
Method Article
Analyse von Membranrohrnetze in Herzmuskelzellen von Vorhöfe und Kammern
* Diese Autoren haben gleichermaßen beigetragen
In diesem Artikel
Zusammenfassung
In Herzmuskelzellen, röhrenförmigen Membranstrukturen intrazellulären Netzwerken. Wir beschreiben optimierte Protokolle für i) Isolierung von Myozyten aus Maus Herz einschließlich der Qualitätskontrolle, ii) Lebendzellfärbung für state-of-the-Art-Fluoreszenzmikroskopie, und iii) die direkte Bildanalyse, um die Komponente Komplexität und die Plastizität der intrazellulären Membran quantifizieren Netzwerken.
Zusammenfassung
In cardiac myocytes a complex network of membrane tubules - the transverse-axial tubule system (TATS) - controls deep intracellular signaling functions. While the outer surface membrane and associated TATS membrane components appear to be continuous, there are substantial differences in lipid and protein content. In ventricular myocytes (VMs), certain TATS components are highly abundant contributing to rectilinear tubule networks and regular branching 3D architectures. It is thought that peripheral TATS components propagate action potentials from the cell surface to thousands of remote intracellular sarcoendoplasmic reticulum (SER) membrane contact domains, thereby activating intracellular Ca2+ release units (CRUs). In contrast to VMs, the organization and functional role of TATS membranes in atrial myocytes (AMs) is significantly different and much less understood. Taken together, quantitative structural characterization of TATS membrane networks in healthy and diseased myocytes is an essential prerequisite towards better understanding of functional plasticity and pathophysiological reorganization. Here, we present a strategic combination of protocols for direct quantitative analysis of TATS membrane networks in living VMs and AMs. For this, we accompany primary cell isolations of mouse VMs and/or AMs with critical quality control steps and direct membrane staining protocols for fluorescence imaging of TATS membranes. Using an optimized workflow for confocal or superresolution TATS image processing, binarized and skeletonized data are generated for quantitative analysis of the TATS network and its components. Unlike previously published indirect regional aggregate image analysis strategies, our protocols enable direct characterization of specific components and derive complex physiological properties of TATS membrane networks in living myocytes with high throughput and open access software tools. In summary, the combined protocol strategy can be readily applied for quantitative TATS network studies during physiological myocyte adaptation or disease changes, comparison of different cardiac or skeletal muscle cell types, phenotyping of transgenic models, and pharmacological or therapeutic interventions.
Einleitung
Bei gesunden gestreiften Muskelzellen, sind röhrenförmige Membranstrukturen mit "quer" Orientierungen (T-Tubuli) senkrecht zu den Hauptzellenachse reichen. Folglich haben T-Tubuli als kontinuierliche Verlängerungen der Hauptmuskelzelle "seitlich" Oberflächenmembran (Sarkolemm), die tief in das Cytosol der Zelle zu durchdringen Zentrum gekennzeichnet. Die physiologische Rolle der T-Tubuli kontinuierlich mit der Außenfläche der Membran eine schnelle elektrische Kopplung der Fernintrazellulären Kompartimenten durch SER Organell Kontaktbereiche im gesamten verhältnismäßig großen Herzmuskelzellvolumens von nanometrischen Nähe Kopplung von spannungsaktivierten L-Typ-Ca 2 + gebildet Kanäle (Cav1.2) nach innen (I Ca) Aktivieren benachbarten Ryanodinrezeptor (RyR2) SER Ca 2 +-Releases. In ventrikulären Myozyten (VMs), die nicht-kontinuierliche Membran Kontakte ("junctions") zwischen den Knoten-SER-Domänen und T-tubules sind gedacht, um Tausende von Einzelintrazelluläre Ca2 + Freisetzung Nanodomänen in jeder Zelle 1 zu steuern.
Für jede gegebene Kontakt Domäne die nebeneinander liegenden Membranteile jeder der T-Tubuli und dem Peripheriegerät (junctions) SER etwa 15 nm nahe zueinander, also als Nanodomänen definiert. Dadurch werden sehr kleine zytoplasmatische einzelnen Teilräume getrennt, welche quasi-autonomen Zellenabteil Verhalten ermöglichen. Wenn ein eingehender Aktionspotential aktiviert Cav1.2-Kanäle in den T-Tubuli von VMs, einem relativ kleinen Ca 2 + Einstrom wird rasch zunehmen den Unterraum Ca 2 +-Konzentration [Ca2 +] S in der Größe Attolitervolumina Nanodomänen 1. Nächsten, aktiviert der S Zunahme [2 + Ca] Ca 2 + -regulierte Ryanodinrezeptoren (RyR2) in Nanometer in der Nähe nebeneinander SER Membran Kreuzung, und das Kupplungsvorgang in ganz elektrisch Paar auftrittd myocyte Nanodomänen. RyR2s auftreten als dichte Multi-Channel-Cluster mit einem geschätzten Stöchiometrie von 1 Cav1.2-Kanal für 5-10 RyR2 Kanäle 2. Seit der SER-to-Cytosol [Ca 2 +] Steigung ist sehr steil (Verhältnis 10 4: 1) und RyR2s funktionieren als Hoch Leitfähigkeit Ca2 +-Kanäle in Release funktionell gekoppelt Cluster, RyR2 Aktivierung führt zu einer quantitativ großen Ca 2 + Auslösestrom von T-Tubuli gekoppelt junctional SER Domains Erhöhung der lokalen Unterraum [Ca 2 +] S bis 100 um oder mehr innerhalb von 1-2 ms 3,4. Diese Herzsignalverstärkung Verhalten wird auch als Ca 2 + induzierten Ca2 + Freisetzung (ZPK) bezeichnet. Zusammengenommen T-Tubuli sind wesentliche Membranstrukturen, die während der Erregung und Kontraktion (EG) Kupplung schnell aktivieren Ca2 + Freisetzung Signale durch junktionale Nanodomänen SER Kontakte und Zell-breite IKRK.
Neben T-Tubuli, axial Tubuluss (A-Tubuli) mit deutlich unterschiedlicher Ausrichtung parallel zu den Haupt (längs) Zellenachse durch Elektronenmikroskopie (EM), konfokale und 2-Photonen-Mikroskopie Studien dokumentiert. So wurde beispielsweise eine Zelle weiten kontinuierlichen Gitter von A-Tubuli zwischen Myofibrillen mit T-Tubuli in der Nähe von Sarkomer Z-Leitungen miteinander verbunden durch extrazelluläre Tracer und EM-Bildgebung des Anlage Meerschweinchen VMs 5 gezeigt. Mit extrazellulären Dextran-linked Fluoreszeinfärbung und leben 2-Photonen-Imaging von Ratten VMs, wurde eine komplexe netzartige 3D-Röhrchen Netzwerk visualisiert, bestehend aus ~ 60% T-Tubuli und ~ 40% A-Tubuli 6. Diese Studie nicht nur 3D-Visualisierung von reichlich A-Röhrchen geführt, sondern auch zu der Erkenntnis, dass die Schneiden zum EM-Visualisierung ist inhärent für die Analyse von komplexen und dynamischen Membran Netzwerke wie queraxialer Tubulus System (TATS) begrenzt. Folglich konfokale Live Cell Imaging TATS Membranen direkt mit Bunt di-8-ANNEPS entwickelt. Wenn Live CellTATS Netzwerke werden durch Fourier-Transformation analysiert werden, wird der reguläre Auftreten von T-Tubuli Komponenten im Raum in der Nähe Sarkomerlänge Z-Linien von dem Ensemble Leistungsspektrums von einem Bereich der quergestreiften Signale 7 reflektiert. Diese indirekte Analyse Strategie wurde verwendet, um die Zellweit regionale Veränderungen in TATS Komponente Regelmäßigkeit in Krankheitsmodellen 7 zu erkennen. Zum Beispiel verursacht shRNA vermittelten junctophilin-2 knock-down Herzinsuffizienz und die Isoform spezifische Protein-Mangel führte zu T-Tubuli Reorganisation mit Nanodomänen Ca 2 + Release 8 Dysfunktion. Wir haben vor kurzem erweitert die Analyse von Membran TATS Netze durch direkte quantitative Ansätze und weiter durch Superresolution-Mikroskopie an lebenden Zellen der einzelnen Komponenten in TATS Maus VMs, die stimulierte Emission Depletion (STED) Nanoskopie 9. Nanometrischen Bildauflösung für die direkte Analyse von kleineren Einzel TATS Komponenten, die eine 50:50 Verteilung der Quernähern erlaubtgegenüber dem axialen Röhrchen Orientierungen, quantitativ bestätigt zwei reichlich noch unterschiedlich orientierte Einzel TATS Komponenten bei gesunden Mäuseherzen 9. Diese Strategien werden in dem Protokoll weiter unten beschrieben werden.
Während die physiologische Rolle der reichlich vorhandenen A-Tubulus Komponenten im erwachsenen Herzen hat rätselhafte blieb, haben EM-Studien SER mit A-Tubuli was darauf hindeutet, endogenen Ca2 + Freisetzung Nanodomänen in Meerschweinchen und Ratte VMs 5,10 verbundenen Membranstrukturen dokumentiert. Konfokale Analyse der Cav1.2 und RyR2 fanden eine hohe Kolokalisation bei A-Tubulus Gänge 10. Da ca. 20% der spontanen Ca2 + Funken in Ratten VMs entstand relativ weit entfernt von Z-Linie Streifen, wo T-Tubuli treten typischerweise auf, hat ein Argument gewesen, dass A-Röhrchen verbunden Nanodomänen kann in der Tat existieren und funktionieren, wie Ca2 + Freisetzung Websites 11,12. Interessant ist, dass T-Tubuli Bildung und Reifung ocCURS erst nach der Geburt und verläuft parallel zum Wachstum der Herzzellen, beispielsweise durch Austreiben von Vorläufer sarkolemmalen Einstülpungen an P5 und unreife verzweigte TATS Netzwerk Baugruppen bei P10 in 13 Mäuse. Es scheint, dass Junctophilin-2 ist besonders wichtig für postnatalen Reifung TATS Netzwerk seit shRNA knock-down verhindert die Verankerung der T-Tubuli Membranen SER Gänge, die zu verzögerten Ca2 + Freisetzung und pathologischen TATS Organisation, die mit unreifen A-Tubulus dominiert Architekturen VMs 13. Diese Beobachtungen können letztlich zu Proof-of-Concept, die T-Tubuli bilden durch Membranprozesse während eine Einstülpung-Tubuli kann durch zusätzliche oder sogar alternative intrazelluläre Mechanismen 14 morphen führen.
Charakterisierung von Membran TATS Veränderungen der Herzkrankheit ist zu einem wichtigen Forschungsbereich für pathophysiologische Fragen. Erste Berichte in einem Hundemodell der Stimulation-induzierte Herz failure zeigte einen Verlust von T-Tubuli und Cav1.2 (I Ca) 15. Ein Schweinemodell des ischämischen Kardiomyopathie zeigte T-Tubuli Dichten reduziert und ein reduziertes Synchronität der intrazellulären Ca 2 + frei 16. Mit Hilfe eines spontan hypertensiven Ratten (SHR) Modell der Herzinsuffizienz, wurde ein Verlust von T-Tubuli mit reduzierter Nanodomänen Kopplung von Cav1.2 und RyR2 durch die vorgeschlagene Mechanismus der "RyR2 verwaisen" assoziiert 7. Ein Verlust der T-Tubuli hat auch in der Human VMs von ischämischen, erweiterte und hypertrophe Kardiomyopathie Proben 17 gezeigt. Darüber hinaus wurde eine Zunahme in A-Tubuli in Gewebeschnitten von menschlichen Kardiomyopathie 18 gemeldet. Nach Myokardinfarkt, zeigten wir eine Differentialmechanismus TATS Reorganisation in der Maus VMs mit einer signifikanten Abnahme der T-Tubuli im Gegensatz zu Erhöhungen der A-Tubulus Komponenten 9. Wichtig ist, Verbesserung der lokalen Membran dagegen durch Lebendzell superre erreichtLösung der STED-Mikroskopie konnten detaillierte quantitative Komponentenanalyse durch direkte Messungen, die erhebliche Vermehrung der A-Tubuli mit insgesamt steigt der TATS Netzlänge und Verzweigung Komplexität 9 zeigte. Darüber hinaus wurde gezeigt, dass körperliches Training kann T-Tubuli Umbau bei Ratten nach Myokardinfarkt und 19, dass die kardiale Resynchronisationstherapie kann dazu führen, Umbau der T-Tubuli bei Hunden mit Vorhof tachypacing induzierten Herzinsuffizienz 20 rückwärts umzukehren. Zusammengenommen werden Studien sowohl in erkrankten menschlichen und tierischen VMs sowie mögliche therapeutische Interventionen wohl aus hochwertigen Zellisolationsverfahren und detaillierte quantitative Analyse Strategien profitieren, wie im Protokoll beschrieben und Ergebnisse Abschnitten.
