics cross-bridge and titin-based stress in unactivated cells. J Gen Physiol. 137 (1), 81-91 (2011).
- Schindelin, J., et al. Fiji, an open-source platform for biological-image analysis. Nat Methods. 9 (7), 676-682 (2012).
- Arganda-Carreras, I., Fernandez-Gonzalez, R., Munoz-Barrutia, A., Ortiz-De-Solorzano, C. 3D reconstruction of histological sections Application to mammary gland tissue. Microsc Res Tech. 73 (11), 1019-1029 (2010).
- Liu, Z. Q. Scale space approach to directional analysis of images. Appl Opt. 30 (11), 1369-1373 (1991).
- Powell, T., Twist, V. W. A rapid technique for the isolation and purification of adult cardiac muscle cells having respiratory control and a tolerance to calcium. Biochem Biophys Res Commun. 72 (1), 327-333 (1976).
- Watanabe, Y., et al. Inhibitory effect of 2,3-butanedione monoxime (BDM) on Na(+)/Ca(2+) exchange current in guinea-pig cardiac ventricular myocytes. Br J Pharmacol. 132 (6), 1317-1325 (2001).
- Kohl, T., Westphal, V., Hell, S. W., Lehnart, S. E. Superresolution microscopy in heart - cardiac nanoscopy. J Mol Cell Cardiol. 58, 13-21 (2013).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。