Darüber, wie kürzlich durch Seitenfläche demonstriert gegen TATS Membrantransport von KATP-Kanal-Isoformen 21, ist es wichtig, Vorhof m betrachtenyocytes (AMS) als biologisch verschieden sowie vergleichende Herzzellmodell gegenüber VMs. T-Tubuli wurden kürzlich bei Schafen und menschlichen BM 22 dokumentiert. Aktuelle Hinweise darauf, dass einige T-Tubuli existieren in AM-Zellen und in der Regel in größeren Säugetieren wie Schafe und Menschen, aber nicht in 23 kleine Nagetiere. Im Gegensatz zu VMs, AMS intrazelluläre Ca2 + Freisetzung wird durch Diffusion in Richtung der Zellenmitte, die in räumlicher und zeitlicher gekennzeichnet Ca 2 +-Gradienten führt zu 23 von der Zelloberfläche ausbreitet, auftritt. In diesem Rahmen erscheint es wichtig, die Mechanismen der intrazellulären Ca 2 +-Signalisierungs Instabilität für die gemeinsame Krankheitsformen wie Vorhofflimmern 24 aufzuklären. Zusammenfassend werden sowohl AM und VM Zellisolierung und jeweils für gesunde und kranke Herz üblicherweise verwendeten Protokolle. Nur wenn der Zellisolierung richtig gemacht wird, wie durch mikroskopische Dokumentation ausreichender Zellqualität erachtet wird, sollte AM und VM Proben ca seinrried vorwärts für die quantitative Analyse TATS. Dementsprechend sind die folgenden Abschnitte Protokoll entscheidend von Qualitätszelle Isolate aus der Maus oder anderen Spezies, gefolgt von Lebendzellmikroskopie zu intakten Membranen TATS analysieren. Wie zuvor erwähnt, Charakterisierung von Membranen TATS ist eine anspruchsvolle Forschungsbereich mit einem Hang zur Fixierung und Vorbereitung Artefakte 6, Membranveränderungen durch osmotische Änderungen und Auflösung Grenzen der konventionellen Lichtmikroskopie 9. Wir stellen fest, dass die jüngsten state-of-the-Art-Protokolle für die Isolierung von humanen BM für Ca2 + Imaging und Patch-Clamp-und Ratten VMs für die Zellkultur wurden bereits in dieser Zeitschrift veröffentlicht 25,26.
Protokoll
HINWEIS: Alle Tier Verfahren wurden überprüft und von der Institutional Animal Care und Use Committee der Universitätsmedizin Göttingen in Übereinstimmung mit der humanen Pflege und Verwendung von Labortieren zugelassen.
1. Isolierung von atrialen und ventrikulären Myozyten von der Maus Herz
- Hand Tiere so schonend wie möglich und nach anerkannten Protokollen, um Stress im Allgemeinen zu minimieren und speziell auf mögliche unbeabsichtigte Effekte aus leistungsstarken neurohormonalen Exzesse an isolierten Herzzellen zu vermeiden. Zusätzlich zu injizieren jede Maus mit Heparin (500 IU / kg Körpergewicht sc) mindestens 20 min vor der Herz Extraktion Blutgerinnung und Mikroembolien, die die Ausbeute und die Integrität der Herzzellen während der Isolierung erheblich beeinträchtigen können, zu verhindern.
- Betäuben Mäuse im Alter von 12 Wochen oder älter von Isofluran Inhalation, Abwesenheit von Schmerz Rückzug Reflexe zu bestätigen, und einschläfern Tiere durch Genickbruch.
- Entpacken Sie die Herzen schnell nach vorher festgelegten Experte Protokolle (siehe zB Kaestner et al. Louch 26 und et al. 27). Vermeiden Sie jede unbeabsichtigte Beschädigung der Vorhöfe durch unnötige Quetschen oder Stretching.
- Sorgfältig bewahren das Gewebe des proximalen aufsteigenden Aorta mit einem Paar Stumpf Pinzette und Schere gerade, eine kontinuierliche Querschnittkante durch die Gefäßwand der Aorta, die wichtig für die erfolgreiche Kanülierung und Perfusion des Herzens ist zu etablieren.
- Übertragen Sie die ausgeschnittenen Herzen sofort in eiskaltem nominell Ca2 + frei Perfusionspuffer (Lösungen siehe Tabelle 1). Halten Sie die großen Gefäße während der Übertragung durch die Luft in den Puffer eingespannt, um eine unbeabsichtigte Luftembolie zu vermeiden, bis das Herz vollständig untergetaucht. Verwenden BDM in den Puffer und eisgekühlten Lösungen für Herzkontraktion hemmen.
- Verwenden Sie ein Fernglas Zoom-Mikroskop mit ausreichender 3D Illumfung.
- Kanülieren die Aorta unter Panorama-Stereo des Herzens mit einer glatten, polierten Oberfläche 21 G Kanüle (Außendurchmesser 0,81 mm; für normale Maus Herzgewicht), die vollständig mit Puffer gefüllt werden müssen. Sicherzustellen, dass sich keine Luftblasen in der Kanüle durch Verbinden eines proximalen Lösungsreservoir (beispielsweise eine Spritze) über ein 2-Wegeventil, das Luer zur schnellen Lösungsflusskontrolle.
- Bestätigung unter Fernglases, daß die Kanüle richtig in die Aorta, die etwa 1 mm oberhalb der Aortenklappe und Koronararterie Verzweigungen positioniert ist. Vermeiden Sie unbedingt Durchgang durch oder unbeabsichtigte Perforation der Aortenklappen mit der Kanüle (dies dauerhaft stören Aortenklappenverschlusses und damit stören Herzdurchblutung).
- Binden Sie die Aorta vorsichtig, um maßgeschneiderte, Umfang ausgerichteten Anti-Rutsch-Rillen in der Nähe des Ende der Kanüle mit zwei Seidenfäden. Der Herzkranz arteri wegspülenES mit Nachdruck an einer beliebigen Stelle. Schließen Sie die Lösung gefüllt Kanüle in die Aorta und dem Herzen verbunden, um eine eng anliegende Ablaufstutzen eines maßgeschneiderten und vor-kalibriert Perfusion System, auch bekannt als der modifizierten Langendorff-Setup (entweder mit konstantem Druck oder konstanten Fluss, siehe auch Diskussion weiter unten).
- Perfusion des Herzens so bald wie möglich für 4 min mit Sauerstoff angereicherten Perfusions-Puffer bei 37 ° C (Solldurchblutungsrate: 4 ml / min). Beginnen die Verdauung durch Umschalten der Perfusion Kollagenase enthaltenden Aufschlusspuffer (600 U / ml Collagenase Typ II) für 8-10 min bei 37 ° C aufweist. Überwachen Sie den Fortschritt von Gewebe Verdauung durch ähnliche Gewebeveränderungen einschließlich der Erhöhung Undurchsichtigkeit, Weichheit und Schlaffheit in der gesamten Herzoberfläche offensichtlich bestätigt.
- Sezieren die Herzkammern bei Bedarf nach der Verdauung. Legen Sie die Kanüle eingeführt Herz unter dem Binokular und visualisieren die hintere Herzwand. Sezieren Rest nicht-kardialen Gewebe zB Lunge einnd zu Zelle Kontamination in der Verdauungspuffer mit Mikro Scheren zu vermeiden Gefäßteile (zB Feder Schere mit 8 mm gerade Klingen), wie in Abbildung 1 dargestellt.
- Befolgen Sie eine Checkliste, die insbesondere Kammern, Regionen und / oder Zellen der Kollagenase verdaut Herz sollte geerntet werden (Abbildung 1): links und / oder rechts Atrium, kostenlose linken und / oder rechten Herzkammerwand, und / oder der Kammerscheidewand.
- Für die Präparation von bestimmten Herzgewebe, mit einem relativ breiten und flachen Dissektion Bad mit mehreren mm dicken Schicht aus Silikon-Elastomer Kunststoff beschichtet. Befestigen Sie die Spitze des Herzens mit einem feinen Insektenstahlstift auf den Boden Elastomerschicht.
- Lenken den rechten Herzohr und sezieren den rechten Vorhof direkt über den AV-Klappen. Fahren Sie mit der Dissektion mit dem linken Vorhof. Sezieren und entsorgen Sie die Faserventilvorrichtung. Schließlich sezieren die links und rechts frei ventrikulären Wände und das Septum und / oder kleineER Gewebeteile, wie gebraucht.
- Hinweis nur für Auszubildende: die Praxis zu gewinnen, beginnen Sie mit der Maus nicht verdaut Herzen. Zur Vereinfachung der anatomischen Orientierung, üben das Gewebe Umgang mit allen aufeinanderfolgenden Schritte Dissektion unter binokularen Sehens, wie in Abbildung 1 dargestellt. Sobald der 3D-Anatomie, manuelle Handhabung unter binokularen Sehens und Dissektion Schritte ausreichend vertraut, fahren Sie mit der Maus Kollagenase verdaut Herzen als oben beschrieben.
- Für Ventrikelmyocyten (VM) Zelltrennung: überweisen Sie den ventrikulären Gewebe in 2,5 ml frischem Verdauungspuffer. Wenn gleichzeitige Zellisolierungen aus mehreren Organteilen, beispielsweise Vorhöfe und Kammern versucht wird, kann eine zweite Person über einen Schenkel des Zelldissoziationsmedium Verfahren beteiligen, sowohl zu minimieren und die Verwendung von Mäusen durch koordinierte Handhabung von mehreren Herzgewebe. Fahren Sie mit Schritte 1.9.1-1.9.4, danach 1,10.
- Sezieren entweder die gesamte Kammergewebeoder bestimmte Teile davon (zB LV, RV, freie Wände und / oder Septum) in etwa 1 mm 3 Stück in 2,5 ml Verdauungspuffer mit einer scharfen Schere (zB Feder Schere mit 8 mm gerade Klingen) in einer 60 mm Petrischale .
- Sanft distanzieren VMs in Zellsuspension durch langsame Verreiben der Gewebestücke mit einer Transferpipette. Vermeiden Sie Luftblasenbildung in der Zellsuspension.
- 8 ml Stoppuffer zu dem VM Zellsuspension und übertragen die Zellsuspension in einem 15 ml konischen Röhrchen. Lassen Sie die restlichen Gewebestücke an der Unterseite ca. 15 Sekunden absetzen, aber kurz genug, für isolierte Zellen in Suspension bleiben. Als nächstes ernten die VM Suspension über Transfer von der überstehenden Volumen auf ein neues 15-ml-Tube. Wenn übermäßige Gewebeteile vorhanden sind, alternativ eine minimal 200 um voneinander Nylonnetz, um die Gewebestücke aus der Zellsuspension zu trennen.
- Lassen Sie die VM Zellsuspension in den Boden eines 15 ml Co absetzenschen Rohr durch die Schwerkraft für 8 min.
- Waschschritt: Entfernen Sie den Überstand und resuspendieren sanft die verbleibende VM Pellet in 10 ml Perfusionspuffer. Wiederholen Waschschritt 1.9.5 (Optional: Fügen Sie zusätzliche Waschschritte schrittweise Erhöhung der Ca2 +-Konzentration nach Bedarf).
- Resuspendieren siedelt VM Pellet in 10 ml Perfusionspuffer und verteilen Sie die restlichen Zellsuspension in 1,5 ml Volumen-Mikrozentrifugenröhrchen (ca. 50.000 Zellen pro VM Rohr).
- Für Vorhof Myozyten (AM) Zelltrennung: überweisen Sie den verdaut / zerlegt Vorhofgewebe in 1 ml frischem Verdauungspuffer.
- Schneiden Sie die teilweise verdaute Vorhofgewebe in etwa 1 mm 3 Stück in 1 ml Verdauungspuffer mit Mikro Schere in eine kleine Petrischale (zB Durchmesser 60 mm). Sanft dissoziieren AM-Zellen aus der verdauten Gewebestücke in Zellsuspension mittels Verreiben mit einer 1 ml-Plastikpipette mit einer Schnittspitze schädlichen Fluidstrahlen zu vermeiden. WährendVerreiben strikt vermeiden Sie Luftblasenbildung in der Zellsuspension. Nach mechanischer Bewegung, 4 ml Stop-Puffer (50 uM CaCl 2, 10% BCS), um alle verbleibenden Kollagenase-Aktivität in Zellsuspension zu verhaften.
- Übertragen Sie die Uhr Zellsuspension in einem 15 ml konischen Röhrchen. Die Aufnahme der verbleibenden Gewebestücke an der Unterseite etwa 15 Sekunden zu regeln, aber kurz genug für die isolierten Zellen in Suspension verbleiben. Ernten Sie die überstehende Band mit den freien AM-Zellen über-Lösung an einem neuen 15-ml-Tube.
- Zentrifugieren Sie die Uhr Zellsuspension, zB 2 min bei 20 g bei RT oder - bevorzugt Membranstudien - lassen sich die Zellen absetzen langsam durch die Schwerkraft für 20 min in einem 15 ml konischen Röhrchen.
- Waschschritt: den Überstand verwerfen und sanft resuspendieren Uhr Pellet in 5 ml Perfusionspuffer. Wiederholen 1.10.4.
- Resuspendieren Uhr Zellen vorsichtig in 5 ml Perfusionspuffer. Verteilen Sie die Zellsuspension in 1,5 ml Volumen ReaktionsgefäßeES (ca. 1000 Uhr Zellen pro Röhrchen).
- Analysieren und dokumentieren die isolierten Zellpopulation Qualität für jeden Herz einschließlich der Zellausbeute mit Trypanblaufärbung.
- Hierzu verdünnte 500 ul der Zellsuspension zu 1: 1 vol / vol mit Trypanblau-Lösung (Endkonzentration 0,02%) unter Verwendung von 1 ml-Pipettenspitzen geschnitten. Mischen die Zellen und die Trypanblau sanft durch sehr langsam nach oben / unten Pipettieren. Das Trypanblau haltigen Zellsuspension auf eine Neubauer-Typ Durchflusszytometer verbessert sofort an und rechnet auf die intakten Myozyten mit einem inversen Mikroskop.
- Schließen Sie alle Zellen mit offensichtlichen Schäden, Membran Blasen, Schlieren gestört, Kontrakturen und Zellen ansammeln intrazellulären Trypanblau (Abbildung 2). Auch ausschließen spontan kontra Zellen, die anfällig für nachfolgende Zelltod sind. Um die Anzahl der intakten Zellen in Suspension zu bewerten, verwenden Sie nur Herzmuskelzellen mit regelmäßigen Streifen, die während Trypanblau auszuschließendas Zellvolumen.
- Beurteilen die Integrität der einzelnen atriale und ventrikuläre Myozyten von Durchlichtmikroskopie. Speichern Sie die hellen Bildfeld als Tif-Datei für die Dokumentation und die weitere Analyse.
- Verwenden Sie die folgenden Kriterien für die Analyse der Herzzellintegrität:
- bestätigen das Vorhandensein von regelmäßigen Streifen über das sichtbare Zellvolumen;
- bestätigen die kontinuierliche Integrität der Seitenfläche Membran auf beiden Zellenseiten parallel zu den Myofilamenten;
- visualisieren scharfen Zacken an den Glanzstreifen auf beiden Seiten Zelle, die die Integrität der spezifischen Oberfläche Membranstrukturen zu reflektieren; und
- visualisieren die Fluoreszenzsignale von TATS Membranen (oder immun Caveolin-3-Protein oder anderen Membran Marker), wie in Abschnitt 3 für die Lokalisierung Korrelation neben dem zugrunde liegenden zellspezifischen Hellfeld Bild Morphologie (zB kombinieren beide Bilder mit ImageJ als Overlay-Verbindung beschrieben image).
- Bestimmen Sie die Sarkomerlänge von den hellen Bereich Bilder. Für mittlere Sarkomerlänge, messen Sie den Abstand von sequentiell ausgerichtet Sarkomer Streifen, und teilen Sie den Abstand durch die Anzahl der Sarkomere. Messung an mindestens zwei Stellen pro Zelle. Führen Sie die Analyse mit kommerzieller Software oder offline mit ImageJ.
HINWEIS: Wenn mit Trennmitteln behandelt, intakte entspannt VMs von Mäuseherzen zeigen eine mittlere Sarkomerlänge von ~ 1,9 um 28.
- Verwenden Sie die folgenden Kriterien für die Analyse der Herzzellintegrität:
- Quantifizierung der Morphologie und Abmessungen der einzelnen atriale und ventrikuläre Myozyten aus dem übertragenen Bild der Lichtmikroskopie. Bedenken Sie, dass AM und VM-Zellen unterscheiden sich deutlich in der Größe. Messen Sie die Zelle Länge, Breite und Fläche, und berechnen Sie die Länge: Breite-Verhältnis.
- Analysieren Sie die 2D-Zelldimensionen von der Durchlichtbild in ImageJ mit den Befehlen Polygon-Auswahl-Werkzeug und in den ROI-Manager, wie in Abbildung 3. Wenn morphologischen cHanges innerhalb bestimmter Kontexte Studie erwartet weiteres Dokument für alle Zellen die spezifische Mausstamm, Alter, Geschlecht, Herzgröße und alle Interventionen für die anschließende Datenklassifizierung.
2. Die Färbung der TATS Membranen in Wohn atriale und ventrikuläre Myozyten
- Legen Sie die Imaging-Kammer (zB POC-R2) mit einem 42 mm Deckglas. Für eine stabile Befestigung an der Myozyten Deckglas, bereiten 20 ul Laminin-Lösung von 1:10 Verdünnung des Laminin Lager in physiologischen Perfusion Puffer (Endkonzentration 0,2 mg / ml). Verbreiten Sie 20 ul der Laminin-Lösung gleichmäßig auf dem Deckglas.
- Planen 800 ul einer 50 uM di-8-ANEPPS Lösung in Perfusionspuffer. Dazu verdünnen 20 ul 2 mM di-8-ANEPPS Stammlösung in 780 ul physiologische Puffer.
- VMs Fleck, lassen sich die Zellen absetzen durch die Schwerkraft für 8 min in einem 1,5 ml Reaktionsgefäß. AMs zu färben, verwenden Sie entweder die Schwerkraft Sedimentation oder spin der Zellsuspension für 2 Minuten (siehe 1.10.3). Für beide AMs und VMs, entfernen Sie vorsichtig den Überstand unter Vermeidung unnötiger Aufregung des Zellpellets und sanft Zellpellet in 800 ul von Di-8-ANEPPS haltigen Lösung (50 um). Die di-8-ANEPPS / myocyte Suspension unmittelbar übertragen auf die Laminin beschichteten Deckglas in der bildgebenden Kammer.
- Flecken auf der VM Suspension für 15 min bei RT im Dunkeln.
- Langsam überschüssiges Volumen über die Aufwärtsfluidmeniskus an den Seiten der Abbildungskammer mit einer manuellen Pipette. Zu bestätigen, dass die Mehrzahl von Di-8-ANEPPS gefärbten Myozyten bleibt fest mit dem Laminin beschichteten Deckglas befestigt und werden erst an der Luft. Weiter, wäscht den beigefügten Myozyten Suspension einmal durch langsame Zugabe von 1 ml Puffer, gefolgt von Perfusion Entfernen jeglicher überschüssiger Flüssigkeit mit nicht-adhärenten Zellen.
- Sorgfältig überlagern die gefärbten und Fläche befestigt Myozyten mit 1 ml Perfusionspuffer langsam von der Seite des Ter Imaging-Kammer. Setzen Sie die Druckkammer auf dem Mikroskoptisch.
3. Imaging TATS Membrane Structures in Wohn atriale und ventrikuläre Myozyten
- In der Regel sorgfältig auswählen, die bestmögliche Option Fluoreszenzmikroskop (e) für TATS Membran-Bildgebung zur Verfügung. Für die konfokale Bildgebung, betrachten letzten Generation, moderne Fluoreszenzmikroskopen mit optimierten PMT-Array-Detektoren und Recycling Photonenwege, die Fluoreszenzsignalintensität zu maximieren. Für die konfokale Bildgebung von kleineren Details von TATS Membranstrukturen verwenden eine 63X 1,4 NA Öl-Objektiv oder - je nach Verfügbarkeit - mit einem STED-Mikroskop mit Superauflösung für kleinste Details TATS wie für Myozyten-spezifische Anwendungen von Kohl et al 34 Für allgemeine Grundsätze des Hochprüft. Auflösung der Fluoreszenzmikroskopie, beziehen sich auf den Diskussionsteil.
- Stellen Sie die Aufnahmeparameter, die meisten erkennen, idealerweise alle Di-8-ANEPPS gefärbten intrazellulären memBranen innerhalb einer bestimmten myocyte Abbildungsebene. Verwenden Sie die folgenden Parameter als Ausgangspunkt für die konfokale Laserscanmikroskopie: Anregung 458 nm zB bei 3% der maximalen Laserleistung; detektieren des emittierten Signals zwischen 550 nm und 740 nm; Detektorverstärkung (zB Master-Befehl 800); und Lochblende 1 AU für eine optische Schichtdicke von 900 nm auf. Diese Parameter einzustellen, um das Signal-zu-Rausch-Optimierung.
- Verwenden Sie das Hellfeld-Modus, um eine intakte Uhr oder VM Zelle als angemessen (4A und 4B) zu wählen. Siehe für relevante Kriterien, wie Zellintegrität richten zusammengefasst, um Zellen auszuwählen Punkt 1.12: Zellweit regelmäßige Rillen und gleich Sarkomer Abstand, scharfe Kanten und Zacken Oberfläche auf allen vier Zellseiten, kontinuierliche Integrität der Seitenfläche Membran und des Fehlens von keine Membran Bläschen.
- Nehmen Sie ein Beispielbild von einer zentralen intrazellulären myocyte Abschnitt. Zur Einstellung der ROI mit der "Ernte"-Funktion →; Stellen Sie die Schnittfenster → die endgültige Pixelgröße misst 100 nm x 100 nm auf. Stellen Sie die x-Achse des Schnittfensters, um mit dem großen (axial / Längsachse) der Myozyten entsprechen.
- Wählen Sie die endgültige Bildebene. Benutzen Bildrahmen, um den entsprechenden Bildebene in der z-Richtung manuell auswählen. Bestätigen, dass die TATS Membranen, darunter T-Tubuli und A-Tubulus Komponenten sind in der Brennebene visuell erkennbar. Beachten Sie, dass eine typische intrazelluläre Abbildungsebene kann einen Kern als intrazelluläre Referenzpunkt gehören. Beziehen sich auf die Beispiele in den 4A und 4B.
HINWEIS: In der Regel halten Zelle Bestrahlung mit Laserlicht so kurz wie möglich. Wenn möglich, verwenden Sie einzelne Bildrahmen, um die optimale Fokusebene in Herzmuskelzellen zu bestimmen. - Einstellen der Pixel Verweilzeit auf etwa 0,5 us. Wählen 16x Mittelung und speichert das Bild als Momentaufnahme. Wiederholen Sie den Snapshot-Image Schritt, um den entsprechenden Bildebene ein etablierens für TATS Membranstrukturen in 3,5 nach Bedarf beschrieben.
- Speichern Sie das fertige Bild, und bestätigen, dass die Datei in den Zielordner gespeichert. In der Regel speichern Sie alle Bilddateien im gleichen Format (zB LSM) für eine einheitliche Anwendung der Analyse-Software. Vor einer Bildanalyse, bestätigen einmal mehr ausreicht, Zellintegrität off-line (unter Schritt betrachten 1.12.1 aufgeführten Kriterien) und schließen alle geschädigten Zellen von der Analyse. Beziehen sich auf die Beispiele in den Figuren 4C und 4D.
4. Analyse der Membran TATS Netzwerk und seine Komponenten
Die folgenden Bildverarbeitungsschritte für die direkte Analyse von TATS Membrankomponenten werden als Top-down-Workflow-Diagramm in 5A und 5B zusammengefasst.
- Öffnen Sie die Image-Datei einer di-8-ANEPPS gefärbt Myozyten in Fidschi (http://fiji.sc/), eine frei verfügbare Variante des ImageJ, die für Analyse-Plugins enthältBildverarbeitung. Für weitere Informationen wenden Sie sich bitte an Schindelin et al 29 beziehen.
- Speichern Sie das Bild → Datei → Speichern unter → Tif.
- Um die TATS Membrankomponenten zu analysieren, wählen Sie die entsprechende ROI ohne die äußere Oberflächenmembran-Signal, dann benutzen Sie die "Polygonauswahl" Werkzeug, um den ROI Grenze ohne die äußere Oberflächenmembran (Sarkolemm) und einschließlich der intrazellulären Membranen Teile der TATS abzugrenzen, wie in 5A (ROI). Fügen Sie die ausgewählte ROI auf die "ROI-Manager", indem Analysieren → Extras → ROI-Manager.
- Um einen bestimmten ROI-Analyse für die Ausrichtung von TATS Komponenten auszuwählen, richten Sie den Hauptlängszellenachse und das Bild der x-Achse parallel. Wenn die Zelle leicht gebogen ist, wählen Sie mehrere ROIs und individuell auszurichten jede ROI. Schließen Sie alle Kerne aus der Analyse. Schließen Sie alle übermäßig gekrümmten Zellen von der Analyse, weil eine genaue Ausrichtung der ROI wird zunehmend schwieriger und während der Analyse zu erhöhen Orientierungsfehler.
- Löschen Sie unerwünschte Signalinformationen von der Außenfläche Membran: Bearbeiten → Löschen außen nach "gewählt ROI" (5A) zu generieren. Sicherzustellen, dass die ausgewählte ROI enthält nur die intrazelluläre Membranabschnitte, die mit dem Netzwerk TATS entsprechen.
- Nach der folgenden Kette von Bildverarbeitungsschritten (Befehle) vor der anschließenden quantitativen Analyse wie in 5A dokumentiert.
- Klicken Sie auf → Prozess → Subtrahieren Hintergrund. Stellen Sie die Rollkugelradius auf 5 Pixel.
HINWEIS: Stellen Sie die Rollkugelradius auf 5 Pixel, wenn das Bild analysiert hat eine Pixelgröße von 100 nm x 100 nm werden. Für andere Pixelgrößen setzen Sie die Rollkugelradius auf die Anzahl der Pixel, die in etwa einem physischen Umkreis von 500 nm entspricht. - Klicken Sie auf → Prozess → Verbessern Local Contrast (ClahE). Stellen Sie die Blockgröße auf 49, die Histogramm-Bins bis 256, die maximale Steigung bis 3 und die Maske auf "keine".
- Klicken Sie auf → Prozess → glatt.
- Klicken Sie auf → Plugins → Segmentierung → Statistische Region Merging. Stellen Sie die Parameter Q100 bis → klicken Sie zeigen Durchschnittswerte.
- Bestätigen Sie die Komplettbearbeitung des statistischen Zusammenführen Region durch automatische Präsentation eines neuen Bildrahmen angezeigt und bestätigen, dass das Etikett "SRM Q = 100" angezeigt wird. Weiterhin die folgenden Schritte mit dieser Image-Datei. Klicken Sie auf → Bild → Typ → 8-Bit.
- Klicken Sie auf → Bild → Stellen → Schwellwert. Wählen Sie die Schwelle niedrig genug, um die meisten erkennen, idealerweise alle TATS Strukturen, insbesondere vermeiden Ausschluss von TATS Komponenten mit geringer Signalintensität (mit einem Schwellenwert von 40 als Ausgangspunkt). Konsultieren Sie bitte den Abschnitt "Repräsentative Ergebnisse" für eine detaillierte Datenausgabe und die Beispiele in Abbildung 6(Oberer Schwellenwert von 255). Dokumentieren Sie die endgültige Wahl der Schwellenparameter. Beachten Sie, dass die richtige Wahl der Schwelle sollte nur bestimmte TATS Membranstrukturen falsch-positive Signale durch Hintergrundgeräusche erzeugen aber nicht.
- Bestätigen Sie die ordnungsgemäße Überlagerung von Bilddetails TATS gegen extrahiert Skelett Daten, insbesondere für mittlere und hohe Fluoreszenzsignalpegel, der die (Dis) Kontinuität der Skelettstruktur entsprechen sollte. Sobald ein geeigneter Schwellwert festgestellt wurde, gilt das gleiche Schwellenwert für alle Bilder bei der Analyse und wiederholen den Overlay Vergleich des Originalsignals im Verhältnis Gerippedaten konsistent dieses potenzielle Fehlerquelle zu minimieren.
- Klicken Sie auf Übernehmen →: die Bilddaten wird binär wie in Abbildung 5 unter "Threshold" gezeigt.
- Klicken Sie auf → Plugins → Skeleton → Skelettierung (2D / 3D). Speichern Sie das Bild als 2D skelettiert Tif-Datei (wie in Abbildung 5 dargestellt)Klicken Sie auf → Datei → Speichern unter → Tif. Analysieren Sie den skelettierten Bilddatei für die quantitative Datenausgang, indem Sie auf → Plugins → Analysieren Skeleton (2D / 3D). Wählen Sie Prune Zyklusmethode: keine. Bestätigen automatische Erzeugung des resultierenden Datentabelle.
- Die automatisch generierte Datentabelle als txt-Datei. Wählen Sie die entsprechende quantitative Parameter wie zB die Gesamtzahl der Verzweigungspunkte oder die durchschnittliche Zweiglänge, wie durch beispielhafte Daten in Abbildung 7 dargestellt. Betrachten weitere Datenanalyse mit komplementären / unterstützende Software-Tools wie Excel und als angemessen.
- Klicken Sie auf → Prozess → Subtrahieren Hintergrund. Stellen Sie die Rollkugelradius auf 5 Pixel.
- Betrachten Sie die automatisierte Bildverarbeitungsroutinen verwenden, wann immer möglich auch alle unter 4.5 beschrieben, um die Analyse zwischen den einzelnen Bildern und / oder Bildserien zu harmonisieren notwendigen Schritte. Verwenden Sie das Beispiel-Code Ergänzende Datei, die eine für die Bildverarbeitung programmiert Fidschi Makro (stellen Sie die Programmierung nach Bedarf).
- Analysieren Sie die inindividuelle Orientierungen aller oder wählen TATS Netzwerkkomponenten aus den skelettierten Bilddaten durch den Fidschi-Plugin "Direktionalität". Erzeugen eine Richtungs Histogramm, wo die jeweiligen axial ausgerichteten A-Tubuli oder quer ausgerichtet T-Tubuli Komponenten durch die 0 ° oder 90 ° Tonnen vertreten. Beachten Sie den richtigen Referenzbildorientierung und dass es wichtig ist, die x-Achse des Bildes, um eng mit den wichtigsten (längs) 0 °-Achse des VM Zelle entsprechen, wie in 8 und Repräsentative Ergebnisse gezeigt.
- Klicken Sie auf → Analysieren → Direktionalität → Definieren Methode: Fourier-Komponenten, Nbins 180, Histogramm beginnen -45 → klicken Sie auf Tabelle anzeigen.
- Speichern Sie die neu generiert und angezeigt Ergebnistabelle einschließlich der zugehörigen Histogramm-Daten als txt-Datei. Betrachten Sie die weitere Analyse der txt-Datei Daten durch kostenlose Software-Tools wie Excel und nach Bedarf.
- Unterklassen zu generierenDaten gruppiert Datensätze zB für alle unter den gleichen Bedingungen (und möglicherweise anderen Bedingungen) behandelten Zellen, wiederholen Sie die Schritte 4.1-4.7 die Analyse für alle relevanten Bilder als angemessen. Importieren Sie die skelettierten Datenparameter von allen Bildern kombiniert in einem Excel-Datei, um Durchschnittswerte abzuleiten. Darüber hinaus importieren alle Richtungs Histogramm Daten aus der gleichen Datenmenge zusammengefasst in eine Excel-Datei kombiniert zu berechnen und erzeugen die durchschnittliche Richt Histogramm. Betrachten Weiterverarbeitung von Datensätzen gruppiert, um zusätzliche TATS Netzwerkparameter von Interesse für einzelne oder zwischen verschiedenen Behandlungsgruppen zu analysieren, wie gebraucht.
Ergebnisse
Zusätzlich zu der TATS Membrannetzwerkanalyse eine Reihe von häufig verwendeten Zellbiologie Techniken wie die intrazelluläre Ca2 + Imaging, Patch-Clamp-Elektrophysiologie, oder pharmakologische Dosis-Wirkungs-Studien hängen entscheidend von hochwertigen Primärzellisolierung von den Vorhöfen oder Ventrikeln oder wählen Teile des Herzgewebes Charakterisierung der reifen differenzierten, strukturell und physiologisch intakten Herzmuskelzellen zu ermöglichen. Daher ist die Isolierung und für die Qualitätsbewertung im Kapitel 1. AM und VM-Zellen sind schließlich für viele andere Fragen, einschließlich der hier beschriebenen TATS Netzwerkanalyse, die sich kritisch auf intakter Membran und Zellintegrität abhängt.
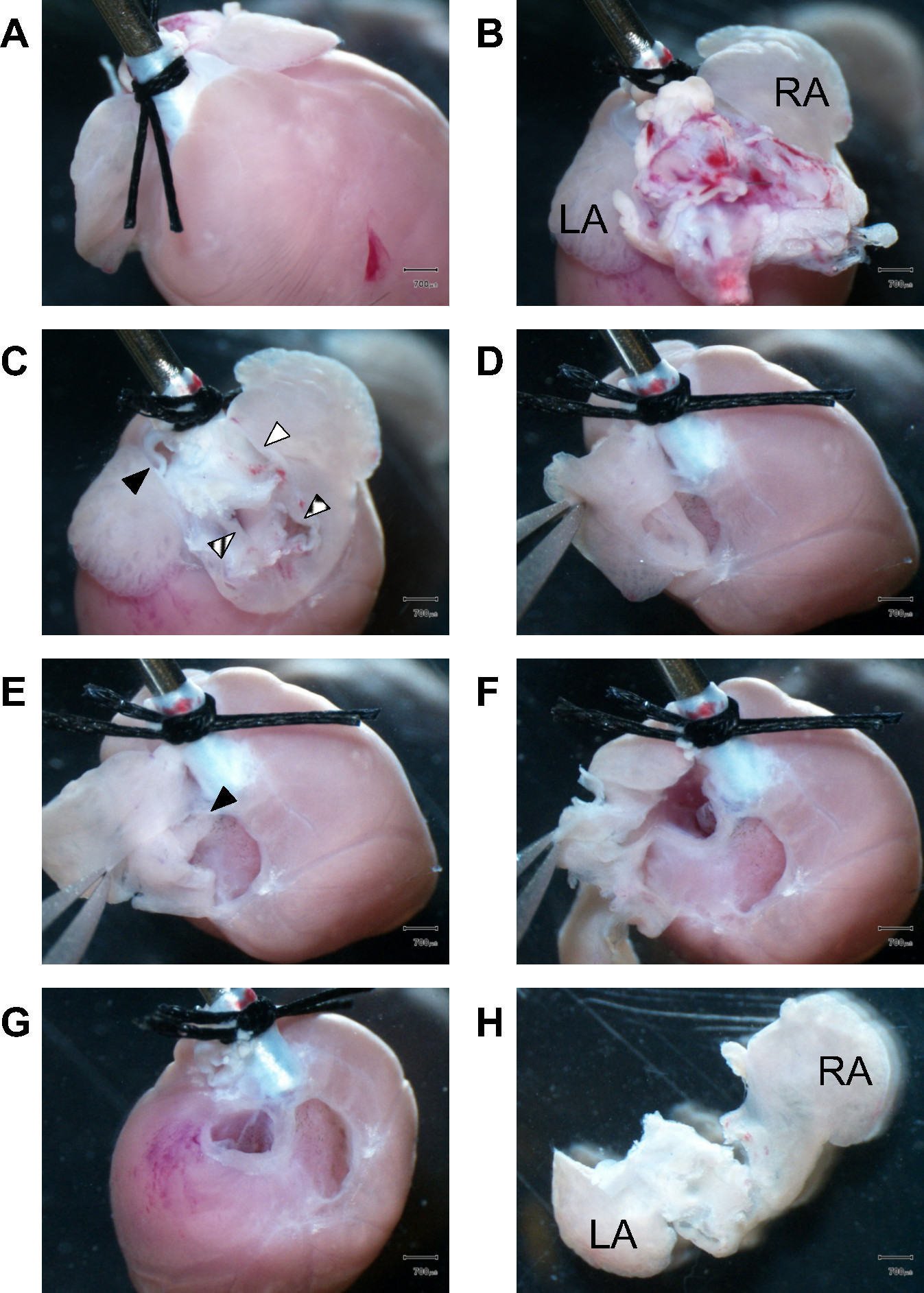
Abbildung 1 enthält eine schrittweise Anleitung, wie man Bilder mit dem Herzgewebe Dissektion beginnend mit den Vorhofkammern in Maus-Herzen kommen. Anschließend werden die ventrikulären Kammern und das Septum ein hergestelltd seziert nach Bedarf. Präzise Auswahl und Vorbereitung der richtigen Gewebeteile ist wichtig, AM und VM Isolierungen zuverlässig etablieren mit ausreichender Zell Reinheit. Folgende Collagenaseverdau es relativ schwierig sein, die korrekte Linie von Dissektion zwischen atrialen und ventrikulären Gewebe zu identifizieren, noch einmal morgens und VM Zellen unkontrolliert in Zellsuspension gemischt werden, ist es unmöglich, die gemischte Zellpopulation umkehren. Daher anatomischen Orientierung, 3D-Gewebe Visualisierung, ausreichend Erfahrungen mit verdauten Gewebebehandlung, korrekte Identifizierung von spezifischen Gewebeteile und deren Präparation Linien werden alle dazu beitragen, den Erfolg der Zellisolierung.

Abbildung 1.Dissection von Vorhofgewebe. (A) Mit Blick auf den vorderen Teil des Herzens, zwei OP-sutures fixieren den proximalen Aorta auf den Stumpf Ende einer 21 G Stahlkanüle. (B) Blick auf den Herz Basis zeigt restlichen Lungengewebe Behinderung der Vorhofkammerblick beim Präparieren. LA, linken Vorhof; RA, rechten Vorhof. (C) Rest lungtissue und der großen Gefäße wurden entfernt, um die Vorhofkammern zugreifen. Gefüllt schwarzes Dreieck, Lungenarterie; gestreiften Dreieck, Lungenvenen; weißes Dreieck, obere Hohlvene gefüllt; Box-Dreieck, untere Hohlvene. (D) Zuerst wird der rechte Vorhofwand seziert, während die Zange halten Sie die Herzohr. (E) Blick in den rechten Vorhof Kammer Höhle. Das schwarze Dreieck markiert den intakten Vorhofscheidewand. (F) Die Dissektion wird fortgesetzt, um die linke Herzkammer Hohlraum eintreten. (G) Nach vollständiger Dissektion des linken und des rechten Vorhofs, die Segelklappen sichtbar. Das faserige Ventilvorrichtung wird seziert und verworfen, um SUBSEQuently ernten nur ventrikuläre Muskelgewebe. (H) Hintere Ansicht der isolierten linken und rechten Vorhof. LA, linken Vorhof; RA, Recht atrium.Scale Bars: 700 um.
Um die Qualität der Zelle bestimmen, Isolierungen, Abbildung 2 zeigt typische Beispiele Zelle während Beurteilung der Ausbeute und Lebensfähigkeit der typischen Stab oder Ziegel geformt quergestreiften Muskelzellen intakt, sowohl für AMS und VMs. Wenig beschädigt, optisch unauffällig und schwer beschädigt Zellen mit abnormen mäßig gekrümmte Morphologien oder abnormale kugelförmige Zellen, können leicht identifiziert werden, einschließlich Trypanblauausschluß, wie in Abschnitt 1.11 beschrieben. Während intakte Muskelzellen bleiben hell und homogen, wenn quergestreiften auf extrazelluläre Trypanblau ausgesetzt, beschädigte Zellen zu zeigen, in der Regel mehrere Membran Bläschen und / oder schnell ansammeln Trypanblau intrazellulär anzeigt Membranschäden. Allerdings kann Trypanblau selbst Zellen durch unnecessari schadenly lange Inkubationszeit und sofortige Zellqualitätsbewertung ist daher zwingend. Beispiele für weitere offensichtlichen Formen von Zellschäden wie myofilament Kontraktur oder grober Oberflächenschäden beeinträchtigen die Integrität der Zelle werden in 4C und 4D gezeigt. Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
{kind=link}

Abbildung 2. Trypanblauausschluss Zellfärbung. Isolierte (A) AM und (B) VM-Zellen in Suspension mit Trypanblau gemischt und durch eine zumin 40Xmagnification gezeigt invertierte Lichtmikroskop sichtbar. Hinweis thatabnormal sphärischen Zellen in (A) und (B) die Aufnahme von Trypanblau, die leaka indicatesmembranege und Strukturschäden. Im Gegensatz dazu sind die centralAM und VM-Zellen mit intakten Membranen Trypanblau auszuschließen, wie gezeigt. Darüber hinaus ist zu beachten, dass intakte Uhr und VM-Zellen zeigen Sarkomer Streifen während ihrer Zellvolumen, nomembrane Bläschen, und scharfe Kanten an den beiden Querseiten und beide Glanzstreifen. Maßstabsbalken: 20 um.
Nach der erfolgreichen Trennung, Zellausbeuten von VMs von 5 x 10. Mai - 10. Juni können von einem einzigen Maus-Herz Verdauung zu erwarten. Die Ausbeute an AMS ist in der Größenordnung von 3 × 10 3 bis 3 × 10 4 stabförmigen, Trypanblau ohne Zellen deutlich geringer. Im Gegensatz zu VM, AM Isolierungen gelegentlich sogar in erfahrenen Händen scheitern. Schritt 1.11 fasst Verfahren, wie man die Ausbeute der isolierten gesunden Zellen in Suspension zu schätzen. Zusätzlich bestimmen die durchschnittliche Zellabmessungen bis Schritt 1.13, wie in Figur 3 für die einzelnen AM oder VM Zellisolaten gezeigten oder AM gegenüber VM Zellpopulationen zu vergleichen Side-by-Side (nach Bedarf). Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
{kind=link}

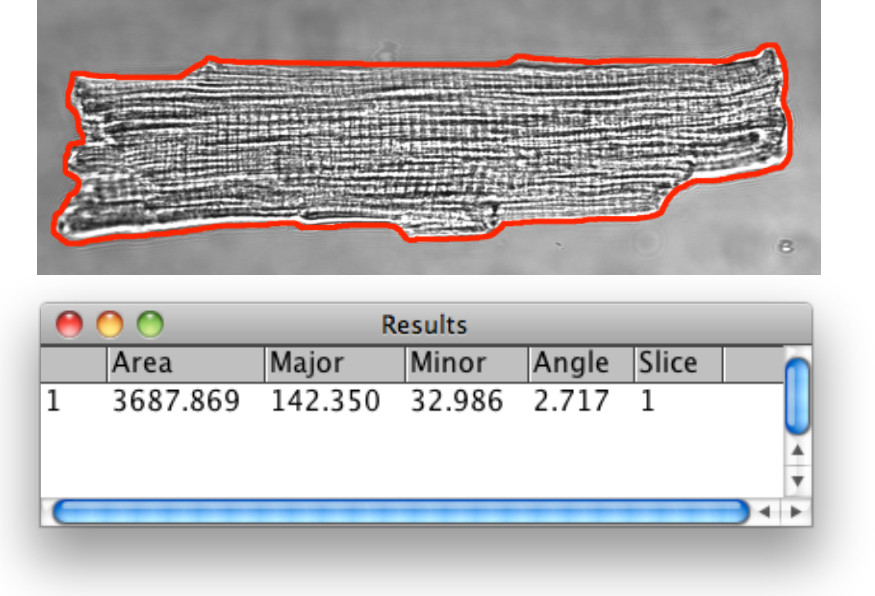
Abbildung 3.Bright Feld morphometrische Analyse der Herzmuskelzellen. Die VM Kontur wurde von den unter Schritt 1.13 beschriebenen Bildanalyse-Tools erkannt. Verwenden Sie das Polygon-Auswahl-Werkzeug, um Marke visuell und definieren die Zelle extrazellulären Grenze durch die Außenfläche Membran für die Analyse definiert. Anzeige der gewählten Region of Interest (ROI) auf die ROI-Manager, gefolgt von 1D-Abstandsmessungen. Für vergleichende Untersuchungen gegen AM VM Abmessungen ist es sinnvoll, die Zellenlänge, Breite und Fläche zu dokumentieren und die Berechnung der Länge: Breite-Verhältnis.
{kind=link}

Abbildung 4.live Membranfärbung von intakten atriale und ventrikuläre Myozyten. Durchlicht und konfokale Bilder der Lebens entsprechenden Di-8-ANEPPS gefärbten intakten (A) AM und (B) VM-Zellen. Im Gegensatz dazu ist eine teilweise zusammengezogen und möglicherweise beschädigt AM mit einem Sarkomerlänge von 1,2 um in (C) gezeigt. Vertraglich Muskelzellen zeigen typischerweise abnorm verkürzt und verzerrt TATS Strukturen, also von der weiteren Analyse ausgeschlossen. Ein wichtiger Indikator für Zellmembrandefekte Membran Bläschen (rote Dreiecke) in einer VM in (D) gezeigt. Membran Bläschen darstellen beschädigte Oberfläche Membranstrukturen und Zellen mit Bläschen von der weiteren Analyse ausgeschlossen TATS werden. Darüber hinaus zeigt die VM Brutto Schäden offenbar fehlt ein ganzes Teil seiner linken unteren Teil (von Sternchen markiert). Zusammenfassend durch den Vergleich Durchlicht und konfokale Bilder, Zellen Morphologie und Oberflächen Unversehrtheit wird dokumentiert und in Kombination mit Fluoreszenzsignal Informationen. 'N' Marken nuCLEI weggelassen aus der Analyse des TATS Membranfleck. Gelbe Balken deuten vergrößerte ROIs aus der gleichen konfokalen Bild oben dargestellt. Maßstabsbalken:. 10 um Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
{kind=link}
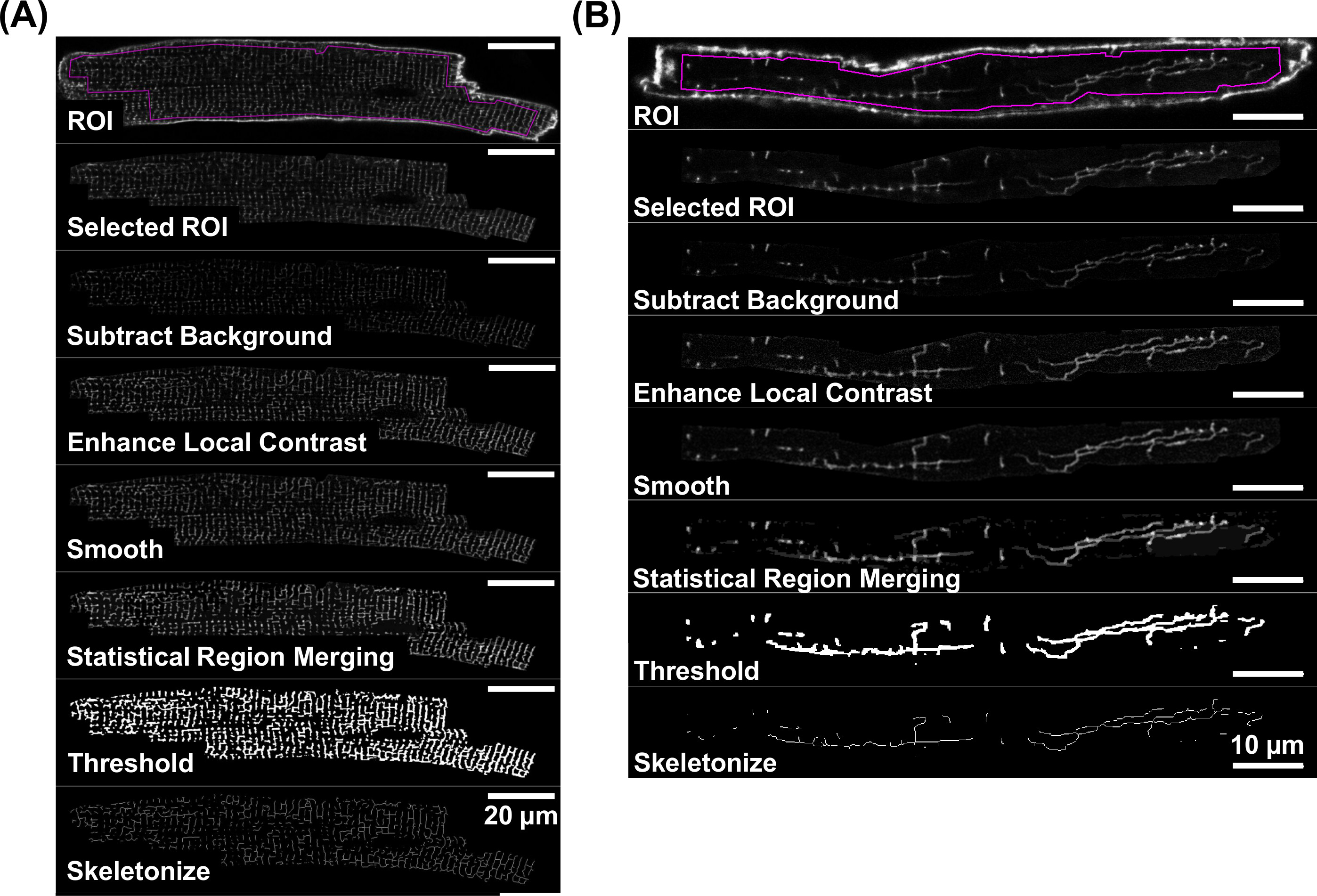
Konfokale Bilder von TATS Membranen mit einem ausreichenden Signal-Rausch-Verhältnis wie in den 4A und 4B gezeigt sind, für eine weitere quantitative Analyse erlaubt. Die Membran TATS Analyse basiert auf Daten von den skelettierten Fluoreszenz geradlinigen Signalkomponenten abgeleitet. 5 zeigt das Ablaufdiagramm der einzelnen Bildverarbeitungsschritte, die im Detail in den Schritten 4.3 bis 4.5 beschrieben. Diese Schritte erzeugen skelettiert Bilder, die geradlinigen TATS Membran Netzwerke als für jede VM isoliert (5A) gezeigt und AM-Zellen (Figurieren 5B).

Figur 5.Workflow zur Skelettierung des Fluoreszenz TATS Bilder. Bildverarbeitungsschritte, die zu einer skelettierten Bild des TATS Netzwerk führen, werden von den einzelnen Schritt-für-Schritt-Bild Beispiele sowohl für einen di-8-ANEPPS dargestellt gefärbten VM (A) und AM (B). Für einzelne Bildverarbeitungsschritt, finden Sie in Abschnitt 4 verwiesen. Hinweis Unterschiede in Umfang Bars. 20 um (A) und 10 um (B) Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
{kind=link}
Ein entscheidender Schritt bei der Bildverarbeitung, die geeignete Schwelle für Binarisierung Daten, wie in Abschnitt 4.5 beschrieben.6. Das resultierende Binärbild sollte nur wahre Membran Signale von der TATS Netzwerk, aber nicht durch fehlerhafte Schwellen aus dem Hintergrund Signalrauschen abgeleitet falsche Strukturen umfassen. Dennoch ist es wichtig, dass die Schwelle niedrig genug ist, um alle wahren TATS Strukturen wie wahr TATS Komponenten nicht irrtümlich während der Bildanalyse verloren erfassen. 6 zeigt den Vorgang, wie die Schwelle bei der Daten Digitalisierung auszuwählen. Während ein Schwellenwert von 40, wie in den 6B und 6C gezeigt ist angebracht alle wahren TATS Strukturen einzeln zu erfassen, die Wahl eines höheren Schwellen Beispiel von 60, wie in 6A gezeigt, nicht schwächer axialen Membranstrukturen (ATS) zu erfassen, wie durch gelbe Dreiecke . Im Gegensatz, die Wahl eines niedrigeren Schwellen zB von 20, wie in 6D gezeigt führt zu fehlerhaften Erfassung von Hintergrundgeräuschen, wie falsch-positive TATS Strukturen, wie durchgelbe Dreiecke.

Abbildung 6.How, um die Signalschwelle während Skelettierung von TATS Bilddaten zu bestimmen. Die Beispiele zeigen, TATS Skelette jeweils für unterschiedliche Schwellenwerte während der Daten Binarisierung (in Schritt 4.5 beschrieben) erzeugt. Ober Bilder: zeigen die Schwelle Einstellungen mit Fidschi. Nieder Bilder: Überlagerung von skelettierten Bilder mit der entsprechenden Bildeingabe Fluoreszenz und vergrößert Regionen wie angegeben. Eine hohe Schwelle zB 60 in der angewandten (A) ist offenbar nicht geeignet, um alle wahren TATS Strukturen zu erkennen, wie durch gelbe Dreiecke in der vergrößerten Abschnitt angegeben. Ein Schwellenwert von 40 in (B) und (C) angewendet erkennt alle TATS Strukturen und richtig nicht Hintergrundgeräusche zu erkennen, während eine niedrige Schwelle zB., 20 in (D) fälschlicherweise identifiziert Hintergrundgeräusche wie TATS Strukturen und produziert falsch-positiven Signale der nicht vorhandene Membranstrukturen dabei. Falsch-positive Signale werden durch gelbe Dreiecke in der vergrößerten Einsatz angezeigt. Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
{kind=link}
Die "Analyse Skeleton (2D / 3D)" Plugin unterstützt die detaillierte Analyse der skelettierten TATS Strukturen. Einmal ausgeführt, das Plugin erzeugt eine Datentabelle mit den folgenden Parametern Skelett: #branches, #junctions, # Endpunkt Voxel, die durchschnittliche Zweiglänge, #triple Punkte, #quadruple Punkte, und die maximale Länge Zweig. Für eine detaillierte Beschreibung aller möglichen Ausgabeparameter finden Sie unterhttp: //fiji.sc/wiki/index.php/AnalyzeSkeleton und verwandten Artikeln 29-31. Ein typisches Datentabelle ausgegeben wird, in 7A gezeigt.
Die Parameter können ferner verwendet werden, um die Gesamtskelettlänge pro Fläche oder der Anzahl der Übergänge pro Fläche abzuleiten. Beispielhafte Berechnung der Anzahl der Filialen multipliziert mit der durchschnittlichen Zweig Länge gibt die Gesamtlänge des kontinuierlichen Skelett in 2D:Gesamtlänge pro Skelett ROI:
Σ (x #branches durchschnittliche Zweiglänge) = 5155 px = 515,5 um
Die Gesamtlänge der Skelettfaktoren auf die Bildfläche. Für das in Figur 7 der normierte Skelettlänge von 0.64μm gezeigten Beispiel / um 2 und die Summe aller Knotenpunkte berechnet werden, wie unten gezeigt:
Normalisierten Skelettlänge:
515,5 um / 803,6 um 2 = 0,64 um / um 2
Normalisierte Anzahl von Knotenpunkten:
155 Kreuzungen / 803,6 um 2 = 0,19 Gänge / um 2

Abbildung 7.Automated Datenausgabe von skelettierten Bilder. (A) Ein von der "Analysieren Skeleton (2D / 3D)" Plugin von der in (B) gezeigt skelettiert Bild erzeugt typische Daten Tabelle. Für eine detaillierte Beschreibung der möglichen Ausgangsparameter finden Sie unter http://fiji.sc/wiki/index.php/AnalyzeSkeleton . Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
{kind=link}
Die unter 4.5 beschrieben und in 5 dargestellt Bildverarbeitungsschritte können unter Verwendung eines Makro Fiji als Ergänzungscodedatei vorgesehen automatisieren . Die Befehle definieren Wiederholungen der Bildverarbeitung durch reiterative Schritten. Das Makro kann angewendet werden, um Stapel von durch die Schritte 4.3 und 4.4 produziert Eingangsbilder zu vervollständigen. Das Makro kann für die Analyse der kompletten Datensatz Gruppen vorteilhaft sein, beispielsweise durch Herstellung von einzelnen Eingangsbildstapel für jede unabhängige Behandlungsgruppe für die automatisierte Analyse unter Verwendung der Makrobefehle.
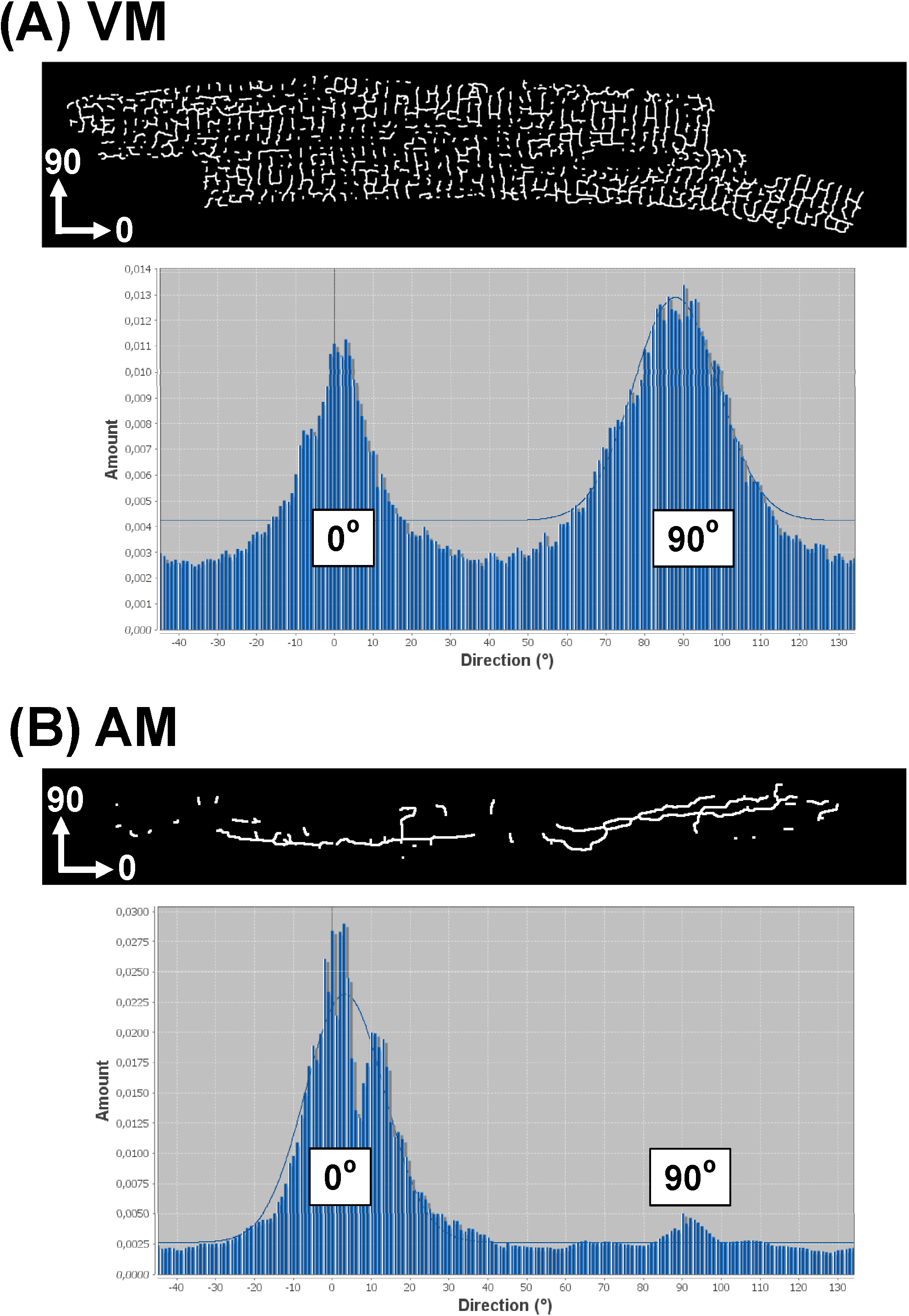
Das skizzierte Software-Strategie ermöglicht ferner die TATS Netzwerk Orientierungsanalyse für alle Komponenten. Dazu benutzen Sie die "Direktionalität"-Plugin (http://fiji.sc/Directionality) 29,31, die Histogramm-Daten, die die Orientierungsverteilung aller TATS Komponente Orientierungen produziert. Wenn der X-Achse des Eingangsbildes entspricht, um die Hauptachse einer gegebenen AM oder VM Zelle, die axial (longitudinal) TATS Komponenten werden von der 0 ° Bin repräsentiert werden, wobei die Querkomponenten durch die 90-dargestellt werden# 176; bin. Abbildung 8 zeigt beispielhafte Richt Histogramme von skelettierten TATS Bilder für eine VM (8A) gegenüber einem AM (8B) Zelle. Während die Direktionalität Histogramm eines typischen VM zeigt eine Doppelspitzenverteilung bei 0 ° und 90 °, das AM-Histogramm zeigt eine einzelne dominante Peak bei 0 °. Diese Beispiele sind in Übereinstimmung mit früheren Beobachtungen, dass die einzelnen Komponenten der TATS VMs fast gleichmäßig zwischen T-Tubuli und A-Tubuli verteilt, während die TATS Komponenten AMS kann überwiegend aus A-Tubuli zusammengesetzt sein.

Abbildung 8.Representative Richt Histogramm von TATS Netzwerke von einzelnen Zellen. Direktionalität Histogramme wurden aus skelettierten Bilder der einzelnen VM (A) gegenüber der Uhr erzeugt (B ) TATS Netzwerken. Da die x-Achse des analysierten Bildes entspricht Haupt (Längsachse) der Myozyten wie dargestellt, sind A-Tubulus-Komponenten von der 0 ° Behälter dargestellt, während T-Tubuli Komponenten durch die 90 ° Behälter dargestellt. Gauß passt nur zu den großen Histogramm peakasshown als Hintergrund Graphen. Bitte klicken Sie hier, um eine größere Version dieser Figur zu sehen.
{kind=link}
| Perfusionspuffer | mM |
| NaCl | 120,4 |
| KCl | 14,7 |
| KH 2 PO 4 | 0,6 |
| Na 2 HPO 4 | 0,6 |
| MgSO 4 | 1.2 |
| HEPES | 10 |
| NaHCO 3 | 4.6 |
| Taurin | 30 |
| 2,3-Butandion-Monoxim | 10 |
| Glucose | 5.5 |
| pH-Wert 7,4 | |
| Verdauungspuffer | mm (wenn nicht angegeben) |
| NaCl | 120,4 |
| KCl | 14,7 |
| KH 2 PO 4 | 0,6 |
| Na 2 HPO 4 | 0,6 |
| MgSO 4 | 1.2 |
| HEPES | 10 |
| NaHCO 3 | 4.6 |
| Taurin | 30 |
| 2,3-Butandion-Monoxim | 10 |
| Glucose | 5.5 |
| Collagenase Typ II | 600 U / ml |
| pH-Wert 7,4 | |
| Anschlagpuffer | mm (wenn nicht angegeben) |
| NaCl | 120,4 |
| KCl | 14,7 |
| KH 2 PO 4 | 0,6 |
| Na 2 HPO 4 | 0,6 |
| MgSO 4 | 1.2 |
| HEPES | 10 |
| NaHCO 3 | 4.6 |
| Taurin | 30 |
| 2,3-Butandion-Monoxim | 10 |
| Glucose | 5.5 |
| CaCl2 | 0,0125 |
| Kälberserum | 10% |
| pH-Wert 7,4 | |
Tabelle 1.Buffer solutIonen. Der Inhalt von drei unterschiedlichen physiologischen Pufferlösungen zur Zellisolierung und Abbildungs zusammengefasst.
Diskussion
Obwohl Herzmuskelzellen wurden isoliert und untersucht seit Jahrzehnten 32, eine aktuelle Überprüfung ergab, dass eine gleichbleibend hohe Qualität myocyte Zellisolierungen bleiben 27-. Dies spiegelt relativ komplexe Protokolle für die Isolierung von primären Herzmuskelzellen gegenüber einem Mangel an gemeinsamen Standards Ansätze, gemeinsame Metadaten und transparente Zelle Qualitätsdokumentation. Zellisolierung Protokolle werden in der Regel von einzelnen Gruppen angepasst, produzieren unterschiedliche Ergebnisse von Zell isoliert, hängen von individuellen Modelleinstellungen (zB, Art, Alter, Herzerkrankungen nebeneinander), und sind in der Regel für bestimmte Versuchsbedingungen angepasst. Im Rahmen der quantitativen TATS Membran Studien und Protokolle hier vorgestellt, ein wesentlicher Niveau der Qualitätsbewertung und Dokumentation betrifft die konfokale Mikroskopie oder Superauflösung der einzelnen Zelle Membranstrukturen anfällig für metabolische und Isolation Protokoll abhängige Veränderungen, sowohlin AM oder VM. Wichtig ist, auch wenn hohe Ausbeuten an Zell Isolate vorschlagen gesunde, intakte Muskelzellen, müssen die Ermittler zu dokumentieren und kritisch jede einzelne Zelle sorgfältig gegen morphologischen Kriterien der Oberfläche und TATS Membranintegrität gegenüber nicht-spezifische Schäden durch Isolierungsverfahren gegen bestimmte Veränderungen durch verschiedene Arten zu beurteilen von Interventionen im Vergleich zu Bedingungen zu kontrollieren. Eine wichtige Größe bei Herzzellisolierung ist die spezifische Aktivität einer gegebenen Menge Kollagenase. Um eine neue Menge von Kollagenase zu wählen, sollten die enzymatische Aktivität der Kollagenase mehrere Proben gegeneinander durch die Auswertung Kardiomyozyten-Ausbeute und Qualität, und nach den Anweisungen des Herstellers getestet werden. Idealerweise wird eine neue Charge von Collagenase mit Kollagenase-Aktivität gegenüber den Vorjahren erfolgreich viele identifiziert (für erweiterte Evaluierung möglicher Enzymaktivitäten finden Sie in der "Collagenase viel Auswahl-Werkzeug" in der Materialien und Methoden Tabelle). Taken zusammen, quantitative Ansätze der Visualisierung TATS Membran hängen entscheidend von Zellisolierung Qualität und, umgekehrt, Herzzellisolierungen, die zu unspezifischen Membranschädigung durch TATS Mikroskopie dokumentiert sollten kritische Überprüfung und Korrektur der Isolationsverfahren auslösen. Da Zellisolierung Qualität und TATS Membran Visualisierung und Quantifizierung sind untrennbar miteinander verbunden, die in diesem Artikel beschriebenen Protokolle decken alle wichtigen Aspekte einer kontinuierlichen Strategie.
Eine zusätzliche Herausforderung und gemeinsame Ausgabe von Herzuntersuchungen, Zellschäden und / oder Zellverlust entstehen durch metabolisch Kompromisse Interventionen zB nach Herzinfarkt 9, noch brauchen, um gegen mögliche unbeabsichtigte Beschädigung zB beurteilt werden, unbemerkt folgenden Luftembolie während der Zellisolierung. Isolierung von Herzmuskelzellen von erkrankten Herzen zu zusätzlichen, erheblichen Zellverlust führen und verminderte Zellausbeuten. Daher ComparIson der Gesamtzahl der isolierten intakten Zellen zwischen Kontrolle und kranken Herzen können sinnvoll sein, wenn der Zellisolierung und Zählung ist ständig durch standardisierte Protokolle eingesetzt. Folglich ist es wichtig, die Integrität der Zelle durch eine entsprechende Kontrollgruppe, die die bestmögliche myocyte Zellisolierung Qualität spiegelt beurteilen. Wichtig ist, dass einzelne Zellqualität und Lebendzellmikroskopie von gesunden gegenüber kranken gegenüber Muskelzellen versehentlich durch die Isolierung beschädigt Verfahren kann die Analyse von Membran TATS Netzwerke wesentlich beeinflussen. Die hier vorgestellten Protokolle betonen daher die Integrität und Stabilität des physiologischen Membrankomponenten während der Zellisolierung und Lebendzellmikroskopie von intakten Membranen. Der gesamte Workflow wird als eine kontinuierliche Strategie zu erreichen und zu bewahren intakt TATS Membrankomponenten unter Ausschluss beschädigte Zellen, da diese zeigen Isolierung abhängige Membran Artefakte wie gestört Membran Tubuli, membrane Bläschen, und veränderte TATS Netzwerke irrtümlich unter Kontrollbedingungen und Kompromisse weitere quantitative Analyse. Umgekehrt, sind die gleichen Strategien entscheidend für Interventionsstudien mit dem Potenzial, TATS Membranen, die sich kritisch auf aussagekräftigen Vergleich zwischen Kontrollen wahr gesunde gegenüber wahr erkrankten Zelle mit TATS Membran Veränderungen hängen zu stören.
Darüber hinaus beschäftigen wir uns mit Verfahren, um die technisch viel anspruchsvoller Isolierung von AM Zellen zu erreichen. Trotz der Fortschritte und verbesserte Protokolle, ist es wichtig zu betonen, dass es nicht trivial ist, um qualitativ hochwertige Zell Isolierungen von VMs reproduzieren und noch weniger zuverlässig für AMS. Dies ist aufgrund der insgesamt geringeren Ertrag von AM Zellen, in denen auch kleine Fehler oder Abweichungen in der Zellisolierung kann zum Ausfall der Zellisolierung Uhr abzuschließen, während ein milder Grad der VM Zellschäden können in Zellsuspension weniger offensichtlich sein, aufgrund der relativ hohen Zell Zahlen im Vergleich zu Uhr. Seit AM Zellen könnten c werdennach der Isolierung urved, kann die Analyse durch mehrere ROIs von Vorteil sein, wie unter Schritt 4.3 beschrieben. Nach einer detaillierten Verfahren der Zellisolierung Schritte, bieten wir ein Protokoll für den direkten integralen Membranfärbung und konfokaler Bildgebung oder STED-Superauflösung von TATS Netzwerke sowohl für VMs und AMS. Diese Protokolle ermöglichen sowohl quantitative Analyse und Differenzierung von ausgewählten Komponenten der TATS Membranen durch vorher festgelegten Parametern. Im Vergleich zu VMs, sind die 3D-Organisation und funktionale Verhaltensweisen des Vorhof TATS Netzwerk AMS derzeit weniger verstanden.
Die Verfahren, die Bild TATS Membranen in lebenden Zellen (Schritte 3.1 bis 3.7) wurden mit kommerziellen konfokalen (Tabelle der Materialien / Ausrüstung) und maßgeschneiderte STED Fluoreszenzmikroskope 9 entwickelt. Um die Einstellungen für Fluoreszenzmikroskop Bilderzeugung und quantitative Analyse TATS zu optimieren, von allgemeiner Bedeutung, die die folgenden Punkte:
- Ziel
Um kleine Details TATS Membranstrukturen lösen, empirisch testen, welche Zielsetzung die höchste Bildqualität bietet bei gleichzeitiger Konzentration mehrere Mikrometer tief in der Zelle. Bestimmte konfokale Mikroskope können besser mit Wasser oder Glycerol Ziele durchzuführen, im Gegensatz zu dem 63X 1,4 NA Öl-Objektiv verwendet. Ziele mit 100-facher Vergrößerung für Superresolution-STED-Mikroskopie verwendet wird, den Handel aus einer kleineren Sichtfeld für Nanometerauflösung 34. - Anregung und Gewinn
Die optimalen Einstellungen der Erregerleistung und Detektorverstärkung hängen von Lichtwegs im Mikroskop, Laser-Leistung und Probeneigenschaften. Im Idealfall, Laserleistung und Verstärkung eingestellt werden, um die gesamte Bandbreite des Detektors zu nutzen, noch zu vermeiden Bildsättigung. Handels Mikroskopie-Software-Pakete bieten in der Regel Lookup-Tabellen, die die untere und obere Grenze des Dynamikbereichs zu visualisieren. Darüber hinaus minimiert Farbbleichung beschäftigen die bestmöglichen lAser Macht, die immer noch eine ausreichende Struktur TATS Membran Details. Auch sollte Erregerleistung niedrig genug sein, um zu vermeiden, kumulative Foto Schäden, die zu myocyte Kontrakturen und Tod. - Pixelgröße
Verwenden Sie einen Pixelgröße mit Nyquist-kompatibel, etwa die Hälfte der mit den gegebenen Einstellungen erreichte Auflösung. Für die konfokale Abbildung eine Pixelgröße von 100 nm x 100 nm kompatibel ist, was auch begrenzen Bleichen. Superresolution-Mikroskopie für deutlich kleinere Pixelgrößen sind zB 20 nm x 20 nm für die STED-Mikroskopie 9 verwendet. - Verweilzeit
Konfokale Mikroskope bieten eine Mittelungsfunktion. In der Regel verwenden die kürzest mögliche Pixelverweilzeit Bleich in Kombination mit Signalmittelung zB vermeiden, Line-Mittelung ≥ 8, um das Signal-zu-Rausch-Verhältnis zu verbessern. - Dokumentieren Sie die Angewandte Mikroskopie Einstellungen Durch Meta-Daten
Sobald die Einstellungen wieBilddetails TATS Membranstrukturen wurden auf einem bestimmten konfokalen Mikroskop optimiert, sicher und / oder dokumentieren Sie die Einstellungen (Protokoll Meta-Daten). Erwerben Sie alle Bilder in einem (oder zwischen) Gruppe (n) von Zellen mit dem gleichen Ziel, Erregerleistung, Verstärkung, Pixelgröße, Pixel Verweilzeit und Mittelungsfunktion. Gleich Abbildungsbedingungen ermöglichen den direkten Vergleich und die Quantifizierung in einem (oder zwischen) Gruppe (n) der Zellen. - Zur allgemeinen Orientierung und weitere Details zu Grundlagen und Anwendungen der konfokalen Mikroskopie beziehen sich auf das Handbook of Biological konfokale Mikroskopie (Pawley JB, 3. Auflage, 2006, Springer Science + Business Media, LLC).
Im Gegensatz zu den direkten Analyse Strategien hier vorgestellten bisherigen Publikationen beschreiben TATS Membranen und krankheitsbedingte Veränderungen wurden regionale Gesamtanzeigen von T-Tubuli Dichte wie quantitative Strategie 16,17 oder indirekte regionale Strategien basierend auf Fourier transf verwendetormationen Analyse der quergestreiften Membran Signale, um T-Tubuli Komponente Regelmäßigkeit 7 zu bewerten. Im Gegensatz dazu sind die hier beschriebenen quantitativen Ansätzen direkt an einzelne TATS Komponenten bezogen und bieten eine Reihe von zusätzlichen Parametern, einschließlich Membranwerkeigenschaften und spezifische Komponenten wie der Anteil der A-Tubuli. Weiterhin kann die Netzwerkdichte TATS als normierte Länge des gesamten Skeletts extrahiert pro ROI-Bereich quantifiziert werden. Die Anzahl der Dreifachübergänge der drei einzelnen, kontinuierlich verbunden Tubulus Komponenten als Maß für die Komplexität der Verzweigungs TATS Membran Netzwerk verwendet werden. Wir stellen fest, dass eine Analyse der kleinsten TATS Komponenten hängt von den Färbeverfahren. Nach unserer Erfahrung sind 800 ul einer 50 uM di-8-ANEPPS Lösung ausreicht, um komplette TATS Netzwerke in einer Zelle Pellet mit 50.000 VM 9 Zellen färben. Wenn jedoch das Zellpellet enthält, eine geringere Anzahl von HerzmuskelzellenWenn leistungsstarke Fluoreszenzdetektoren vorhanden sind, und wenn die konfokale Abbildung der gesamten TATS Netzverteilung statt kleinsten Membrandaten und quantitative Veränderungen von Interesse sind, niedriger Farbstoffkonzentrationen können anhand von empirischen Tests verwendet werden. Schließlich kann ein Software-Makros für die beschriebene Analyse geschrieben werden die Bildverarbeitungsschritte zur Analyse großer Datenmengen, die für den Vergleich zwischen den verschiedenen Behandlungsgruppen (zum Beispiel Arzneimittel), Zelltypen, besonders nützlich ist, zu erleichtern automatisieren (zB AM gegen VM ) und pathophysiologischen Interventionen (zB Schein gegen Myokardinfarkt).
Für die Bildanalyse von TATS Netzwerken wird die folgende Sequenz von Schritten aufgetragen Prinzip: 1) rollenden Ball Hintergrund-Subtraktion (4.5.1), um räumliche Schwankungen in der Hintergrundintensität zu entfernen; 2) lokale Kontrastverstärkung (4.5.2). 3) Bildglättung (4.5.3); 4) statistische Region Verschmelzung (4.5.4); 5) definiert, dieSchwelle von Bild Binarisierung (4.5.6); und 6) Berechnung der Skelettdaten (4.5.8). Ein entscheidender Schritt bei der Skelettierung von Fluoreszenzbildern TATS ist die in 6 gezeigte Bild Binarisierung. Die zugehörigen Schwellen Schritten letztlich bestimmen, welche wahre Membranstrukturen erkannt werden, um die darunterliegende Komponenten gegen TATS durch Fehler vom Hintergrundrauschen erkannt potentiell falsche Strukturen darstellen. Die Ermittlung der richtigen Schwelle für binäre Bildanalyse sollte mit den wahren TATS Membranstrukturen, die auf einem ausreichend hohen Signal-zu-Rausch-(SNR) jedes der konfokalen Mikroskopie und Hochauflösungs-Ansätze hängt entsprechen. Daher sollte eine ausreichende Bildqualität und anschließend mit ersten kritischen Urteil von Einzelzellqualität einschließlich Dokumentation von Hellfeld-Bildern kombiniert, wie beschrieben eingerichtet werden. Alternativen, um die Bildsegmentierung Protokoll für einen gegebenen Objektdatenausgang und / oder anzupassen physiologische Fragen sind Bilddekonvolution und andere Schwellenwertverfahren wie "Otsu" oder "Iso-Daten", wie ImageJ Plugins zur Verfügung. Unabhängig von der letzten Segmentierungsverfahren halten wir den Vergleich zwischen extrahiert und Rohdaten von Bild-Overlay die obligatorische Qualitätskontrolle Schritt. Zusammenfassend wird morphologischen und Membranintegrität der einzelnen isolierten Myocyten ausreichende Färbung von intrazellulären Membranen TATS, Parameteroptimierung für die Fluoreszenzbildgebung und Lagerungssteuer extrahierter Gerippedaten tragen zur Qualität von Fluoreszenz TATS Bildern und quantitative Ergebnisse.
Wenn größere Arten als Maus zur Zellisolierung verwendet wird, kann die Protokolle ohne weiteres entsprechend angepasst werden. Für die nächsten größeren Arten kann Rattenherzen mit einer stumpfen Kanüle 14 G (Außendurchmesser 2,1 mm) kanüliert und 8 ml / min durchströmt werden. Deutlich älter oder krank Herzen können auch größere Kanüle Größen erfordern. In der Gen-RAL kann Herzperfusion entweder mit konstantem Druck durchgeführt werden, zB unter Verwendung einer 1 m hohen Wassersäule zwischen dem Behälter und der Aorta oder durch konstante Strömung unter Verwendung einer peristaltischen Pumpe. Zur Zellisolierung von kleinen Nagetieren Herzen wie Mäusen und Ratten kann konstant Fluss vorteilhaft sein, da Kollagenaseverdau schließlich stören koronaren Widerstandsgefäße, die zu übermäßiger Perfusionsraten aus undichten Gefäßbetten, die zu einem gewissen Grad durch konstante Durchflussprotokolle gesteuert wird. Im Gegensatz dazu ist konstanter Druck Perfusion vorteilhaft, wenn die Überwachung der Durchflussrate und korrekte Kanülierung eine Priorität, die zur Intervention Modelle mit veränderten Blutgefäßwiderstandsverhalten als auch für die Ausbildung der Zellisolierungsverfahren vorteilhaft ist.
Wie oben dargelegt, ist ausreichend Zellqualität für quantitative Studien der endogenen Membransystemen sehr wichtig. Doch während der Herzdurchblutung und Kollagenaseverdau zahlreichen Faktoren CRitically Einfluss auf die Qualität der Zellisolation, die nie während der Protokolloptimierung oder Fehlersuche, 27 zu unterschätzen ist. Insbesondere sollte die Aktivität einer gegebenen Menge Collagenase für den spezifischen Gewebe von Interesse zB Atrien oder Ventrikel vor der Ausführung der Versuchs bona fide Studien zur Isolierung Bedingungen fest bestimmt den Rest der Studie aufrechterhalten wird. Außerdem wird die Wasserqualität, pH, Temperatur, Optimierung und Reinigung der Perfusion Einrichtung die Gefahr einer unbeabsichtigten Beschädigung durch Verunreinigungen und Embolien zu minimieren, und möglicherweise weiteren Faktoren überwacht werden müssen, um eine optimale homöostatischen Bedingungen während der Zellisolierung zu schaffen. BDM (2,3-Butandion-monoxim) ein reversibler Inhibitor der Myosin-ATPase Querbrücken wird üblicherweise während Gewebedissektion und Verdauung verwendet, um die Herzmuskelentspannung, die die Ausbeute an Zellisolierungen erhöht aufrechtzuerhalten. Dennoch müssen die Ermittler to beachten Sie, dass BDM kann unspezifische Phosphatase-Aktivitäten, die zu Off-Target-Effekte zB Hemmung der Na + / Ca 2 + Wechselströme unter bestimmten Bedingungen 33 auszuüben. Bei einigen Experimenten es vorteilhaft, BDM durch blebbistatin als Kardioplegie-Lösung, einem Inhibitor mit einer hohen Affinität für Myosin in mikromolaren Konzentrationen, ist jedoch toxisch und relativ teuer zu ersetzen sein könnte und kann andere Nebeneffekte haben. Ruhen gesunden Herzmuskelzellen sollten keine Kontraktionen in Abwesenheit von elektrischer Stimulation und solche Zellen von der weiteren Analyse ausgeschlossen werden, zeigen. Auf der anderen Seite, Kardiomyozyten-Kontraktion und Relaxation als Reaktion auf elektrische Stimulation bei physiologischen extrazellulären Ca 2 +-Konzentrationen können verwendet werden, um normale kontraktile Verhalten als zusätzliche Maßnahme zu schaffen, um funktionelle Zellqualität und / oder Verhaltensstörungen in der Herzkrankheit und gesunden Kontroll beurteilen Zellen.
Zusammenfassend sind die Protokolle für die Einzelzellisolierung und quantitative hier beschriebenen Bildanalyse wurden erfolgreich für die konfokale Mikroskopie und Superresolution des TATS Membran-Netzwerk in VM 9 Zellen 21 sowie für die quantitative Analyse der Mikrotubuli-Netzwerke in der angewandten und AM feste Herzmuskelzellen (Daten nicht gezeigt). Diese und zukünftige Anwendungen der Protokolle kann Wege für eine Vielzahl von experimentellen Fragen wie die Charakterisierung von Membranen TATS in verschiedenen Entwicklungsstadien oder die Analyse von membranassoziierten Protein oder Organellen Strukturen, die die TATS Netzwerk Kontakt offen auszuüben stark lokalisierten, Domain-spezifische Signal Funktionen in AM und VM-Zellen.
Offenlegungen
The authors declare that no conflict of interest exists.
Danksagungen
This work received support through Deutsche Forschungsgemeinschaft SFB 1002 (subprojects A05 and B05 to S.E.L.) and KFO 155 (subproject 4 to S.E.L.), a Halbach Foundation award to S.E.L. supporting E.W.; a grant from the German Cardiac Society to S.B.; and a DAAD exchange program supporting T.K. as visitor at the University of Maryland. The research leading to these results has received funding from the European Community’s Seventh Framework Program FP7/2007-2013 under grant agreement No. HEALTH-F2-2009-241526, EUTrigTreat (to S.E.L.). S.E.L. is a principal investigator of the German Center of Cardiovascular Research (DZHK).
Materialien
| Name | Company | Catalog Number | Comments |
| Chemicals and Enzymes | |||
| 2,3-Butanedione monoxime | Sigma-Aldrich, Munich, Germany | B0753 | |
| Bovine calf serum | Thermo Scientific, Schwerte, Germany | SH30073 | Triple 0.1 µm sterile filtered. |
| CaCl2 | Sigma-Aldrich, Munich, Germany | 21115 | Diluted 1:10 in MQ water to obtain 100 mM CaCl2 stock concentration. |
| Collagenase type II | Worthington via Cell Systems, Troisdorf, Germany | on request | Enzymatic activity depends on individual collagenase batches. Collagenase II and other enzyme activities (Caseinase, Clostripain, Tryptic) can be assessed in the "collagenase lot selection tool". Determine cell yield and quality individually for each new lot of collagenase. |
| Glucose | Carl Roth, Karlsruhe, Germany | HN06.1 | |
| Heparin | Rotexmedica, Trittau, Germany | PZN-03862340 | Diluted in 0.9% NaCl and injected subcutaneuosly in abdominal skin. |
| HEPES | Carl Roth, Karlsruhe, Germany | 9105.4 | |
| Forene 100% (V/V) | Abbott, Libertyville, IL, USA | B506 | Active agent: isoflurane, 250 ml. Use approximately 2 Vol% in air/oxygen dispenser instrument. |
| KCl | Carl Roth, Karlsruhe, Germany | 6781.3 | |
| KH2PO4 | Carl Roth, Karlsruhe, Germany | 3904.2 | |
| Laminin (2 mg/ml) | BD Biosciences, Heidelberg, Germany | 354232 | Lamination is described under step 2.1. |
| MgCl2·6H2O | Carl Roth, Karlsruhe, Germany | 2189.2 | |
| MgSO4·7H2O | Carl Roth, Karlsruhe, Germany | 8283.2 | |
| Na2HPO4·2H2O | Carl Roth, Karlsruhe, Germany | 4984.2 | |
| NaHCO3 | Carl Roth, Karlsruhe, Germany | HN01.1 | |
| Taurin | Carl Roth, Karlsruhe, Germany | 4721.2 | |
| Dyes | |||
| Di-8-ANEPPS | Molecular Probes, Life Technologies, Darmstadt, Germany | D-3167 | Stock solution 2 mM in DMSO |
| Trypan blue | Sigma-Aldrich, Munich, Germany | T8154 | Trypan blue is gently mixed 1:1 via tip-cut 1 ml plastic pipette with cell suspension prior to cell counting in Neubauer cytometer. |
| Langendorff Perfusion Setup | |||
| Circulation thermostat | Lauda, Lauda-Königshofen, Germany | Please refer to Louch et al. (JMCC 2011). Heat up thermostat und buffers in perfusion tubing to 37 °C 15 min prior to use. | |
| Flexible silicone tubing Tygon for peristaltic pump | VWR, Darmstadt, Germany | 224-2252 | Tubing needs to be changed regularly. |
| Flexible silicone tubing Tygon for thermostat | VWR, Darmstadt, Germany | 228-4340 | |
| Heating coil surroundung perfusion tubing | Rettberg, Göttingen, Germany | custom-made | Heating coil and tubing needs to be cleaned thoroughly via MQ water after using. Do not use detergents. Glass components should be bathed regularly in 10 mM NaOH overnight. |
| Peristaltic pump | Ismatec, Wertheim, Germany | ISM830 | |
| Three way stop cock Discofix C Luer Lock 10 cm | Braun, Melsungen, Switzerland | 16500C | |
| Three way stop cock Discofix 3SC | Braun, Melsungen, Switzerland | 4095146 | |
| Instruments | |||
| 42 mm glass coverslips | Menzel Gläser via Thermo Scientific, Schwerte, Germany | on request | 0.13-0.16 mm thickness |
| Cannula 21 G | Becton, Dickinson and Company, Franklin Lake, NY, USA | 304432 | Cut to a length of ~5 mm, roughened with sandpaper. |
| Coverslips for Neubauer cytometer 24 x 24 mm | Menzel Gläser via Thermo Scientific, Schwerte, Germany | on request | 0.38-0.42 mm thickness |
| Graefe forceps, 0.5 mm tips, slight curve | Fine Science Tools, Heidelberg, Germany | 11151-10 | |
| LSM 710 NLO | Carl Zeiss, Jena, Germany | 63X 1.4 NA oil objective | |
| Neubauer improved cytometer | Labor Optik, Friedrichsdorf, Germany | 1100000 | Counting procedure: Wipe cytometer and coverslip provided with the counting chamber with 70 % ethanol. Press coverslip gently on the counting chamber so that the two glass surfaces are in contact and Newton's rings can be observed. Subsequently, 10-20 µl cell suspension can be applied to the edge of the coverslip to be sucked into the void by capillary action. Count the intact vs. defect myocytes using the squares of the cytometer grid which reflects 100 nl. Repeat counting procedure on the second grid provided on the cytometer. Calculate the density of cells in your original cell suspension by taking account of any dilutions and counting shortcuts. |
| POC-R2 Imaging Chamber | Pecon, Erbach, Germany | Cell suspension volume: 800 µl; desired plating density: ~1,000 AM and ~10,000 VM | |
| Spring scissors, 8 mm blades straight, blunt | Fine Science Tools, Heidelberg, Germany | 15025-10 | |
| Student dumont #7 forceps, inox | Fine Science Tools, Heidelberg, Germany | 91197-00 | |
| Student iris scissors, curved, 11.5 cm | Fine Science Tools, Heidelberg, Germany | 91461-11 | |
| Student iris scissors, straight, 11.5 cm | Fine Science Tools, Heidelberg, Germany | 91460-11 | |
| Student surcigal scissors, straight, sharp, 12 cm | Fine Science Tools, Heidelberg, Germany | 91402-12 | |
| Tissue forceps, 1 x 2 teeth, slim, 10 cm | Fine Science Tools, Heidelberg, Germany | 11023-10 | |
Referenzen
- Prosser, B. L., Ward, C. W., Lederer, W. J. Subcellular Ca2+ signaling in the heart the role of ryanodine receptor sensitivity. J Gen Physiol. 136 (2), 135-142 (2010).
- Wehrens, X. H., Lehnart, S. E., Marks, A. R. Intracellular calcium release and cardiac disease. Annu Rev Physiol. 67, 69-98 (2005).
- Cheng, H., Cannell, M. B., Lederer, W. J. Propagation of excitation-contraction coupling into ventricular myocytes. Pflugers Arch. 428 (3-4), 415-417 (1994).
- Williams, G. S., Chikando, A. C., Tuan, H. T., Sobie, E. A., Lederer, W. J., Jafri, M. S. Dynamics of calcium sparks and calcium leak in the heart. Biophys J. 101 (6), 1287-1296 (2011).
- Sperelakis, N., Rubio, R. orderly lattice of axial tubules which interconnect adjacent transverse tubules in guinea-pig ventricular myocardium. J Mol Cell Cardiol. 2 (3), 211-220 (1971).
- Soeller, C., Cannell, M. B. of the transverse tubular system in living cardiac rat myocytes by 2-photon microscopy and digital image-processing techniques. Circ Res. 84 (3), 266-275 (1999).
- Song, L. S., et al. et al. ryanodine receptors in the failing heart. Proc Natl Acad Sci USA. 103 (11), 4305-4310 (2006).
- Oort, R. J., et al. Disrupted junctional membrane complexes and hyperactive ryanodine receptors after acute junctophilin knockdown in mice. Circulation. 123 (9), 979-988 (2011).
- Wagner, E., et al. Stimulated emission depletion live-cell super-resolution imaging shows proliferative remodeling of T-tubule membrane structures after myocardial infarction. Circ Res. 111 (4), 402-414 (2012).
- Asghari, P., Schulson, M., Scriven, D. R., Martens, G., Moore, E. D. Axial tubules of rat ventricular myocytes form multiple junctions with the sarcoplasmic reticulum. Biophys J. 96 (11), 4651-4660 (2009).
- Lukyanenko, V., Ziman, A., Lukyanenko, A., Salnikov, V., Lederer, W. J. Functional groups of ryanodine receptors in rat ventricular cells. J Physiol. 583 (Pt 1), 251-269 (2007).
- Shacklock, P. S., Wier, W. G., Balke, C. W. Local Ca2+ transients (Ca2+ sparks) originate at transverse tubules in rat heart cells. J Physiol. 487 (Pt 3), 601-608 (1995).
- Reynolds, J. O., et al. Junctophilin-2 is necessary for T-tubule maturation during mouse heart development. Cardiovasc Res. 100 (1), 44-53 (2013).
- Di Maio, A., Karko, K., Snopko, R. M., Mejia-Alvarez, R., Franzini-Armstrong, C. T-tubule formation in cardiac myocytes two possible mechanisms. J Muscle Res Cell Motil. 28 (4-5), 231-241 (2007).
- He, J., et al. Reduction in density of transverse tubules and L-type Ca(2+) channels in canine tachycardia-induced heart failure. Cardiovasc Res. 49 (2), 298-307 (2001).
- Heinzel, F. R., et al. Remodeling of T-tubules and reduced synchrony of Ca2+ release in myocytes from chronically ischemic myocardium. Circ Res. 102 (3), 338-346 (2008).
- Lyon, A. R., et al. Loss of T-tubules and other changes to surface topography in ventricular myocytes from failing human and rat heart. Proc Natl Acad Sci USA. 106 (16), 6854-6859 (2009).
- Crossman, D. J., Ruygrok, P. N., Soeller, C., Cannell, M. B. Changes in the organization of excitation-contraction coupling structures in failing human heart. PLoS One. 6 (3), e17901(2011).
- Kemi, O. J., et al. The effect of exercise training on transverse tubules in normal, remodeled, and reverse remodeled hearts. J Cell Physiol. 226 (9), 2235-2243 (2011).
- Sachse, F. B., et al. Subcellular structures and function of myocytes impaired during heart failure are restored by cardiac resynchronization therapy. Circ Res. 110 (4), 588-597 (2012).
- Arakel, E. C., et al. Tuning the electrical properties of the heart by differential trafficking of KATP ion channel complexes. J Cell Sciences. 127 (Pt 9), 2106-2119 (2014).
- Richards, M. A., et al. Transverse tubules are a common feature in large mammalian atrial myocytes including human. Am J Physiol Heart Circ Physiol. 301 (5), H1996-H2005 (2011).
- Trafford, A. W., Clarke, J. D., Richards, M. A., Eisner, D. A., Dibb, K. M. Calcium signalling microdomains and the t-tubular system in atrial mycoytes potential roles in cardiac disease and arrhythmias. Cardiovasc Res. 98 (2), 192-203 (2013).
- Greiser, M., Schotten, U. Dynamic remodeling of intracellular Ca2+ signaling during atrial fibrillation. J Mol Cell Cardiol. 58, 134-142 (2013).
- Voigt, N., Zhou, X. B., Dobrev, D. Isolation of human atrial myocytes for simultaneous measurements of Ca2+ transients and membrane currents. J Vis Exp. (77), 10-3791 (2013).
- Kaestner, L., et al. Isolation and genetic manipulation of adult cardiac myocytes for confocal imaging. J Vis Exp. (31), (2009).
- Louch, W. E., Sheehan, K. A., Wolska, B. M. Methods in cardiomyocyte isolation, culture, and gene transfer. J Mol Cell Cardiol. 51 (3), 288-298 (2011).
- King, N. M., et al. Mouse intact cardiac myocyte mechanics cross-bridge and titin-based stress in unactivated cells. J Gen Physiol. 137 (1), 81-91 (2011).
- Schindelin, J., et al. Fiji, an open-source platform for biological-image analysis. Nat Methods. 9 (7), 676-682 (2012).
- Arganda-Carreras, I., Fernandez-Gonzalez, R., Munoz-Barrutia, A., Ortiz-De-Solorzano, C. 3D reconstruction of histological sections Application to mammary gland tissue. Microsc Res Tech. 73 (11), 1019-1029 (2010).
- Liu, Z. Q. Scale space approach to directional analysis of images. Appl Opt. 30 (11), 1369-1373 (1991).
- Powell, T., Twist, V. W. A rapid technique for the isolation and purification of adult cardiac muscle cells having respiratory control and a tolerance to calcium. Biochem Biophys Res Commun. 72 (1), 327-333 (1976).
- Watanabe, Y., et al. Inhibitory effect of 2,3-butanedione monoxime (BDM) on Na(+)/Ca(2+) exchange current in guinea-pig cardiac ventricular myocytes. Br J Pharmacol. 132 (6), 1317-1325 (2001).
- Kohl, T., Westphal, V., Hell, S. W., Lehnart, S. E. Superresolution microscopy in heart - cardiac nanoscopy. J Mol Cell Cardiol. 58, 13-21 (2013).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten